Антитела: функция, виды, формы взаимодействия с антигенами
Федеральное государственное бюджетное
образовательное учреждение
Высшего профессионального образования
РЕФЕРАТ
на тему:
"Антитела: функция, виды, формы взаимодействия с
антигенами"
г.
Содержание
Введение
1. Природа антител
2. Молекулярное строение антител
3. Структурно-функциональные особенности иммуноглобулинов различных
классов
4. Антигенность антител
5. Механизм взаимодействия антитела с антигеном
6. Свойства антител
7. Генетика иммуноглобулинов
8. Динамика антителопродукции
9. Теории разнообразия антител
Заключение
Библиографический список
Введение
Термин "антитело" был введён в
употребление в конце XIX века. В 1890 году Беринг (Behring) и Китасато
(Kitasato) провели эксперименты, в которых они изучали на морских свинках действие
дифтерийного и столбнячного токсинов. Они вводили животным сублетальную дозу
токсина, через некоторое время брали у них сыворотку и вводили её вместе с
летальной дозой токсина другим животным, в результате чего животные не
погибали. Был сделан вывод, что после иммунизации токсином в крови животных
появляется вещество, способное нейтрализовать его и тем самым предотвратить
заболевание. Данное вещество получило название антитоксина, а затем был введён
более общий термин - антитело; вещества вызывающие образование антител стали
называть антигенами.
Только в 1939 году Тизелиус (Tiselius) и
Кэбет (Kabat) показали, что антитела содержатся в определённой фракции белков
сыворотки. Они иммунизировали животное овальбумином и из полученной сыворотки
взяли две пробы, в одну из них был добавлен овальбумин и образовавшийся осадок
(комплекс антитело-овальбумин) удалили. Электрофорез выявил, что в пробе куда
добавлялся овальбумин, содержание г-глобулинов значительно ниже чем в другой
пробе. Это указывало на то, что антитела являются г-глобулинами. Чтобы отличить
их от других белков, содержащихся в этой фракции глобулинов, антитела были
названы иммуноглобулинами. Сейчас известно, что антитела обнаруживаются в
значительных количествах также во фракциях б - и в-глобулинов.
Структура антител была установлена в ходе
разнообразных экспериментов. В основном они заключались в том, что антитела
обрабатывались протеолитическими ферментами (папаин, пепсин), и подвергались
алкилированию и восстановлению меркаптоэтанолом. Затем исследовались свойства
полученных фрагментов: определялась их молекулярная масса (хроматографией),
четвертичная структура (рентгеноструктурным анализом), способность связываться
с антигеном и т.п. Также использовались антитела к данным фрагментам:
выяснялось, могут ли антитела к одному типу фрагментов связываться с
фрагментами другого типа. На основе полученных данных была построена
описываемая ниже модель молекулы антител.
антитело антиген иммуноглобулин молекулярный
1.
Природа антител
Одной из филогенетически наиболее древних форм иммунной
защиты является биосинтез антител - белков, специфически реагирующих с
антигенами. Антитела относятся преимущественно к γ-глобулиновой фракции
белков плазмы крови, на долю которых приходится 15-25% ее белкового содержания,
что составляет примерно 10-20 г/л. Поэтому антитела получили название иммуноглобулинов,
и их обозначают символом Ig. Следовательно антитела - это γ-глобулины плазмы крови, способные специфически связываться с
антигеном и участвовать во многих иммунных реакциях.
Антитела синтезируются В-лимфоцитами и их потомками -
плазматическими клетками и в циркулирующей форме, и в виде рецепторных молекул
на иммунокомпетентных клетках. Циркулирующие антитела подразделяются на
сывороточные и секреторные. К антителам могут быть также отнесены белки
Бенс-Джонса, которые являются фрагментами молекулы Ig (его легкая цепь) и
синтезируются в избытке при миеломной болезни.
Строение и функцию антител изучали многие видные ученые: П.
Эрлих (1885) предложил первую теорию гуморального иммунитета, Э. Беринг и С.
Китазато (1887) получили первые антитоксические сыворотки к дифтерийному и
столбнячному токсинам, А. Безредка (1923) разработал метод безопасного введения
пациентам лечебных иммунных сывороток. Большая заслуга в расшифровке молекулярного
строения Ig принадлежит Д. Эдельману и Р. Портеру (1959), а разгадка
многообразия антител - Ф. Бернету (1953) и С. Тонегаве (1983).
2.
Молекулярное строение антител
Иммуноглобулины - это белки сыворотки крови. Они
секретируются плазматическими клетками в ответ на антиген. Молекулы Ig имеют
универсальное строение (рис.1). Они состоят из 2 пар полипептидных цепей: двух
тяжелых (550-660 аминокислотных остатков, молекулярная масса - 50 кД) и двух
легких (220 аминокислотных остатков, молекулярная масса - 20-25 кД). Обозначают
их как H - (от англ. heavy - тяжелый) и L - (от англ. light - легкий)
цепи. Тяжелые и легкие цепи связаны между собой попарно дисульфидными связями
(-S-S-). Между тяжелыми цепями также есть дисульфидная связь - это так
называемый шарнирный участок. Такой тип межпептидного соединения позволяет
молекуле Ig легко менять свою конформацию в зависимости от окружающих условий и
состояния. Область шарнирного участка ответственна за взаимодействие с первым
компонентом комплемента (С1) и его активацию по классическому пути.
