|
Рис. 102. Липиды архебактерий: 1 — фитаниловый
диэфир глицерина; 2 — бифитаниловый тетраэфир диглицерина; 3 — изопреноид
из полярных липидов, содержащий пятичленные циклические группировки; обведены
полярные области; R — остатки фосфорной кислоты или сахара
|
|
|
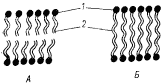
Риc. 103. Схема бислойной и монослойной мембран
архебактерий, образованных соответственно ди- (А) и тетраэфирами (Б)
глицерина: 1 — молекула глицерина; 2 — углеводородные цепи
разной длины (по Воробьевой, 1987)
|
Существенные отличия
выявлены у архебактерий в строении генома, аппаратов репликации, транскрипции и
трансляции. Прежде всего исследователи обратили внимание на то, что именно в
группе архебактерий наименьший геном среди свободноживующих форм прокариот: у Thermoplasma
acidophilum — 0,8x109 Да, изученных метанобразующих бактерий —
порядка 1x109 Да (для сравнения молекулярная масса генома Е. coli
— 2,5x109 Да). Однако у галобактерий величина генома оказалась
больше, чем у Е. coli. Особенность генома архебактерий — наличие
многократно повторяющихся нуклеотидных последовательностей, а в генах,
кодирующих белки, тРНК и рРНК, — интронов, что характерно для организации
генетического материала эукариот. У некоторых архебактерий обнаружены основные
гистоноподобные белки, связанные с ДНК. Функция их предположительно заключается
в обеспечении определенной упаковки ДНК в нуклеоиде.
Помимо хромосомной ДНК в
клетках архебактерий обнаружены типичные для эубактерий фаги, плазмиды,
мигрирующие элементы.
Существование механизмов
переноса генетической информации с помощью фагов и плазмид позволяет
предполагать, что архебактерии должны каким-то образом защищать собственный
генетический материал от чужеродного. У эубактерий эта проблема решена с
помощью системы рестрикции-модификации. У эукариот такой системы нет, они
выработали иные механизмы генетической изоляции. Найдено, что архебактерии
обладают системой рестрикции-модификации, аналогичной эубактериальной.
Информация об аппарате
репликации архебактерий в основном ограничивается данными о выделенной из
ограниченного числа видов ДНК-зависимой ДНК-полимеразе, по некоторым свойствам
близкой к эукариотному типу. Генетический код архебактерий такой же, как у
других организмов.
ДНК-зависимая
РНК-полимераза архебактерий сочетает свойства, характерные для эукариот и
эубактерий. У всех изученных представителей архебактерий РНК-полимераза одной
формы, осуществляющая, как и в случае эубактерий, транскрипцию всех генов. (У
эукариот, например дрожжей, существуют 3 формы РНК-полимеразы, различающиеся
функционально, компонентным составом, чувствительностью к ингибиторам). Фермент
архебактерий отличается структурной сложностью, в его состав входят от 5 до 11
отдельных субъединиц. (РНК-полимеразы эубактерий состоят из 4 — 8 компонентов,
а эукариот — 10 — 14 субъединиц). Характерным для РНК-полимераз всех эубактерий
является их чувствительность к антибиотикам, специфически ингибирующим
инициацию (рифампицин) и элонгацию (стрептолидигин 1) транскрипции.
Архебактериальная и все РНК-полимеразы эукариот не чувствительны к этим
антибиотикам.
Недавно у архебактерий
описан известный только у эукариот процессинг: вырезание из первичного продукта
транскрипции определенных нуклеотидных участков, укорачивание и образование
зрелых молекул РНК.
Процесс трансляции у
архебактерий происходит по тому же принципиальному пути, что и у других
организмов, но обнаружены многочисленные особенности в организации трансляционного
аппарата. Рибосомы архебактерий сочетают свойства, присущие эубактериям и
эукариотам: по размерам они схожи с рибосомами эубактерий (имеют константу
седиментации 70S, а их субъединицы — 30S и 50S), по форме ближе к 80S рибосомам
эукариот.
Состав рибосомальных РНК
архебактерий типично эубактериальный (5S, 16S и 23S рРНК), но их первичные
структуры отличны от эубактериальных и эукариотных. Изучение нуклеотидных
последовательностей 16S (18S) рРНК разных представителей живого мира и привело
к выявлению среди прокариот группы архебактерий. Значения коэффициента сходства
(SAB), отделяющие рРНК эубактерий, архебактерий и эукариот друг от
друга, лежат в области 0,1 (SAB, равный 1, соответствует полной
гомологии нуклеотидных последовательностей; SAB порядка 0,02 —
уровень случайного совпадения).
Архебактериальные 5S
рРНК по нуклеотидной последовательности также заметно отличаются от
соответствующих рРНК эубактерий и эукариот. Вторичные структуры этих РНК у
различных представителей архебактерий проявляют наличие эубактериальных,
эукариотных и уникальных черт в разных соотношениях и по своему разнообразию
охватывают широкий спектр структур от типично эубактериальной до типично
эукариотной.
