|
Ферменты
|
Реакции
|
Переносимая
группа
|
|
Глутаматмутаза
|
L-трео-р-метиласпарат L-глу - тамат
|
-СН (NHa) СООН
|
|
Метил
малонил-КоА-му-
|
сукцинил КоА +
Ь^-метилмало-
|
-COS-KoA
|
|
Таза
|
нил-КоА
|
|
|
2 -
Метилепглутаратму-
|
метилитаконат -
2-Метиленглута-
|
-С (=С На) СООН
|
|
Таза
|
Рат
|
-NHa
|
|
L-P-Лизинмутаза
|
|
|
D-a-Лизинмутаза
|
2,5-диаминогексаноат
D-a-ли - зин
|
-NHa
|
|
О-а-Орнитчшмутаза
|
2,4-диаминогексаноат
- "-"■ D-a ор - нитин
|
-NHa
|
|
L-a-Лейцинмутаза
|
3-аминоизокапронат
L-a лейцин
|
-NHa
|
|
1,2-Диолдегидратаза
|
этиленгликоль -
*■ ацетальдегид ■++ НаО
|
-ОН
|
|
1,2-пропандиол
- *■ пропионовый альдегид + Н20
|
-ОН
|
|
Глицериндегидрогеназа
|
Глицерин
3-оксипропионовый альдегид + НаО
|
-ОН
|
|
Этиленгликоль -
> ацетальдегид + + н;. о
|
-ОН
|
|
1,2-Пропандиол
- *■ пропионовый альдегид - ) - НаО
|
-он
|
|
Этаноламиндезамииаза
|
Этаноламин
ацетальдегид + +
nh4
|
-NHa
|
|
2-аминопропанол
- ► пропионовый альдегид + NH4
|
-NH2
|
|
Рибоиуклеотидредук-
|
D-рибонуклеотид - ► D-2-дезокси-
|
Нет
|
|
Таза
|
рибонуклеотид 4
- НгО
|
|
Глава 2.
Методы исследования
2.1
Продуценты витамина В12
В природе витамин В12 и родственные корриноидные
соединения находят в клетках микроорганизмов, в тканях животных и некоторых
высших растениях (горох, лотос, побеги бамбука, листья и стручки фасоли).
Однако происхождение витамина В12 в высших растениях окончательно не
установлено. Такие низшие эукариоты, как дрожжи и мицелиальные грибы,
корриноиды, по - видимому, не образуют. Организм животных не способен к
самостоятельному синтезу витамина. Среди прокариота способность к биосинтезу
корриноидов широко распространена. Активно продуцируют витамин В12
представители рода Propionibacterium. Природные штаммы пропионовокислых
бактерий образуют 1,0 - 8,5 мг/л корриноидов, но получен мутант P. shermanii М
- 82, с помощью которого получают до 58 мг/л витамина.
В семействе Propionibacberiaceae есть и другие
представители, способные к высокому накоплению витамина В12 в
клетках. Это прежде всего Eubacterium limosum (Butyribacterium rettgerii). Как
продуценты витамина практический интерес имеют многие представители
актиномицетов и родственных микроорганизмов. Истинный витамин В12 в
значительных количествах синтезирует Nocardia rugosa. Путем
мутаций и отбора получен штамм N. gugosa, накапливающий до 18
мг/л витамина В12. Активные продуценты витамина обнаружены среди
представителей рода Micro - monospora: М. purpureae, М. echinospora,
М. halophitica.М. fused, М. chalceae. Высокой
кобаламинсинтезирующей активностью обладают метаногенные бактерии, например, Methanosarcina
barkeri, М. vacuolata и отдельные штаммы галофильного вида Methanococcus
halophilus. Последний организм синтезирует более 16 мг корриноидов на
грамм биомассы. Столь высокого содержания корриноидов не отме ieno ни у одного
другого из изученных микроорганизмов. Причина высокого содержания корриноидов у
метаногенных бактерий не установлена. Корриноиды синтезируют строго анаэробные
бактерии из рода клостридий. У Clostridium tetanomorphum и Cl.
sticklandii аденозилкобаламин входит в состав ферментных систем,
катализирующих специфические реакции изомеризации таких аминокислот, как
глутаминовая, лизин и орнитин. В значительных количествах образуют витамин В12
ацетогенные клостридии Cl. thermoaceticum, Cl. for
- micoaceticum и Acetobacter ivoodi, синтезирующие ацетат
из С02.
Продуценты витамина B12 культивируют в средах, приготовленных на основе
пищевого сырья: соевой муки, рыбной муки, мясного и кукурузного экстракта. В
последние годы выявлены микроорганизмы, образующие высокие качества корриноидов
при утилизации непищевого сырья. Achromobacter sp., используя изопропиловый
спирт как источник углерода и энергии, накапливает до 1, 1 мг/л провитамина,
Pseudomonas sp. синтезирует витамин В12 в среде с метанолом или
пропандиолом (до 160 мкг/л), факультативный метилотроф (FM=02T) образует в среде с
метанолом до 2,6 мг/л витамина. Выделен штамм Klebsiella 101, образующий
большое количество корриноидов в клетках только при росте на среде с метанолом
как единственном источнике углерода и энергии.
2.2 Биосинтез
витамина В12
Далее образуется порфобилиноген, возникающий при конденсации
двух молекул б - АЛК, а при конденсации четырех молекул
порфобилиногенауропорфириноген III (УПГ III). Последовательность реакций между УПГ
III и кобириновой кислотой получила экспериментальное подтверждение лишь в
последние годы. Показано, что при нарушении процесса амидирования клетки
Propionibacterium shermanii выделяют в среду соединения, названные
коррифиринами - метилированные восстановленные производные уропорфирина III. К
настоящему времени выделено и охарактеризовано три соединения: коррифирин-1
(метилкоррифирин), коррифирин-2, или сирогидрохлорин (диме-метилкоррифирин).
