Функции белков в организме. Стероидные гормоны. Липиды
КОНТРОЛЬНАЯ
РАБОТА
по
дисциплине «Биохимия»
г.
План
Строение,
состав и физиологическая роль отдельных органелл клетки
Напишите
структурные формулы дипептидов карнозин (бета-аланилгистидин) и ансерин (бета-
аланил-1- метилгистидин), опишите их функции в организме
Функции
белков в организме
Классификация
белков по степени сложности. Липопротеиды. Состав, строение, биологическая роль
Напишите
последовательность оснований в м-РНК, образованной на цепи ДНК с
последовательностью ГТГ-ГЦТ-ГЦА-ТАЦ-ЦЦГ-АГЦ-ГТГ-ГАТ-ААГ-ГТА-ГЦГ-ГЦЦ-ТЦГ-ТАГ-ТГТ-ЦТГ-ЦЦГ-АЦТ-ГЦА-ТЦА.
Установите последовательность аминокислот в пептиде
Привести
структурную формулу витамина С, указать его физико-химические характеристики
(агрегатное состояние, растворимость в воде и органических растворителях,
влияние температуры, кислотности среды и солнечного облучения на стабильность
витамина), пояснить, в какой кофермент оно включается в организме или не
включается, подчеркнуть биологическую роль витамина, а также его минимальную
норму потребления в сутки в составе продуктов питания, указать источник
витамина, признаки гипер - и авитаминоза
Привести
структурную формулу витамина В6 (пиридоксин), указать его физико-химические
характеристики (агрегатное состояние, растворимость в воде и органических
растворителях, влияние температуры, кислотности среды и солнечного облучения на
стабильность витамина), пояснить , в какой кофермент оно включается в организме
или не включается, подчеркнуть биологическую роль витамина, а также его минимальную
норму потребления в сутки в составе продуктов питания, указать источник
витамина, признаки гипер- и авитаминоза
Механизм
ферментативного катализа
Стероидные
гормоны. Мужские половые гормоны. Строение и биологическая роль гормонов
Вода
в живом организме. Состояние воды в живых тканях, ее функции. Эндогенная вода
Полисахариды
морских водорослей. Состав, строение. Функции в организме
Классификация
липидов. Сложные липиды. Фосфолипиды. Строение. Распространение. Биологическая
роль
Литература
гормон липид белок органелла
Строение, состав и физиологическая роль
отдельных органелл клетки
Основными компонентами клетки являются: наружная
клеточная мембрана, цитоплазма и ядро. Клеточная мембрана - это оболочка
клетки, выполняющая следующие функции:
разделение содержимого клетки и внешней среды;
регуляция обмена веществ между клеткой и средой;
место протекания некоторых
биохимических реакций (в том числе фотосинтеза
<#"870672.files/image001.gif">
Они увеличивают амплитуду мышечного
сокращения, предварительно сниженную утомлением. Имидазолсодержащие дипептиды
не влияют непосредственно на сократительный аппарат, но увеличивают
эффективность работы ионных насосов мышечной клетки.
Карнозин участвует в процессах
фосфорилирования, происходящего в мышцах при образовании фосфатных соединений.
Карнозин является естественным дипептидом, который находится в мышечных
волокнах обоих типов - 1 и 2 (медленные и быстрые) . Но в значительно более
высоких концентрациях он содержится в волокнах 2 типа. Именно этот тип является
наиболее чувствительным к мышечному росту.
Ансерин также связан с физиологической
функцией мышц: появление его в мышечной ткани совпадает по времени с
формированием их сократительной функции; мышцы, несущие большую функциональную
нагрузку, содержат больше ансерина.
Функции белков в организме
Белки занимают центральное место в
метаболизме организма человека, выполняя ряд важных функций:
) структурную или пластическую
(структурная основа клеток, органеллы, фибриллярные белки);
) транспортную (липопротеиды,
гемоглобин, альбумин);
) сократительную (мышечные белки -
актин, миозин);
) каталитическую (ферменты);
) регуляторную (гормоны);
) защитную (иммуноглобулины,
антитела, интерферон; белки системы свертывания крови и фибринолиза);
) энергетическую (распад аминокислот
обеспечивает до 18% потребляемой энергии).
Пример:
В плазме крови содержится 7% всех
белков организма при концентрации 60 - 80 г/л. Белки плазмы крови выполняют
множество функций. Альбумин, транстиретин, транскортин, трансферрин и некоторые
другие белки вьшолняют транспортную функцию.
|
руппа
|
Белки
|
Концентрация
в сыворотке крови, г/л
|
Функция
|
|
Альбумины
|
Транстиретин
|
0,25
|
Транспорт
тироксина и трийодтиронина
|
|
|
Альбумин
|
40
|
Поддержание
осмотического давления, транспорт жирных кислот, билирубина, жёлчных кислот,
стероидных гормонов, лекарств, неорганических ионов, резерв аминокислот
|
|
|
ЛПВП
|
0,35
|
Транспорт
холестерола
|
|
|
Транскортин
|
0,03
|
Транспорт
кортизола, кортикостерона, прогестерона
|
|
|
Кислый
α1-гликопротеин
|
1
|
Транспорт
прогестерона
|
|
|
Тироксинсвязывающий
глобулин
|
0,02
|
Транспорт
тироксина и трийодтиронина
|
|
α2-Глобулины
|
Церулоплазмин
|
0,35
|
Транспорт
ионов меди, оксидоредуктаза
|
|
|
1
|
Связывание
гемоглобина
|
|
|
α2-Макроглобулин
|
2,6
|
Ингибитор
плазменных протеиназ, транспорт цинка
|
|
|
Ретинолсвязывающий
белок
|
0,04
|
Транспорт
ретинола
|
|
|
Витамин
D связывающий белок
|
0,4
|
Транспорт
кальциферола
|
|
β-Глобулины
|
ЛПНП
|
3,5
|
Транспорт
холестерола
|
|
|
Трансферрин
|
3
|
Транспорт
ионов железа
|
|
|
Транскобаламин
|
25×10-9
|
Транспорт
витамина B12
|
|
|
Глобулин
связывающий белок
|
20×10-6
|
Транспорт
тестостерона и эстрадиола
|
Классификация белков по степени сложности.
