Физиология человека и животных
Сургутский
Государственный Университет
Контрольная
работа
по
физиологии человека и животных
Выполнил: ст.
Шабанова Патимат Гаджиевна
Сургут,
2011
1. Проведение возбуждения по нервным волокнам
Нервные волокна - это отростки нервных клеток. В
состав нерва входят длинные отростки нервных клеток (аксоны), которые несут
возбуждение в центральную нервную систему или от нее на периферию. Нервы, как
правило, являются смешанными. В их состав входят двигательные и чувствительные
волокна.
Нервные волокна, как и все возбудимые структуры,
обладают следующими физиологическими свойствами: возбудимостью, проводимостью,
рефрактерностью, лабильностью. При сравнении основных, физиологических свойств
нервной и мышечной ткани установлено, что возбудимость и лабильность нервного
волокна выше, а рефрактерный период короче, чем в мышечной ткани. Это связано с
более высоким уровнем обменных процессов в нерве.
Проводимость - способность живой ткани проводить
волны возбуждения - биоэлектрические импульсы.
Для обеспечения гомеостатического единства все
структуры организма (клетки, ткани, органы и т.д.) должны иметь возможность
пространственного взаимодействия. Распространение возбуждения от места его
возникновения до исполнительных органов - один из основных способов такого
взаимодействия. Возникший в месте нанесения раздражения потенциал действия
является причиной раздражения соседних, невозбужденных участков нервного (или
мышечного) волокна. Благодаря этому явлению волна потенциала действия создает
ток действия, который распространяется по всей длине нервного волокна. В
безмиелиновых нервных волокнах возбуждение проводится с некоторым затуханием -
декрементом, а в миелиновых нервных волокнах - без затухания. Проведение
возбуждения также сопровождается изменением обмена веществ и энергии.
Проведение возбуждения, или нервных импульсов,
является специализированной функцией нервных волокон. Скорость проведения
возбуждения в основном зависит от диаметра и гистологических особенностей
строения нервных волокон. Чем больше диаметр нервного волокна, тем скорость
распространения возбуждения в нем выше. Так, скорость распространения
возбуждения по нервному волокну с диаметром 12-22 мкм составляет 70-120 м/с, а
с диаметром 8-12 мкм - 40-70 м/с.
В зависимости от гистологического строения
нервные волокна делятся на миелиновые (мякотные) и безмиелиновые (безмякотные).
Миелиновое волокно (рис. 53) состоит из осевого
цилиндра и покрывающей его миелиновой и шванновской оболочек. Миелиновая
оболочка состоит из жироподобных веществ, имеет высокое удельное сопротивление
и выполняет роль изолятора. Миелиновая оболочка через промежутки равной длины
прерывается, оставляя открытыми участки осевого цилиндра шириной около 1 мкм.
Эти участки называются перехватами узла (перехваты Ранвье). Длина
межперехватных участков зависит от диаметра волокна и колеблется в пределах от
0,2 до 1-2 мм. Поверхность осевого цилиндра представлена плазматической
мембраной, а его содержимое аксоплазмой.
Безмякотные нервные волокна не имеют миелиновой
оболочки, они покрыты только леммоцитами (шванновскими клетками).
Между леммоцитами и осевым цилиндром имеется
щель в 15 нм (150 А), заполненная межклеточной жидкостью. В связи с этим
поверхностная мембрана осевого цилиндра сообщается с окружающей нервное волокно
средой (межклеточной жидкостью). Основоположником учения о проведении
возбуждения по нервным волокнам является немецкий физиолог Герман (1385). Он
полагал, что возбуждение по нервным волокнам распространяется за счет малых
круговых токов, которые возникают внутри волокна и в окружающей его жидкости
(рис. 54).
нерв волокно рефлекс сетчатка


Между возбужденным и невозбужденным участками
нервного волокна в аксоплазме и в окружающей жидкости ток распространяется от
положительно заряженного участка к отрицательно заряженному. Это приводит к
возникновению так называемых малых, или круговых, токоз, которые, выходя из
нервного волокна, последовательно возбуждают его участки (1, 2 и т. д.). По
мере удаления от очага возбуждения (участки 3 и 4) раздражающее действие
круговых токов ослабевает и они становятся неспособными вызвать возбуждение.
