Исследование пространственно-демографической структуры населения лесной мыши гетерогенных местообитаний
Министерство образования и науки
Республики Казахстан
Костанайский социально-технический
университет
имени академика Зулхарнай Алдамжар
Факультет «Педагогический»
Кафедра Естественных наук
Специальность 050608 «Экология»
Допускается к защите
зав. кафедрой
Естественных наук
Белан О.Р.
«___»
______________2009г.
ДИПЛОМНАЯ РАБОТА
на тему: Исследование
пространственно-демографической структуры населения лесной мыши гетерогенных
местообитаний
Дипломник:
Дмитриева Маргарита Евгеньевна
Руководитель
дипломной работы: доцент, к.б. н. Белан О.Р.
Костанай, 2009
СОДЕРЖАНИЕ
Введение
Анализ литературных данных по пространственно-демографической
структуре и миграционной активности популяций мелких млекопитающих
Район, материалы и методы исследования
.1 Характеристика районного исследования
.2 Материалы и методы исследования
Динамика численности и демографическая структура населения
лесной мыши
.1 Демографическая структура населения лесной мыши на
площадке 1
.2 Демографическая структура населения лесной мыши на
площадке 2
Динамика миграционной активности населения лесной мыши
Пространственная структура населения лесной мыши
.1 Пространственное распределение лесной мыши на площадке 1
.2 Пространственное распределение лесной мыши на площадке 2
Заключение и выводы
Список использованной литературы
ВВЕДЕНИЕ
Актуальность темы. Закономерное распределение особей в пространстве имеет
важное биологическое значение и, по существу, является основой всех форм
нормального функционирования популяции. Прежде всего, пространственная
структурированность определяет наиболее эффективное использование ресурсов
среды (пищевых, защитных, микроклиматических и др.), снижая хаотичностъ их
использования и, как следствие, уровень конкурентных отношений особей внутри
популяции. На этой основе открывается возможность поддержания высокого уровня
биомассы и биологической активности вида. Это укрепляет позиции данной
популяции в ее взаимоотношениях с другими видами биоценоза и повышает
вероятность ее устойчивого существования [1,2,3,4,5,6]. Второй аспект
биологической роли пространственного структурирования заключается в том, что
она служит основой устойчивого поддержания необходимого уровня
внутрипопуляционных контактов между особями. Осуществление функций популяции -
как видовых (размножение, расселение и др.), так и биоценотических (участие в
круговороте, создание биологической продукции, воздействие на популяции других
видов) - возможно только на основе устойчивых, закономерных взаимодействий,
между отдельными особями и их группами. Поддержание таких взаимодействий в
структурированной системе обеспечено намного надежнее, нежели это возможно при
хаотическом, случайном распределении элементов популяции в пространстве
[7,8,9]. Таким образом, пространственная структурированность популяций представляет
собой «морфологическую» основу популяционного гомеостаза, определяя снижение
уровня конкуренции и поддержание устойчивых внутрипопуляционных контактов как
функциональных, так и информационных.
Определяя собой ведущие стороны биологии популяций, пространственная
структура, в свою очередь, определяется особенностями, местности (пространства)
в биологии данного вида [10,11,12].
Понимание процессов формирования динамики численности и пространственной
структуры мелких млекопитающих связано с изучением миграционных процессов, в
частности, таких проблем, как зависимость миграционной активности от плотности
населения, степени неоднородности биотопических условий, половозрастного
состава популяций.
Таким образом, изучение особенностей пространственной структуры актуально
для понимания основ функционирования популяций в естественных условиях.
Цель исследования. Цель работы состоит в комплексном анализе
пространственно-демографической структуры населения лесной мыши гетерогенных
местообитаний.
Задачи исследования.
.Исследовать сезонные изменения демографической структуры популяции
лесной мыши.
. Проанализировать пространственное распределение и миграционную
активность населения лесной мыши.
.Выявить биотопические особенности пространственно-демографической
структуры и миграционной активности населения лесной мыши.
1. АНАЛИЗ ЛИТЕРАТУРНЫХ ДАННЫХ ПО ПРОСТРАНСТВЕННО-ДЕМОГРАФИЧЕСКОЙ
СТРУКТРУРЕ ПОПУЛЯЦИЙ
Пространственная структура популяции выражается характером размещения
особей и их группировок по отношению к определенным элементам ландшафта и друг
к другу.
Различают следующие принципиальные типы пространственного распределения
особой в популяциях равномерный (регулярный), диффузный (случайный) и
агрегированный (групповой, мозаичный) [13,14].
Равномерный тип распределения в идеале характеризуется равным удалением
каждой особи от всех соседних; величина расстояния мёжду особями соответствует
порогу, за которым начинается взаимное угнетение. Таким образом, теоретически
этот тип распределения в наибольшей степени соответствует задаче полного
использования ресурсов при минимальной степени конкуренции. В действительности
же равномерное распределение особей встречается в природе достаточно редко.
Близкий к этому характер распределения свойствен, например, одновидовым зарослям
некоторых растений, встречается в уплотненных популяциях некоторых сидячих
беспозвоночных. Практическая невозможность реализации этого «идеального» типа
распределения определяется прежде всего неоднородностью среды обитания;
нарушают равномерность распределения и свойственные многим видам формы
взаимосвязей.
Диффузный тип распределения особей, встречается в природе значительно
чаще, при нем особи распределены в пространстве неравномерно, случайно. В этом
случае расстояния между особями неодинаковы, что определяется, с одной стороны,
вероятностными процессами, а с другой - определенной степенью неоднородности
среды. Такой тип распределения широко представлен среди растений и многих
таксонов животных. В частности, диффузное распределение характерно для животных,
у которых социальная связность в пространстве выражена относительно слабо.
Агрегированный (мозаичный) тип распределения выражается в образовании
группировок особой, между которыми остаются достаточно большие незаселенные
территории. Биологически это связано либо с резкой неоднородностью среды, либо
с выраженной социальной структурой, действующей на основе активного сближения
особой. Последнее особенно характерно для высших животных (многие позвоночные,
насекомые, образующие полиморфные колонии, и некоторые другие).
Характер территориального распределения животных видоспецифичен и
определяет возможность наиболее эффективного использования ресурсов среды и
устойчивого осуществления внутрипопуляционных взаимодействий особей и их
группировок. Эти взаимодействия составляют сущность функционирования популяции,
как целого [15].
Во всех случаях тип пространственного распределения в популяциях
определяется, с одной стороны, гетерогенностью среды, а с другой соотношением
процессов пространственной дифференциации, снижающей степень конкурентности, и
функциональной интеграции особей, обеспечивающей целостность популяции как
функциональной системы. Особенно четко это выявляется при изучении популяций
животных.
Для животных как подвижных организмов ведущее значение в определении
характера пространственной структуры популяции имеет степень привязанности к
территории. В крайних вариантах это свойство выражается либо оседлым, либо
номадным (кочевым) образом жизни; между этими вариантами имеется ряд
промежуточных.
Нередко пространственная структура рассматривается как система
индивидуальных участков [16,17,18,19]. Так, В.В. Демидов описывает
пространственную структуру населения мелких млекопитающих как упакованную с
максимальной плотностью мозаику индивидуальных участков, где обитает большая
часть животных, составляющих популяцию [20]. С количеством свободных участков
часто связывают репродуктивную активность животных. В размножении, как правило,
принимают участие особи, обладающие собственной территорией. Зверьки, которые не
имеют индивидуального участка, составляют "резерв" популяции и
приступают к размножению только тогда, когда смогут завладеть гнездовым
участком [17,21,22,23,24,25].
Видам, для которых характерен оседлый образ жизни, как правило, свойствен
интенсивный тип использования территории, при котором отдельные особи или их
группировки (главным образом семейные) в течение длительного времени
эксплуатируют ресурсы на относительно ограниченном пространстве. Для видов
отличающихся номадным образом жизни, характерен экстенсивный тип использования
территории, при котором кормовые ресурсы используются обычно группами особей
(подчас весьма многочисленными), постоянно перемещающимися в пределах обширной
территории.
Различий в особенностях биологии и в нагрузке на биомассу кормовых
объектов в этих двух вариантах существенным образом определяют общий характер
пространственной структуры популяций. Такая структура формируется и
поддерживается в результате сложных взаимоотношений между особями.
Пространственная структура популяций представляет собой не только статическую
картину размещения особей по территории, но и динамическую систему
взаимодействий, выражающуюся в многообразных упорядоченных формах поведения
каждого животного в ответ на информацию о местонахождении и поведении :других
особой. Если топография расположения особей - это «морфологический» аспект
пространственной структуры популяции, то система взаимоотношений
(«этологическая структура») -это функциональный ее аспект. Сочетание этих двух
аспектов составляет‚ биологическую сущность понятия
пространственно-этологической структуры популяций животных. Представление о
пространственно-этологической структуре позволяет объективно определить
механизмы, формирующие динамическую устойчивость и адаптивный характер
структуры популяций разных видов животных.
Для видов с оседлым одиночно-семейным образом жизни принцип
пространственной организации популяций заключается в формировании, системы
индивидуальных (семейных) участков обитания, используемых в течение длительного
времени. Такой тип пространственного распределения ведет к рациональному
использованию ресурсов территории уровне популяций в целом: отдельные особи
распределены в пространстве относительно равномерно; на каждом участке обитания
обеспечены нее условия для жизни. В результате уровень конкуренции за корм,
убежища и другие ресурсы сведен к минимуму, каждая особь, имеет шансы на
выживание и воспроизведение, а популяция в целом получает более широкие
перспективы роста и захвата территории. Кроме изучения процессов формирования и
функционирования скоплений организмов (агрегаций или мерусов), для понимания
пространственной структуры популяции имеет большое значение выявление
особенностей использования территории отдельными животными. В настоящее время
накопилось множество сведений о видовых особенностях, сезонных и географических
изменениях участков обитания. При этом отмечается высокая степень
вариабельности их размеров и конфигураций. Причинами таких изменений могут быть
как факторы внешней среды (сезон года, кормность и защищенность биотопа), так и
внутрипопуляционные. Современными исследованиями выявлены механизмы образования
индивидуальных участков, половые и возрастные особенности размеров и степени
перекрывания участков обитания мелких млекопитающих в разные сезоны [26,27,28,29,30].
Привязанность к ограниченной территории дает особям-резидентам ряд
биологических преимуществ. Большое значение, в частности, имеет знакомство
животных со своей территорией, ее освоенность. В пределах участка обитания
животное перемещается в системе знакомых ориёнтиров. Многие виды прокладывают
на участке систему троп, связывающих места расположения убежищ, кормовых
участков, запас пищи и т.п.; нередко животные метят пути постоянных перемещений
запахом или визуальными метками. В результате повседневная активность животных
осуществляется как бы автоматически: особь резидент кратчайшим путем достигает
мест кормежки, отдыха, укрытия от хищников или непогоды и т. п., не затрачивая
дополнительного времени и энергии на исследовательское поведение. Многочисленные
наблюдения за разными видами животных (главным образом млекопитающих),
постоянно обитающими на данном участке, показывают, что они перемещаются по
участку быстро, направленно, стереотипными путями; при опасности
особи-резиденты быстро скрываются в ближайшем укрытий, В отличие от этого
животные, перемещающиеся во незнакомой территории, передвигаются суетливо,
неупорядоченно, часто принимают позы настороженности, убежища находят лишь
случайно; в их поведении явно доминирует ориентировочная реакция.