Меченая 6-АЛК и метионин включаются в коррифирины, а последние - в витамин В12.
Метилирование УПГ III с одновременным декарбоксилированием
боковой цепочки уксусной кислоты при С12 кольца С приводит к
расхождению путей биогенеза витамина В12 и других тетрапирролов. В
результате дальнейшего метилирования и образования С - С связи между кольцами А
и Д синтезируется кобириновая кислота.
Донором семи метильных групп, включаемых в УПГ III, является S-аденозилметионин.
Характер включения атома Со и образования непосредственной связи между кольцами
А и Д неизвестен. На следующем этапе биосинтетического пути кобириновая кислота
превращается в кобинамид (фактор В), при этом остатки карбоновых кислот
кобириновой кислоты амидируются, а к остатку пропионовой кислоты кольца Д
присоединяется аминопропанол (рисунок 2).
На этой же стадии происходит присоединение к корриновому кольцу
5-дезоксиаденозильной группы и кобириновая кислота (рисунок 3) становится
5-дезоксиаденозилкобинамидом (коферментная форма фактора В). Аминопропанол,
видимо, образуется при декарбоксилировании L-треонина.
Далее кобинамид фосфорилируется с образованием кобинамидфосфата
и, реагируя с гуанозинтрифосфатом (ГТФ), дает кобинамидгуанозиндифосфат. В
коферментную форму кобинамидгуанозиндифосфата происходит включение нуклеотида и
образование кобаламин-5'-фосфата.
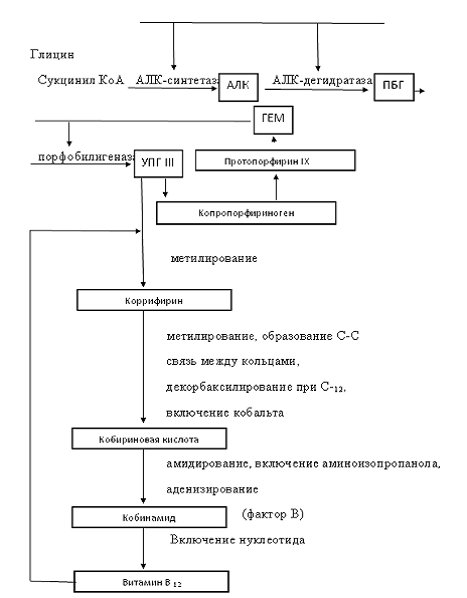
Рисунок 2. Биосинтез
витамина В12
Витамин В12, как уже отмечалось выше, содержит
специфическое азотистое основание - 5,6-ДМБ, которое в природе встречается
только в этом соединении. Непосредственным предшественником 5,6-ДМБ служит
рибофлавин. В молекулу витамина В12 5,6-ДМБ включается в виде
а-рибазол-5'-фосфата. При ферментативном дефосфорилировании
кобаламин-5'-фосфата образуется кобаламин.
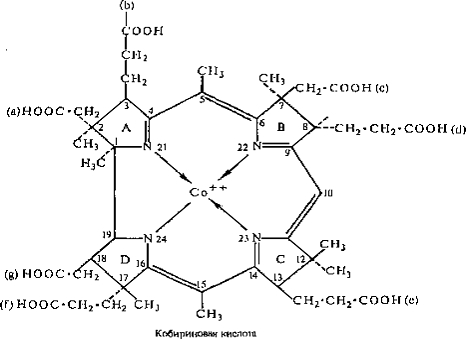
Рисунок 3 - Строение кобириновой кислоты
Витамин В12, как уже отмечалось выше, содержит специфическое
азотистое основание - 5,6-ДМБ, которое в природе встречается только в этом
соединении. Непосредственным предшественником 5,6-ДМБ служит рибофлавин. В
молекулу витамина В12 5,6-ДМБ включается в виде а-рибазол-5'-фосфата. При
ферментативном дефосфорилировании кобаламин-5'-фосфата образуется кобаламин.
Последовательность реакций при синтезе кобаламина из
кобинамида выглядит следующим образом:
) кобинамид + АТФ кобинамид-Ф + АДФ;
) кобинамид-Ф + ГТФ - *ГДФ-кобинамид + ФФИ;
) кобинамид-ГДФ + а-рибазол-5'-фосфат кобаламин-5' - фосфат +
ГМФ;
) кобаламин-5'-фосфат кобаламин + Ф".
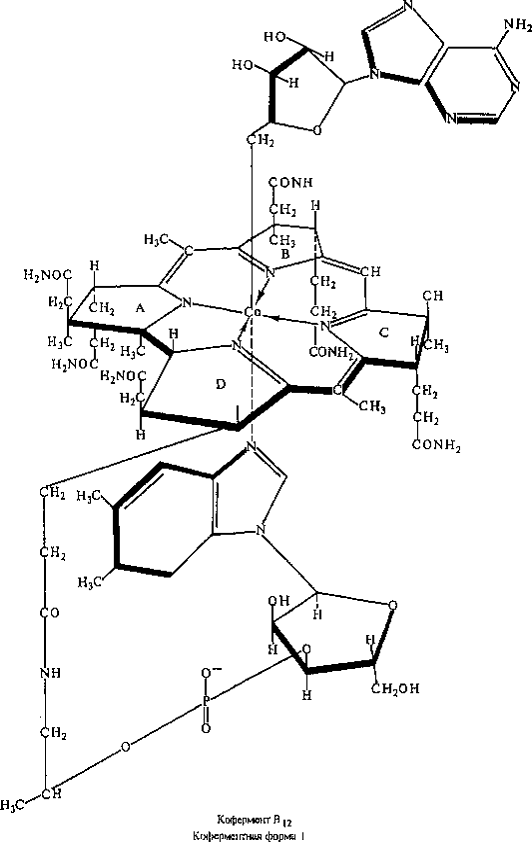
Рисунок 4 - Строение коферментной формы
Известны две коферментные формы витамина В12:
аденозилкобаламин (рисунок 4) и метилкобаламин (коферментная форма II). Более
80% всех синтезированных пропионовокислыми бактериями корриноидов находится в
коферментной форме I, которая, как уже указывалось, в качестве лиганда содержит
5-дезоксиаденозил.