Липопротеиды. Состав, строение, биологическая роль
Простые белки состоят только из аминокислот, а
сложные белки содержат, кроме аминокислот, другие небелковые вещества,,
например, углеводы, липиды, нуклеиновые кислоты и др.
Липопротеиды.
Этот класс сложных беков состоит из белка и
простетической группы, представленной каким-либо липидом. Белковая группа
гидрофильна, а простетическая гидрофобна, что и играет роль в организации
мембран и их избирательной проницаемости. В частности, в составе липопротеинов
открыты нейтральные жиры (триглицериды), свободные жирные кислоты, фосфолипиды,
холестериды. Липопротеины широко распространены в природе и выполняют
разнообразные биологические функции. Они входят в состав клеточной мембраны и
внутриклеточных биомембран ядра, митохондрий, микросом (структурированные
липопротеины), а также присутствуют в свободном состоянии (главным образом в
плазме крови). К липопротеинам относятся, кроме того, тромбопластический белок
ткани легких, липовителлин желтка куриного яйца, некоторые фосфолипиды молока и
т.д. Установлено, что липопротенины участвуют в структурной, комплексной
организации миелиновых оболочек, нервной ткани, фоторецепторной и
электронно-транспортной систем, палочек и колбочек сетчатки и др.
Большинство ЛП синтезируется в печени или в
слизистой оболочке кишечника. Они содержат гидрофобное липидное ядро,
окруженное полярными липидами и оболочкой из белков, получивших название
апобелки. Обычно ЛП содержат до 5% углеводов (глюкоза, галактоза, гексозамины,
сиаловая кислота), поэтому некоторые из них являются и гликопротеинами.
Триглицериды, холестерин, эфиры холестерина и
фосфолипиды транспортируются в форме водорастворимых липопротеидов.
Липопротеиды - это сферические частицы, состоящие из гидрофобной сердцевины и
гидрофильной оболочки. Сердцевина содержит неполярные липиды - триглицериды и
эфиры холестерина. Оболочка построена из полярных липидов - холестерина и
фосфолипидов, причем заряженные концы этих молекул обращены наружу. Кроме того,
в состав оболочки входят белки, нековалентно связанные с фосфолипидами и
холестерином, - апопротеины. Апопротеины поддерживают структуру липопротеидных
частиц и обеспечивают их взаимодействие с рецепторами липопротеидов.
Липопротеиды подразделяют на несколько классов в зависимости от их плотности и
подвижности при электрофорезе. Плотность липопротеидной частицы определяется
отношением апопротеины/липиды: чем больше это отношение, тем выше плотность. В
крови, взятой натощак, присутствуют только ЛПОНП, ЛПНП и ЛПВП, тогда как другие
липопротеиды (хиломикроны, остаточные компоненты хиломикронов, а также ЛППП)
выявляются только после еды или при нарушениях обмена липидов. ЛПОНП образуются
главным образом в печени. Это основная транспортная форма эндогенных триглицеридов,
синтезирующихся в печени. ЛПОНП - главный поставщик триглицеридов для скелетных
мышц и миокарда. В состав ЛПОНП входят апопротеины C, E и B100. По сравнению с
хиломикронами, ЛПОНП содержат меньше триглицеридов, но больше холестерина и
эфиров холестерина. Известно, что развитие атеросклероза ускоряется на фоне
повышенной концентрации ЛПОНП при сахарном диабете и болезнях почек.
ЛППП. В капиллярах скелетных мышц, миокарда и
жировой ткани ЛПОНП расщепляются липопротеидлипазой с образованием ЛППП. ЛППП
содержат апопротеины E и B100. В печени половина ЛППП превращается в ЛПНП под
действием триацилглицероллипазы, а оставшиеся ЛППП связываются с рецепторами
гепатоцитов, подвергаются эндоцитозу и разрушаются.
ЛПНП являются конечными продуктами расщепления
ЛПОНП и ЛППП. ЛПНП - это самые атерогенные липопротеиды. В составе ЛПНП
содержится примерно две трети всего холестерина плазмы, преимущественно в виде
эфиров холестерина. В отличие от других липопротеидов, ЛПНП содержат только
один апопротеин - B100. Избыток ЛПНП - один из главных факторов риска
атеросклероза.
ЛПВП синтезируются в печени и кишечнике и
содержат эфиры холестерина и фосфолипиды. Важнейшая функция ЛПВП - удаление
холестерина из тканей. Связывание ЛПВП с рецепторами вызывает выброс
холестерина из клеток. Первоначально холестерин встраивается в оболочку ЛПВП,
но затем под действием лецитинхолестерин-ацилтрансферазы он этерифицируется и
перемещается в сердцевину ЛПВП. В печени ЛПВП связываются с рецепторами и
разрушаются. Таким образом, ЛПВП - это антиатерогенные липопротеиды.
Напишите последовательность оснований в м-РНК,
образованной на цепи ДНК с последовательностью
ГТГ-ГЦТ-ГЦА-ТАЦ-ЦЦГ-АГЦ-ГТГ-ГАТ-ААГ-ГТА-ГЦГ-ГЦЦ-ТЦГ-ТАГ-ТГТ-ЦТГ-ЦЦГ-АЦТ-ГЦА-ТЦА.
Установите последовательность аминокислот в пептиде
Генетический код - система записи генетической
информации в ДНК в виде определенной последовательности нуклеотидов.
Последовательность нуклеотидов определяет последовательность включения АК в
синтезируемый белок. 3 нуклеотида - триплет - кодон - кодируют 1 АК.
Совокупность триплетов и составляет генетический код.
в ДНК содержится тимин, а в РНК - урацил.
В качестве азотистых оснований в РНК
присутствуют урацил, цитозин, аденин и тимин
Заменяем цитозин на гуанин, гуанин на цитозин,
тимин на аденин, аденин на урацил
ГТГ-ГЦТ-ГЦА-ТАЦ-ЦЦГ-АГЦ-ГТГ-ГАТ-ААГ-ГТА-ГЦГ-ГЦЦ-ТЦГ-ТАГ-ТГТ-ЦТГ-ЦЦГ-АЦТ-ГЦА-ТЦА.