Длительное обитание на одном участке у многих видов животных
сопровождается определенными формами благоустройства территории: строительство
разного рода убежишь, формирование системы переходов, кормовых столиков,
запасов корма и т. п. Все это повышает эффективность жизнедеятельности при
одновременном уменьшении энергозатрат и времени на отдельные ее формы, а также
способствует снижению пресса хищников.
Предположение о том, что оседлые особи в меньшем количестве погибают от
хищников, чем особи того же вида, но не имеющие постоянных участков и кочующие
по незнакомой территории ( «внутрипопуляционные мигранты»), высказывалось уже
давно. В настоящее время имеется ряд наблюдений и экспериментов, объективно
подтверждающих это предположение.
Так, в Институте экологии Польской Академии наук на опытном участке леса,
где все оседлые особи рыжих полевок Clethrionomys glareolus были индивидуально помечены, выпустили несколько гадюк Vipera berus. Повторные отловы этих змей позволили изучить состав
полевок, оказавшихся их добычей. Выяснилось, что гадюкам чаще удастся поймать
зверьков, передвигающихся по незнакомой местности, (чем оседло живущих
особей-резидентов. В свою очередь, показало, что успешность охоты гадюк
определяется знакомством с участками, которые у этих змей очень постоянны и
хорошо освоены.
В условиях эксперимента изучалась охота ушастой совы Аsiооtus на хомячков Реromyscus leucopus. При этом часть хомячков
предварительно знакомили с обстановкой экспериментальной камеры, в которой были
размещены укрытия, а часть запускали в помещение без такой подготовки. Опыт
заключался в том, что в камеру, где заранее была помещена голодная сова,
запускали однополые пары хомячков, состоящие из знакомого с территорией
(«оседлого) и незнакомого с ней («мигрант) животных. В результате из 20 особей
каждой категории добычей совы стали 2 «оседлых» зверька и 11 «мигрантов» -
разница статистически достоверна.
Аналогичные опыты, проведенные с пенсильванскими полевками Мicrotus pennsylvanicus, показали, что если на поимку одного
животного, знакомого с территорией, сова затрачивает от 10 до 43 ч то
незнакомый с экспериментальным помещением зверек становится ее добычей в
течение 4-14 ч. Опыты Ю.М. Смирина (1974) также показали, что добычей ушастой
совы скорее становятся только что выпущенные в экспериментальное помещение
грызуны (лесные мыши, рыжие полевки), чем зверьки, заранее освоившие обстановку
и установившие определенные взаимоотношения между собой. Последнее
обстоятельство подчеркивает биологическое значение системы внутрипопуляционных
отношений особей в группах [31,32].
Косвенным образом значение знакомства с территорией для выживания
показывают и некоторые полевые наблюдения.
Формирование участка обитания. Величина и форма участков обитания широко
варьируют у разных видов и даже популяций одного вида. Размеры участков
определяются комплексом факторов; включающих как внешние (особенности среды),
так и внутренние (особенности биологии данного вида). Одно из важнейших
условий, определяющих приуроченность участка к определенному, месту и его
размеры,- обеспеченность кормом. Связь площади участков с обилием кормовых
ресурсов хорошо известна для многих видов животных. Обратные соотношения
площади участков с обилием корма отмечены для ряда видов грызунов, хищных
млекопитающих, птиц и других животных.
Отмеченная закономерность ярко демонстрируется полевыми экспериментами с
внесением в природные биотопы искусственной подкормки. В различных опытах
такого рода регулярно фиксировалось уменьшение размеров участка и подвижности
особей, увеличение плотности населения, а иногда и интенсификация размножения и
рост численности.
Поскольку потребность в пище определяется уровнем обмена веществ, создано
немало математических моделей, демонстрирующих корреляцию величины участка с
размерами тела. На примере млекопитающих было показано, что зависимость уровня
обмена от массы проявляется и в соответствующих сдвигах размеров участка
обитания. Впрочем, подобные обобщения всегда оказываются достаточно
приблизительными: реальный уровень обмена определяется не только размерами
тела; обилие пищи, выраженное в се биомассе, не отражает ее энергетической
ценности, разные виды пищи отличаются по доступности и по затратам энергии на
кормодобывание и т. д. По-видимому существенное значение имеет и величина
энергозатрат на удержание территории: при определенных условиях захват и
удержание большого участка энергетически не оправданы.
Помимо обилия корма, на размеры участка оказывает влияние и ряд других
факторов. Для позвоночных животных, например, вёсьма большое значение имеет количество
и распределение естественных убежищ или мест, природных для сооружения гнезда,
норы и т. п. Поэтому для млекопитающих-норников очень важным оказывается
микрорельеф.
По-видимому столь же важна «структурированность» участка, степень его
неоднородности: чем сложнее местность, тем больше возможность осуществления
всех форм деятельности на относительно меньшем пространстве. Наконец, очень
существенным фактором оказывается свойственный виду тип коммуникации.
Дистанция, в пределах которой может быть воспринята информация от соседей,
устойчивость сигналов, особенности среды, влияющие на распространение
коммуникативных сигналов,- все это имеет прямое отношение как к средним видовым
показателям величины участков, так и к установлению их конкретных границ в тёх
или иных вариантах местности.
В целом расположение и величина участков формируются под воздействием
всего комплекса перечисленных факторов. Можно полагать, что у высших
позвоночных расположение естественных убежищ или мест, пригодных дал их
сооружения, определяет места формирования «центров активности» участков
обитания, обилие и дисперсия кормовых объектов - минимальные размеры участков,
а условия коммуникации - их максимальные размеры. Конкретные структурные
особенности среды корректируют эти параметры.
В процессе освоения занятого участка формируется стереотип поведения
особи в пределах знакомой территории. В дальнейшем этот стереотип выступает в
качестве механизма, удерживающёго. животное в пределах участка, поскольку выход
за его границы немедленно вызывает комплекс ориентировочных реакций,
стимулирующих стремление вернуться в пределы системы знакомых ориентиров.
Наряду с привязанностью к территории, особям-резидентам свойствен
комплекс территориального поведения, направленного на предотвращение вторжения
других особой своего вида: активная защита участка и различные формы маркировки
его границ. Эти механизмы индивидуализации территории входят в число проявлений
популяционного гомеостаза и будут подробнее рассмотрены в следующей главе.
Пространственная дифференциация оседлых животных связана с одиночным
(семейным) образом жизни. Противоположный тип образа жизни - групповой, при
котором особи постоянно или периодически образуют плотные скопления. При
большом скопления особей на малом пространстве возрастает конкуренция между
ними. Поэтому отчетливый групповой образ жизни в наибольшей степени развит у
кочующих (номадных) животных, подвижный образ жизни, которых снижает нагрузку
на кормовые ресурсы, а соответственно и уровень пищевой конкуренции.
Существование в составе группы имеет свои выгоды. В группе животные легче
обеспечивают себя кормом и затрачивают меньше энергии на добывание пищи.
Добывание корма в составе скоплений болёе результативно и требует меньших
энергозатрат - опять-таки отчасти потому, что в группе особь меньше времени
тратит на ориентировочные реакций.
Велико значение группового образа жизни в защите от хищников Скопление
большого числа особей существенно повышает вероятность, раннего обнаружения
опасности; свойственная групповым животным система взаимного оповещения делает
этот факт достоянием всей группы. Эффективность реакции группы на хищника
прослежена в ряде наблюдений и экспериментов. Оповещение об опасности может
быть активным. Специфичность сигналов тревоги подчас обнаруживает их особое
биологическое значение.
Характерно, что при высокой видоспецифичности других звуковых сигналов
крики тревоги воспринимаются, однозначно многими видами. Обнаруживаются и
пассивные формы оповещения об опасности. Аналогичного типа реакции описаны для
грызунов. Обнаружен «запах страха» в у домовых мышей: если обычно в группах
этих грызунов запах отдельных особей имеет привлекающие свойства, то запах
мышей, предварительно напуганных (обдуванием или встряхиванием), вызывал
отчетливую защитную реакцию.
Своевременное обнаружение опасности и сигнализация о ней обеспечивает
повышение эффективности избежания нападения хищников (убегание,
рассредоточение, затаивание и пр.). При этом и возможности непосредственной
обороны, несомненно, выше в составе группы: в ряде случаев группа вполне
успешно обороняется против хищника, от которого одиночная особь защититься не в
состоянии. Оборона столь аффективна, что хищники избегают встреч с массовыми
скоплениями.
Жизнь в группе сопряжена и с некоторыми другими прёимуществами: экономный
тип энергозатрат (метаболический «эффект группы»), возможность передачи опыта
молодым через подражание и прямое обучение и др. Вместе с тем, как уже
говорилось, в многочисленных скоплениях возрастает вероятность конкуренции.
Поэтому биологические преимущества группового образа жизни должны быть
поддёржаны упорядоченными пространственными взаимоотношёниями особей и
специальными формами интегративного поведения.
У животных, ведущих групповой образ жизни, и пространственная
дифференциация имеет групповой характер и выражается в виде закономерного
распределения отдельных внутрипопуляционных структур -колоний, стай, стад. У
номадных животных степень привязанность к территории отдельных кочующих групп
может быть различной в зависимости от экологических особенностей вида и типа
структурированности внутрипопуляционных отношений. В принципе можно отметить
тенденцию уменьшения величины групповой территории и усиления эффективности ее
маркировки и защиты по мере усложнения внутристадной организация. У видов с наиболее
сложной организацией, позволяющей координировать действия всех особей,
формируется общий групповой участок обитания. Все перемещения укладываются в
пределы, этого участка, который животные активно маркируют и даже защищают. В
этом случае, как и у оседлых животных, складывается система постоянных путей
передвижения, фиксированные места ночевок, водопоев, кормления и т. п.
Внутренняя структурированность подвижной группировки подчиняется задачам
четкой регуляции взаимного расположения в пространстве и высокой степени
синхронизации действий составляющих группу особей. В этологии хорошо
разработано представление о дистанциях, активно поддерживаемых особями в
составе группы. Говорят об индивидуальной дистанции, которую соблюдают особи по
отношению друг к другу. У млекопитающих индивидуальная дистанция всегда больше
спереди, в поле зрения, чем сбоку или сзади. Индивидуальная дистанция отражает
тенденцию к пространственном отделению от других особей. Социальная дистанция,
напротив, подчеркивает связность животных в группе: это максимальное
расстояние, на которое животное может отойти от группы. Взаимодействие этих
двух тенденций приближения и отдаления - формирует «жизненное пространство»
особей, в пределах которого нет других животных; и каждая особь обладаёт
определенной степенью свободы. Находясь внутри этого пространства, особь
взаимодействует с другими животными; расстояние, в пределах которого
осуществляются внутригрупповые связи, называют стадной дистанцией.
Определенная структура («строй») группы поддерживается системой
специальных адаптаций, основывающихся на свойственных животным стереотипах
поведения, направленного на постоянную ориентацию на соседних особей. Этот
процесс облегчается такими морфологическими приспособлениями, как контрастные
элементы окраски, различные формы локации, ольфакторное восприятие соседей и
др. У млекопитающих большую роль в поддержании строя играет активная звуковая
сигнализация: реагируя на звуки соседей, отдельные животные направленно
поддерживают свое положение в стае. Этим обеспечивается сохранение целостности
группы и поддержание устойчивых информационных связей, лежащих в основе
функциональной структуры групп.