Источником аденозила служит АТФ. Коферментная форма содержит
Со-С-связь и является первым кобальторганическим соединением, обнаруженным в
живых системах. Метилкобаламин обычно содержится в клетках в незначительных
количествах, но у М. barkeri метил-фактор III, который содержит в нуклеотидной
части 5-оксибензимидазол, составляет до 80% от суммы всех корриноидов.
Регуляция биосинтеза витамина В12 осуществляется по
принципу репрессии. Витамин В12 угнетает только свой собственный
синтез, не оказывая влияния на образование других тетрапирролов и, как
полагают, действует на стадии метилирования УПГ III. В качестве регулятора
выступает лишь полная нуклеотидсодержащая молекула витамина. Гем оказывает
координированную репрессию синтеза ферментов начальных стадий биогенеза тетра -
пирролов.
Функциональной формой витамина является аденозилкобаламин
(5,6-диметилбензимидазолил) - Со-5-дезоксиаденозилкобамид) и метилкобаламин (а
(5,6-диметилбензимидазолил) - Со - метилкобамид).
Как и большинство витаминов, В12 может существовать в
различных формах и принимать различные наименования. Названия для витамина B12
содержат словоформу "кобальт", так как кобальт - минерал, найденный в
центре витамина: кобринамид, кобинамид, кобамид, кобаламин, гидрокскобаламин,
метилкобаламид, аквакобаламин, нитрокобаламин, и цианокобаламин.
2.3 Получение
витамина В 12
Получение витамина В 12 с помощью
пропионовокислых бактерий
В настоящее время для получения витамина В 12
используют следующие микроорганизмы Prop, freudenreichii ATCC 6207, Prop,
shermanii ATCC 13673, Prop, shermanii BKM-103 и их варианты и
мутанты. Наибольший интерес представляют штаммы, способные к самостоятельному
синтезу 5,6 ДМБ. Поскольку синтез 5,6 ДМБ лучше происходит при доступе воздуха,
осуществляют двустадийный процесс, в котором получают наиболее высокий выход
продукта. На 1 стадии культуру выращивают в анаэробных условиях до полной
утилизации сахара. На 2-й стадии включают аэрацию, тем самым создавая условия
для синтеза 5,6 ДМБ и превращения этиокобаламнна в дезоксикобаламин. Обе стадии
осуществляют в двух разных ферментерах или в одном. Анаэробно выросшие клетки
можно собрать путем центрифугирования и инкубировать густую суспензию на
воздухе и, если нужно, в присутствии 5,6 ДМБ и цианида. Добавление ДМБ
производят только во 2-й стадии ферментации (если бактерии не синтезируют его
самостоятельно), поскольку в его присутствии образуются полные формы витамина,
ингибирующие его синтез. Среда для ферментации обычно содержит глюкозу или
инвертированную-мелассу (10 - 100 г/л), небольшие количества солей Fe, Mn и Mg,
а также Со (10 - 100 мг/л), источники азота. В среду добавляют кукурузный
экстракт (30 - 70 г/л), содержащий молочную и пантотеновую кислоты, усиливающие
рост бактерий. Пантотеновую кислоту, стимулирующую также синтез витамина,
рекомендуют вносить в среду дополнительно. Бактерии культивируют при 30°,
поддерживая рН на уровне 6,5-7,0 путем введения (NH4) OH. Ферментацию
производят в ферментерах на 500 л, содержащих 340 л среды, инокулированных 7 л
посевного материала. В первые 80 ч культура растет под небольшим давлением N2 и
слабым перемешиванием (без аэрации), в следующие 88 ч включают аэрацию (2 м3/ч)
и перемешивание. Возможны некоторые вариации в культивировании. Витамин В12
сохраняется в клетках бактерий, поэтому проводят его экстракцию:
) выделение витамина из клеток и превращение его в
цианокобаламин;
) выделение неочищенного продукта (80% чистоты), который
можно использовать в животноводстве;
) дальнейшую очистку до уровня 91-98% (для медицинских
целей).
Для экстракции витамина из клеток последние нагревают при
80°-120° в течение 10-30 мин при рН 6,1-8,5. Превращение в CN-кобаламин
достигают, обрабатывая горячий раствор или клеточную суспензию цианидом или
тиоционатом, часто в присутствии NaNO2 или хлорамина В. Корриноиды сорбируют на
различных носителях: амберлите IRC-50, А12О3, активированном угле и элюируют
водными спиртами или водно-фенольными смесями. В значительных количествах
образуют витамин В12 ацетогенные клостридии Cl. thermoaceticum, Cl. for -
micoaceticum и Acetobacter ivoodi. Из водных растворов корриноиды экстрагируют
фенолом или крезолом, или смесью этих спиртов с бензином, бутанолом,
углеродистым тетрахлоридом или хлороформом. При упаривании различных
растворителей получают осадок или кристаллы витамина, которые растворяют в
соответствующем растворителе до нужной концентрации. Природные штаммы
пропионовокислых бактерий образуют 1,0 - 8,5 мг/л корриноидов, но получен
мутант P. shermanii М-82, с помощью которого получают до 58 мг/л
витамина.
Но есть патентное сообщение (Франция) о достижении невероятно
высокого выхода - 216 мг/л.
В семействе Propionibacberiaceae есть и другие
представители, способные к высокому накоплению витамина В12 в
клетках. Это прежде всего Eubacterium limosum (Butyribacterium rettgerii). Как
продуценты витамина практический интерес имеют многие представители
актиномицетов и родственных микроорганизмов. Истинный витамин В12 в
значительных количествах синтезирует Nocardia rugosa. Путем мутаций и отбора
получен штамм N. gиgosa, накапливающий до 18 мг/л витамина В12.