Различают структурные варианты легких и тяжелых полипептидных
цепей молекулы Ig. Легкие цепи бывают 2 типов: κ и λ (каппа и лямбда). Тяжелых цепей известно 5 типов: α, γ, μ, ε и δ (альфа, гамма, мю,
эпсилон и дельта). Среди многообразия цепей α-типа выделяют α1 - и а2-подтипы, μ-цепей - μ1 и μ2, γ-цепей - γ1-, γ2-, γ3 - и γ4-подтипы.
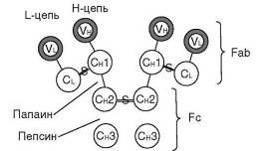
Рис. 1. Схема строения молекулы иммуноглобулина класса G: V -
вариабельный домен; C - константный домен; S - дисульфидная связь шарнирного
участка
Вторичная структура полипептидных цепей молекулы Ig имеет
доменное строение - ее отдельные участки свернуты в глобулы (домены),
стабилизированные внутренней дисульфидной связью. Таких доменов в составе
тяжелой цепи Ig бывает 4-5, в легкой - 2. Каждый домен состоит примерно из 110
аминокислотных остатков.
Домены различаются по постоянству аминокислотного состава.
Выделяют С-домены (от англ. constant - постоянный) с относительно
постоянной структурой и V-домены (от англ. variable - изменчивый)
с переменной структурой. В составе легкой цепи есть по одному V - и С-домену, а
в тяжелой - один V - и 3-4 С-домена. Примечательно, что не весь вариабельный
домен изменчив по своему аминокислотному составу, а лишь его незначительная
часть - гипервариабельная область, на долю которой приходится около 25%.
Вариабельные домены легкой и тяжелой цепей совместно образуют
участок, который специфически связывается с антигеном, - антигенсвязывающий
центр, или паратоп. Гипервариабельные области тяжелой и легкой цепей
определяют индивидуальные особенности строения антигенсвязывающего центра для
каждого клона Ig и многообразие их специфичностей.
Обработка ферментами молекулы Ig приводит к ее гидролизу на
определенные фрагменты. Так, папаин разрывает молекулу выше шарнирного участка
и ведет к образованию трех фрагментов (см. рис.1). Два из них способны
специфически связываться с антигеном. Они состоят из цельной легкой цепи и
участка тяжелой (V - и C-домен), и в их структуру входят антигенсвязывающие
участки. Эти фрагменты получили название Fab (от англ. - фрагмент,
связывающийся с антигеном). Третий фрагмент, способный образовывать кристаллы,
получил название Fc (от англ. - фрагмент кристаллизующийся). Он
ответствен за связывание с рецепторами на мембране клеток макроорганизма
(Fc-рецепторы) и некоторыми микробными суперантигенами (например, белком А
стафилококка). Пепсин расщепляет молекулу Ig ниже шарнирного участка и ведет к
образованию 2 фрагментов: Fc и двух сочлененных Fab, или F (ab) 2.
В структуре молекул Ig обнаруживают дополнительные полипептидные цепи. Так,
полимерные молекулы IgM, IgA содержат J-пептид (от англ. join - соединяю),
который объединяет отдельные мономеры в единое макромолекулярное образование
(см. раздел 11.1.3). Молекулы секреторных Ig обладают S-пептидом (от
англ. secret - секрет). Это так называемый секреторный компонент. Его
молекулярная масса составляет 71 кД, он является β-глобулином и предохраняет
молекулу Ig в секрете слизистых оболочек от ферментативного расщепления.
Рецепторный Ig, локализующийся на цитоплазматической мембране
антителопродуцирующих клеток, имеет дополнительный гидрофобный трансмембранный М-пептид
(от англ. membrane - мембрана). Он прочно удерживает молекулу Ig в
липидном бислое цитоплазматической мембраны и проводит рецепторный сигнал через
цитоплазматическую мембрану внутрь клетки. J - и M-пептиды присоединяются к
молекуле Ig в процессе ее биосинтеза. S-пептид является продуктом эпителиальной
клетки - он присоединяется к J-пептиду полимерной молекулы Ig при ее
транслокации через эпителиальную клетку.
3.
Структурно-функциональные особенности иммуноглобулинов различных
классов
В зависимости от особенностей молекулярного строения тяжелой
цепи, а следовательно, наличия изотипических, или групповых, антигенных
детерминант различают 5 классов или изотипов Ig (рис.2). Молекулы, содержащие
тяжелую цепь α-типа, относят к изотипу, или классу A
(сокращенно IgA), δ-типа - IgD, ε-типа - IgE, γ-типа - IgG и μ-типа - IgM. Различают также подклассы Ig.
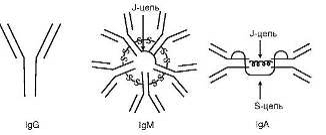
Рис. 2. Схема строения иммуноглобулинов различных
классов (пояснение в тексте)
Для каждого изотипа Ig характерны свои особенности. В
частности, Ig D, E и G имеют мономерное строение, IgM практически всегда
является пентамером, а молекула IgA может быть моно-, ди - и тримером. Наиболее
характерные черты различных изотипов Ig приведены в табл.1.