Рассмотренные формы пространственной дифференциации представляют собой
механизмы увеличения дисперсности в распределении популяции по территории. Их
основное адаптивное значение заключается в снижении конкуренции за ресурсы. Но,
как уже говорилось, «центробежный» процесс пространственной дифференциации не
может быть бесконечным - этому противостоит задача интеграции особей в
пространстве, без чего не могут быть реализованы важнейшие общепопуляционные
функции: воспроизводство, регуляция плотности населения, общепопуляционная
реакция на внешние условия, взаимоотношения с популяциями других видов. Поэтому
в общей системе популяционных адаптаций механизмы, определяющиё устойчивое
поддержание контактов между особями, их интеграцию в единую функциональную
систему имеют важнейшее биологическое значение. Эти механизмы действуют
«центростремительно» у территориальных животных как бы уравновешивая действие
пространственного разобщении в создании суммарной пространственно-этологической
структуры, в которой достаточно низкий уровень конкуренции сочетается с
достаточно высокой контактностью, обеспечивающей бесперебойное функционирование
популяции. У стадных животных поддержание устойчивых информативных контактов не
только для сохранении адаптивного построения стада (стаи), но и лежит в основе
поддержания целостности стада как функциональной популяционной системы.
В основе механизмов интеграции лежат два взаимосвязанных процесса:
непрерывная информация о месте локализации отдельных особей и их группировок и
наследственно детерминированный стереотип поведения, стимулирующего животное к
поиску и поддержанию контактов себе подобными.
Информация о присутствии особей своего вида обширна и многообразна. В нее
входят как «личные сигналы», воспринимаемые визуально, на слух и по запаху, так
и различные «попутные» формы информации: следы, тропы, поеди, норы, гнезда,
фекалии и т. п. Немалое значение имеют все формы обозначения занятой
территории, попутно с основной функцией несущие информацию.
Давно известно, что в годы депрессии численности грызунов зверьки
распрёделены группами, которые могут располагаться далеко одна от другой,
сохраняя внутри себя характерные для вида формы расположения участков и
взаимодействия особей. Образованию таких групп предшествует период повышенной
подвижности, который можно характеризовать как «поиск контактов», Так, в
Московской области при резком (десятикратном) снижении численности обыкновенных
полевок Microtus arvalis подвижность зверьков возросла в три раза. Более
высокая подвижность при низкой, плотности была зарегистрирована и у
американских «белоногих мышей» Peromtscus leucopus.
Наблюдения в естественных популяциях полевых мышей в Польше показали, что
количество расселяющихся зверьков, оседающих в данном месте, прямо
пропорционально числу оседло живущих здесь особей. Эти наблюдения показывают
роль интегративного поведения в формировании естественных внутрипопуляционных
группировок.
Реальные пути формирования таких групп прослежены, например, при работе
по борьбе с грызунами в природных очагах инфекций. Так, монгольские песчанки,
выжившие после обработки ратицидами больших территорий, оставили свои участки и
перешли к ненаправленным перемещениям, в процессе которых встречались с другими
животными и уже группами вновь переходили к оседлому образу жизни. Характерно,
что в период кочевок между песчанками явно преобладали мирные, контактные формы
поведения. «Тяга к контактам» показана и в эксперйментах с грызунами, хотя в
целом этот вопрос изучен еще явно недостаточно. Не исключено, что это свойство
неодинаково выражены у представителей разного пола и возраста. В опытах с
древесной ящерицей в естественных популяциях удаление самок вызывало резкое
повышение подвижности самцов: лишь 5 % их остались на своих участках; в тех же
условиях удаление самцов не вызывало увеличения подвижности самок.
В сложившихся популяциях активный поиск информации и контактов составляет
одну из характерных сторон исследовательского поведения: «патрулирование»
участков, а также спонтанные выходы за их пределы с последующим возвращением
характерны для хорошо изученных в этом отношении грызунов, а также других
млекопитающих, птиц в репродуктивный период некоторых других животных.
В результате стремления к установлению контактов с особями своего вида у
оседлых животных с интенсивным типом использования территории полное
пространственное разобщение участков обитания, встречается не так часто. У большинства
млекопитающих и у многих других животных участки соседних особей в той или иной
степени пёрекрываются. При этом активная охрана участка проявляется лишь в
отношении территории, на которую не налегают участки соседей, зоны же
совместного использования маркируются всеми животными-соседями, а активно
защищаются только от «чужаков». Области перекрывания участков представляют
собой своеобразные зоны контактов», в которых между соседними животными:
устанавливаются нейтральные или даже дружелюбные отворения на основе
индивидуального опознавания. так; наблюдения за малыми сусликами показали, что
на общей часта территории зверька-соседи мирно кормятся рядом, при опасности
прячутся в соседние защитные норы, совместно прогоняют с этой территории
«чужаков».
Аналогичные наблюдения имеются и по другим видам. Показано, например, что
пойманные в природе полевки-экономка при парном объединении в экспериментальной
камере демонстрировали преобладание агрессивных контактов, если это были
животные, пойманные в разных местообитаниях, и дружелюбных - если это
оказывались зверьки е соседних участков. Такие же результаты ранее были
получены в опытах хлопковыми крысами.
Дифференцированное проявление территориальности с выделением в пределах
участка активно охраняемой части открывает возможность «маневра» в
использовании пространства: у грызунов, например, при увеличении плотности
населения отмечается уменьшение размеров активно охраняемой территории,
возрастание степени перекрывания участков и уменьшения их площади. Такая реакций
адаптивна на уровне популяции как целого: это предохраняет популяцию от потерь
на время, пока включаются и реализуются мёханизмы приведения плотности
населения в соответствии о конкретной экологической обстановкой.
Усилению контактности способствует и различное отношение
особей-резидентов к представителям того же вида, проникающим на территорию их
участка, активная охран территорию не абсолютно. Известно, что рецептивные
самки бурундуков и бурундуковых бёлок перестают защищать свои участки от
самцов, которых до начала течки на свою территорию не допускают. В большинстве
же случаев во все периоды года агрессия по отношению к особям противоположного
пола отсутствует или проявляется вяло. У многих видов грызунов участки самок,
как и самцов, практически изолированы друг о друга тогда как участки особей
разного пола могут широко перекрываться.. Исключение представляют виды, у
выражена только у самою в этом случае в период выкармливания агрессия самок по
отношению к самцам.
У грызунов и некоторых других позвоночных в ряде случаев регистрируется
терпимое отношение к молодым животным. Показано, что у грызунов это
определяется спецификой их запаха. Этим объясняется слабая выраженность стресса
у молодых расселяющихся животных в отличие от взрослых внутрипопуляционных
мигрантов неизбежную агрессию со стороны резидентов. Таким путем обеспечивается
важная общепопуляционная функция дисперсии молодых особей, способствующей
увеличению занятого популяцией пространства.
На основе рассмотренных механизмов интеграции общая пространственная
структура популяции реализуется в виде системы отдельных внутрипопуляционных
группировок, в каждой из которых особи находятся тесном информационном и
функциональном контакте, поддерживая устойчивые взаимоотношения. У высших
животных эти отношения строятся на личном опознавании членов группы, и
пространственно-этологическая структура служит основой стабильности и
«управляемости» популяции как биологической системы.
В настоящее время наиболее широко обсуждаются вопросы пространственного
распределения по территории не отдельных особей, а их группировок разного
уровня (мерусы, парцеллы, агрегации). Так, В.Е. Флинт идентифицирует
пространственную структуру как типичную и закономерную для данной популяции
пространственно - временную динамику мерусов, слагающих ее [7]. Здесь мерусы -
совокупность особей, обладающих фенотипическим единством и представляющих собой
таксон биохорологической структуры вида. Фенотипическое единство животных
основано на тонкой адаптированности к конкретным и неустойчивым условиям
ограниченного участка земной поверхности на коротком отрезке времени. И.А.
Шилов [9] описывает отдельные группировки животных внутри популяции, в которых
особи находятся в устойчивых информационных и функциональных контактах. Такие
группы в литературе называются демами или парцеллами [4,5,33].
Исходя из этого, пространственную структуру населения можно представить
как закономерное чередование пятен, характеризующихся определенным обилием
животных. Каждое такое пятно это площадь территории, занятая частью популяции,
причем независимо от уровня численности все такие части имеют один и тот же
иерархический внутрипопуляционный ранг (Смуров, 1989). Степень агрегированности
можно оценить путем статистического анализа распределения животных по
территории [34,35,36]. В зависимости от расположения этих пятен выделяют
различные типы пространственного распределения животных. Так В.Е. Флинт [7]
описал четыре типа пространственной структуры популяций: эквальный (диффузный),
инсулярный (мозаичный), циклический (переложный), пульсирующий.
Многочисленные наблюдения за различными видами житных из разных таксонов
показывают, что даже в относительно однородных условиях местообитания
равномерное или диффузное распределение особей встречается практически как
исключение; у самых разных видов явно преобладает агрегированный тип
распределения, при, котором в составе популяции формируются группировки особей;
более тесно связанных друг с другом, чем с другими аналогичными группировками.
Их называют демами, субпопуляциями, парцеллярными группировками, мерусами,
внутрипопуляционными группировками. Не даваясь в тонкие отличия этих понятий;
констатируем лишь, что внутрипопуляционные группировки представляют собой
совокупность особей и семей, живущих в ближайшем соседстве и непосредственно
связанных друг с другом теми или иными формами сигнализации. Они возникают
активно, и их биологическая роль заключается в различных типах взаимопомощи.
Дальнейшие исследования показали, что роль внутрипопуляционных
группировок еще более значительна: они представляют собой элементарную
структурную единицу популяции, и именно на их основе обеспечивается адаптивный
ответ популяции в целом на изменения вне- и внутрипопуляционных условий.
Несмотря на индивидуальную неустойчивость самих таких группировок (они могут у
ряда видов распадаться формироваться вновь; состав индивидов в каждой такой
группировке может все время меняться), именно их существование обеспечивает
устойчивость популяции в целом.
Интеграция особей в составе различного рода внутрипопуляционных группировок
всегда сопровождается определенной степенью их пространственного сближения
(перекрывание участков, тяга к контактам). Это ведет к некоторому повышению
вероятности обострения внутрипопуляционной конкуренции. Ослабление негативных
отношений, повышение рациональности использования ресурсов среды идет в этом
случае через упорядочение взаимных отношений особей в группах. У высших
животных это связано с проявлением специфических форм социального поведения,
комплекс которого рассматривается как этологическая структура
внутрипопуляционных групп.
В общей форме характер взаимоотношений особей во внутрипопуляционных
группировках можно определить как более или менее выраженную систему типа
«доминирование -подчинение», при котором отдельные особи отличаются в различных
видах деятельности. Конкретное выражение форм доминирования - подчинения широко
варьирует у разных видов, но всегда основывается на неравнозначности особей,
входящих в состав группировки. Такую систему взаимоотношений принято называть
иерархической. При такой системе в составе группы выделяются особи разных
рангов: «доминанты», «субдоминанты» (подчиняются доминантам, но господствуют
над другими) и различные ступени «подчиненных» особей (субординанты).
Такал система иерархии имеет место у многих видов. В частности, она
изучена у лабораторных белых мышей. В природных условия: линейный тип иерархии,
по-видимому, свойствен серой белке; у пустынных кавий в Аргентине участки
самцов широко перекрываются, и на совместно обитаемой территории среди них устанавливается
линейный тип доминирования подчинения.