Активные продуценты витамина обнаружены среди представителей рода
Micro-monospora: М. purpureae, М. echinospora, М. halophitica.М. fused, М.
chalceae. Высокой кобаламинсинтезирующей активностью обладают метаногенные
бактерии, например, Methanosarcina barkeri, М. vacuolata и отдельные штаммы
галофильного вида Methanococcus halophilus. Последний организм синтезирует
более 16 мг корриноидов на грамм биомассы. Столь высокого содержания
корриноидов не отмечeнo
ни у одного другого из изученных микроорганизмов. Причина высокого содержания
корриноидов у метаногенных бактерий не установлена.
Получение витамина В12 с помощью
бактерии Pseudomonas denitrificans
Ряд штаммов рода Pseudomonas образует в значительных
количествах B12, но чаще всего используют мутант Ps.
denitriflcans, у которого в результате мутагенеза уровень витамина В12
удалось поднять с 0,6 мг/л (дикий штамм) до 60 мг/л. Бактерии культивируют с
аэрацией и перемешиванием в периодических (или проточных) условиях в среде (а)
следующего состава: свеклосахарная меласса - 100 г, дрожжевой экстракт - 2 г,
(NHtb НРО4 - 5 г, MgS04 - 3 г, MnS04 - 200 мг, CoNO3 - 188 мг, 5,6 ДМБ - 25 мг,
ZnSO4 - 20 мг, Na2MoO3 - 5 мг, вода водопроводная - до 1 л, рН 7,4. Меласса
богата бетаином и глутаминовой кислотой, оказывающими положительный эффект на
выход витамина. Бетаин стимулирует синтез б-АЛК и, возможно, также изменяет
проницаемость мембраны. Культуру сохраняют в лиофилизированном состоянии,
поддерживают в вышеприведенной среде. Пересев осуществляют в пробирку с плотной
средой (б). Состав среды (б): свеклосахарная меласса - 60 г, пивные дрожжи - 1
г, N-Z-амин - 1 г, (МН4ЬНРО4 - 2 г, MgSO4 - 1 г, MnSO4 - 200 мг, ZnS04 - 20 мг,
MoSO4 - 5 мг, агар - 25 г, вода водопроводная - до 1 л, рН 7,4. Инкубируют 4
дня при 28°. Далее клетки переносят в 150 мл жидкой среды того же состава (но
без агара), налитую в литровую колбу Эрленмейера. Инкубируют 3 дня при 28° на
качалке. Содержимое колбы вносят в ферментер на 5 л, содержащий 3,3 л среды
(см. выше), стерилизованный 75 мин при 120°. Инкубируют 90 ч при 29° с
перемешиванием (420 об/мин) и аэрацией (0,2 м3/ч). Чистый витамин В12
получают в результате проведения следующих последовательных операций:
Культуральная жидкость с клетками Ps. denitrificans (3,3л)
нагревание 30 мин при 20°С, охлаждение, доведение Рн до 8,5, добавление KCN,
перемешивание 16 ч при 25°, добавление ZnCl2 (200 г), доведение рН до 8,0,
перемешивание, фильтрация.
Фильтрат:
· Трехкратная экстракция 350 мл смесей
· Органический экстракт I
· Водный экстракт
· Трёхкратная экстракция 30 мл. смеси
крезола и
· углеродистого тетрахлорида (1: 2
соотношения)
· Органический экстракт II
· Добавление 200 мл ацетона и 120 мл эфира
· Неочищенный витамин В12
В результате процесса экстракции получают цианкобаламин 98%-й
чистоты и выходом 75%. Конечный выход - 59-60 мг/л, CN-кобаламин - стабильная
форма витамина.
Получение витамина В12 с помощью штамма бактерий Pseudomonas
fluorescens BKM В-2224Д
Изобретение относится к микробиологической промышленности и
касается нового штамма бактерий Pseudomonas fluorescens BKM
В-2224Д, продуцирующего витамин В12, который применяется как
антианемический препарат в медицине и в производстве кормовых добавок для
сельскохозяйственных животных и птицы.
Целью изобретения является получение штамма бактерий,
способных синтезировать витамин В12 в более высокой концентрации,
чем известные микробные продуценты.
Условия биосинтеза витамина В12 штаммом Pseudomonas
fluorescens BKM В-2224Д.
Биосинтез витамина В12 культурой Pseudomonas
fluorescens BKM В-2224Д осуществляется по следующей схеме: посевная
культура на агаризованной среде - -> посевная культура на колбах на жидкой
среде - -> культура на жидкой среде в посевном аппарате - -> биосинтез на
жидкой среде в ферментере.
В качалочную колбу объемом 750 мл с объемом среды 50 мл
вносят культуру из пробирки в количестве 0,1%.
Выращивают культуру при температуре 30oС в течение
48 часов.
Условия ферментации: постоянное перемешивание, температура
28-32oС, при подаче воздуха 02-05 л/л среды в минуту в течение 40-60
часов до оптической плотности больше 0,2.
Полученным посевным материалом в дозе засевают стерильную
ферментационную среду.
Ферментацию проводят при температуре культуральной жидкости
28-32oС, при постоянном перемешивании, избыточном давлении 0,5 ати,
при расходе воздуха 0,2 л/л среды в минуту.
Культура Pseudomonas fluorescens BKM - 2224Д характеризуется
тем, что ее рост происходит параллельно с биосинтезом витамина В12 в
аэробных условиях на протяжении всего процесса ферментации.
Содержание витамина В12в культуральной жидкости к
концу ферментации составляет 120-150 мкг/мл при определении микробиологическим
методом с использованием в качестве тест-культуры Е. соli 113-3 или
спектрофотометрическим методом. При культивировании пропионово-кислых бактерий
содержание витамина B12 в культуральной жидкости равно 40-50 мкг/мл.