М-РНК
ЦАЦ-ЦГА-ЦГУ-АУГ-ГГЦ-УЦГ-ЦАЦ-ЦУА-УУЦ-ЦАУ-ЦГЦ-ЦГГ-АГЦ-АУЦ-АЦА-ГАЦ-ГГЦ-УГА-ЦГУ-АГУ.
Пептид:
ГИС-ТРЕГ-
АРГ-МЕТ-ГЛИ-СЕР-ГИС-ЛЕЙ-ФЕН-ГИС-АРГ-АРГ-СЕР-ИЛЕЙ-ТРЕ-АСП-ГЛИ-СТОП-АРГ-СЕР
Привести структурную формулу витамина С, указать
его физико-химические характеристики (агрегатное состояние, растворимость в
воде и органических растворителях, влияние температуры, кислотности среды и
солнечного облучения на стабильность витамина), пояснить, в какой кофермент оно
включается в организме или не включается, подчеркнуть биологическую роль
витамина, а также его минимальную норму потребления в сутки в составе продуктов
питания, указать источник витамина, признаки гипер - и авитаминоза
По физическим свойствам аскорбиновая кислота
представляет собой белый кристаллический порошок кислого вкуса. Легко растворим
в воде, растворим в спирте. Легко тразлагается при нагревании, в кислой среде
более устойчива. Аскорбиновая кислота или витамин С получила название
антискорбутного, антицинготного фактора, предохраняющего от развития цинги -
болезни, принимавшей в средние века характер эпидемии. По химической структуре
аскорбиновая кислота представляет собой лактон кислоты со структурой, близкой структуре
L-глюкозы. Аскорбиновая кислота относится к сильным кислотам; кислый характер
ее обусловлен наличием двух обратимо диссоциирующих енольных гидроксилов у 2-го
и 3-го углеродных атомов.
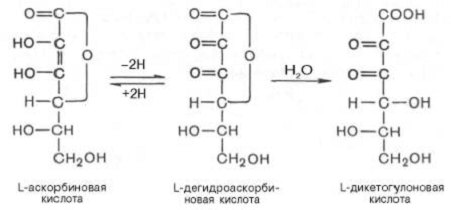
Из представленных структурных формул
видно, что наиболее важным химическим свойством аскорбиновой кислоты
<#"870672.files/image003.gif">
Все формы витамина В6 используются в организме
для синтеза коферментов: пиридоксальфосфата и пиридоксаминфосфата. Коферменты
образуются путём фосфорилирования по гидроксиметильной группе в пятом положении
пиримидинового кольца при участии фермента пиридоксалькиназы и АТФ как
источника фосфата.

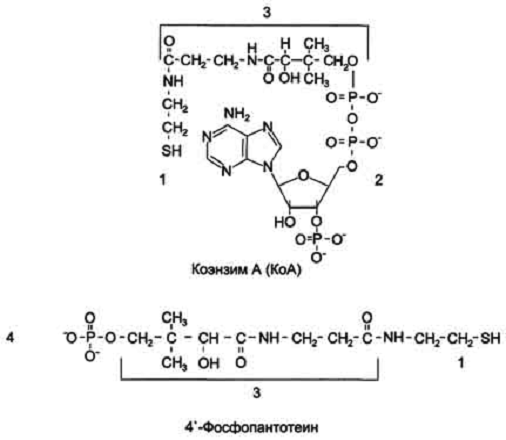
Строение КоА и 4'-фосфопантотеина. 1 -
тиоэтаноламин; 2 - аденозил-3'-фосфо-5'-дифосфат; 3 - пантотеновая кислота; 4 -
4'-фосфопантотеин (фосфорилированная пантотеновая кислота, соединённая с
тиоэтаноламином).
Все 3 формы витамина - бесцветные кристаллы,
хорошо растворимые в воде. Пиридоксин довольно устойчив даже при нагревании в
присутствии кислот и щелочей, он хорошо растворим в этиловом спирте и воде.
Однако при воздействии солнечных лучей витамин В6 быстро разрушается. Все формы
витамина В6 (пиридоксин, пиридоксаль и пиридоксамин) реагируют как основания с
кислотами, образуя соли. Он обладает горьковато-кислым вкусом.
Пиридоксалевые ферменты играют ключевую роль в
обмене аминокислот: катализируют реакции трансаминирования и
декарбоксилирования аминокислот, участвуют в специфических реакциях метаболизма
отдельных аминокислот: серина, треонина, триптофана, серосодержащих
аминокислот, а также в синтезе гема. Пиридоксальфермент катализирует реакцию
переаминирования. В результате серии реакций, включающих в себя образование
фермент-субстратных комплексов с гистидинаминотрансферазой, гистидин переходит
в оксогистидин, а глиоксилевая килотая - в глицин. Над стрелкой внизу -
схематически группа лизина от ферментного белка
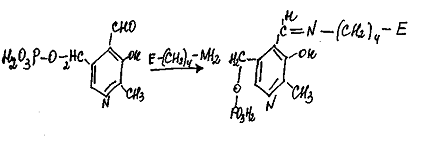
Взаимодействие между субстратом
<#"870672.files/image007.gif">
Пиридоксаминфосфат на второй стадии
реакции реагирует с α-кетокислотой
- глиоксилевой, что через те же стадии образования промежуточных соединений
(идущих в обратном направлении) приводит к синтезу новой аминокислоты
<#"870672.files/image008.gif">
Авитаминоз В6 у детей проявляется
повышенной возбудимостью ЦНС, периодическими судорогами, что связано, возможно,
с недостаточным образованием тормозного медиатора ГАМК, специфическими
дерматитами. У взрослых признаки гиповитаминоза В6 наблюдают при длительном
лечении туберкулёза изониазидом (антагонист витамина В6). При этом возникают
поражения нервной системы (полиневриты), дерматиты.
Источники витамина В6 для человека -
такие продукты питания, как яйца, печень, молоко, зеленый перец, морковь,
пшеница, дрожжи. Некоторое количество витамина синтезируется кишечной флорой.