Однако в большем числе случаев линейная иерархия замещается групповой,
при которой если и выделяется единственный доминант (по крайней мере, в
экспериментальных условиях), то остальные особи не составляют линейного ряда
соподчинения, а распадаются на две-три группы, в пределах которых животные
более или менее равнозначны. Так, в частности, складывается структура отношений
у многих видов полевок и других мелких грызунов.
Есть и иные варианты внутрипопуляционной иерархии. Так, у нильских
крокодилов популяция, по-видимому, распадается лишь на две категории особей:
доминантные самцы захватывают участки на отмели и защищают их, все остальные
особей участков не имеют. У отдельных видов наблюдается «деспотическое» доминирование
одной особи при относительно равнозначном подчинении остальных. Такая структура
отношений характерна, например, для хомячков, по крайней мере, в условиях
искусственного ограничения территории; близкая к ней - у черных крыс. Особенно
отчетливо, хотя и в своеобразной форме, такой тип взаимоотношений проявляется у
диких домовых мышей, заселяющих замкнутые пространства построек человека.
Отношения доминирования-подчинения чаще наблюдаются только среди самцов,
хотя у ряда видов параллельно формируются и среди самок, а у некоторых животных
возникает общая система иерархии. Главное биологическое значение
структурированности взаимоотношений заключается в том, что при этом повышается
степень организованности и «управляемости» группы, что важно при нарастающей по
мере совмещения участков вероятности конкуренции. Видимо, поэтому выраженность
структуры взаимоотношений тем более отчетлива, чем в большей степени
объединяются территории отдельных особей. В частности, деспотическое
доминирование одного самца, в группах домовых мышей напрямую связано с
обитанием в замкнутых ограниченных пространствах; при поселении в природных
биотопах этот вид формирует структуру в виде участков отдельных особей с
мягкими формами доминирования-подчинения группах.
В целом связь систем взаимоотношений в территориальных группах с
характером использования пространства можно представить следующей схемой:
а) система неперекрывающихся участков обитания. Отчетливо выраженная
охрана и маркировка территории; однородность особей по эколого-физиологическим
свойствам; отсутствие доминантно - подчиненных отношений между ними;
б) группа перекрывающихся участков обитания. Разделение участков на
охраняемую частъ и «зоны контактов»; отношения типа доминирования-подчинения,
иерархия;
в) общая групповая территория. Жесткая иерархия с деспотическим
доминированием (домовая мышь в постройках).
Иерархическая соподчиненность особей в группах определяет различия в их
поведении. Особи-доминанты обладают явными преимуществами и свободно, проявляют
различные формы деятельности. Более крупные самцы, имеющие высокий
иерархический статус, чаще занимают территории, имеющие, укрытия. В
экспериментальных группах грызунов доминанты неограниченно передвигаются по
всей территории, беспрепятственно подходят к корму, используют любые убежища и
т.д. Подчиненные животные, напротив, ограничены в своих действиях поведением
более высокоранговых: их перемещения лимитируются присутствием доминанта, то же
относится к использованию укрытий и мест кормления. Чаще всего низкоранговые особи
избегают контактов с доминантам, но иногда последний активно сдерживает
передвижения подчиненных особей, отгоняет их от кормушек и т.п. При встрече с
особями более, высокого ранга субординанты демонстрируют особые формы
«поведения подчинения», сдерживающие агрессию со стороны доминатов. Вообще,
ритуальные формы поведения играют большую роль в снятии ‚агрессии и
нормализации деятельности группы в целом.
Нередко в результате таких взаимоотношений у наиболее низкоранговых
животных смещается тип суточной активности: в опытных группах такая особи
проявляют активность, в том числе кормовую, в периоды, когда неактивны
доминантные животные. Нечто подобное можно отметить и в природе: наблюдения за
мечеными грызунами показывают, что, в частности, активность молодых самцов,
обычно занимающих низшие ранги, не совпадает активностью взрослых
высокоранговых особей либо во времени, либо в пространстве. В некоторых опытах
у подчиненных животных отмечалась повышенная исследовательская активность, что
можно рассматривать как стимул к миграция. Это подтверждается и наблюдениями в
природе: низкоранговые особи, покинувшие свои участки и вошедшие в состав
«внутрипопуляционных мигрантов»,- обычное явление.
В поведении доминантных особей отмечаются специфические элементы, которые
можно рассматривать как проявления некоторых общегрупповых функций. Отмечено,
например, что доминирующие самцы домовых мышей активно маркируют и патрулируют
территорию группы; то же наблюдается у монгольских песчанок и ряда других видов
грызунов и зайцеобразных. У сумчатых летяг самцы-доминанты метят только
территорию, но и подчиненных особей; таким образом индивидуальный запах
доминанта приобретает, роль фактора групповой интеграции. Подобным же образом
поддерживается целостность «кланов (групп, заселяющих единую систему нор у
барсуков с той разницей, что в этом случае мечение в группе взаимное, без четко
выраженной иерархии.
У некоторых видов существенна роль доминатов во взаимоотношениях с
«чужими» особями. Так, у домовых мышей именно доминирующие самцы активно
охраняют групповую территорию вторжения животных извне. Остальные особи, если и
вовлекаются в борьбу, то обычно в меньшей степени; при этом активность
нападений на «чужаков» коррелирует с положением данной особи в структуре
группировки. В семейных группах монгольских песчанок функцию охраны групповой
территории выполняют доминантные самки. Аналогичное «разделение обязанностей» в
территориальных группах описано и у некоторых других видов.
Как показывают многочисленные наблюдения за искусственными популяциями
лабораторных и диких домовых мешей, а также других видов млекопитающих,
доминирующие самцы имеют определенное преимущество в половых контактах - как
правило, тем большее, чем более, четко выражена иерархическая структура. Таким
образом, значение внутрипопуляционных группировок как элементарных
размножающихся единиц («демон») связано в первую очередь с высокоранговыми
самцами.
Иерархическая соподчиненность животных в группах накладывает, отпечаток и
на их физиологические особенности. В опытах с белыми мышами выяснилось, что у
подчиненных особей уровень обмена веществ был ниже, чем у доминантов; при
удалении доминантов из групп уровень потребления кислорода у животных, занявших
их место, повышался; при рассаживании поодиночке в течение недели повышался
уровень обмена и у тех особей, которые не сумели занять высокоранговое
положение. Это показывает, что снижение уровня обмена вызвано угнетающим
влиянием высокоранговых особей [37].
Аналогичные данные получены в опытах с хомяками («оленьи мыши»): уровень
потребления кислорода был выше у доминантов, особенно ночью, в период
активности. У подчиненных самцов, изъятых из групп, уровень и ритм потребления
кислорода постепенно возвращались к норме.
В сложившихся группах различных видов грызунов и некоторых других
млекопитающих подчиненные особи всегда проявляют определенно выраженную реакцию
стресса, тогда как у доминантов стресс не выражен идя выражён слабо. В
природных условия во внутрипопуляционных группировках изученных в этом
отношении видов грызунов также отмечается дифференциация особей по уровню
стресса, которая в целом соответствует различиям в размерах участков и их
расположении. Поэтому есть все основания считать, что и в природе
внутрипопуляционные группировки строятся по иерархическому принципу на основе
физиологической разнокачественности особей.
При всем различии особей, составляющих внутрипопуляционную группировку,
последняя, несомненно, представляет собой единое целое. Это видно уже из того,
что, невзирая на определенную степень взаимного угнетения, отношения в группе
более стабильны и терпимы, чем отношение тех же животных к вторгающимся в
группу «чужакам». Агрессия по отношению к чужим особям зарегистрирована у
многих видов как в экспериментах, так и в природной обстановке. Показано, в
частности, что в природе грызуны, проникающие в достав оседлых группировок,
характеризуются повышенным уровнем стресса. В приближенных к естественным
условиям опытах с красно-серой полевкой в Японии было выяснено, что
внедряющиеся в сложившиеся группировки новые особи, особенно самцы, занимают в
них низшие ранги.
Целостность сложившейся группировки прослеживается и в более прямой
форме. Так, изучение свободно размножающейся группы кроликов на большом (18 га)
выгоне показало, что при нарастании численности популяции, происходящей от трех
исходных пар, практически все население было сконцентрировано в три группы;
обмен особями между ними был ограничен, расселения кроликов по свободной части
выгона не наблюдалось. По другим наблюдениям свободно живущая популяция домовых
мышей, заселяющая чердачное помещениё, распадалась на две группировки, которые
не смешивались, несмотря на отсутствие препятствий к передвижению и отдельные
переходы зверьков из группы в группу. Подобная же реакция прослежена в
эксперименте: при предоставлении двум сложившимся группам домовых мышей доступа
друг к другу объединения этих групп не произошло, и особи чужой группы
неизменно вызывали агрессивную реакцию. В аналогичных опытах с лабораторными
мышами объединение групп состоялось после схватки между доминантами; в
образовавшейся таким путем группе все мыши из группировки проигравшего схватку
доминанта заняли подчиненное положение.
Таким образом, в результате тесных взаимодействий в территориальных
группировках позвоночных животных формируется определенная разнокачественность
особей по физиологическим свойствам, поведению и «вкладу» в общепопуляционные
функции. Такая функциональная структурированность лежит в основе
авторегуляторных реакций на уровне популяции как интегрированной целостной
системы.
Изучение механизмов формирования и функционирования пространственной
структуры необходимо, прежде всего, потому, что именно пространственная
дифференциация населения может являться первым этапом в становлении популяции
как единицы эволюционного процесса. Она служит основой, на которой возникает
дифференциация населения по биологическим свойствам и определяет генетическую
неоднородность популяции [2,3,4]
2. РАЙОН,
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
2.1 Характеристика района исследования
Ауликольский
район расположен на севере тургайской впадины в южной части соленого озера
Кушмурун. Территория района характеризуется континентальным климатом,
проявляющемся в резких сменах температур дня и ночи, зимы и лета, а также в
быстрых переходах от зимы к лету.
Зима продолжительная,
малоснежная и холодная, с сильными периодическими ветрами, метелями. Ввиду
маломощности снежного покрова (около 250 мм) почва сильно промерзает (более
1,5м).
Весна отличается быстрым ростом тепла и высокой контрастностью дневных и
ночных температур, а также сильными ветрами, вызывающими быстрое иссушение
поверхности почвы.
Лето сухое и жаркое. Как и весной летом довольно часты суховеи и засухи.
Жаркий период со среднесуточной температурой выше +200С длится не более месяца.
Осень характеризуется постепенным похолоданием, первые заморозки
наступают в конце сентября.
Среднегодовое количество осадков для Ауликольского района - 284мм, что
указывает на дефицит влаги. Поэтому, для выращивания сельскохозяйственных
культур в регионе, необходимо проводить агротехнические мероприятия,
направленные на накопление и сохранение влаги.
Ауликольский район расположен вдоль Тургайской ложбины в центре
Тургайского прогиба. На севере и западе района расположена Северо-Тургайская
равнина, рельеф которой плоскоравнинный или слабоволнистый.
На юге ауликольского района расположено Терсекско-Адаевское плато,
которое имеет плоскую поверхность и ограничено с севера высоким уступом,
изрезанными балками. С востока к району примыкает Восточно-Тургайское плато,
которое представлено равниной, слабо расчлененной руслами мелких пересыхающих
речек, а также террасовидные уступы вдаются в Тургайскую ложбину и сильно
изрезаны глубокими балками. В районе поселка Аманкарагай в 5 км от центра
района расположен Аманкарагайский бор с огромным песчаным массивом. В
приборовой полосе на границе леса со степью расположены высокобугристые пески.