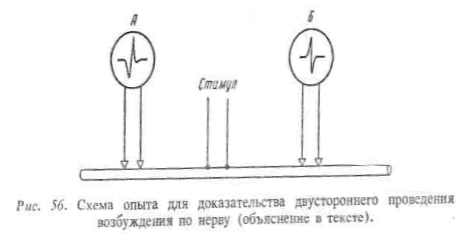
При нанесении раздражения на нервное волокно
происходит двустороннее распространение возбуждения в центростремительном и
центробежном направлениях (закон двустороннего проведения возбуждения по
изолированному нервному волокну). Это доказывается следующим опытом. К нервному
волокну прикладывают две пары электродов А и Б, связанных с
электроизмерительными приборами (рис. 56), Раздражение наносят между
электродами А и Б с помощью раздражающих электродов (стимул). В результате
двустороннего проведения возбуждения вдоль клеточной мембраны приборы
зарегистрируют прохождение нервных импульсов как под электродом А, так и под
электродом Б.
Двустороннее проведение возбуждения по нервному
волокну впервые описано русским ученым Р. И. Бабухиным (1887) и Клоне (1886).
Оно не противоречит одностороннему распространению нервных импульсов в
целостном организме, которое объясняется местом возникновения нервных импульсов
(рецептор или нервный центр), а также наличием специальных образований -
синапсов.
Возбуждение проводится только по одному нервному
волокну, не распространяясь на соседние волокна, что обусловливает
осуществление строго координированной рефлекторной деятельности (закон
изолированного проведения возбуждения по нервному волокну). Периферический
нервный ствол обычно состоит из большого количества нервных волокон. Так, в
состав седалищного нерва входят тысячи нервных волокон: мякотные и безмякотные,
афферентные и эфферентные, соматические и вегетативные. В случае
неизолированного проведения возбуждения наблюдалась бы хаотическая ответная
реакция. Изолированное проведение возбуждения в мякотных нервных волокнах
обеспечивается миелиновой оболочкой, в безмякотных - высоким удельным
сопротивлением окружающей нервное волокно жидкости. Доказать наличие
изолированного проведения возбуждения можно в эксперименте на препарате задней
лапки лягушки, раздражая отдельные корешки седалищного нерва. Более строгое
доказательство может быть получено при отведении тока действия от отдельных
нервных волокон, входящих в состав нервного ствола.
Утомление нервного волокна. Н.Е. Введенский в
1883 г. в опытах на нервно-мышечном препарате лягушки впервые установил, что
нерв мало утомляем. Раздражая седалищный нерв индукционным током в течение 6-8
ч (о возбуждении нерва он судил по сокращению мышцы), он обнаружил, что нерв
длительно (на протяжении многих часов) сохранял способность возбуждаться и
проводить возбуждение. Данные экспериментов Н.Е. Введенского подтверждены
современными исследованиями в опытах с регистрацией биотоков нерва. Малая
утомляемость нервных волокон объясняется тем, что энергетические затраты в них
при возбуждении незначительны, а восстановительные процессы протекают быстро.
В целостном организме малой утомляемости нервных
волокон способствует их работа с постоянной недогрузкой. Так, двигательное
нервное волокно обладает высокой лабильностью (может воспроизводить до 2500
импульсов в 1 с, из нервных же центров на периферию обычно проводится
прерывисто не более 50-100 волн возбуждения в 1 с). Таким образом, практическая
неутомляемость нервных волокон связана с низкими энергетическими затратами при
возбуждении, с высокой лабильностью нервных волокон и постоянной работой их с
недогрузкой [1].
. Условные рефлексы
Рефлекс - это осуществляемая при участии нервной
системы ответная реакция организма на раздражение, исходящее из внешней или
внутренней среды.
Рефлексы являются приспособительными реакциями
живого организма в осуществлении его взаимосвязей с внешней средой, а также
взаимодействия между его органами и системами; эти реакции обеспечивают
целостность организма и постоянство его внутренней среды (рефлекторная
регуляция давления крови, уровня глюкозы в крови и т. п.).
Рефлексы лежат в основе деятельности нервной
системы. По И.М. Сеченову, “все акты сознательной и бессознательной жизни по способу
своего происхождения - суть рефлексы”.
Каждый рефлекс осуществляется при помощи
рефлекторной дуги, строение которой отражает этапы филогенетического развития
данной реакции. В процессе филогенеза наиболее простыми и древними формами
являются спинальные рефлексы безусловно-рефлекторных двигательных реакций.
Каждая рефлекторная дуга состоит из афферентного
(чувствительного) звена, начинающегося рецепторным аппаратом, и эфферентного
(двигательного), заканчивающегося рабочим органом (эффектором). Довольно часто
между двумя указанными звеньями имеются один-два вставочных нейрона,
воспринимающих импульсы от рецепторных аппаратов и перерабатывающих их в
центробежные импульсы, идущие к исполнительному органу.