Содержание пиридоксина в 100 граммах
продукта составляет: в печени - 0,9 мг, в постном мясе (говядина, телятина) -
примерно 0,3 мг, в мясе птицы - 0,17 мг, в рыбе - не менее 0,39 мг, в соевых
бобах - около 0,86 мг, в грецких орехах - не менее 0,68 мг, в пророщенных
зёрнах пшеницы - 0,72 мг, в бананах - не менее 0,34 мг, в авокадо - 0,22 мг, в
шпинате - 0,25 мг, в отрубях - 0,17 мг.
Суточная потребность составляет 2-3
мг.
Микроорганизмы кишечника жвачных
животных активно синтезируют его; эти животные не нуждаются во введении
витамина В6 с пищей. Микрофлора кишечника человека также синтезирует витамин
В6, но в недостаточных количествах, поэтому для человека и большинства животных
основным источником витамина являются продукты питания.
Механизм ферментативного катализа
В процессе катализа реакции в
контакт с субстратом вступает не вся молекула фермента, а определенный ее
участок, который называется активным центром. Эта зона молекулы не состоит из
последовательности аминокислот, а формируется при скручивании белковой молекулы
в третичную структуру. Отдельные участки аминокислот сближаются между собой,
образуя определенную конфигурацию активного центра. Важная особенность строения
активного центра - его поверхность комплементарна поверхности субстрата, т.е.
остатки АК этой зоны фермента способны вступать в химическое взаимодействие с
определенными группами субстрата. Можно представить, что активный центр
фермента совпадает со структурой субстрата как ключ и замок.
В процессе ферментативной реакции
различают несколько стадий: присоединение молекулы субстрата к ферменту,
преобразование первичного промежуточного соединения в один или несколько
последовательных (переходных) комплексов и протекающее в одну или несколько
стадий отделение конечных продуктов реакции от фермента.
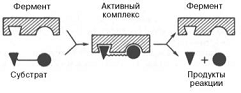
Образование нестойкого
фермент-субстратного комплекса согласно теории Э. Фишера «ключ-замок».

Участок фермента, содержащий такие
функциональные группы, называется активным центром. В активном центре условно
различают так называемый каталитический участок, непосредственно отвечающий за
превращения субстратной молекулы, и контактную площадку, которая обеспечивает
специфическое сродство к субстрату и формирование его комплекса с ферментом -
сорбционный участок. Ферменты, связываясь с субстратом, образуют промежуточный
фермент-субстратный комплекс, который затем распадается на свободный фермент и
продукт реакции.
Под изоферментами понимают ферменты,
катализирующие одну и ту же реакцию, но различающиеся по физическим, химическим
и иммунохимическим свойствам (ранее описаны изоферменты ЛДГ).
Мультиферментные комплексы
Скорость реакций можно повысить, не
увеличивая концентрации субстратов, если собрать различные участвующие в
последовательных реакциях ферменты в большой мультиферментный комплекс. При
таком способе организации продукт фермента А переходит непосредственно к
ферменту Б и т. д. до конечного продукта, причем лимитирующая стадия диффузии
отсутствует даже при очень низких внутриклеточных концентрациях промежуточных
соединений.
Мультиферментный комплекс оксидаз
митохондрийсостоит из 3-х компонентов.
Флавиновые дегидрогеназы. Содержат
ФАД. Наиболее обычный субстрат для них - НАДФН2.
Железо-серный белок. Содержит
негеминовое железо с переменной валентностью.
Цитохром Р450. Его строение
отличается от строения цитохромов цепи митоходриального окисления.
Мультиферментный комплекс формирует
цепь переноса электронов и протонов, в конце ее происходит активация кислорода.
Активированный кислород присоединяется к активному центру цитохрома Р450, и на
него переносятся электроны, а затем этот кислород включается в молекулу
субстрата
Ферменты являются высокоспецифичными
катализаторами; это их уникальное свойство. Ферменты проявляют специфичность в
отношении типа катализируемой химической реакции. Каждый фермент катализирует
определенную реакцию. Ферменты проявляют отчетливо выраженную субстратную
специфичность; некоторые ферменты действуют только на один субстрат, другие -
на группу родственных субстратов.
Механизмы обеспечения высокой специфичности
ферментов: комплементарность, баланс между прочностью и лабильностью структуры
активного центра, индуцированное связывание, многоточечность связывания
субстрата в активном центре, повышение специфичности по принципу «двойного
сита» в двух-субстратных реакциях.
При действии на субстраты,
содержащие асимметричные центры, ферменты, как правило, проявляют
стереоспецифичность. Она оказывается абсолютной в тех случаях, когда в
ферментативную реакцию вовлекается асимметрический центр. Так, например, L-лактатдегидрогеназа
действует только на L-лактат, в то время как D-лактатдегидрогеназа - только на
D-лактат. В тех случаях, когда субстрат не содержит, а продукт содержит
асимметрический атом углерода, происходит асимметрический синтез и при
восстановлении пирувата L-лактатдегидрогеназой образуется только L-лактат.
Акт катализа складывается из трех
последовательных этапов.
. Образование фермент-субстратного
комплекса при взаимодействии через активный центр.
. Связывание субстрата происходит в
нескольких точках активного центра, что приводит к изменению структуры
субстрата, его деформации за счет изменения энергии связей в молекуле. Это
вторая стадия и называется она активацией субстрата. При этом происходит
определенная химическая модификация субстрата и превращение его в новый продукт
или продукты.
. В результате такого превращения
новое вещество (продукт) утрачивает способность удерживаться в активном центре
фермента и фермент-субстратный, вернее уже фермент-продуктный комплекс
диссоциирует (распадается).
Ферментативные эффекторы - вещества,
изменяющие скорость ферментативного катализа и регулирующие за счет этого
метаболизм. Среди них различают ингибиторы - замедляющие скорость реакции и
активаторы - ускоряющие ферментативную реакцию.