В целом по району рельеф равнинных массивов способствует применению
сельскохозяйственной техники. Почвенный покров Ауликольского района
разнообразен в преобладающем большинстве это темно-каштановые почвы с небольшим
процентом других почв. Чаще всего это примеси солончаков, так как реки и озера
здесь в основном соленые. В районе Аманкарагайского бора почвы
дерново-подзолистые по механическому составу песчаные. Примерно половина всех
земель составляют почвы легкосуглинистые. Очень небольшой процент занимают
черноземы южные карбонатные и южные карбонатные луговые.
На вышеперечисленных почвах пахотные угодья не целесообразно располагать.
Поэтому в Ауликольском районе занимаются скотоводством и частично выращиванием
бахчевых культур в северо-восточной части района.
Так как район расположен в зоне засушливой степи, а почвенный покров в
основном не пригоден для выращивания зерновых культур, то большая часть
территории занята пастбищными угодьями. Распаханы не большие участки вокруг
поселков, озер, рек. Наиболее распространенными растительными комплексами
являются разнотравно-песчаноковыльно-красноковыльные степи и
разнотравно-песчаноковыльные степи - темнокаштановых почвах, а на каштановых
почвах и солонцах комплексами являются типчаково-ковыльные степи с
типчаково-полынными сообществами. Основу травостоя образуют: ковыль волосатик,
типчак, тонконог, полынь австрийская. Использовать данные пастбища
рекомендуется вечной, летом и осенью для выпаса всех видов скота.
Животный мир представлен несколькими классами из них:
§ пресмыкающие - ящерица живородящая;
§ земноводные - жаба обыкновенная;
§ птицы - жаворонок полевой, перепел, трясогузка белая, каменка
обыкновенная, воробей полевой, ласточка деревенская, а также хищники - коршун
черный, кобчик обыкновенный, лунь луговой, некоторые виды ястребиных;
§ млекопитающие - полевка серая, полевая мышь, некоторые виды сусликов,
заяц беляк и т. д.
из класса птиц многие хищные и кукушкообразные отряды, отряд сов и широко
распространены птицы умеренных широт: грачи, воробьи, сороки, жаворонки,
встречается иволга, коршун. Из мелких млекопитающих - лесная мышь, полевка
обыкновенная, полевая мышь, бурозубка обыкновенная.
Гордостью района является Аманкарагайский бор, состоящий из соснового и
березово-соснового леса на песках и слабосформированных дерново-подзолистых
почвах.
Засушливый климат и равнинный рельеф Ауликольского района определяет
слабое развитие речной сети. Основная река Убаган имеет снеговое питание. Для
нее характерны резкие колебания среднегодовых расходов воды в различные по
водности годы. До 80% - 90% годового стока приходится на весну (апрель - май).
В долине реки много озер долино-руслового происхождения. Площадь водной поверхности
самых крупных озер - Кушмурун и Сары-Моин - составляет 465 и 126 км2
соответственно. Река Убаган и приток Ащибуй, а также озеро Кушмурун, соленые.
Большинство рек и озер летом пересыхают, и образуется множество плесовых
блюдец, в которых вода держится весь пастбищный период. Уровень воды
поддерживается родниками, атмосферными осадками и грунтовыми водами, которые
находятся на глубине 3 - 3,5 м.
Исследуемые нами участки расположены в степном и лесном биотопах в
непосредственной близости друг от друга. Степь находится в 2-3 км к югу от
озера Кушмурун. Растительность степи представлена следующими видами:
§ семейство злаковые - ковыль, пахучий колосок, тимофеевка луговая;
§ семейство мотыльковые - клевер луговой, чина луговая, люцерна
серповидная;
§ семейство зонтичные - синеголовник плоский;
§ семейство сложноцветные - пижма обыкновенная, тысячелистник мелкоцветный,
полынь горькая, полынь бело-земельная;
§ семейство норичниковые - коровяк фиолетовый.
Огромное влияние на данный биотоп оказывает человек своей
сельскохозяйственной и промышленной деятельностью.
В недалеком прошлом лет 8-10 назад степь подвергалась распашке, что
повлекло за собой изменение видового состава растительности, а находящийся в 5
км от степи карьер пагубно влияет как на животный, так и растительный мир. Если
после прекращения распашки земель растительность начала восстанавливаться, то
резкое увеличение пастбищной нагрузки нарушило экологический баланс.
Растительность ксерофитизируется, становится редкой и низкорослой.
Второй участок расположен в ольховом лесу. Древесный ярус представлен
ольхой, осиной и березой. Средний - это ива, черемуха, дикая вишня, шиповник,
ирга. Нижний - лопух, лапчатка, крапива, папоротник. А завершает все ярус мхов
и лишайников.
На почве всегда есть остатки растений, опавших листьев, сухие ветки,
образующие лесную подстилку, богато населенную микроорганизмами и грибами,
вызывающими минерализацию опада. Поэтому в ольшанике почва плодородная, рыхлая,
влажная. Наличие обильной стабильной кормовой базы делает фауну богатой и
разнообразной.
2.2 Материалы и методы исследования
Исследования проводились в мае-сентябре 2007 г. в степном и лесном
биотопах. Основной метод сбора материала отлов лесной мыши стандартными линиями
давилок (ловушек Геро) [38]. За указанный период отработано 600 ловушко-суток и
отловлено 215 животных. Пойманных зверьков взвешивали, определяли пол,
репродуктивное состояние, возраст согласно методики Н.В. Тупиковой [39].
Нами выделялись три возрастные группы: прибылые неполовозрелые - juvenis;
прибылые половозрелые - subadultus; половозрелые животные прошлого года
рождения - adultus. Отловы проводились сериями по 4 дня, серии отловов названы
турами (табл. 1).
Таблица 1
|
Тур
|
Дата
|
|
1
|
11.05.-14.05.
|
|
2
|
02.06.-05.06.
|
|
3
|
15.07.-18.07.
|
|
4
|
10.08.-13.08.
|
|
5
|
02.09.-05.09.
|
Для анализа миграционной активности животных в работе использована
методика, предложенная О. А. Лукьяновым [40,41,42] позволяющая на основе
последовательных суточных отловов животных количественно оценить интенсивность
потока перемещающихся через площадку особей.
Для характеристики обилия и пространственной структуры населения животных
использовали следующие показатели.
. Общее обилие (I),
выражаемое в числе отловленных особей на 100 л/с, рассчитывали обычным способом
по формуле: I = (c/d)*100, где c - число отловленных особей за 4
суток, d - число отработанных ловушко-суток (d=a*t, где a - общее число ловушек, t - число суток отлова, в данном
случае t=4). Этот показатель отражает
совокупное обилие животных на территории, включающей все типы микроучастков,
часть из которых непригодна для обитания вида и не заселена им. Общее обилие
выражает в некотором смысле емкость и пригодность среды обитания в целом для
населения вида.
. Частное обилие (А) - обилие вида на микроучастках, заселенных
животными, выражено числом отловленных особей на 100 л/с, рассчитано по
формуле: А = (с/f)*100, где с -
количество отловленных животных за 4 суток, f - число отобранных ловушко-суток в микроучастках (f = b*t, где b - частное число ловушек из общего
количества d, отлавливавших животных, отражает
число микроучастков территории, занятых зверьками, t - количество суток отлова). Этот показатель отражает обилие
животных на подходящих микроучастках территории, которые они реально заселяют.
. Заселенность территории (F)
отражает долю (%) от обследованной территории, заселенную животными.
Заселенность рассчитывается по формуле: F=b/a*100, где а - общее количество
ловушек, b - частное число ловушек,
отлавливавших животных. Этот показатель отражает совокупную емкость всей
территории для особей данного вида.
. В качестве меры агрегированности (Ag), или степени скученности населения мы используем индекс
Уитфорда (Whitford, 1949), который в данном случае
равен отношению частного обилия (А) к заселенности территории (F): Ag=A/F.
Этот показатель принимает наименьшее значение при выраженном размещении
животных по территории и возрастает с увеличением мозаичности (когда небольшое
число ловушек попадает на микроучастки, плотно заселенные животными, а
оставшиеся - на территорию, не заселенную зверьками).
Для оценки доверительных интервалов, в пределах которых лежат
действительные значения параметров, мы вычисляли стандартные ошибки показателей
обилия и пространственной структуры.
Статистическая обработка проведена посредством прикладного
статистического пакета "Statistica". Достоверность различия средних
оценивалась с помощью t-критерия. Различия считались достоверными на уровне
значимости 5% (р=0.05).
3. ДИНАМИКА ЧИСЛЕННОСТИ И ДЕМОГРАФИЧЕСКАЯ СТРУКТУРА НАСЕЛЕНИЯ ЛЕСНОЙ МЫШИ
3.1 Демографическая структура населения лесной
мыши на площадке 1
С весны 2006 г. обилие лесной мыши увеличивалось до сентября (табл. 2),
причем рост численности обусловлен, главным образом, иммиграцией молодых неполовозрелых
животных (их средняя доля за летний период составляла 79,4%). Характеризуя
интенсивность размножения нужно отметить, что общая доля половозрелых
размножающихся самок в среднем за сезон невысока, она составляла 4,1%. Кроме
того, в популяции лесной мыши практически на протяжении всего сезона
размножения преобладают неполовозрелые животные (табл. 2). Средняя доля
неполовозрелых за сезон размножения - 79,4%. Высокая доля молодых
неполовозрелых зверьков в составе населения лесной мыши, при низком репродуктивном
потенциале может объясняться только миграциями этих животных из переуплотненной
окружающей площадку лесной полосы.
Анализируя сезонные изменения соотношения полов отмечается относительно
высокая доля самок в начала репродуктивного сезона 66,7%, а затем снижается к
осени до 43,2%. Половозрастная структура и динамика численности лесной мыши
скорее свидетельствуют о том, что животные используют первую площадку для
временного расселения и проживания молодняка.
3.2 Демографическая структура населения лесной мыши на площадке 2
Общий уровень численности лесной мыши значительно выше, чем на первой
площадке (табл. 3). Обилие с мая до середины августа увеличивалось, достигнув
максимального значения (39); основную долю прироста составляли неполовозрелые животные.
К началу сентября численность снизилась в два раза, что, по-видимому, является
следствием торможения репродуктивных процессов и миграциями животных,
вызванными перенаселением площадки. Косвенно это подтверждается продолжающимся
увеличением численности лесной мыши в соседнем биотопе.