Таблица 1. Основные характеристики
иммуноглобулинов человека
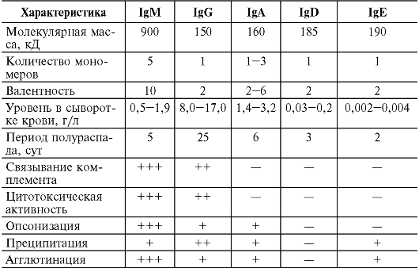

Иммуноглобулин класса G составляет основную массу
Ig сыворотки крови, на его долю приходится 70-80% всех циркулирующих Ig, при
этом 50% содержится в тканевой жидкости. Среднее содержание IgG в сыворотке
крови здорового взрослого человека - 12 г/л, что достигается к 7-10-летнему
возрасту. Период полураспада IgG 21 день.- мономер, имеет 2 антигенсвязывающих
центра, может связать 2 молекулы антигена подряд. Молекулярная масса около 160 кД,
константа седиментации 7S. Синтезируется зрелыми В-лимфоцитами (Βγ) и плазматическими клетками. Хорошо определяется
в сыворотке крови на пике первичного и при вторичном иммунном ответе. Обладает
высокой аффинностью.
Различают подтипы G1-G4. IgG1 и G3 связывают комплемент,
причем G3 активнее. IgG4 подобно IgE обладает цитофильностью (тропностью, или
сродством, к тучным клеткам и базофилам) и участвует в развитии аллергической
реакции I типа.
Легко проходит через плацентарный барьер и обеспечивает
гуморальный иммунитет новорожденного в первые 3-4 мес после рождения, в том
числе обнаруживается в молоке. IgG обеспечивает нейтрализацию и маркирование
антигена, осуществляет запуск комплементопосредованного цитолиза и АЗКЦТ.
Иммуноглобулин класса M - наиболее крупная молекула
из всех Ig. Это пентамер, который имеет 10 антигенсвязывающих центров. Его
молекулярная масса около 900 кД, константа седиментации 19S. Различают подтипы
М1 и М2. Тяжелые цепи молекулы IgM, в отличие от других изотипов, построены из
5 доменов. Являясь полимерной молекулой, содержит J-цепь. Период полураспада 5
дней.
На его долю приходится 5-10% всех циркулирующих Ig. Среднее
содержание IgM в сыворотке крови здорового взрослого человека около 1 г/л.
Этого уровня человек достигает уже к 2-4-летнему возрасту. IgM филогенетически
наиболее древний иммуноглобулин. Образуется в начале первичного иммунного
ответа.
Обладает высокой авидностью, наиболее эффективный активатор
комплемента по классическому пути. Большая часть нормальных антител и изоагглютининов
относится к IgM. Не проходит через плаценту. Обнаружение высоких титров
специфических антител изотипа M в сыворотке крови новорожденного указывает на
бывшую внутриутробную инфекцию или дефект плаценты. IgM обеспечивает
нейтрализацию и маркирование антигена, осуществляет запуск
комплементопосредованного цитолиза и АЗКЦТ. Является маркером острого
инфекционного процесса.
Иммуноглобулин класса A существует в сывороточной
и секреторной формах. Около 60% всех IgA содержится в секретах слизистых
оболочек.
Сывороточный IgA. На его долю приходится около 10-15%
всех циркулирующих Ig. В сыворотке крови здорового взрослого человека
содержится около 2,5 г/л IgA, максимум достигается к 10 - летнему возрасту.
Период полураспада 6 дней.- мономер, имеет 2 антигенсвязывающих центра,
молекулярную массу около 170 кД и константу седиментации 7S. Различают подтипы
A1 и A2. Синтезируется зрелыми иммунными В-лимфоцитами (Βα) и плазматическими клетками. Хорошо определяется
в сыворотке крови на пике первичного и при вторичном иммунном ответе. Обладает
высокой аффинностью. Не связывает комплемент. Не проходит через плацентарный
барьер. IgA обеспечивает нейтрализацию и маркирование антигена, осуществляет
запуск АЗКЦТ.
Секреторный IgA (sIgA) существует в полимерной
форме в виде диили тримера (4 - или 6-валентный), несет 4 или 6 паратопов и
содержит J - и S-пептиды. Молекулярная масса 350 кД и выше, константа
седиментации 13S и выше.
Синтезируется В1-лимфоцитами, плазматическими клетками и,
возможно, В1-лимфоцитами в пределах слизистых оболочек и выделяется в их
секреты. Объем продукции может достигать 5 г в сутки. Пул sIgA считается самым
многочисленным в организме - его количество превышает суммарное содержание IgM
и IgG. В сыворотке крови sIgA не обнаруживается.
Формирование четвертичной структуры молекулы sIgA происходит
при ее транслокации через эпителиальную клетку. На базальной и латеральной
поверхности эпителиальная клетка несет рецептор к J-цепи полимерной молекулы Ig
(JR). Присоединяясь к рецептору, IgA эндоцитируется клеткой в виде везикулы и
переносится к апикальной поверхности эпителиоцита, где JR подвергается
ферментативному расщеплению. В результате IgA высвобождается в слизистый секрет
просвета органа уже в секреторной форме, так как оставшийся прикрепленным к
молекуле Ig фрагмент JR становится S-цепью.
