Фізико-хімічні методи одержання наноматеріалів
Зміст
Вступ
1. Виробництво наноматеріалів за допомогою мікроорганізмів
1.1 Використання специфічних властивостей мікроорганізмів
1.1.1 Віруси
1.1.2 Бактеріальні клітини
1.1.3 Гриби
1.2 Отримання сировини методами генної інженерії
1.3 Використання продуктів життєдіяльності мікроорганізмів
2. Нанобіоелектроніка
2.1 Біокомп’ютери
2.1.1 Елементи ДНК-комп’ютера
2.1.2 Запам’ятовуючі пристрої на вірусній основі
2.1.3 Клітинний біокомп’ютер
2.1.4 Модульні логічні вентилі з бактеріальних геномів
2.2 Мікробні клітинні біосенсори
2.2.1 Біосенсори на основі генетично модифікованих бактеріальних
клітин
2.2.2 Люмінесцентні біосенсори
2.2.3 Системи ферментів мікроорганізмів в якості промоторів
Висновок
Список літератури
Вступ
Останні десятиліття визначаються бурхливим розвитком
нанотехнології, фізико-хімічні методи одержання наноматеріалів вже не можуть
забезпечити необхідну якість сировини для зборки нанопристроїв.
Учені констатують необхідність вивчення
механізмів, що відбуваються в біологічних системах, і використання
фундаментальних принципів їх дії, оскільки в живих клітинах ефективно
функціонують молекулярні наномашини, на вдосконалення яких природа витратила
3,5 млрд років еволюції. Мікроорганізми - найдавніші організми, що існують на
Землі і досі процвітають в біосфері, оскільки пристосувалися до всіх можливих
умов існування: екстремальні температурні умови, високі концентрації різних
хімічних сполук, аеробні та анаеробні умови; що означає широкий спектр хімічних
і біохімічних процесів, що проходять всередині організму.
З іншого боку, методи генної інженерії дають
можливість не тільки використовувати вже закладені природою властивості
мікроорганізмів, а і власноруч програмувати необхідні процеси. Це є підставою
для пошуків методів синтезу наночастинок, наностержнів, нанотрубок,
нанопроволок, нанокомпозитів, нановолокон, нанокапсул тощо.
Для утримання мікроорганізмів не використовуються
високі температури, тиски і агресивні середовища, що забезпечує потенціальну
екологічність і безпечність технологій на їх основі.
Вивчення даної теми є необхідним, тому що
використання мікроорганізмів може призвести до революції у нанотехніці, наномедицині та інших галузях.
Метою даної роботи є визначення фундаментальних
принципів, методів, перспектив розвитку і застосування нанотехнологій з
використанням мікроорганізмів та продуктів їх життєдіяльності.
1.
Виробництво наноматеріалів за допомогою мікроорганізмів
Вимоги до компонентів наноелектронної
промисловості стають все більше. Машини, які традиційно використовуються для
виготовлення деталей незграбні на таких малих масштабах, в той час як природа
має ідеальні, вдосконалені часом і еволюцією інструменти інструменти для обходу
цієї проблеми. Більшість об'єктів вивчення мікробіології можна класифікувати як
нанооб'єкти: бактерії, розмір яких знаходиться в інтервалі між 1 і 10 мкм,
відносять до об'єктів мезоскопічних масштабів; віруси з розмірами від 10 до 200
нм знаходяться у верхній частині діапазону наночастинок; і відповідно елементи
їх будови. Ті ж мікроорганізми, що не входять в межі наномасштабів, можуть бути
розглянуті як фабрики з виробництва наноматеріалів.
1.1
Використання специфічних властивостей мікроорганізмів
1.1.1
Віруси
З точки зору матеріалознавців, віруси можна
розглядати як біорганічні наночастки. Вони складаються з невеликої кількості
різних біополімерів: білків і нуклеїнових кислот. Багато вірусів оточені
ліпідною мембраною, вони не мають власного метаболізму, але використовують
метаболічний апарат живої клітини для самовідтворення; їхня поверхня несе
специфічні засоби для подолання бар'єрів клітин-господарів. Розмір і форма
вірусів, а також кількість і характер функціональних груп на їхній поверхні
точно визначені. Саме ці властивості вірусних клітин і є підставою для
використання їх в матеріалознавстві як каркасів для ковалентно зв'язаних
модифікацій поверхні. Особливістю вірусів є те, що вони можуть бути адаптовані
шляхом спрямованої еволюції, використовуючи їх вбудовані колокалізації генотипу
і фенотипу. Потужні методи стають основою інженерних підходів до наноматеріалів,
відкриваючи широкий діапазон застосувань далеко за межами біології та медицини
[1].