Количество рибосомальных
белков у архебактерий больше, чем у эубактерий, но меньше, чем у эукариот.
Получены данные, свидетельствующие об уникальности первичной структуры ряда
рибосомальных белков архебактерий. В то же время наиболее интенсивно изучаемый
рибосомальный белок А архебактерий по аминокислотной последовательности сходен с
соответствующим белком эукариот. На 70S архебактериальной рибосоме отсутствуют
места связывания ингибиторов, специфичных для 70S рибосом эубактерий
(хлорамфеникол, стрептомицин), но есть места для связывания ингибиторов,
специфических в отношении 80S рибосом эукариот.
Для тРНК архебактерий
характерно сходство с остальными организмами по общим принципам организации, но
значительные различия по многим частным деталям. Это проявляется, например, в
специфической модификации ряда нуклеотидов в молекуле архебактериальной тРНК,
происходящей после окончания транскрипции. Модификации могут быть уникальными
для архебактерий, общими для эубактерий и эукариот, характерными только для
эубактерий или только для эукариот. В целом тРНК архебактерий отличается от
тРНК эубактерий и эукариот в такой же степени, как последние различаются между
собой.
Несмотря на выявленные
многочисленные различия в аппаратах транскрипции и трансляции между
эубактериями и архебактериями, было показано, что архебактериальная ДНК,
перенесенная с помощью плазмиды в клетку эубактерий, может считываться в ней,
результатом чего является синтез функционально активных ферментов.
Метаболизм
архебактерий. В
группе архебактерий известны организмы с хемоорганогетеротрофным,
хемолитоавтотрофным, хемолитогетеротрофным и фотогетеротрофным типом питания.
Источником углерода для разных представителей могут служить сахара,
аминокислоты, органические кислоты, C1-соединения и CO2.
У архебактерий, метаболизм которых связан с молекулярной серой, автотрофная
фиксация CO2 происходит по восстановительному ЦТК, функционирующему
также в группе зеленых серобактерий (см. рис.
76); виды, способные расти на среде с CO2 + H2,
фиксируют углекислоту по недавно открытому нециклическому ацетил-КоА-пути,
описанному у ацетогенных эубактерий (см. рис.
62). Центральным метаболитом этого пути является ацетил-КоА, который через
фосфоенолпируват превращается в 3-ФГА и далее в молекулу фруктозо-6-фосфата.
Таким образом, у архебактерий, способных к автотрофии, фиксация CO2
происходит по восстановительному пути карбоновых кислот в различных его
модификациях. Ни в одном случае не обнаружено функционирование
восстановительного пентозофосфатного цикла. В сравнении с последним и
восстановительным ЦТК ацетил-КоА-путь фиксации CO2 более экономичен
с точки зрения потребления АТФ, но требует большего количества восстановленных
кофакторов71.
Исследование других
реакций конструктивного метаболизма архебактерий, как правило, обнаруживает
пути, функционирующие у эубактерий. Так, глюконеогенез, начиная с пирувата,
идет по тому же механизму, что у эубактерий. Ассимиляция аммония, синтез
изопреноидных липидов, нуклеотидов происходит по обычным для эубактерий путям.
Катаболизирование
архебактериями Сахаров происходит по путям, свойственным эубактериям: гликолиз,
окислительный пентозофосфатный путь, ЦТК и путь Энтнера–Дудорова. Эти
катаболические пути найдены не у всех представителей группы. У многих
архебактерий, например, отсутствует гликолиз. Таким образом, анаболические и
катаболические пути превращения углеродных соединений у архебактерий сходны с
эубактериальными путями.
71 Для синтеза 1 молекулы триозы в
восстановительном пентозофосфатном цикле требуется 9 молекул АТФ, в
восстановительном ЦТК — 5, а в ацетил-КоА-пути — только 3.
Способы получения
архебактериями энергии включает бесхлорофилльный фотосинтез, брожение, аэробное
и анаэробное дыхание, при котором конечными акцепторами электронов могут быть
CO2 и другие C1-соединения, молекулярная сера, NO3
– , Fe3+ и Mo6+. У организмов, получающих энергию с
использованием электронного транспорта, в качестве электронпереносящих
компонентов обнаружены ферредоксины, хиноны, цитохромы. Электронный транспорт
сопряжен с трансмембранным переносом протонов. Механизм окислительного
фосфорилирования архебактерий соответствует хемиосмотическому принципу и сходен
с аналогичным механизмом эубактерий и митохондрий. В то же время следует
подчеркнуть, что архебактериям свойственны типы энергетического метаболизма, не
встречающиеся у эубактерий и эукариот. Это бесхлорофилльный фотосинтез и особый
тип анаэробного дыхания, в процессе которого происходит образование метана.
Таблица
33. Свойства архебактерий (по Дуде, 1985)
|
Признаки, уникальные для архебактерий
|