Секреторная форма IgA - основной фактор специфического
гуморального местного иммунитета слизистых оболочек желудочнокишечного и
респираторного тракта, мочеполовой системы. Благодаря S-цепи он устойчив к
действию протеаз. sIgA не активирует комплемент, но эффективно связывается с
антигенами, нейтрализует их и препятствует адгезии микробов на эпителиальных
клетках.
Иммуноглобулин класса E называют также реагином.
Содержание в сыворотке крови крайне невысоко - примерно 0,00025 г/л.
Молекулярная масса около 190 кД, константа седиментации примерно 8S, мономер.
На его долю приходится около 0,002% всех циркулирующих Ig. Этот уровень
достигается к 10-15 годам жизни.
Синтезируется зрелыми В-лимфоцитами (Βε) и плазматическими клетками преимущественно в
лимфоидной ткани бронхолегочного дерева и желудочно-кишечного тракта. Не
связывает комплемент. Не проходит через плацентарный барьер. Обладает
выраженной цитофильностью - тропностью к тучным клеткам и базофилам. Участвует
в развитии гиперчувствительности немедленного типа - реакция I типа.
Иммуноглобулин класса D практически полностью
содержится в сыворотке крови в концентрации около 0,03 г/л (около 0,2% общего
количества циркулирующих Ig). IgD имеет молекулярную массу 160 кД и константу
седиментации 7S, мономер. Не связывает комплемент. Не проходит через
плацентарный барьер. Экспрессируется на предшественниках В-лимфоцитов.
Рецепторные иммуноглобулины, или мембранные,
локализуются на цитоплазматической мембране В-лимфоцитов и выполняют
функции их антигенспецифических рецепторов. Имеют те же
изотип и специфичность, что и синтезируемые в межклеточную среду антитела.
Содержат особый дополнительный M-пептид, благодаря которому молекула
рецепторного Ig фиксируется в цитоплазматической мембране иммунокомпетентной
клетки.
Нормальные антитела, или естественные, -
совокупность Ig сыворотки крови человека различной специфичности, формирующих
их базальный уровень. К ним относят изогемагглютинины - антитела к
эритроцитарным антигенам групп крови (например, система АВ0), антигенам
бактерий кишечной группы, кокков и некоторых вирусов. Эти антитела постоянно
образуются в организме без видимой антигенной стимуляции. Отражают готовность
макроорганизма к иммунному реагированию, а также свидетельствуют об отдаленном
контакте с антигеном.
Моноклональные антитела. Каждый В-лимфоцит и его
потомки, образовавшиеся в результате клеточного деления (т.е. клон), способны
синтезировать антитела с паратопом строго определенной специфичности. Такие
антитела получили название моноклональных. В естественных условиях
макроорганизма получить моноклональные антитела практически невозможно, так как
на одну и ту же антигенную детерминанту одновременно реагируют до 100 различных
клонов В-лимфоцитов, незначительно различающихся антигенной специфичностью.
Поэтому в результате иммунизации даже монодетерминантным антигеном мы всегда
получаем поликлональныеантитела.
Принципиально получение моноклональных антител выполнимо,
если провести предварительную селекцию антителопродуцирующих клеток и их
клонирование, т.е. получение необходимых клонов. Однако задача осложняется тем,
что число генераций В-лимфоцитов, как и других эукариотических клеток,
ограничено. Тем не менее проблема была успешно решена Д. Келлером и Ц.
Мильштайном (1975). Исследователи получили гибриды иммунных В-лимфоцитов и
миеломных (опухолевых) клеток, которые обладали свойствами антителопродуцента и
"бессмертием" раково-трансформированной клетки. Такой вид клеток
получил название гибридом. В ходе дальнейшей селекции были отобраны
клоны с наивысшей продуктивностью и аффинностью специфических антител.
Гибридомные моноклональные антитела нашли широкое применение при создании
диагностических и лечебных иммунобиологических препаратов.
Полные и неполные антитела. Такое подразделение
основано на способности образовывать в реакции агглютинации или преципитации (in
vitro) хорошо различимый глазом результат. Таким свойством обладаютполные
антитела. К ним относятся IgМ, а также некоторые IgA и G.
Неполные антитела лишены такой способности,
несмотря на то, что они специфически связываются с антигеном - их еще называют
неагглютинирующими, непреципитирующими или блокирующими антителами.
4.
Антигенность антител
Иммуноглобулин, как и всякий белок, обладает антигенностью и
выраженной иммуногенностью. В молекуле Ig различают 4 типа атигенных
детерминант: видовые, изотипические, аллотипические и идиотипические. Видовые
антигенные детерминанты характерны для Ig всех особей данного вида
(например, кролика, собаки, человека). Они определяются строением легкой и
тяжелой цепей. По этим детерминантам можно идентифицировать видовую
принадлежность антител.