По окончании процесса ферментации культуральную жидкость
подкисляют серной кислотой до значения рН 5,8 ед., проводят лизис культуры при
температуре 90 - 10oС. Для получения готового продукта культуральная
жидкость может быть переработана на распылительной сушилке с наполнителями
(мел, соль, отруби и др.) или отправлена на стадии получения и выделения
витамина B12 в чистом виде.
Получение витамина B12 с помощью
метаногенных бактерий
В клетках метанобразующих бактерий витамин В12
присутствует от 4,1 нмоля/мг сухих клеток у Methanosarcina barkeri до
0,65 наномолей/мг сухих клеток у Metanobacterium formicum. Биосинтез
кобаламинов архебактериями (изучали на М. barkei) сходен с биосинтезом
корриноидов у анаэробных эубактерий. У метанотрофа Mtb. thermoautotrophicum большая
часть клеточного кобамида локализована во фракции мембран и связана с
мембранным белком. Предполагают, что содержащий кобамид интегральный мембранный
белковый комплекс играет существенную роль в метаболизме этих бактерий при
утилизации H2 + CO2, которая, видимо, сводится к переносу электронов. Корриноиды
у метанобразующих бактерий участвуют также в катаболизме ацетата и метанола.
Превращение метанола в метан у Mis. barkeri происходит через образование
СНз-СоМ, в метилировании которого за счет метанола участвуют две
метилтрансферазы, зависимые от кобамида. Корриноид, видимо, служит
простетической группой фермента. Во Франции из ила сточных вод выделили
мезофильные метаногенные бактерии и инкубировали их в ассоциации с другими
бактериями в полупроточном режиме в среде, содержащей метанол (3-12 г/л),
мелассу, кукурузный экстракт, NH4, Co, ортоксилидин и 5,6 ДМБ. Ферментацию
ведут при 35° в ферментере на 1000 м3 с ежесуточной заменой 10% бродящего
субстрата свежей средой. Биомассу отделяют на сепараторах и высушивают на
распылительной сушилке. Высушенный концентрат усредняют мелом до стандартной
активности 1000 мкг/г препарата, который используют в таком виде как добавку к
кормам. Сухой концентрат до усреднения содержит - 3000 мкг/г витамина B12 что
составляет 45-50% суммы корриноидов, фактор III - 10-15% и другие неполные
корриноиды - 40-50%. Термофильные штаммы метанотрофных бактерий родов Methanobacillus и Methanobacterium образуют 2 мг/л
кобаламина при содержании в среде 8 г/л метанола. В России производство
кормового препарата витамина B12 основано в основном на переработке барды
(отхода ацетонобутилового или спиртового производства) биоценозом бактерий, осуществляющих
термофильное метановое брожение сточных вод. Используют сложный консорциум
анаэробных микроорганизмов, включающих углеводсбраживающие, аммонифицирующие,
сульфатвосстанавливающие и метанобразующие бактерии. К, барде добавляют метанол
- до 2%, СоСl2-6Н2О - 10 г/м3, мочевину - 300 г/м3 и сухие кормовые дрожжи -
230 г/м3. Дозировку обогатителей производят автоматически. Барду подают в
нижнюю часть ферментатора метантенка (па 4-5 тыс. м3), в котором автоматически
регулируют параметры процесса, обеспечивая контроль температуры (55-57°С), рН
(7,5-8,0) и длительности брожения. Брожение ведут непрерывно, ежесуточно
заменяя 20-25% бродящей жидкости на свежую барду. В качестве пеногасителя
используют рыбий жир. Для получения кормового препарата бражку выпаривают и
сушат. Поскольку витамин В12 неустойчив при тепловой обработке, особенно в
щелочной среде, его стабилизируют. Для этого получаемую в процессе брожения
жидкость перед выпариванием подкисляют до рН 5,0-5,3 и добавляют к ней сульфит
натрия (0,1-0,25%). Содержание витамина В12 в исходной сброженной жидкости -
4,4 г/м3. Сгущение сброженной барды осуществляют на выпарных аппаратах (до
14-17% содержания сухих веществ), а сушку в распылительной сушилке.
Концентрация витамина B12 в высушенном препарате - 500-600 мг/кг. Истинный
витамин составляет 20-25% от суммы корриноидов, фактор III - 35-40%, фактор В и
другие - 40-45%. Получаемый препарат называют КМБ-12.
Схема представлена на рисунке 5. Ацетоно-бутиловая барда из
нижней части бражной колонны поступает в сборник барды 1 и насосом подается в
декантатор 3. Отстой барды собирается в сборнике 4 и используется на корм
скоту. Декантат, охлажденный до температуры 55-57°С, метанол и хлористый
кобальт поступают в ферментер 12. Сброженную массу из верхней части ферментера
отбирают и направляют в реактор 19, где осуществляют стабилизацию витамина B12 путем добавления
сульфита натрия и соляной кислоты, смешанных в смесителе 18. Из
стабилизированной бражки удаляют газы в сепараторе газов 22, бражку упаривают в
выпарной установке 24 и собирают в сборниках 26. Сгущенная метановая бражка
перекачивается насосом 27 в сборник метановой бражки 28, а оттуда насосом 29 в
распылительную сушилку 31.
В качестве теплоносителя для сушки используют газы брожения,
сжигаемые в печи 39. Сухой порошок поступает в бункер 33 и расфасовывается в
полиэтиленовые мешки, вложенные в крафт-пакеты.
Отсутствие промышленных отходов, доступность сырья,
непрерывность метода, не требующего стерильных условий, делают его экономичным.
Кормовой витамин В12 получают в Советском Союзе на
Грозненском ацетоновом и Ефремовском биохимическом заводах. Для производства
витамина В12 предусмотрены и за последнее время внедрены системы
автоматизации основных стадий технологического процесса.
Сухой концентрат КМБ-12, помимо витамина В12 (100
мг/кг препарата), содержит ряд других ростстимулирующих веществ. Особенно
хорошие результаты в животноводстве получают при сочетании витамина B12 с малыми дозами
антибиотиков, в частности с биомицином. Реализация в нашей стране описанного
способа и получение концентрата витамина В12 позволили полностью
обеспечить животноводство этим витамином.