В зависимости от механизма
торможения реакции различают конкурентные и неконкурентные ингибиторы. Строение
молекулы конкурентного ингибитора сходно со структурой субстрата и совпадает с
поверхностью активного центра как ключ с замком (или почти совпадает). Степень
этого сходства может даже быть выше чем с субстратом.Концентрация способного к
катализу фермента при этом снижается и скорость образование продуктов реакции
резко падает .В качестве конкурентных ингибиторов выступает большое число
химических веществ эндогенного и экзогенного происхождения (т.е. образующихся в
организме и поступающих извне - ксенобиотики, соответственно). Эндогенные
вещества являются регуляторами метаболизма и называются антиметаболитами.
Второй вид ингибиторов - неконкурентные. Они взаимодействую с ферментом вне
активного центра и избыток субстрата не влияет на их ингибирующую способность,
как в случае с конкурентными ингибиторами. Эти ингибиторы взаимодействуют или с
определенными группами фермента (тяжелые металлы связываются с тиоловыми
группами Цис) или чаще всего регуляторным центром, что снижает связывающую
способность активного центра. Необратимые ингибиторы инактивируют фермент,
образуя с его АК или другими компонентами структуры химическую связь. Обычно
это ковалентная связь с одним из участков активного центра. Такой комплекс
практически недиссоциирует в физиологических условиях. В другом случае
ингибитор нарушает конформационную структуру молекулы фермента - вызывает его
денатурацию. Действие обратимых ингибиторов может быть снято при переизбытке
субстрата или под действием веществ, изменяющих химическую структуру
ингибитора. Конкурентные и неконкурентные ингибиторы относятся в большинстве
случаев к обратимым.
Помимо ингибиторов известны еще
активаторы ферментативного катализа. Они:
защищают молекулу фермента от
инактивирующих воздействий,
образуют с субстратом комплекс,
который более активно связывается с активным центром Ф,
взаимодействуя с ферментом, имеющим
четвертичную структуру, разъединяют его субъединицы и тем самым открывают
доступ субстрату к активному центру.
Стероидные гормоны. Мужские половые
гормоны. Строение и биологическая роль гормонов
Стероиды - производные
восстановленных конденсированных циклических систем -
циклопентанпергидрофенантренов. В организме человека основной стероид -
холестерол, остальные стероиды - его производные. Мужские половые гормоны
вырабатываются в основном в мужских половых железах - в интерстициальных
клетках Лейдига семенников (95%). Небольшое количество андрогенов образуется в
коре надпочечников.
Путь биосинтеза андрогенов в яичках
и коре надпочечников одинаков. Предшественником андрогенов, как и других
стероидных гормонов, служит холестерол, который либо поступает из плазмы в
составе ЛПНП, либо синтезируется в самих железах из ацетил-КоА. Отщепление боковой
цепи холестерола и образование прегненолона - скорость-лимитирующая реакция.
Однако, в отличие от аналогичной реакции, протекающей в надпочечниках, эта
стадия стимулируется ЛГ (а не АКТГ). ЛГ, связываясь с рецептором плазматической
мембраны клеток Лейдига, активирует аденилатциклазу, увеличивая тем самым
внутриклеточную концентрацию цАМФ, что в конечном итоге вызывает активацию
фермента, который расщепляет боковую цепь холестерола между С-20 и С-22.

Мужские половые гормоны.
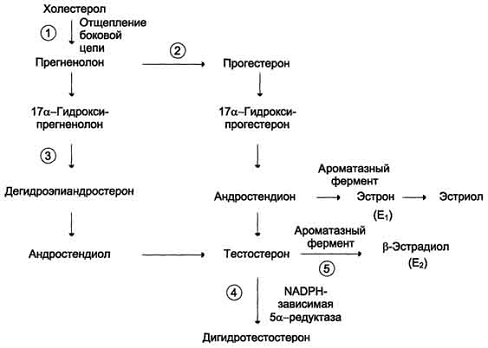
Схема синтеза половых гормонов
Суточная секреция тестостерона у
мужчин составляет в норме примерно 5 мг и сохраняется на протяжении всей жизни
организма. Гормон циркулирует в крови в связанном с белками плазмы состоянии:
альбумином (40%) и специфически связывающим половые гормоны глобулином
(называемым секс-гормонсвязывающим глобулином, СГСГ). Лишь 2% от общего
количества гормона в крови транспортируется в свободном виде, и именно такие
молекулы проявляют биологическую активность.
Дигидротестостерон. В семенных
канальцах, предстательной железе, коже, наружных половых органах тестостерон
служит предшественником более активного адцрогена - дигидротестостерона. Это
превращение, в котором участвует примерно 4% тестостерона, происходит в
результате восстановления двойной связи кольца А и 3-кетогруппы при участии
цитоплазматического фермента - NADPH-зависимой 5α-редуктазы.
Семенники человека секретируют в сутки до 50-100 мкг дигидротестостерона.
Однако большее количество гормона - следствие периферических превращений, и
суммарная суточная секреция дигидротестостерона составляет 400 мкг, что почти в
10 раз меньше уровня секреции тестостерона.
В некоторых периферических тканях
небольшое количество тестостерона превращается в эстрадиол. В качестве побочных
продуктов клетки Лейдига также постоянно секретируют эстрадиол и прогестерон,
хотя роль этих гормонов в развитии и поддержании функций размножения и
формирования полового поведения у мужчин до настоящего времени не выяснена.
В препубертатный период секреция
андрогенов подавляет по механизму отрицательной обратной связи секрецию
гонадотропина до начала пубертатного периода, когда гипофизарные клетки
становятся менее чувствительными к ингибирующему действию циркулирующих в крови
андрогенов. Эта потеря чувствительности приводит к циклически импульсному
освобождению ЛГ и ФСГ. ЛГ, связываясь с рецепторами клеток Лейдига, стимулирует
образование тестостерона интерстициальными клетками Лейдига, а ФСГ, связываясь
с рецепторами клеток Сертоли в семенниках, стимулирует сперматогенез.
Тестостерон замыкает отрицательную обратную связь на уровне гипофиза и
гипоталамуса, уменьшая частоту секреторных импульсов ЛГ. Торможение секреции
ФСГ аденогипофизом происходит под действием белка ингибина, вырабатываемого
клетками Сертоли. ФСГ стимулирует синтез этого белка, который по механизму
отрицательной обратной связи тормозит дальнейшую секрецию ФСГ.