Изотипические антигенные детерминанты являются
групповыми. Они локализуются в тяжелой цепи и служат для дифференцировки Ig на
5 изотипов (классов) и множество подклассов.
Аллотипические антигенные детерминанты являются
индивидуальными, т.е. присущими конкретному организму. Они располагаются в
легкой и тяжелой полипептидных цепях. На основании строения аллотипических
детерминант можно различать особи внутри одного вида.
Идиотипические антигенные детерминанты отражают
особенности строения антигенсвязывающего центра самой молекулы Ig. Они
образованы V-доменами легкой и тяжелой цепей молекулы Ig. Обнаружение
идиотипических антигенных детерминант послужило основанием для создания теории
идиотип-антиидиотипической регуляции биосинтеза антител.
5.
Механизм взаимодействия антитела с антигеном
В процессе взаимодействия с антигеном принимает участиеантигенсвязывающий
центр молекулы Ig, или паратоп, который способен связываться со
строго определенной антигенной детерминантой. Эта связь осуществляется за счет
слабых взаимодействий (ван-дер-ваальсовы силы, водородные связи,
злектростатические взаимодействия) и отличается неустойчивостью -
образовавшийся иммунный комплекс (ИК) может легко диссоциировать: АГ + АТ ↔
ИК.
Аффинность - сила специфического взаимодействия
антитела с антигеном (или энергия их связи). Аффинность определяется степенью
стерического (пространственного) соответствия эпитопа и паратопа. Чем больше
образуется связей между эпитопом и паратопом, тем выше будут устойчивость и
продолжительность жизни образовавшегося иммунного комплекса. Иммунный комплекс,
образованный низкоаффинными антителами, чрезвычайно неустойчив и имеет малую
продолжительность существования.
Установлено, что в условиях макроорганизма с одной и той же
антигенной детерминантой способны одновременно прореагировать и образовать
иммунный комплекс около 100 различных клонов антител. Все они будут отличаться
структурой антигенсвязывающего центра, специфичностью и аффинностью. Аффинность
антител существенно меняется в процессе иммунного ответа в связи с селекцией
наиболее специфичных клонов В-лимфоцитов. Наименее аффинными считаются
нормальные антитела. По расчетам общее количество различных
антигенспецифических клонов В-лимфоцитов достигает 106-107.
Другой характеристикой Ig является авидность. Под этим
термином понимают прочность связывания антитела и антигена. Эта характеристика
определяется аффинностью Ig и числом антигенсвязывающих центров. Наибольшей
авидностью обладают антитела класса М, так как они имеют 10 антигенсвязывающих
центров.
Эффективность взаимодействия антитела с антигеном существенно
зависит от условий, в которых происходит реакция, прежде всего от pH среды,
осмотической плотности, солевого состава и температура среды. Оптимальными для
реакции антиген-антитело являются физиологические условия внутренней среды
макроорганизма: близкая к нейтральной реакция среды, присутствие фосфат-,
карбонат-, хлорид - и ацетат-ионов, осмолярность физиологического раствора
(концентрация раствора 0,15 М), а также температура 36-37°С.
6. Свойства
антител
Благодаря уникальной способности специфически связываться с
антигенными детерминантами антитела выполняют в организме ряд важнейших
функций.
К прямым эффектам антител относится нейтрализация - связывание
и блокирование паратопом иммуноглобулина активного центра биологически активной
молекулы, например токсина, рецептора, лекарственного препарата и пр. Эффект
имеет обратимый характер в случае распада иммунного комплекса. На этом принципе
основан механизм действия антитоксических, противовирусных и многих других
лечебных иммунных сывороток.
Другим прямым эффектом является энзиматическое действие
антител. Благодаря реликтовой протеазной или нуклеазной активности
иммуноглобулины способны вызывать деструкцию молекулы антигена (например,
расщепление отдельных пептидов или ДНК). Запуск системы комплемента по
классическому пути также представляет собой результат ферментолиза.
В большинстве случаев взаимодействие антител с антигеном в
организме не влечет за собой его структурную или функциональную модификацию.
Прочно связываясь с эпитопом, антитела маркируют молекулу антигена - обозначают
его как мишень для других факторов иммунитета (фагоцитоз, лизис).
К непрямым эффектам относятся:
индукция комплементопосредованного лизиса чужеродных клеток
(см. раздел 9.2.3.3), наилучшими свойствами обладает IgM (IgM > IgG3 >
IgG1);
запуск антителозависимой клеточно-опосредованной
цитотоксичности АЗКЦТ;
индукция гиперчувствительности немедленного, или I, типа;
опосредование иммунного фагоцитоза.
Клеточно-опосредованные эффекты иммуноглобулинов реализуются
благодаря экспрессии на мембране иммунокомпетентных клеток рецепторов к
Fc-фрагменту молекулы иммуноглобулина (FcR). Эти рецепторы являются
трансмембранными белковыми молекулами и различаются по специфичности к
определенному изотипу тяжелой цепи молекулы Ig. Различают также высокоаффинные
и низкоаффинные FcR. Первые могут взаимодействовать с интактной молекулой
иммуноглобулина. В некоторых случаях она используется как ко-рецепторный фактор
(базофилы, тучные клетки). Низкоаффинные FcRсвязываются уже с иммунным
комплексом, их называют непрямыми иммунорецепторами.