В США почти все вырабатываемые комбикорма для свиней и птиц
обогащают витамином В12. Показано, что белок животного происхождения
можно заменить растительным белком при условии обогащения кормосмесей витаминов
В12 в дозе 60 мкг/кг.
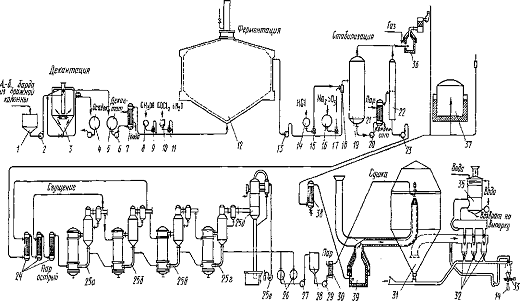
Рисунок 5 - Технологическая схема получения концентрата витамина
В12 с помощью смешанной культуры метанообразующих бактерий: 1 - сборник барды; 2 - насос для барды;
3 - декантатор барды; 4 - сборник сгущенной барды; 5 - сборник декантата барды;
6 - насос для декантата барды; 7 - холодильник для охлаждения декантата барды,
8 - сборник-мерннк метанола; 9 - насос-дозатор метанола; 10 - сборннк-мерннк
раствора хлористого кобальта; 11 - насос-дозатор раствора хлористого кобальта;
12 - ферментатор для метанового брожения; 13 - насос для метановой бражки; 14 -
сборник-мерннк соляной кислоты; 15 - насос-дозатор соляной кислоты; 16 - сборник-мерник
раствора сульфита натрия; 17 - насос-дозатор раствора сульфита натрия; 18 -
смеситель метановой бражки, соляной кислоты и раствора сульфита натрия, 19 -
реактор для стабилизации витамина Bi2 в
метановой бражке; 20 - насос для стабилизированной метановой бражки; 21 -
подогреватель стабилизированной метановой бражки; 22 - сепаратор газов,
выделяющихся из метановой бражки; 23 - насос для подачи стабилизированной
метановой бражки на выпарную установку; 24 - подогреватели метановой бражки; 25
- выпарная установка для сгущения метановой - бражки (а-1 корпус, 6-II корпус, s - III корпус, г-IV корпус, д-барометрический конденсатор,
е - вакуум-насос); 26-сборник сгущенной метановой бражки; 27-насос для
сгущенной метановой бражки; 28 - сборник сгущенной метановой бражки
(передаточный); 29 - насос для сгущенной метановой бражки; 30 - подогреватель
сгущенной метановой бражки; 31 - распылительная сушилка с центробежным
распылением; 32 - циклоны распылительной сушилки; 33 - бункер сухого
концентрата; 34 - расфасовка в мешкн; 35 - скруббер для очистки дымовых газов
сушилки от порошка концентрата; 36 - установка для каталитического сжигания
газов, выделяющихся при подкисленни и нагревании метановой бражки; 37 -
газгольдер для газов брожения; 38 - холодильник для отделения воды из газов
брожения; 39 - газовая печь распылитель ной сушилки.
Украинским научно-исследовательским институтом спиртовой и
ликеро-водочной промышленности разработана технология получения кормового
концентрата витамина В12 путем сбраживания мелассно-спиртовой барды
смешанной культурой метанообразующих бактерий. Предварительно на
мелассно-спиртовой барде выращивают кормовые дрожжи. После сепарирования
дрожжей получают культуральную жидкость, содержащую 7-8 % сухих веществ. На
этой жидкости выращивают метанообразующие бактерии и получают с 1 м1
исходной барды 1,5-2 г витамина В12.
Культуральную жидкость упаривают до содержания 60-70 % сухих
веществ, смешивают с наполнителем и высушивают. В качестве наполнителя
используют кукурузную муку, отруби и т.п. Сухой концентрат размалывают и
расфасовывают в мешки. В 1 кг кормового концентрата содержится: витамина B12 18 - 20 мг, В2 41 мг, РР 146 мг и другие
витамины. Срок хранения 12 месяцев.
Мезофильные и термофильные метаногенные бактерии, в том числе Metanobacterium thermoautotrophi-сит, Mb. thermoformicuin, Mb. bryantii, Metanosarcina barkeri, Ms. vacuolata,
Ms. mazei, Methanococcus hatopilus,
синтезируют исключительно
фактор III. Истинный витамин B12 образуют неспорообразующие
метилотрофы: Eubacterium limosum, близкий к нему Butyribacterium methylotrophicum и Acetobacter woodi. Путем
создания искусственных биоценозов и подбора условий ферментации возможно
целенаправленное регулирование процесса биосинтеза витамина В12.
Новые разработки. Для удешевления производства витамина B12 и
утилизации дешевого возобновляемого сырья изучалось образование корриноидов
бактериями Prop, atidipropionici ATCC 25562 при росте на ксилозе
как главной составной части гидролизатов гемицеллюлоз. Используя ксилозу,
бактерии аккумулировали 0,35 мг корриноидов в одном литре среды без внесения
солей Со. Для продукции корриноидов из ксилозы больше всего подходит
UFR-реактор, работающий с ультрафильтрационной рециркуляцией клеток.
Иммобилизованные клетки в Японии сразнивали стабильность и продуктивность
биокатализатора при включении клеток Propionibacterium sp. в гели каппа
- каррагенана, Na-альгината, агара и форполимерные уретановые смолы.