К мишеням тестостерона относят
эмбриональные вольфовы структуры, сперматогонии, мышцы, кости, почки, мозг.
Подобно другим стероидным гормонам, андрогены образуют внутри клетки комплекс с
рецептором, который связывается с определённым участком хроматина, активируя
специфические гены, белковые продукты которых опосредуют биологические эффекты
андрогенов.
Физиологическое действие андрогенов
различно в разные периоды жизни организма. У эмбриона под действием андрогенов
из вольфова протока образуются придаток яичка (эпидидимис), семявыносящий
проток и семенной пузырёк. У плода мужского пола происходит маскулинизация
мозга. Поскольку андрогены в организме обладают мощным анаболическим действием
и стимулируют клеточное деление, повышенный уровень андрогенов в препубертатный
период приводит к скачкообразному увеличению линейных размеров тела, увеличению
скелетных мышц, росту костей, но одновременно способствуют и остановке роста,
так как стимулируют сращение эпифизов длинных костей с их стволами. Андрогены
вызывают изменение структуры кожи и волос, снижение тембра голоса вследствие
утолщения голосовых связок и увеличения объёма гортани, стимулируют секрецию
сальных желёз.
Вода в живом организме. Состояние
воды в живых тканях, ее функции. Эндогенная вода
Вода. За небольшим исключением
(кость и эмаль зубов), вода является преобладающим компонентом клеток - в
среднем 75-85%. В клетке вода находится в свободном и связанном состоянии.
Биологическое значение воды: растворитель (для полярных веществ, неполярные
вещества в воде не растворяются); среда для реакций, участник реакций
(расщепление белков), участвует в поддержании теплового равновесия клетки;
источник кислорода и водорода при фотосинтезе; основное средство передвижения
веществ в организме.
Вся вода, что есть в организме,
разделяется на внутриклеточную и внеклеточную, каждая из которых существует в виде
двух фракций:
а) фракция воды, способная к обмену;
б) фракция воды, связанная в
коллоидных системах с молекулами органических веществ (белков, жиров,
углеводов).
Около 66 % всей воды организма
входит в состав внутриклеточной жидкости и 33 % находится вне клеток.
Внеклеточная вода является основой внутрисосудистых жидкостей (лимфы, плазмы
кровь) и межклеточной жидкости.
Функции воды.
Вода принимает участие в:
формировании внутриклеточных
структур;
формировании пространственных
конформаций молекул белков (вторичной, третичной);
многих биохимических реакциях
гидролиза, гидратации, окисления, возобновление и др.;
осуществляет транспортную функцию за
счет высокой растворимой способности и текучести;
принимает участие в выведении из
организма продуктов разпада;
является средой для осуществления
почти всех химических реакций в организме;
принимает участие в теплорегуляции
путем испарения из легких и из поверхности кожи;
осуществляет механическую функцию -
является компонентом смазывания трущихся поверхностей в суставах, добавляет
упругости хрящам и межпозвонковым дискам;
является компонентом секретов
организма: пота, мочи, слюны, молока и тому подобное.
Ткани и клетки используют два вида
воды: экзо и эндогенную.
Потребность организма в воде зависит
от возраста, интенсивности обменных процессов, мускульной деятельности,
функционального состояния почек, температуры окружающей среды, состава еды. У
взрослых она составляет в среднем 40 г на 1 кг массы тела, для детей эта
величина приблизительно втрое больше - от 70 до 150 г на 1 кг массы тела. За
сутки к организму в нормальных условиях поступает около 2,5 л воды, включая
экзо- и эндогенную. Последняя усваивается в организме при распаде белков,
углеводов и особенно жиров. При том, что при окислении 100 г жира образуется
107 мл воды, 100 г белка - 41 мл воды, 100 г углеводов - 55 мл воды.
Всасывание воды, что поступает с
едой, происходит по всему желудочно-кишечному тракту: небольшое количество воды
всасывается в ротовой полости и пищеводе, часть - в желудке, часть - в толстом
кишечнике, основная масса - в тонком кишечнике.
Вода с питательными веществами путем
диффузии и осмоса, а частично пиноцитоза и активного транспорта, проникает
внутрь эпителия слизистых оболочек кишечника. Эндоплазматичной сетью она
перемещается в клетки и поступает в межклеточное пространство, потом в
межклеточную жидкость, капилляры, венулы, подэпителиальную и пидслизистую
венозной сетки кишечной ворсинки, воротную вену, печень и большой круг
кровообращения. Часть воды поступает через лимфатическую систему. Выведение
воды из организма осуществляется с мочой, калом, потом и выдыханием.
|
Водный
баланс
|
|
Употребление
воды
|
мл
|
Выделение
воды
|
мл
|
|
Жидкости
|
1500
|
Моча
|
1600
|
|
Вода
с пищи
|
800
|
900
|
|
Вода
с обмена веществ
|
300
|
Кал
|
100
|
|
Итого
|
2600
|
Итого
|
2600
|
Почки являются основным органом выведения воды и
электролитов из организма. В обычных условиях за сутки выделяется через кожу и
почки почти 1500 мл воды, через легкие - до 400 мл и через кишечник -
приблизительно 100 мл. Вся вода организма обновляется примерно за 4 недели. За
1 мин. вода крови успевает обновиться на 73 %.
Полисахариды морских водорослей. Состав,
строение. Функции в организме
Полисахариды морских растений объединяют
полисахариды, выделяемые из красных и бурых морских водорослей. В пищевой
промышленности широко используются альгинаты, каррагинаны и агароиды.
Альгиновая кислота ( Е400) и ее соли (E40I--E405) относятся к полисахаридам
бурых морских водорослей родов Laminaria и Macrocystis (от лат. alga -
водоросль), которые построены из остатков маннуроновой и гулуроновой кислот,
находящихся в пиранозной форме и снизанных в линейные цепи (1,4)-гликозидными
связями.

Агар (агар-агар) Е406 - смесь полисахаридов
агарозы и агаропектина. Основная фракция агарозы - линейный полисахарид,
построенный из чередующихся остатков β-D-галактопиранозы
и З.6-ангидро-β-L-raлактопиранозы,
связанных попеременно β -(1,4)- и
β-(1,3)-связями.