Помимо эффекторных свойств, антитела являются активными
регуляторами иммунореактивности. Так, Ig являются антигенспецифическими
рецепторами В-лимфоцитов.
Специфическое связывание эпитопов специфическими антителами
может блокировать развитие как гуморального, так и клеточного иммунного ответа.
Этот эффект используется в клинической практике, например, для профилактики
гемолитической болезни новорожденных в результате резус-конфликта. Антитела,
специфичные к идиотипическим антигенным детерминантам Ig, могут управлять силой
антительного иммунного реагирования.
7. Генетика
иммуноглобулинов
Для структуры молекул Ig характерно уникальное генетическое
кодирование. Методами молекулярной генетики было доказано, что структура
молекулы Ig контролируется большим числом генов, которые имеют фрагментарную
организацию, образуют три группы, располагаются в трех различных хромосомах и
наследуются независимо.
Первая группа генов кодирует первичную структуру легкой цепи λ-типа, вторая - легкой цепи κ-типа, а третья - всех
типов тяжелых цепей (α, δ, ε, γ и μ). Гены, относящиеся к каждой группе, находятся на соответствующей
хромосоме в непосредственной близости друг от друга, располагаются
последовательно и разделены интронами.
Участок ДНК, кодирующий строение легкой цепи λ-типа, содержит 2 V-сегмента (контролируют структуру
V-доменов) и 4 C-сегмента (контролируют структуру C-доменов). Между C -
и V-сегментами располагается J-сегмент (от англ. join - соединяющий).
Легкая цепь κ-типа кодируется несколькими сотнями
V-сегментов ДНК, 4 J-сегментами и одним C-сегментом. Группа генов,
контролирующая структуру тяжелых цепей, имеет еще более сложное строение.
Наряду с V-, C - и J - сегментами ДНК

Рис. 3. Схема строения генов иммуноглобулинов
(пояснение в тексте)
в их состав входят 20 D-сегментов (от англ. divercity
- разнообразие). Кроме того, имеется M-сегмент, который кодирует
биосинтез мембраноассоциированного участка молекулы рецепторного Ig.
Созревание пре-В-лимфоцитов сопровождается перестройками в их
генетическом аппарате. Происходят произвольное сближение отдельных фрагментов
ДНК и сборка в пределах соответствующих хромосом единых функциональных генов.
Этот процесс называется сплайсинг (от англ. splicing - сращивание,
состыковывание). Пропущенные участки ДНК исключаются из дальнейшего считывания.
С функциональных генов в дальнейшем транскрибируется про-мРНК, а затем
окончательная мРНК, кодирующая первичную аминокислотную последовательность L -
и H-цепей молекулы Ig. Параллельно со сплайсингом в отдельных участках
V-сегментов генов иммуноглобулинов могут происходить точечные мутации и
нематричная достройка олигонуклеотидов. Эти участки ДНК получили название гипермутабельных
областей.
Сплайсинг и мутационный процесс в генах Ig носят случайный
характер. Они происходят в каждом лимфоците независимо друг от друга и
уникальны, что в бесконечное количество раз повышает разнообразие V-доменов и в
конечном счете структуры паратопов и идиотипических антигенных детерминант
молекулы Ig. Поэтому в организме всегда существуют или в любой момент могут появиться
В-лимфоциты, специфичные практически к любому антигену. Этот тезис составляет
основу молекулярно-генетической теории
происхождения многообразия специфичностей антител,
разработанной С. Тонегавой (1983).
В процессе первичного иммунного ответа размножение
В-лимфоцитов также сопровождается рекомбинационными перестройками в пределах
иммуноглобулиновых генов, но уже в пределах С-сегментов. Это проявляется
последовательной сменой класса Ig: на ранних этапах дифференцировки В-лимфоциты
синтезируют Ig классов М и D, на более поздних - классов G, A или E (редко).
8. Динамика
антителопродукции
Иммунная система реагирует на появление во внутренней среде
макроорганизма антигена усилением биосинтеза специфических антител. Это
достигается размножением клонов антигенспецифических
клеток-антителопродуцентов. При этом антиген выступает в роли как пускового,
так и селектирующего фактора: преимущественно активируются клоны с наивысшей
специфичностью, т.е. наибольшей аффинностью рецепторных молекул Ig. Параллельно
с размножением идет процесс дифференцировки В-лимфоцитов. Наблюдаются
перестройка в геноме клеток и переключение их биосинтеза с крупной
высокоавидной молекулы IgM на более легкие и экономичные высокоаффинные IgG или
IgA.
Антителопродукция в ответ на антигенный стимул имеет
характерную динамику. Ее можно проследить на примере сывороточных Ig (рис.4).
Выделяют латентную (индуктивную), логарифмическую, стационарную фазы и фазу
снижения. В латентную фазу антителопродукция практически не изменяется и
остается на базальном уровне.
В этот период происходят переработка и представление антигена
иммунокомпетентным клеткам и запуск пролиферации антигенспецифических клонов
клетокантителопродуцентов. Ввиду того что клетки делятся дихотомически (т.е.
надвое), прирост их количества происходит в логарифмической зависимости и
поэтому после первых циклов деления оно изменяется незначительно.