Оптимальной подложкой служит форполимер PU-9, полимерная матрица которой не
снижала активности включенных в нее клеток. В оптимальных условиях ферментации
5 г иммобилизованных клеток вновь синтезировали и экскретировали 900 мкг
витамина за 18 дней повторной периодической ферментации, продемонстрировав
возможность проведения многоступенчатого сложного синтеза (подобных примеров
известно немного). Усовершенствование штаммов-продуцентов. В последние годы
усовершенствование штаммов было достигнуто путем мутаций и селекции. Этим
методом в 50 раз увеличена продуктивность по витамину у Ps. denitrificans. Для
грамположительных бактерий Propionibac-terium, Bacillus, Streptotnyces
применимо слияние протопластов, для грамотрицательных бактерий, например, Pseudomonas,
доступны конъюгативные плазмиды. Пока весомых практических результатов
этими новыми и мощными методами не получено, но начало таким работам положено.
Клонировали 11 генов, кодирующих ферменты биосинтеза витамина B12 у
бактерии Вас. megaterium. Полагают, что в геноме содержится всего 20-30
таких генов. Поэтому DNA Вас. megaterium подвергли фрагментированию и
крупные фрагменты встраивали в плазмиды, которыми далее трансформировали
мутанты-ауксотрофы по В12. Такие мутанты приобрели способность
синтезировать витамин В12. Метод может быть использован для
получения штамов-продуцентов в производственных масштабах. В бактерии Е.
coll клонированы гены Prop, technicum, ответственные за синтез
витамина В12. Бактерия Prop. technicum не содержит плазмид,
поэтому из этого штамма выделили, очистили и частично разрушили ДНК, получив
фрагменты 15 - 20 килобаз. Эти фрагменты включили в расщепленную плазмиду pBR
322 и полученной гибридной плазмидой трансформировали Е. соli. Новые
трансформанты отличались от контрольного штамма в отношении морфологических и
физиологических признаков.
2.4
Применение витамина В12
Цианокобаламин обладает выраженным липотропным действием, он
предупреждает жировую инфильтрацию печени, повышает потребление кислорода
клетками при острой и хронической гипоксии. Витамин B12 участвует в
процессах трансметилирования, переноса водорода. Цианокобаламин принимает
непосредственное участие в процессе синтеза метионина. Он принимает участие в
синтезе пуриновых и пиримидиновых оснований, нуклеиновых кислот, необходимых
для процесса эритропоэза, активно влияет на накопление в эритроцитах
соединений, содержащих сульфгидрильные группы.
Ампулы с витамином B12 - это один из лекарственных
препаратов, применяющийся в традиционной медицине для лечения различных
гематологических заболеваний (рисунок 6). В частности, к ним относятся:
хронические анемии, сопровождающиеся дефицитом витамина B12
(алиментарная макроцитарная анемия, болезнь Аддисона-Бирмера); железодефицитная
анемия; апластическая анемия; геморрагическая анемия; анемии, вызванные
воздействием токсичных веществ или сильнодействующих лекарственных препаратов.
Цианокобаламин помогает нормализовать пониженное кровяное
давления. Стимулируя синтез протеина в человеческом организме, витамин В12
является своеобразным анаболиком. В самое последнее время получены данные, что
витамин B12 имеет важное значение и для образования костей. Рост
костей может происходить лишь в том случае, когда в остеобластах (клетках, из
которых создаются кости) имеется достаточный запас витамина. Витамин В12
увеличивает количество лейкоцитов и ускоряет их деятельность, то есть
воздействует на ретикулоэндотелиальную систему, благодаря данному препарату
повышается сопротивляемость организма всем возможным заболеваниям. Недостаток
витамина B12 значительно сокращает жизнь больных СПИДом. Витамин В12
стимулирует деятельность кровеносных органов и оказывает существенное влияние
на способность человека к восприятию и усвоению новой информации. Его
недостаток вызывает депрессивное состояние и рассредоточение внимания. Витамин
B12 является одним из веществ, необходимых для здоровья
репродуктивных органов. Очень важен для поддержания в хорошей форме женского и
мужского здоровья он способен корректировать снижение содержания сперматозоидов
в семенной жидкости.
Витамин B12 принимает непосредственное участие в
процессе кроветворения: именно он способствует нормальному созреванию
эритроцитов и увеличивает их толерантность к гемолизу. В результате применения
витамина B12 активируется свертывающая система крови и повышается
способность тканей организма к регенерации. Помимо этого, данный лекарственный
препарат оказывает благотворное воздействие на функцию нервной системы и
печени, а также понижает концентрацию холестерина в крови.
Кобаламин стимулирует деление клеток, от наличия его в
организме зависит благополучие тех тканей, которые наиболее подвержены
интенсивному делению: иммунные клетки, клетки крови и кожи, а также клетки,
составляющие верхнюю часть кишечника. Витамин B12 влияет на
миелиновую оболочку (покрытие нервов), и нехватка витамина вызывает необратимые
разрушения нервов.

Рисунок 6 - Формы отпуска витамина В12
из аптек
Заключение
В ходе анализа литературы было рассмотрено несколько
продуцентов витамина В 12. Наиболее активно продуцируют
представители вид Propioni bacterium, с помощью которого получают до 58 мг/л
витамина. Продуценты витамина B12 культивируют в средах, приготовленных на основе
пищевого сырья: соевой муки, рыбной муки, мясного и кукурузного экстракта. В
последние годы выявлены микроорганизмы, образующие высокие качества корриноидов
при утилизации непищевого сырья.
Одним из основных методов получения витамина В12
выбран был метод с помощью пропионовокислых бактерий, так как этот метод
представляет штаммы, способные к самостоятельному синтезу 5,6 ДМБ. Он наиболее
выгоден и даёт получить довольно большой выход продукта, например, в России
составил 58 мг/л витамина, а во Франции достигли невероятного высокого выхода -
216 мг/л. Изучены были недавно открыты новые аналоги витамина В12:
три из них получены при частичном синтезе (фтор-метилфосфито-Р-кобаламид,
диметилфосфито-Р-кобаламин, Fe-кобаламин) и один фактор VA выделенный из
активного ила.