Наразі вчені розглядають можливість використання
вірусів у якості тривимірних платформ синтезу композитних матеріалів, на
противагу нанотехнологічним методам розміщення молекул у двох вимірах. Такі
системи можуть бути використані в синтезі металевої нанопроволоки, попередниками якої є розташовані в порожнині вірусу наночастинки [2,3].
Центральний канал з вірусу тютюнової мозаїки
(ВТМ) використаний як матриця для синтезу нікелевих і кобальтових нанопроволок
діаметром в кілька атомів і довжиною до діапазону мікрометра у роботі [3].
Самоасоційовані циліндричні частинки дикого типу
та рекомбінантний вірус тютюнової мозаїки (ВТМ) були використані в якості
органічних шаблонів для контрольованого осадження та організації наночастинок
Pt, Au, або Ag [2]. Хімічне руйнування [PtCl6] 2 - або [AuCl4] -
комплексів при низьких значеннях рН призвело до специфічного покриття
зовнішньої поверхні дикого типу ВТМ металевими наночастками менше 10 нм за
розміром. На відміну від цього, внаслідок фотохімічного розкладання солей Ag
(I) при рН=7 утворилися дискретні Ag наночастинки, які створили форму полого
канала з внутрішнім діаметром 4нм. Число інкапсульованих наночастинок Ag
збільшилось при використанні бензоату замість нітрату срібла, так як Ag/бензоат
окисно-відновна пара посилювала на поверхнево-шаблонний ефект карбоксилатов
стінок каналу. Подібні експерименти з використанням мутантного ВТМ зі
зменшенням негативного заряду вздовж центральної порожнини підтвердили, що
глутамінова і аспартатна кислотні групи залучені в осадженні по конкретних
ділянках [2].
Одним із найсучасніших прикладів є розробка
дослідницької групи А. Белхер 2015 року [4]. Вони використовують вірус M13 як
універсальний біологічний каркас, який був генетично створений, щоб розміщувати
різні матеріали на мезопористих структурах, заснованих на нанопроволоці (НП). У
цьому дослідженні, НП з високим вмістом діоксиду титану синтезовані з
використанням вірусів М13 в якості шаблонів, і НП зібрані в напівпровідникові
мезопористі мережі зі здатною до перебудови структурою. Щоб зрозуміти вплив
різних морфологій на продуктивність фотоелектрики, мережі були встановлені в
якості фотоанодів в рідкофазних сенсибілізованих барвником сонячних елементах
(DSC). У порівнянні з традиційними фотоанодами на основі наночасток,
нанопроволочні фотоаноди продемонстрували значно вищі довжини дифузії
електронів при збереженні порівнянної потужності світлопоглинання, що
призводить до підвищеної ефективності перетворення енергії. Крім того,
напівпровідникові мезопористі тонкі плівки на основі НП здатні завантажити
достатньо органосвинцево йодид перовскітних матеріалів в сполучені пори, і
плівки, покриті перовскітом використовуються в якості ефективних фотоанодів для
твердофазних органосвінцевих йодид перовскітних гібридних сонячних елементів
для досягнення ефективності перетворення енергії вище, ніж у рідкофазній DSC
[4].
1.1.2
Бактеріальні клітини
Бактерії - найдавніша з відомих груп організмів,
що існують на Землі і досі процвітають в біосфері. За 3,5 млрд років бактерії
пристосувалися до всіх можливих умов існування: екстремальні температурні
умови, високі концентрації різних хімічних сполук, аеробні та анаеробні умови;
що означає широкий спектр хімічних і біохімічних процесів, що проходять
всередині організму. До того ж збільшення популяції бактерій відбувається з
високою швидкістю і потребує мінімальних витрат на створення умов. Ці
властивості роблять бактерії незамінними "біомашинами" нанотехнології.
березня 2015р вчені з Іллінойського університету
в Чикаго опублікували результати роботи, в якій закріпили набір квантових точок
на базі графену на бактерії, зробивши з неї пристрій для вимірювання вологості.
Оболонка бактеріальних спор розширюється при впливі молекул води, що змінює
провідність точок графена і електродів. Бактерії здатні перетворюватися в
спору: клітина втрачає воду, об'єм і форму, утворюючи під зовнішньою мембраною
щільну сферичну оболонку. Ця оболонка витримує механічні, температурні і
хімічні навантаження, але реагує на них певним чином. Присутність молекул води
змушує оболонку розширюватися. Вчені закріпили на бактеріальну спору сінної
палички Bacillus subtilis точки на базі графену. При зміні рівня
вологості навколо бактерії її об'єм змінюється, що призводить до зміни
провідності точок графена. Така бактерія в десять разів швидше реагує на зміну
вологості, ніж використовувані зараз датчики, і крім цього здатна працювати у
вакуумі. Нанорозмірні біосенсори на зразок пристрою NERD можуть бути корисні в
консервуванні харчових продуктів та моніторингу корозії, де має значення
низький рівень вологості в процесі роботи. Також сенсори можуть бути
використані в космонавтиці, завдяки тому що бактеріальна спора здатна виживати
десятки років, так що датчик на її основі зможе працювати протягом тривалого
часу [5].