Параллельно происходят дифференцировка пре-В-лимфоцитов в
зрелые формы и плазматические клетки и переключение синтезируемых изотипов Ig. Во
времялогарифмической фазы наблюдается интенсивный прирост количества
антигенспецифических В-лимфоцитов, что находит отражение в существенном
нарастании титров специфических антител. Встационарной фазе количество
специфических антител и синтезирующих их клеток достигает максимума и
стабилизируется. Освобождение макроорганизма от антигена устраняет антигенный
стимул, поэтому вфазе снижения наблюдается постепенное уменьшение
количества клонов специфических антителопродуцентов и титров соответствующих
антител.

Рис. 4. Динамика антителообразования при первичном (I) и
вторичном (II) иммунном ответах. Фазы антителообразования: а - латентная; б -
логарифмического роста; в - стационарная; г - снижения
Динамика антителообразования существенно зависит от
первичности или вторичности контакта с антигеном. При первичном контакте с
антигеном развивается первичный иммунный ответ. Для него характерны
длительные латентная и логарифмическая (7-15 сут) фазы. Первые диагностически
значимые титры специфических антител регистрируются на 10-14-е сутки от момента
иммунизации. Стационарная фаза продолжается 15-30 сут, а фаза снижения - 1-6
мес.
В течение первичного иммунного ответа происходят созревание,
размножение клонов и дифференцировка антигенспецифических В-лимфоцитов, а также
переключение биосинтеза Ig с изотипа M на изотопы G, A или Е. В итоге
первичного иммунного реагирования формируются многочисленные клоны
антигенспецифических антителопродуцирующих клеток и клеток иммунологической памяти,
а во внутренней среде макроорганизма в высоком титре накапливаются
специфические IgG и/или IgA. Таким образом обеспечиваются активное
противодействие иммунной системы внедрению в макроорганизм антигена и высокая
готовность к повторной с ним встрече.
Со временем антительный ответ угасает. Элиминация антигена
исключает новое стимулирование к клонообразованию, а появившиеся ранее
плазматические клетки имеют короткую продолжительность жизни. Вместе с тем
В-лимфоциты иммунологической памяти надолго остаются циркулировать в организме.
Повторный контакт иммунной системы с тем же антигеном ведет к
формированию вторичного иммунного ответа. Его латентная фаза значительно
укорочена, а логарифмическая фаза отличается более интенсивной динамикой
прироста и более высокими титрами специфических антител. Для стационарной фазы
и фазы снижения свойственна затяжная динамика (несколько месяцев или даже лет).
При вторичном иммунном ответе организм сразу же в подавляющем большинстве
синтезирует IgG. Это обусловлено подготовленностью иммунной системы к повторной
встрече с антигеном за счет формирования иммунологической памяти:
многочисленные клоны антигенспецифических В-лимфоцитов, оставшиеся после
первичного иммунного реагирования, быстро размножаются и интенсивно включаются
в процесс антителогенеза.
Для развития гуморального иммунитета слизистых оболочек
характерны те же процессы и динамика антителообразования. Однако в данном
случае в слизистых оболочках в подавляющем большинстве созревают и размножаются
В-лимфоциты, продуцирующие полимерные молекулы IgA.
Явление интенсивного антителообразования при повторном
контакте с антигеном широко используется в практических целях, например, вакцинопрофилактике.
Для создания и поддержания иммунитета на высоком защитном уровне схемы вакцинации
предусматривают многократное введение антигена для формирования и поддержания
иммунологической памяти.
Этот же феномен используют при получении высокоактивных
лечебных и диагностических иммунных сывороток (гипериммунных). Для этого
животным или донорам производят многократные введения препаратов антигена по
специальной схеме.
Динамика и интенсивность антителообразования в значительной
степени зависят от иммуногенности антигена: дозы, способа и кратности его
введения, а также от состояния макроорганизма. Попытка повторного введения
антигена в латентной фазе может привести к иммунологическому параличу -
иммунологической неотвечаемости на антиген в течение определенного периода
времени.
. Теории
разнообразия антител
Для объяснения механизмов антителопродукции и разнообразия
специфичности антител было предложено множество гипотез и теорий. Только
немногие из них получили практическое подтверждение, большинство представляет
исторический интерес.
Первую принципиально важную концепцию боковых цепей выдвинул
П. Эрлих (1898). Согласно этой концепции, клетки органов и тканей имеют на
своей поверхности рецепторы, способные в силу химического сродства связывать
антиген и инактивировать его. Затем они отделяются с поверхности клетки и
замещаются вновь синтезированными. Эта теория заложила основные представления о
гуморальном иммунитете и рецепторах иммунокомпетентных клеток.
Заслуживают внимания инструктивные или матричные теории.
Согласно концепциям, предложенным Ф. Брейнлем и Ф. Гауровитцем (1930), Л.
Полингом (1940), антиген является матрицей, с которой штампуется молекула
антител. Эти теории оказались тупиковыми в связи с открытием Д. Уотсоном и Ф.
Криком (1953) механизма кодирования в ДНК генетической информации.