Таблица 2
Демографическая
структура лесной мыши на площадке 1
|
тур
|
Общее обилие, экз.
|
Доля неполо- возрелых,%
|
Доля поло-возрелых,%
|
Доля полово- зрелых
пере-зимовавших,%
|
Доля самцов,%
|
Доля самок,%
|
|
1
|
2
|
66,7
|
0
|
3,3
|
33,3
|
66,7
|
|
2
|
7
|
57,1
|
7,2
|
35,7
|
57,1
|
42,9
|
|
3
|
19
|
84,5
|
15,5
|
0
|
50
|
50
|
|
4
|
18
|
88,6
|
5,7
|
5,7
|
51,4
|
48,6
|
|
5
|
22
|
100
|
0
|
0
|
56,8
|
43,2
|
Таблица 3
Демографическая
структура лесной мыши на площадке 2
|
турОбщее обилие, экз.Доля
неполо- возрелых,%Доля поло-возрелых,%Доля полово- зрелых
пере-зимовавших,%Доля самцов,%Доля самок,%
|
|
|
|
|
|
|
|
1
|
24
|
0
|
76,6
|
52
|
48
|
|
2
|
19
|
23,7
|
31,6
|
44,7
|
65,8
|
34,2
|
|
3
|
20
|
43,6
|
43,6
|
12,8
|
59
|
41
|
|
4
|
39
|
87
|
10,4
|
2,6
|
57,1
|
42,9
|
|
5
|
20
|
92,3
|
7,7
|
0
|
56,4
|
43,6
|
Характеризуя сезонную динамику возрастного состава исследуемого вида
нужно отметить, что в начале сезона преобладали половозрелые животные, а к
сентябрю неполовозрелые составили 92,3%. Значительна средняя доля
неполовозрелых сеголеток - 54,1%.
Соотношение полов у лесной мыши на протяжении всего периода размножения
смещено в пользу самцов. Средняя за сезон доля половозрелых самок составляла
22,8%. Интересным оказался тот факт, что в середине мая при достаточно близком
соотношении прибылых и перезимовавших зверьков прибылые животные неполовозрелы.
Подобная картина может наблюдаться в лишь том случае, если размножение
начинается значительно позже.
4. ДИНАМИКА
МИГРАЦИОННОЙ АКТИВНОСТИ НАСЕЛЕНИЯ ЛЕСНОЙ МЫШИ
Дифференцированность территориальных участков предполагает наличие
миграционных потоков. Целостность населения популяции, разобщенного
незаселенными пространствами, поддерживается, как правило, иммиграцией
неполовозрелых животных с территории благоприятных биотопов и станций
переживания через транзитные участки и образованием временных репродуктивных
поселений [29,43]. Способность животных к активным перемещениям обусловливает
высокую лабильность пространственной организации их популяций, чем определяется
особая ее роль, как механизма быстрого реагирования на изменения условий среды
обитания. Определение мотиваций миграционного поведения животных и факторов,
влияющих на его активность, имеет большое значение для понимания
внутрипопуляционных процессов и функционирования популяции как целого.
Под миграциями подразумевается процесс покидания индивидуального участка,
перемещения и занятие новой территории, что наблюдается практически у всех
видов млекопитающих.
Еще в 30-50 годы было показано, что население локальных территорий
состоит из животных, относящихся к двум основным группам: постоянные обитатели,
перемещающиеся в пределах этой территории и посетители, пересекающие ее
[44,45,46]. В настоящее время эти две группы животных называют: оседлые -
совершающие регулярные перемещения в пределах своего индивидуального участка
обитания; мигранты - совершающие непривязанные к конкретной территории
перемещения. Сформировалось два взгляда на явления "оседлости -
миграции" у мелких млекопитающих. Ряд исследователей считает, что для
мелких млекопитающих нехарактерно перемещение на далекие расстояния, связанные
с покиданием индивидуального участка. Поэтому миграции они рассматривают как
случайные перемещения, а их влияние на динамику численности и демографическую
структуру популяций - минимальным. Другая часть авторов полагает, что для
мелких млекопитающих характерна значительная миграционная активность,
являющаяся одним из механизмов регуляции численности популяции [47,48,49]. При
этом одни из них предполагают, что каждое животное в популяции на протяжении
жизни имеет статус либо "мигранта", либо "оседлого". С
точки зрения вторых, миграционное поведение свойственно практически каждой
особи, и в течение жизни ее переход из статуса "мигрант" в статус
"резидент" может происходить много раз.
Вопрос о биологической специфичности мигрантов и оседлых животных
рассматривался с разных сторон (этологии, морфологии, фенетики, генетики и
т.д.). Но до сих пор не имеет окончательного решения, хотя все большее число
исследователей склоняется к тому, что резиденты и мигранты разнокачественны по
целому ряду показателей.
Предполагается, что оседлый образ жизни дает ряд преимуществ. В первую
очередь к ним относится знакомство с территорией, позволяющее животным наиболее
рационально использовать кормовые и защитные условия местообитаний. Кроме того,
оседлый образ жизни позволяет животному регулировать свою "социальную
среду" на основе направленных контактов между особями. Знакомство с
территорией и относительное постоянство социального окружения позволяют
поддерживать ранг животного в системе иерархии данной группировки и
доминирование особей, обладающих участками, над мигрантами.
В современной литературе известны разные классификации перемещений. Мы
придерживаемся классификации, предложенной Лидикером [50], для мелких
млекопитающих. Немиграционные перемещения: номадизм; экскурсии.
Миграции: собственно миграции; миграции насыщения; миграции донасыщения:
а) сезонные; б) онтогенетические; в) колонизирующие; г) конкурентные.
В качестве немиграционных здесь рассматриваются два типа перемещений. К
ним относятся номадизм, при котором участок обитания не организуется и экскурсии,
связанные с исследовательскими выходами за пределы участка обитания, при этом
животные оставляют свой участок на короткий срок, после чего возвращаются
обратно. Такие "походы" могут стать началом миграционного поведения
или попытками найти партнера для спаривания. Конечно, если животное не
возвращается из такой экскурсии по какой-либо причине, экскурсия автоматически
переходит в разряд миграционных перемещений. Такие перемещения отмечены для
мелких млекопитающих многими исследователями.
Перемещения считаются миграционными в тех случаях, когда животное
покидает свой индивидуальный участок или навсегда, или, по крайней мере, на
значительный период времени. Собственно миграции включают в себя процесс
покидания индивидуального участка, переход на значительные расстояния с
организацией нового участка обитания с возможным последующим возвращением
обратно. Как правило, такие явления сезонно упорядочены и включают в себя
перемещения большей части популяции. Наиболее известны сезонные изменения
местообитаний у леммингов, эти данные обобщены Стенсетом [51]. Известны случаи
регулярных сезонных перемещений грызунов в периодически затопляемых
местообитаниях. Миграции насыщения - вспомогательный механизм, направленный на
недопущение переуплотнения населения и обеспечивающий оптимальное использование
территории. Такие процессы в литературе описаны достаточно широко [50]. Миграционная активность животных
рассматривается при этом как механизм снижения плотности. Считается, что
мигранты в таких условиях представлены особями низкого социального ранга и
обречены на гибель. Как правило, это неполовозрелые животные, расселяющиеся
из-за недостатка жизненных ресурсов.
В зависимости от мотиваций миграционные перемещения формируются разными
внутрипопуляционными группировками. У некоторых видов беременные или
лактирующие самки мигрируют на новые участки, оставляя прежние последнему
выводку. В то же время, для миграций другого типа характерно преобладание среди
мигрантов неполовозрелых животных. Среди мелких млекопитающих, как правило,
самцы более подвижны, хотя известны многочисленные случаи, когда оба пола
мигрируют одинаково [50]. Соотношение полов в составе мигрирующих животных так
же зависит от типа миграций. Например, среди мигрантов насыщения различий между
миграционной активностью самцов и самок может не наблюдаться.
Мигранты донасыщения - как правило, половозрелые животные низкого
социального ранга, принужденные к расселению агрессией доминантов. Лидикер [50] предлагает четыре типа миграций
донасыщения. Эти случаи не являются исчерпывающими и взаимоисключающими.
. Сезонные миграции. Регулярно происходят в определенные сезоны года, не
зависят от плотности и других потенциально важных факторов. Примером является
реорганизация структуры популяций мелких млекопитающих перед началом сезона
размножения.
. Онтогенетические миграции. Имеют место на конкретной стадии развития
животных. У мелких млекопитающих, например, наибольший уровень миграционной
активности отмечен для неполовозрелых животных.
. Колонизирующие миграции. Миграции, при которых животные перемещаются на
незанятые территории и организуют там собственные участки обитания. Эти
перемещения могут являться продолжением других типов миграций.
. Конкурентные миграции. Миграции, которые вызываются присутствием других
видов, делающих среду обитания менее благоприятной. Обычно такими видами
являются хищники, паразиты или конкуренты.
Очевидно, что миграции представляют собой очень разнородное явление и
могут иметь множество причин и последствий, разнообразную мотивацию и графики
протекания.
Одним из важнейших вопросов является изучение непосредственной связи
миграций с численностью населения. Существуют различные гипотезы,
подчеркивающие потенциальную роль миграций в регуляции плотности популяций и
наличие линейной или нелинейной связи этих параметров [41,42]. В литературе
описаны примеры как положительной, так и отрицательной зависимости миграционной
активности животных от уровня их численности [52,53,54].
Анализ литературных данных показывает наличие взаимосвязей между
миграциями, пространственной структурой популяций и уровнем общей численности.
В настоящее время актуальным является изучение характера таких взаимосвязей
[55,56,57,58].
Согласно литературным данным, доля мигрантов в популяции может достигать
существенных значений и это, несомненно, делает миграционную активность
животных одним из факторов, влияющих на структуру популяции в цело.
Дифференцированность территориальных участков предполагает наличие миграционных
потоков, оказывающих непосредственное влияние на формирование пространственного
распределения животных. Интенсивность миграционной активности определяется
комплексом условий, наиболее существенные среди которых биотопические условия,
уровень численности и репродуктивное состояние животных.
Учитывая роль миграционных процессов в формировании популяционной
структуры, представляется необходимым проведение анализа миграционной
активности животных с целью выявления характера связи интенсивности
миграционных перемещений с динамикой демографических характеристик, пространственной
структуры и биотопическими условиями территории.
Средняя доля мигрантов лесной мыши на первой площадке составляла 37,2%.
Миграционная активность населения снижалась с начала сезона к осени на фоне
роста численности (рис.1). Значительный прирост численности неполовозрелых
животных при низкой доле размножающихся зверьков в течение всего летнего сезона
свидетельствует о том, что территория первой площадки используются лесной мышью
в качестве зоны расселения неполовозрелых сеголеток.
Демографическая структура лесной мыши на второй площадке характеризует
этот биотоп как благоприятный для ее обитания и размножения. Миграционная
активность лесной мыши на второй площадке почти в два раза ниже, чем на первой
(средние за сезон 2000 г. доли мигрирующих животных 37,2% и 19,8%
соответственно). С июня до конца июля отмечалось снижение миграционной
подвижности населения на фоне увеличения общей численности при наименьшей за
сезон доле мигрантов среди половозрелых животных (2,8%). В августе численность
лесной мыши достигла сезонного пика, что привело к увеличению частного обилия и
доли используемой территории до максимальных значений, и снижению доли
мигрантов до минимума (рис.2). Снижение численности к сентябрю сопровождалось
увеличением миграционной активности. Возможно перемещение животных в соседние
биотопы, в частности со второй площадки на первую. Среди мигрирующих животных
преобладают неполовозрелые особи (в среднем за сезон 30%).