Агаропектин - смесь полисахаридов сложного строения, содержащая глюкуроновую
кислоту и эфирносвязанную серную кислоту.

Звено агаробиозы
Агар-агар получают из красных морских водорослей
(Gracilaria, Gclidium, Ahnfeltia), произрастающих на Белом море, Тихом и
Атлантическом океанах. Агар незначительно растворяется в холодной воде, но
набухает в ней. В горячей воде он образует коллоидный раствор, который при
охлаждении дает хороший прочный гель, обладающий стекловидным изломом. У агара
этот процесс осуществляется за счет образования двойных спиралей и их
ассоциации независимо от содержания катионов, сахара или кислоты.
Гелеобразующая способность агара в 10 раз выше, чем у желатина. При нагревании
в присутствии кислоты способность к гелеобразованию снижается. Гели стабильны
при рН более 4,5 и термообратимы.
Каррагинаны (Е407) объединяют семейство
полисахаридов (известное также под названием ирландский мох), содержащихся,
наряду с агаром, в красных морских водорослях Chondrus Crispis, Eucheuma
Spinosum, Gigartina species и др. По химической природе каррагинаны близки к
агароидам и представляют собой неразветвленные сульфатированные гетерогликаны.
Обычные каррагинаны являются гелеобразователями, а лямбда-тип - загустителем.
Классификация липидов. Сложные липиды.
Фосфолипиды. Строение. Распространение. Биологическая роль
Простые липиды:
ацилглицеролы (жиры, триглицериды и т.п.);
воска.Сложные липиды:
фосфолипиды
• глицерофосфолипиды (фосфатидилхолин,
фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозитол, фосфатидилглицерол,
кардиолипин, плазмалоген);
• сфингофосфолипиды (сфингомиелин);
стероиды (холестерол, эргостерол, ланостерол,
стигмастерол, экдистероиды);
гликолипиды (цереброзиды, ганглиозиды,
сульфатиды).
Фосфолипиды делят на глицерофосфолипиды, основу
которых составляет трёхатомный спирт глицерол, и сфинго-фосфолипиды -
производные аминоспирта сфингозина. Фосфолипиды имеют амфифильные свойства, так
как содержат алифатические радикалы жирных кислот и различные полярные группы.
Благодаря своим свойствам фосфолипиды не только являются основой всех клеточных
мембран, но и выполняют другие функции: образуют поверхностный гидрофильный
слой липопротеинов крови, выстилают поверхность альвеол, предотвращая слипание
стенок во время выдоха. Некоторые фосфолипиды участвуют в передаче
гормонального сигнала в клетки. Сфингомиелины являются фосфолипидами,
формирующими структуру миелиновых оболочек и других мембранных структур нервных
клеток.
Для всех фосфолипидов характерно, что одна часть
их молекул (радикалы R1и R2) обнаруживает резко выраженную гидрофобность, тогда
как другая часть гидрофильна благодаря отрицательному заряду фосфорной кислоты
и положительному заряду радикала R3.
Если в третьем положении имеется только
фосфорная кислота, то глицерофосфолипид называется фосфатидной кислотой. Её
остаток называют фосфатидил; он входит в название остальных
глицерофосфолипидов, после которого указывают название заместителя атома
водорода в фосфорной кислоте, например фосфатидилэтаноламин, фосфатидилхолин и
т.д.
Фосфатидная кислота в свободном состоянии в
организме содержится в небольшом количестве, но является промежуточным
продуктом на пути синтеза как три-ацилглицеролов, так и глицерофосфолипидов. У
глицерофосфолипидов, как и у триацилгли-церолов, во второй позиции находятся
преимущественно полиеновые кислоты; в молекуле фосфатидилхолина, входящего в
структуру мембран, это чаще всего арахидоновая кислота. Жирные кислоты
фосфолипидов мембран отличаются от других липидов человека преобладанием
полиеновых кислот (до 80-85%), что обеспечивает жидкое состояние гидрофобного
слоя, необходимое для функционирования белков, входящих в структуру мембран.
В зависимости от характера азотистого основания,
присоединенного к фосфорной кислоте, глицерофосфолипиды подразделяют на
фосфатидилхолины (лецитины), фосфатидилэтаноламины (кефалины) и
фосфатидилсерины. Участвуют в построении клеточных мембран.
В молекуле фосфатидилхолина одна из трех
гидроксильных групп глицерина связана не с жирной, а с фосфорной кислотой.
Фосфорная кислота в свою очередь соединена эфирной связью с азотистым
основанием - холином [НО-СН2-СН2-N+(CH3)3].

Холин (основание) -
(триметил-2-гидроксиэтиламмоний)
Основными фракциями коммерческих лецитинов
являются фосфатидилхолины, т. е. собственно лецитины (ло 25%), фосфатид и
этаноламины (до 25%). фосфатидилсерины (до 15%). фосфатидилинозиты,
фосфатидовые (фосфатидные) кислоты (5-10%). Основным источником промышленного
получения лецитинов для пищевой промышленности являются масличные культуры
(главным образом, соя, реже - подсолнечник), откуда их выделяют при гидратации
масел. Особенности эмульгирующих свойств фосфолипидов обусловлены способностью
образовывать и поддерживать в однородном состоянии как прямые, так и обратные
эмульсии, что распространяет их использование на все виды пищевых эмульсий: от
майонезов и различных салатных соусов (прямые эмульсии) до маргаринов
различного жирно-кислотного состава и разного содержания жировой фазы (обратные
эмульсии). Другой отличительной особенностью фосфолипидов как пищевых
эмульгаторов является их способность образовывать липосомы (липидные везикулы)
- частицы, формируемые концентрическими замкнутыми липидными бислоями с внутренним
водным слоем, изолированным от внешней среды и содержащим, в зависимости от
назначения липосом, различные включения, например, пептиды или белки.