В вегетативной нервной системе рефлекторные
эффекты могут иметь место и при наличии лишь одного нейрона. Имеется в виду
аксон-рефлекс, осуществляемый без участия центральной нервной системы и
представляющий собой возбуждение одной ветви аксона с распространением
проксимально до места ветвления и затем снова дистально по другой его ветви.
Различают простые и сложные, приобретенные и
врожденные, безусловные и условные рефлексы.
Безусловные рефлексы - это врожденные,
наследственно закрепленные рефлексы, выработанные в процессе филогенеза,
условные - это непостоянные, индивидуальные рефлексы, приобретенные в
онтогенезе в результате взаимодействия организма с внешней средой, выработанные
на базе безусловных рефлексов.
Интеграция и сложное переплетение безусловной
рефлекторной и условно-рефлекторной деятельности создают единую целостную
картину поведения, в частности двигательного поведения - непроизвольных, а
также произвольных целенаправленных двигательных актов, приобретенных в течение
индивидуальной жизни (профессиональные
<#"564362.files/image004.gif">
Рефлекторная дуга - путь нервного импульса от
рецепторов до рабочего органа. От рабочего органа в центральную нервную систему
опять поступают нервные импульсы - это обратная связь. Рефлекторная дуга -
сложный структурно-функциональный комплекс, замкнутый в кольцо. Она состоит <#"564362.files/image005.jpg">
Рис. Зрительный анализатор
- Поле зрения; 2 - Сетчатка; 3 - Зрительный
нерв; 4 - Зрительный перекрест (хиазма); 5 - Левый зрительный тракт (волокна,
идущие от левых половин сетчатки); 6 - Правый зрительный тракт (волокна, идущие
от правых половин сетчатки); 7 - Наружные коленчатые тела (подкорковый центр -
первичная обработка зрительной информации); 8 - Четверохолмие (подкорковая
регуляция работы органа зрения); 9 - Проводящие волокна; 10 - Комиссурные
(межполушарные) волокна; 11 - зрительный центр коры больших полушарий.
Сетчатка - это внутренняя оболочка глаза,
имеющая сложную многослойную структуру. Она содержит два вида различных по
своему функциональному значению фоторецепторов (палочки, колбочки и несколько
видов нервных клеток). Под воздействием света в фоторецепторах идут
фотохимические реакции, суть которых заключается в изменении структуры
светочувствительных зрительных пигментов. Последнее вызывает возбуждение
фоторецепторов и последующее синаптическое возбуждение связанных с ними нервных
клеток. Нервный аппарат глаза передает зрительную информацию в виде
электрических импульсов в центры головного мозга. Поэтому сетчатку можно
рассматривать как бы частью мозга, вынесенной на периферию. Кратко рассмотрим
структуру и функцию всех слоев сетчатки.
Основы физиологии сетчатки.
Показано, что различные цвета воспринимаются
лучше всего при действии световых раздражителей на центральную ямку, где
расположены в основном колбочки. К периферии сетчатки восприятие цвета
становится хуже. Периферия сетчатки, где находятся только палочки, не способна
воспринимать цвета. Чувствительность к свету колбочкового аппарата сетчатки в
несколько раз меньше аппарата палочек. Потому в условиях слабой освещенности,
(центральное) колбочковое зрение резко снижается. Преобладающим становится
периферическое (палочковое) зрение. В связи с неспособностью палочек
воспринимать цвета, в сумерках человек цвета не различает. Нарушение функций
палочек, возникающее, при недостатке в пище витамина А, вызывает расстройство
сумеречного зрения, так называемую (куриную слепоту) человек слепнет в
сумерках, а днем зрение сохраняется нормальным.
Слепое пятно и нейроны сетчатки, ганглиозные
клетки.
Место входа зрительного нерва в глазное яблоко -
называют слепым пятном, оно не содержит фоторецепторов и нечувствительно к
свету. Внутри, от слоя фоторецепторных клеток, расположен слой биполярных
нейронов, к ним изнутри примыкает слой ганглиозных нервных клеток. Волокна
зрительного нерва образованы аксонами ганглиозных клеток. Возбуждение,
возникающее в колбочке или палочке при действии света, передается на волокна
зрительного нерва через нервные клетки (биполярные и ганглиозные). Передача
импульса с одной клетки на другую совершается с помощью медиатора ацетилхолина.
На 130 миллионов фоторецепторных клеток приходится всего около 1,25 миллионов
волокон зрительного нерва. Значит, импульсы от многих фоторецепторов сходятся к
одной ганглиозной клетке. Показано, что один биполярный нейрон связан со
многими палочками и несколькими колбочками, а одна ганглиозная клетка в свою
очередь связана с несколькими биполярными клетками. Из этого понятно, что
каждая ганглиозная клетка суммирует сигналы, возникающее в большом числе
фоторецепторов. Лишь в районе центральной ямки, каждая колбочка соединена с
одной (карликовой) биполярной клеткой, с которой в свою очередь соединена всего
одна ганглиозная клетка.