Рисунок
1 Миграционная активность лесной мыши на площадке 1
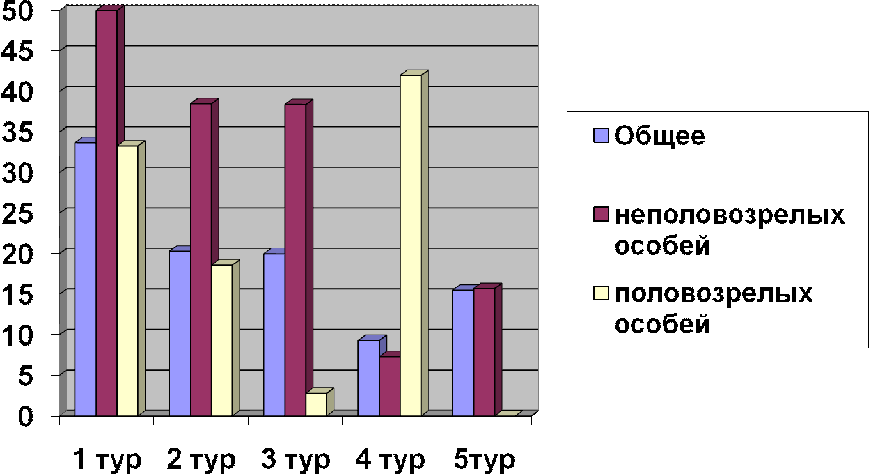
Рисунок 2 Миграционная активность лесной мыши на площадке 2
5. ПРОСТРАНСТВЕННАЯ СТРУКТУРА НАСЕЛЕНИЯ ЛЕСНОЙ МЫШИ
Известно, что распределение животных по территории неравномерно и во
многом определяется биотопическими особенностями. При этом неоднородность
местообитаний входит в число ведущих факторов, определяющих пространственную
структуру популяции, в зависимости от ее выраженности можно наблюдать весь
спектр распределений животных на территории (от равномерного до
агрегированного). В гетерогенных местообитаниях выделяются зоны, существенно
различающиеся по степени их использования животными [59,60,61,62,63]. К таким
территориям относятся: резервации, временные поселения и транзитные участки.
Резервации, или донорные участки, характеризуются наиболее благоприятными для
проживания и размножения условиями среды. Животные обитают на них постоянно.
Временные поселения позволяют размножаться и существовать в них только на
протяжении определенных сезонов. Транзитные участки используются для
проживания, но практически непригодны для размножения.
Таким образом распределение животных по территории в значительной мере
определяется гетерогенностью местообитаний. Сезонные изменения биотопических
условий приводят к перераспределению территории между особями разных возрастных
и генеративных состояний. Такие изменения пространственно-демографической
структуры происходят посредством миграционных перемещений. В связи с этим
особый интерес представляет изучение сезонной динамики пространственного
распределения лесной мыши во взаимосвязи с демографической структурой населения
различных биотопов.
5.1 Пространственное распределение лесной мыши на площадке 1
Особенности демографической структуры лесной мыши показывают, что первая
площадка используется этим видом для расселения неполовозрелых животных. Общая
численность растет на протяжении всего сезона на фоне увеличения доли
используемой территории и снижения индекса агрегированности (рис.4,5 ).
Можно отметить, что в мае животные практически не отлавливались. С мая до
начала июня рост численности населения сопровождался увеличением частного
обилия (с 25 до 33) и площади занимаемой территории (с 1.6 до 5%). Эти
процессы, вероятно, связаны с заполнением наиболее благоприятных микроучастков
животными, приходящими на первую площадку из прилегающего леса. Затем, до
середины августа частное обилие лесной мыши постоянно снижалась одновременно с
увеличением доли используемой территории (рис.3), что обусловлено дальнейшим
расселением приходящих из соседних участков леса животных. Несмотря на
продолжающийся рост общей численности с августа до сентября, частное обилие не
изменялось и оставалось на минимальном уровне (рис.3). Следует отметить, что в
этот период население лесной мыши состояло на 100% из неполовозрелых сеголеток.
Низкое частное обилие поддерживалось за счет увеличения используемой территории
в сторону менее благоприятных для вида участков.
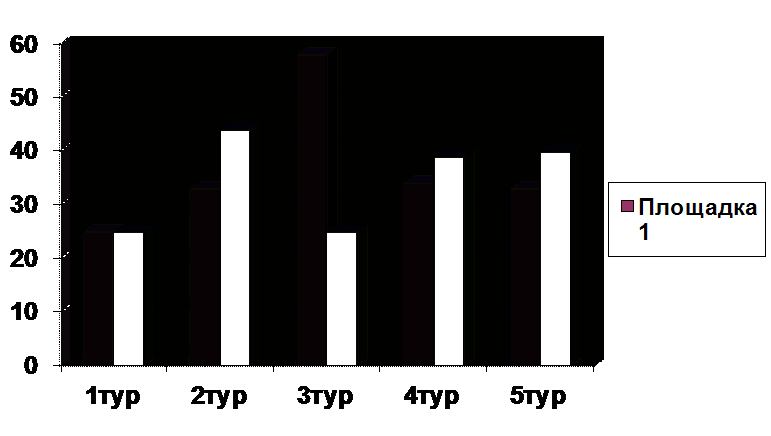
Рисунок
3 Частное обилие лесной мыши
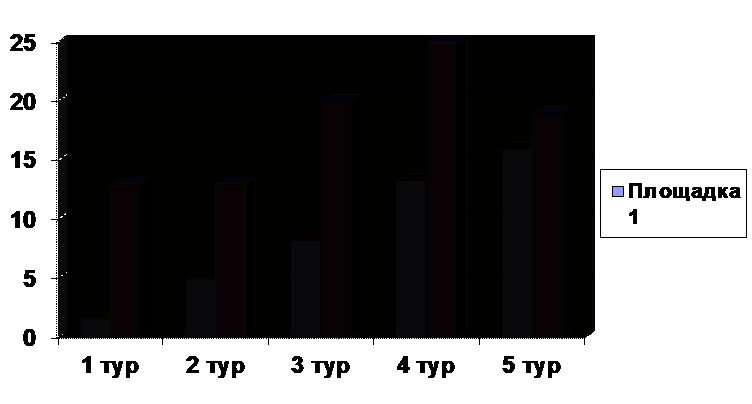
Рисунок 4 Заселенность лесной мыши
Рисунок
5 Агрегированность населения лесной мыши
5.2 Пространственное распределение лесной мыши на площадке 2
лесной мышь демографический миграционный
Показатели, характеризующие пространственную структуру населения лесной
мыши на второй площадке, соответствует предположению об использовании видом
территории первой площадки в качестве зоны временного расселения молодняка, а
участка леса, на котором расположена вторая площадка для постоянного обитания и
размножения. Неодинаковое использование территории определяет различия
демографической структуры и пространственного распределения лесной мыши на
первой и второй площадках.
Так с мая до начала июня уровень частного обилия и степень скученности
лесной мыши возросли на фоне увеличения численности, при этом площадь
используемой видом территории изменилась незначительно (рис. 4). В следующий
период, до конца июля, распределение животных по площадке стало более
равномерным, так как общий уровень частного обилия снизился, а площадь
используемой территории значительно возросла.
Размножение лесной мыши вызвало значительный прирост численности
неполовозрелых животных к августу, что привело к увеличению частного обилия и
доли используемой территории. К сентябрю численность лесной мыши снизилась с 39
до 20. Сопоставляя этот факт с ростом численности на соседней первой площадке,
можно предположить, что высокая численность и значительный уровень частного
обилия лесной мыши на второй площадке при максимальной площади используемой
территории в августе привели к выселению части животных в менее благоприятные,
но приемлемые для обитания участки, прилегающие к лесу. В результате
миграционных перемещений животных численность, как и доля используемой
территории на обеих площадках практически выровнялась - если в августе обилие
населения животных на второй площадке более чем в два раза превысило этот
показатель на первой, то в сентябре численность лесной мыши составляла 20
соответственно.
ЗАКЛЮЧЕНИЕ И
ВЫВОДЫ
Определение характера взаимосвязей между демографической структурой и
пространственным распределением, миграционной активностью животных является
важным аспектом при рассмотрении популяционных процессов.
Согласно существующим классификациям, в гетерогенных местообитаниях
выделяются зоны, существенно различающиеся по степени их использования
животными. Так, участок леса можно отнести к донорному, а степной - к зоне
временного расселения лесной мыши. Вторая площадка используется для постоянного
обитания, а первая - для временного проживания. Сезонные изменения
пространственного распределения определяются совокупным действием различных
факторов, ведущее место среди которых занимают территориальная неоднородность
местообитаний, демографическая структура населения и репродуктивная активность
животных. Популяционные параметры изменяются в течение репродуктивного цикла,
который можно разделить на следующие функциональные состояния: начало
размножения, период интенсивного размножения, сезонный пик численности и конец
сезона размножения.
Для начала размножения характерны низкий уровень численности и
значительные изменения популяционных параметров, в том числе пространственного
распределения. Пространственное распределение лесной мыши в это время можно
охарактеризовать как агрегированное с низкой степенью заселенности.
В период интенсивного размножения по мере роста численности сеголеток
отмечалось снижение степени агрегированности животных и увеличение заселенности
территории в связи с расселением молодняка в участки с меньшей плотностью и,
по-видимому, менее благоприятные для размножения, но вполне пригодные для
обитания. Таким образом, на уровне площадок общий тип распределения животных в
начале сезона размножения можно охарактеризовать как агрегированный, в течение
сезона переходящий в более равномерный по мере роста числа неполовозрелых
сеголеток.
Сезонному пику численности, как правило, соответствует наибольшая доля
используемой территории, высокие значения частного обилия вследствие
максимального использования пригодных для обитания микроучастков.
Снижение общей численности и доли занятой территории в конце сезона
размножения достигается выселением части сеголеток на соседние, незаселенные
участки. Вероятно, миграции неполовозрелых зверьков в этом случае являются
альтернативой их гибели.
В биотопе, используемом для расселения и временного проживания, в связи с
ограниченностью количества пригодных для обитания животных участков, степень
агрегированности выше, чем в местах постоянного обитания (средний индекс
агрегированности у лесной мыши на первой площадке 7,46; на второй - 2,38).
Увеличение скученности лесной мыши к осени на участке, расположенном в степи
обусловлено сосредоточением животных в некоторых локально расположенных
участках территории площадки, биотопические условия которых приемлемы для обитания.
Таким образом, динамику пространственного распределения животных
невозможно объяснить исключительно изменениями уровня численности, как и
другими отдельно взятыми популяционными параметрами.
Простанственно-демографическая структура населения определяется комплексом
факторов, основная роль в котором принадлежит степени гетерогенности среды
обитания и репродуктивной активности животных. В свою очередь, репродуктивные
процессы определяются сложными взаимодействиями большого количества, как
внешних факторов, так и внутрипопуляционных механизмов.
На основании комплексного изучения сезонных изменений демографической
структуры, пространственного распределения и миграционной активности лесной
мыши, обитающей в различных биотопических условиях можно сделать следующие
выводы:
. Миграционные перемещения в течение репродуктивного сезона связаны с
неоднородностью местообитаний. Гетерогенность условий среды вызывает
миграционные потоки, вызывающие снижение уровня численности.
. Наличие участков с резко различающимися биотопическими условиями
вызывают направленные перемещения из благоприятных перенаселенных местообитаний
в менее заселенные, определяя характер пространственного распределения
животных.
. Динамика миграционной активности зависит от репродуктивного состояния животных.
Отмечается высокий уровень миграционной активности в начале сезона размножения,
а в период репродуктивной активности интенсивность перемещений снижена.
. Динамика пространственной структуры зависит от биотопической
неоднородности и репродуктивной активности животных. В начале репродуктивного
сезона отмечается высокая степень агрегированности. В местах постоянного
обитания рост численности сопровождается снижением агрегированности и более
полным использованием территории в результате расселения неполовозрелых особей.