Использование липосомальных систем в пищевых продуктах связано с функциями
зашиты отдельных пищевых ингредиентов от внешнего воздействия (защита дрожжевых
клеток от охлаждения в замороженных мучных полуфабрикатах и пицце), сохранения
влаги (мороженое) или органических, например, вкусовых веществ (хлеб и
бисквиты). И в отличие от большинства других пищевых добавок, препараты
фосфолипидов обладают высокой физиологической эффективностью, связанной с
уменьшением уровня холестерина, улучшением функции печени и состояния
центральной и периферической нервной системы, торможением процессов старения
организма и нормализацией иммунобиологической реактивности организма. И хотя
диетологи не относят фосфолипиды к незаменимым факторам питания, они являются
физиологически ценными компонентами пищи, суточная потребность в которых
составляет около 5 г.
Основное различие между фосфатидилхолинами и
фосфатидилэтаноламинами - наличие в составе последних вместо холина азотистого
основания этаноламина (HO-CH2-CH2-N+H3).
Фосфатидилсерины. В молекуле фосфатидилсерина
азотистым соединением служит остаток аминокислоты серина
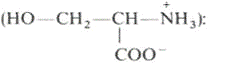
Фосфатидилсерины распространены гораздо менее
широко, чем фос-фатидилхолины и фосфоэтаноламины, и их значение определяется в
основном тем, что они участвуют в синтезе фосфатидилэтаноламинов.
Фосфатидилсерин участвует в организме в синтезе коламинкефалина путем
декарбоксилирования.
Препараты фосфолипидов обладают высокой
физиологической эффективностью, связанной с уменьшением уровня холестерина,
улучшением функции печени и состояния центральной и периферической нервной
системы, торможением процессов старения организма и нормализацией
иммунобиологической реактивности организма.
При дефиците холина нарушается синтез
транспортных форм липидов. Фосфатидилхолин (лецитин), участвует в синтезе
лецитина, который обеспечивает перенос липидов из печени и является важным
компонентом клеточных мембран. Помимо глицерина и жирных кислот, которые входят
в состав нейтральных жиров, для синтеза фосфолипидов необходимы неорганические
фосфаты и азотистые соединения, в частности холин, для синтеза фосфатидхолина.
Неорганические фосфаты в печени имеются в достаточном количестве.
При недостаточном образовании или недостаточном
поступлении в печень холина синтез фосфолипидов из компонентов нейтрального
жира становится либо невозможным, либо резко снижается и нейтральный жир
откладывается в печени. Иными словами, синтез фосфолипидов лимитируется
количеством азотистых оснований, т.е. для синтеза фосфоглицеридов необходим
либо холин, либо соединения, которые могут являться донорами метильных групп и
участвовать в образовании холина (например, метионин).
Плазмалогены - фосфолипиды, у которых в первом
положении глицерола находится не жирная кислота, а остаток спирта с длинной
алифатической цепью, связанный простой эфирной связью.
Характерный признак плазмалогенов - двойная
связь между первым и вторым атомами углерода в алкильной группе. Плазмалогены
бывают 3 видов: фосфатидальэтано-ламины, фосфатидальхолины и
фосфатидаль-серины. Плазмалогены составляют до 10% фосфолипидов мембран нервной
ткани; особенно много их в миелиновых оболочках нервных клеток.
Классификация глицерофосфолипидов и
сфинголипидов
|
Ацилглицеролы
|
Фосфолипиды
|
Сфинголипиды
|
|
Триацилглицеролы
|
Сфингомиелины*
|
|
Глицерофосфолипиды:
|
Гликолипиды:
|
|
Диацилглицеролы
|
Фосфатидилхолин
|
Цероброзиды
|
|
Моноацилглицеролы
|
Фосфатидилсерин
|
Глобозиды
|
|
|
Фосфатидилэтаноламин
|
Сульфатиды
|
|
|
Фосфатидилглицерол
|
Ганглиозиды
|
|
|
Фосфатидилинозитолбисфосфат
|
|
|
|
Фосфатидная
кислота
|
|
|
Кардиолипин
(дифосфатидилглицерол)
|
|
*Сфингомиелины относят как к фосфолипидам, так и
сфинголипидам.
Сфинголипиды
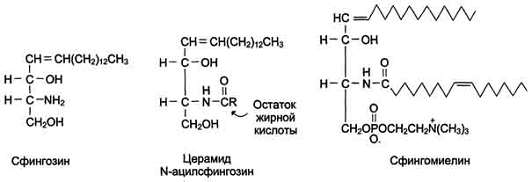
Аминоспирт сфингозин, состоящий из 18 атомов
углерода, содержит гидроксильные группы и аминогруппу. Сфингозин образует
большую группу липидов, в которых жирная кислота связана с ним через
аминогруппу. Продукт взаимодействия сфингозина и жирной кислоты называют
церамид. В церамидах жирные кислоты связаны необычной (амидной) связью, а
гидроксильные группы способны взаимодействовать с другими радикалами. Церамиды
отличаются радикалами жирных кислот, входящих в их состав. Обычно это жирные
кислоты с большой длиной цепи - от 18 до 26 атомов углерода.
Сфингомиелины. В результате присоединения к
ОН-группе церамида фосфорной кислоты, связанной с холином, образуется
сфингомиелин) . Сфингомиелины - основные компоненты миелина и мембран клеток
мозга и нервной ткани. Сфингомиелины, как и глицерофосфолипиды, имеют
амфифильные свойства,
Литература
Березов
Т.Т., Коровкин Б.Ф. Биологическая химия. - М.: Медицина, 1990.
Большая
медицинская энциклопедия.- М.: Советская энциклопедия, 1977.
Комов
В.П., Шведова В.Н. - Биохимия.- М.: Дрофа, 2004.-638 с.
Филиппович
Ю.Б. Основы биохимии. - М.: Высшая школа, 1985. - 503 с.
Андрианова
Л.Е., Авдеева Л.В., Алейникова Т.Л.- Биохимия /под ред. Северина Е.С. - М.: ГЭОТАР-Медиа,
2006.- 784 с.
Ленинджер
А. Биохимия: Пер. с англ. - М.: Мир, 1976. - 957 с.
Пищевая
химия: Учеб. Для вузов / А.П. Нечаев, С.Е. Траубенберг, А.А. Кочеткова и др. /
Под общ. Ред. А.П. Нечаева. - 3-е изд., испр. -СПб.: ГИОРД, 2004. - 640 с.