Реакции в сетчатке.
Примерно в конце 70-х годов прошлого столетия в
сетчатке глаза были открыты светочувствительные пигменты. В палочках сетчатки
человека содержится пигмент родопсин. Колбочки содержат пигмент родопсин, а
также пигменты хлоролаб и эритролаб; первый из них поглощает лучи, соответствующие
зеленой, а второй - красной части спектра. Родопсин состоит из ретиналя -
альдегида витамина А и белка опсина. Под воздействием кванта света происходит
цикл фотофизических и фотохимических реакций: ретиналь изомеризуется, его
боковая цепь выпрямляется, связь ретиналя с белком изменяется, активируются
ферментативные центры белковой молекулы. Конечным этапом превращений является
отщепление ретиналя от опсина. Под влиянием фермента редуктазы ретиналя,
последний переходит в витамин А. При затемнении глаз происходит регенерация
родопсина. Фотохимические процессы в сетчатке происходят крайне экономно, т. е.
при воздействии даже яркого света дегенерирует только малая часть имеющегося
родопсина. Так при действии света интенсивностью 100 лк через 5 секунд в
палочке дегенерирует около 0,006% родопсина.
Фоточувствительные клетки сетчатки ipRGC (RGC -
Retinal ganglion cell) - третий, помимо колбочек и палочек, и относительно
недавно обнаруженный (1991 г.) вид фоторецепторов, которые содержатся в
сетчатке глаза.
Светочувствительные клетки типа ipRGC содержат
меланопсин, светочувствительный пигмент в клетках-фоторецепторах глаза
млекопитающих. В отличие от других нервных ганглиарных клеток, относящихся к
сетчатке глаза, они являются фоточувствительными. Это означает, что они -
третий класс фоторецепторов сетчатки глаза, которые возбуждаются под действием
света. Возбуждаются даже при блокировании «классических» фоторецепторов,
палочек и колбочек, под действием фармакологических агентов, или отделением их
от сетчатки. По сравнению с колбочками и палочками, клетки ipRGC реагируют
более вяло; они сигнализируют о присутствии света в течение достаточно
длительного времени. Их основная функциональная роль - вовсе не
непосредственное создание оптических изображений, чем они существенно
отличаются от палочек и колбочек, которые расположены в сетчатке на фокальной
поверхности и образуют оптическое изображение. В отличие от палочек и колбочек
они обеспечивают устойчивое представление об окружающей световой, цветовой информации
- о величине интенсивности падающего светового потока.
Установлено, что клетки типа ipRGC выполняют по
крайней мере три функции:
) Они играют главную роль в синхронизации
циркадные ритмы в течение суточного светового/темнового цикла, обеспечивая прежде
всего оценку дневного и вечернего вида световой и цветовой информации. Они
посылают информацию об освещённости через ретиногипоталамический тракт
непосредственно циркадному лидеру мозга. Физиологические свойства этих нервных
клеток определяются известной функцией ежедневной светосинхронизации и являются
механизмом, регулирующим циркадные ритмы.
) Фоточувствительные нервные ячейки также
возбуждают другие мозговые области, регулируют диафрагмирование зрачка, нуклеус
среднего мозга. Таким образом, они вносят свой вклад в регулирование размера
зрачка; в мозгу на другие ответные реакции поведенческого типа на возникающие
условия освещения в окружающей среде.
) Они участвуют в светорегулировании и в
процессе подавления «засветок» от внезапного слишком сильного освещения (взгляд
на прямое солнечное освещение, электросварку, луч прожектора и т. д.) благодаря
выделению гормона - мелатонина из шишковидной железы.
Использованная литература
1.
Физиология / Под ред. С.А. Георгиевой. - 2-е изд. - Ф48 М.: Медицина, 1986. -
400 с.
.
Беритов И.С. Общая физиология мышечной и нервной системы, т. 2. - М., 2001. -
643 c.
.
Недоспасов В.О. Физиология центральной нервной системы. - М., 2002 г. - 377 c.
.
Основы физиологии.\ под ред.П. Стерки. - М.: Мир,1984 г.
.
Физиология человека.\ под ред. Р. Шмидта и Г. Тевса. - М.: Мир, 1996 г. - 330
c.
.
<http://21.bewell.ru/m_meh.htm>
.
<http://medivin.ru/default_1309.html>
.
http://ru.science.wikia.com/wiki/Фоточувствительные_клетки_сетчатки_
ipRGC