. В биотопе, используемом для расселения и временного проживания степень
агрегированности выше, чем в местах постоянного обитания животных.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Шварц С.С.
Популяционная структура вида // Зоол. журн. 1967. Т. 46, вып. 10. С. 1456-1469.
. Шварц С.С.
Эволюционная экология животных. Свердловск, 1969. 200 с. (Тр. Ин-та экологии
растений и животных УФАН СССР; Вып. 65.)
. Шварц С.С.
Экологические закономерности эволюции. М.: Наука, 1980. 277 с.
. Шилов И.А.
Принципы организации популяций у животных // Популяционные проблемы в
биогеоценологии. М., 1988. С.5-23.
. Шилов И.А.
Популяционный гомеостаз у животных // Бюл. МОИП. Отд. биол. 1982. Т. 87, вып.
4. С. 23-32.
. Шилов И.А.
Популяция как биологическая система надорганизменного уровня // Экология
популяций: структура и динамика. М., 1995. Ч. 1. С. 3-13.
. Флинт В.Е.
Пространственная структура популяций мелких млекопитающих. М.: Наука, 1977. 183
с.
. Большаков
В.Н., Баженов А.В. Радионуклидные методы мечения в популяционной экологии
млекопитающих. М.: Наука, 1988. 158 с.
. Шилов И.А.
Эколого-физиологические основы популяционных отношений у животных. М.: Изд-во
МГУ, 1977. 262 с.
. Наумов Н.П.
Структура популяций и динамика численности наземных позвоночных // Зоол. журн.
1967. Т. 46, вып. 10. С. 1470-1486.
. Наумов Н.П.
Пространственная структура вида млекопитающих. // Там же. 1971. Т. 50. вып. 7.
С. 965-980.
. Шилов И.А.,
Калецкая М.П., Ивашкина И.Н., Солдатова А.Н. Пространственная структура
популяции полевок-экономок (Microtus oeconomus) в Дарвиновском заповеднике //
Зоол. журн. 1977. Т. 56, вып.2. С.257-267.
. Одум Ю.
Экология: Пер. с англ. М.: Мир, 1986. Т. 2. 376 с.
. Наумов Н.П.
Экология животных. М.: Сов. наука, 1951б. 524 с.
. Никитина Н.
А., Меркова М. А. Использование территории мышами и полевками по данным мечения
//Бюл. МОИП. Отд. биол. 1963. Т. 68, вып.5. С.18-36.
. Никитина
Н.А. О размерах индивидуальных участков грызунов фауны СССР //Зоол. журн. 1972.
Т. 51, вып. 1. С. 119-126.
. Никитина
Н.А. К понятию "индивидуальный участок" у мелких млекопитающих //I
Междунар. териол. конгр.: Реф. докл. М., 1974. Т. 2. С. 71-72.
. Никитина
Н.А. Рыжие полевки // Итоги мечения млекопитающих. М., 1980. С. 189-219.
. Окулова
Н.М., Аристова В.А., Кошкина Т.В. Влияние плотности популяции на размер
индивидуальных участков у мелких грызунов в тайге Западной Сибири и на Дальнем
Востоке // Тез. докл. 3 Всесоюз. териолог. съезда. М., 1982. Т. 1. С. 260-265.
. Демидов
В.В., Хлебалов И.Я. Наблюдения за использованием территории мелкими грызунами в
южной тайге Камского Приуралья. Перм. гос. ун-т. Пермь, 1981. 18 с. Деп. в
ВИНИТИ 20.04.81, N1737-81 Деп.
. Окулова
Н.М. Биологические взаимосвязи в лесных экосистемах: (На прим. природ. очагов
клещевого энцефалита). М.: Наука, 1986. 248 с.
. Никитина
Н.А., Карулин Б.Е., Литвин Ю.В. и др. О размерах суточной территории и
вероятном строении индивидуальных участков у некоторых видов грызунов // Зоол.
журн. 1977. Т. 56, вып. 12. С. 1860-1869.
. Кутенков
А.П. Об особенностях использования территории рыжей полевкой в заповеднике
“Кивач”// Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР.
Петрозаводск, 1978. С. 120-127.
. Хляп Л.А.
Основные проблемы изучения связей мелких млекопитающих с окружающим
пространством // Фауна и экология грызунов. М., 1980. Вып. 14. С. 44-62.
. Хляп Л.А.
Особенности использования и структура участков обитания бурозубок и лесных
полевок // Там же. 1983. Вып.15. С. 162-200.
. Наумов Н.П.
Мечение млекопитающих и изучение их внутривидовых связей // Зоол. журн. 1956.
Т. 35, вып. 1. С. 3-15.
. Никитина
Н.А. О постоянстве использования территории грызунами // Фауна и экология
грызунов. М., 1970. Вып. 9. С. 110-133.
. Крылов Д.Г.
Использование территории Clethrionomys glareolus, Schreber (Rodentia) в условиях средней полосы
Европейской части СССР//I
Междунар. териол. конгр. М., 1974. Реф. докл. Т.1. С. 317.
. Кутенков
А.П. Использование территории рыжей полевкой (Clethrionomys glareolus) // Зоол. журн. 1979а. Т. 58, вып.
1. С. 97-104.
. Ралль Ю.М.
Характер передвижений мышевидных грызунов на небольших площадках // Зоол.журн.
1936. Т. 15, вып. 3. С.472-482.
. Марин Ю.Ф.
Абсолютная численность красно-серой и красной полевок и использование ими
территории в среднегорье Северо-Восточного Алтая // Изв. Сиб. Отд. АН СССР.
Сер. биол. наук. 1976. Т. 15, вып. 3. С. 80-85.
. Кутенков
А.П. Использование территории европейской рыжей полевкой (Clethrionomys glareolus Schreber) в разных типах леса заповедника
«Кивач»: Автореф. дис. канд. биол. наук. М., 1979б. 21 с.
. Бигон М.,
Харпер Дж., Таусенд К. Экология: Особи, популяции и сообщества. М.: Мир, 1989.
Т.1. 667 с.
. Смуров
А.В., Полищук Л.В. Количественные методы оценки основных популяционных
показателей: Стат. и динам. аспекты. М.: Изд-во МГУ, 1989. 208 с.
.
Whitford P.B. Distribution of woodland plants in relation to succession and
clonal growth //Ecology. 1949.
Vol. 30. P. 199-208.
. Смуров А.В.
Новый тип статистического пространственного распределения и его применение в
экологических исследованиях // Зоол. журн. 1975а. Т. 54, вып. 2. С. 283-289.
. Жигарев
И.А. Мелкие млекопитающие рекреационных и естественных лесов Подмосковья.
М:Прометей 2004.232 с.
. Литвинов
Ю.Н Методы изучения сообществ мелких наземных позвоночных животных.
Новосибирск: НГУ, 2001.52с.
.Тупикова
Н.В. Определение возраста грызунов //Фауна и экология грызунов.1957. Вып.5.
. Лукьянов
О.А. Феноменология и анализ миграций в популяциях мелких млекопитающих:
Автореф.дис.док. биол. наук. Екатеринбург, 1997. 46 с.
.Лукьянов
О.А. Анализ процессов миграции в популяциях мелких млекопитающих // Там же.
1993. N1. С. 47-62.
. Лукьянов
О.А. Феноменология и анализ миграций в популяциях мелких млекопитающих //Зоол.
Журнал 2002 г. Т.81 N9 С. 1107-1134
. Груздев
В.В. Территориальные центры и районы благоденствия и прозябания популяций //
Популяционная структура вида у млекопитающих. М., 1970. С. 7-10.
. Ралль Ю.М.
Динамическая плотность грызунов и некоторые методы ее изучения // Бюл. МОИП.
Отд.биол. 1945. Т. 50. вып.5-6. С. 62-64
. Наумов Н.
П. Новый метод изучения экологии мелких лесных грызунов // Фауна и экология
грызунов. М., 1951а. Вып. 4. С. 3-21.
. Наумов Н.П.
Изучение подвижности и численности мелких млекопитающих с помощью ловчих
канавок // Вопросы краевой, общей и экспериментальной паразитологии и
медицинской зоологии. М., 1955. Т. 9. С. 179-202.
.. Щипанов
Н.А., Олейниченко В.Ю., Касаткин М.В. Функции внутрипопуляционных
пространственных структур и синантропизм мелких млекопитающих // Синантропия
грызунов и ограничение их численности. М., 1992. С. 88-110.
. Щипанов
Н.А. Функциональная организация популяции (на прим. мелких млекопитающих):
Автореф. дис.док. биол. наук. Москва, 1996. 47 с.
. Щипанов
Н.А. Оценка плотности населения оседлых и величины потока нетерриториальных
мелких млекопитающих при учетах с безвозвратным изъятием // Там же. 1990. Т. 69, вып. 5. С. 113-123.
.
Lidicker W.Z.(Jr.). The role of dispersal in the demography of small mammals //
Small Mammals: Their Production and Population Dynamic. London, 1975. P.
103-128.
. Белан О.Р.,
Жигальский О.А. Динамика пространственной структуры популяций полевок
Иремельского горного массива // Фауна и экология животных Южного Зауралья и сопредельных
территорий. Екатеринбург, 1995. С. 116-124.
. Бердюгин
К.И., Киселева Е.М., Садыков О.Ф. Грызуны каменистых россыпей Урала //
Экология, методы изучения и организации млекопитающих горных областей.
Свердловск, 1977. С. 76-78.
. Бердюгин
К.И., Дороватовский С.А. Вертикальное распределение грызунов в горах Урала //
Млекопитающие Уральских Гор. Свердловск, 1979. С. 4-5.
.
Млекопитающие Печеро-Илычского заповедника. Сыктывкар: Коми. книж. изд-во,
2004. 463 с.
. Окулова Н.
М., Катаев Г. Д. Многолетняя динамика численности красно-серой полевки в разных
частях ареала //Зоол. жур. 2003. Т82 №9. С. 1045-1111.
. Садыков
О.Ф., Бененсон И.Е. Динамика численности мелких млекопитающих: концепции,
гипотезы, модели. М.: Наука, 1992. 190 с.
. Чернявский
Ф. Б., Лазуткин А.Н. Циклы леммингов и полевок на Севере. Магадан, 2005. 150 с.
. Большаков
В.Н. Пути приспособления мелких млекопитающих к горным условиям. М.: Наука,
1972. 200 с.
. Большаков
В.Н. Высотное распределение мелких млекопитающих Уральских Гор и отличия биопродуктивности
// 3 Всесоюз. Совещ. «Вид и его продуктивность в ареале» (Паланга, 18-19 сент.
1980 г). Вильнюс, 1980. С. 117-118.
. Бердюгин
К.И. Территориальные взаимоотношения грызунов - обитателей каменистых россыпей
Урала // Внутри- и межпопуляционная изменчивость млекопитающих Урала.
Свердловск, 1980. С. 37-53.
. Садыков
О.Ф., Баженов А.В. Очерк экологии лесных полевок Уральских гор // Экология
лесных полевок: (Эколого-биохим. особенности мелких млекопитающих). Свердловск,
1984. С. 3-31.
. Садыков О.Ф.
Пространственная структура горных популяций полевок рода Clethrionomys // Популяционная экология и
морфология млекопитающих. Свердловск, 1984. С. 20-36.