Так же широкое применение получил метод получения витамина В12
с помощью бактерии Pseudomonas denitrificans. Ряд штаммов рода Pseudomonas
образует в значительных количествах витамин B12, но чаще всего
используют мутант Ps. denitriflcans, у которого в результате мутагенеза
уровень витамина В12 удалось поднять с 0,6 мг/л (дикий штамм) до 60
мг/л. Так же новый штамм бактерий Pseudomonas fluorescens BKM
В-2224Д, используют для получения штамма бактерий, способных
синтезировать витамин В 12 в более высокой концентрации, чем
известные микробные продуценты.
Фармацевтические заводы (Борисовский завод медицинских препаратов,
ОАО "Экзон", ОАО Несвижский завод медицинских препаратов, ПРУП
"Минскинтеркапс", Гродненский завод медицинских препаратов, СООО
"Лекфарм") покрывают потребность витамина В12,
отечественного производства по доступной для населения цене и удовлетворяют
потребности препарата для республики в целом.
Список
использованных источников
1. Березовский, В.М. Химия витаминов / B.М. Березовский. - М.: Мир, 1973. -
587 с.
2. Бирюков,
В.В. Оптимизация периодических процессов микробиологического синтеза / В.В. Бирюков,
В.М. Кантере. - М., 1985. - 375 с.
3. Букин,
В.Н. Микробиологический
синтез витаминов / В.Н. Букин. - М., 1972. - 296 с.
4. Букин,
В.Н. Биохимические и
микробиологические основы промышленного получения витамина B12 методом термофильного метанового
брожения. Сб. Витамин В12 и его применение в животноводстве / В.Н.
Букин. - М., 1971. - 376 с.
. Быков,
В.А. Производство белковых веществ / Биотехнология, под ред.Н.С. Егорова и В.Д.
Самуилова; т. 5. - М., 1987. 307с.
. Быховский,
В.Я. Микробиологический синтез витамина B12/В.Я. Быховский. - М., 1984. - Сер.
V.
. Быховский,
В.Я. Промышленное получение витамина B12 методом метанового брожения / В.Я.
Быховский. - М.: Пущино, 1983. - 209 с.
. Викторов,
П.И. Опыт применения продуктов микробиологического синтеза в кормопроизводстве
/ П.И. Викторов. - М., 1979. - 456 с.
. Воробьева,
Л.И. Микробиологический синтез витаминов / Л.И. Воробьева. - М., 1982. - 267 с.
10. Воробьева,
Л.И. Пропионовокислые
бактерии и образование витамина B
12/Л.И. Воробьева. - М., 1976. - 315 с.
11. Гальцова,
Г.Д. Стеринообразование
у дрожжевых организмов / Г.Д. Гальцова. - М., 1980. - 356 с.
. Гусев,
М.В. Микробиология / М.В. Гусев, Л.А. Минеева. - М.: Издат. Центр
"Академия", 2003. - 464 с.
. Голубев,
В.Н. Пищевая биотехнология / В.Н. Голубев. - М.: ДеЛи принт, 2001. - 123 с.
. Деймен,
А. Н Промышленная микробиология / А.Н. Деймен. - М., 1984. - 216 с.
15. Диканская,
Э.М. Биосинтез
флавииов микроорганизмами / Э.М. Диканская. - М., 1972.Т. 3.
. Емцев,
В.Т. Микробиология / В.Т. Емцев, Е.Н. Мишустин. - М.: Дрофа, 2005. - 445 с.
. Елинов,
Н.П. Основы биотехнологии / Н.П. Елинов. - М.: Издательская фирма
"Наука" СПБ, 1995. - 319 с.
. Лысак,
В.В. Микробиология: учебное пособие / В.В. Лысак. - Минск: БГУ, 2007.
. Мосичев,
М.С. Общая технология микробиологических производств / М.С. Мосичев. - М.,
1982. - 256 с.
. Никтин,
Г.А. Биохимические основы микробиологических производств: Учеб. пособие / Г.А.
Никтин. - Киев: Вища школа. Головное изд-во, 1981. - 312 с.
. Письменный,
В.В. Автоматизация процессов производства кормового концентрата витамина
B12/В.В. Письменный. - М., 1985. Сер. IX.
. Поморцсва,
Н.В. Перспективы получения витаминов и коферментов с помощью микроорганизмов /
Н.В. Поморцсва. - М., 1986. № 8. - 968 - 987 с.
23. Робышева,
3.Н. Эргостерин
дрожжей / З.Н. Робышева. - М., 1972. - Т. 3.
. Сартакова,
О.Ю. Промышленная микробиология: учебное пособие по курсу "Основы
микробиологии и биотехнологии" / О.Ю. Сартакова. - Барнаул: Изд. АлтГТУ,
2009. - 239 с.
25. Шавловский,
Г.М. Механизм
регуляции биосинтеза в микроорганизме/Г.М. Шавловский. - М., 1976. № 5. - 34с.
27. Jeter, R., Е s с а
1 а п t е - S e m е г е n a J.С. et al. Synthesis and use of vitamin B12
// Escherichia coli and Salmonell typhimurium / R. Jeter. 1987. Vol. 1. P. -
551-556 c.
28. .
Demaiti A. Z. Riboflavin oversynthesis. Ann. Rev. Microbiol / A. Z. Demaiti. 1972. -
390 c.
29. Vitamin-portal.ru [Электронный ресурс] / Витамин В12
(кобаламин, антианемический витамин). Режим доступа:
<http://www.vitamin-portal.ru>. - Дата доступа 15. 05.2015
30. Studopedia.ru [Электронный ресурс] / Значение витамина
В12 в организме человека. Режим доступа: http://studopedia.ru. -
Дата доступа: 15. 05.2015.
31. Findpatent.ru [Электронный ресурс] / Новый штамм бактерий Pseudomonas fluorescens BKM В-2224Д. Режим доступа: http://www.findpatent.ru/patent/218/2180001.html.
- <http://www.findpatent.ru/patent/218/2180001.html.%20-> Дата доступа: 15. 05.2015.