Ряд теорий исходил из предположения о предсуществовании в
организме антител практически ко всем возможным антигенам (Ерне Н., 1955;
Бернет Ф., 1959). В настоящее время наиболее обоснованной считается теория Ф.
Бернета, которая получила название клонально-селекционной. Согласно
данной теории, лимфоидная ткань состоит из огромного числа клонов
антигенореактивных лимфоцитов, которые специализируются на выработке антител к
определенным антигенам. Клоны уже предсуществуют в новорожденном организме.
Попавший в организм антиген селективно (избирательно) активирует специфичный к
нему клон лимфоцитов, который размножается и начинает вырабатывать специфичные
к данному антигену антитела. Если доза антигена слишком велика, то клон
реагирующих на него лимфоцитов устраняется (элиминируется) из организма - так в
эмбриональном периоде формируется иммунологическая толерантность
(нечувствительность) к собственным антигенам.
Теория Бернета объясняет многие иммунологические реакции
(антителообразование, гетерогенность антител, иммунологическую память,
толерантность), однако она не способна объяснить происхождение всего
многообразия специфичности антител. Бернет предположил, что в организме
существует около 10 тыс. клонов специфических антителопродуцирующих клеток.
Однако мир антигенов оказался на 2-3 порядка обширнее, и организм отвечает на
практически любой из них, в том числе и на искусственно полученный,
несуществующий в природе.
Значительную ясность в представление о разнообразии
специфичности антител внес С. Тонегава (1983), который дал этому явлению
генетическое обоснование. Молекулярно-генетическая теория С. Тонегавы исходит
из того, что в генах иммуноглобулинов постоянно происходят мощные
рекомбинационные и мутационные процессы. В результате возникает огромное
количество вариантов и комбинаций генов, которые кодируют разнообразные
специфичности иммуноглобулинов. Каждый клон антителопродуцирующих лимфоцитов
обладает своим уникальным вариантом гена иммуноглобулина.
Следует также упомянуть теорию сетевой регуляции иммунной
системы. Ее основой является выдвинутая Н. Ерне (1974) идея
идиотип-антиидиотипического взаимодействия. Согласно этой теории, иммунная
система представляет собой бесконечную цепь взаимодействующих антигенных
идиотипов иммуноглобулинов и направленных к ним антиидиотипических антител.
Введение антигена вызывает каскадную реакцию образования антител 1-го порядка.
Это антитело, действуя как антиген, вызывает образование к своему идиотипу
антител 2-го порядка. К идиотипу антител 2-го порядка синтезируются антитела
3-го порядка и т.д. При этом антитело каждого порядка как бы несет внутренний
образ антигена, который передается эстафетно в цепи образования
антиидиотипических антител. Доказательством этой теории являются обнаружение
антиидиотипических антител, способных вызвать в организме иммунитет к соответствующему
антигену, а также существование лимфоцитов, сенсибилизированных к
антиидиотипическим антителам. С помощью теории Ерне можно понять формирование
иммунологичской памяти и возникновение аутоиммунных реакций. Однако она не
способна объяснить много других явлений иммунитета: механизм иммунологического
распознавания "свой-чужой", управления каскадом
идиотип-антиидиотипических реакций и т.д. Данная теория не получила дальнейшего
развития.
Выдающийся отечественный иммунолог П.Ф. Здродовский в 60-е
годы XX столетия сформулировал физиологическую концепцию иммуногенеза -
гипоталамо-адреналовую теорию регуляции иммунитета. Основная идея его теории
сводилась к тому, что продукция антител подчиняется общим физиологическим
законам. Ведущая роль в этом процессе принадлежит гормонам и нервной системе.
Заключение
К любым антигенам в организме животного изначально существуют
антитела. Это предполагает, что каждый организм продуцирует миллионы различных
иммуноглобулинов, различающихся своими центрами связывания антигенов. Такое
разнообразие обеспечивается несколькими механизмами. Лёгкие и тяжёлые цепи
молекул антител кодируются несколькими типами генных сегментов: лёгкая цепь -
тремя типами сегментов (V, J, C), тяжёлая - четырьмя (V, D, J, C). В геноме
обычно присутствует от нескольких до нескольких сотен сегментов каждого типа,
несколько различающихся по нуклеотидной последовательности. Для синтеза
цельного полипептида (лёгкой или тяжёлой цепи) необходимо объединение
нуклеотидных последовательностей сегментов каждого типа. Такое объединение
происходит сначала на уровне ДНК (соматическая рекомбинация), а затем на уровне
матричных РНК (сплайсинг). В результате образуется огромное количество
вариантов мРНК и соответственно полипептидных цепей. Во время соматической
рекомбинации и сплайсинга могут происходить вставки и делеции нуклеотидов, что
вместе с повышенной частотой мутаций в генах антител ещё больше повышает
разнообразие этих уникальных по своим свойствам белков.
Библиографический
список
1.
Иммунология / Ройт А., Бростофф Дж., Мейл Д. - М.: Мир, 2000. - 592 с.
.
Иммунология: В 3-х т.; т.1/Под ред.У. Пола. - М.: Мир, 1987-88. - 476 с.
.
http://vmede.org/sait/?
id=Mikrobiologija_3verev_2010_t1&menu=Mikrobiologija_3verev_2010_t1&page=11