Іншим яскравим прикладом використання
особливостей життєдіяльності і будови мікроорганізмов можуть служити двомірні
бактеріальні S-шари, про використання яких в нанотехнології говориться вже
багато років. Першим структуру, організацію та технологічне застосування таких
збірок досліджував Уве Слейтр з колегами [6]. S-шари поверхневі структури
бактеріальної клітини, виявлені у сотень видів бактерій, що володіють клітинною
стінкою, і у всіх архебактeрій. Архебактерії - це унікальна група прокаріот, що
включає екстремальних термофілів і галофітів. Ця вельми цікава група
філогенетично відособлена від бактерій і володіє деякими рисами подібності з
еукаріотами, особливо щодо механізмів транскрипції і трансляції. Багато
архебактерій екстремофіли (тобто організми, що існують тільки в екстремальних
умовах середовища). Механізми адаптації архей і деяких бактерій до таких
екстремальних умов можна було б використовувати для створення стабільних
наноприладів, здатних працювати довгий час (низька стабільність біомолекул один
з факторів, що обмежують їх використання).
За мільярди років еволюції збірка S-шарів стала
настільки досконалою, що не має собі рівних за впорядкованістю та ефективністю
самозбірки серед біологічних систем. Ці плоскі шари утворюються з одного виду
білків або глікобілків, молекули яких збираються в єдиний шар, що покриває
поверхню клітини. Шар має симетричну порожнисту структуру з відстанню між
вузлами від 3 до 30 нм. Товщина S-шарів варіює від 5 до 15 нм, шари пронизані
порами діаметром 2 - 6 нм.
Встановлено, що S-шари, які утворюються внаслідок
самозбірки, дуже зручні для застосування в літографії. In vitro (У розчині)
очищені будівельні блоки S-шарів спонтанно утворюють високовпорядковані
двомірні кристали. Використовуючи цю властивість, У. Слейтр з колегами показали
можливість перекристалізації субодиниць S-шарів на різних нанотехнологічних
субстратах, таких як чіпи з кремнію або оксиду кремнію. Інші дослідники
виявили, що ці шари можуть формуватися на ліпідних мембранах і на поверхні
розділу повітряного і водного середовищ. Використовувалися S-шари і як матриці
для зв'язування та створення наночастинок, таких як напівпровідникові і
металеві точки [7], і для створення біомолекул - ферментів і антитіл. Такий тип
самозбірки та взаємодії молекул дозволяє застосовувати кристалічні матриці для
нанесення малюнка в нанолітографії.
Також було встановлено, що УФ промені можна
використовувати для нанесення малюнка на S-шари. На сформований па поверхні
кремнієвого чіпа S-шар накладається фотомаска, після чого він піддається УФ
опроміненню. Літографію, засновану на застосуванні таких шарів разом з
біомолекулами і неорганічними сполуками, можна застосовувати для виробництва
так званих лабораторій на чіпі (lab-on-a-chip). S-шари також використовуються
для створення ультрафільтруючих мембран, які пропускають сполуки певної
молекулярної маси, а також для іммобілізації молекул на шари з чітко визначеною
геометрією [7].
Робота групи вчених Масачусинського університету
[8] розглядає способи культивування клітин G. sulfurreducens для
отримання провідних пілів, а також методи їх виділення та модифікації з метою
отримання біологічних нанодротів або біологічних/неорганічніх гібридних
пристроїв з різними функціональними можливостями. Винахід стосується
ізольованих нативних або модифікованих пілей а також клітин, що експресують
нативні або модифіковані пілі.
Основним завданням в наноелектроніці окрім
виробництва нанорозмірних матеріалів є також розробка і виготовлення
нанодротів, оскільки вони є основною зв'язуючою ланкою нанорозмірних приладів.
Хоча раніше білки були в центрі уваги, їхні можливості були в основному
обмежені низьким рівнем провідності, виявлених у багатьох нанодротів на основі
білка, навіть після металізації.
Геопілі, продуковані Geobacter sulfurreducens
пілі, можуть бути застосовані в будь-якому пристрої, де потрібні нанодроти.
Вони включають, але не обмежуються нанодротами з електронними властивостями,
починаючи від металевих до напівпровідникових, для контролю заряду електронів в
наноприладах, біологічних і хімічних біосенсорах. Переваги геопілів в тому, що
їх масове виробництво є порівняно нескладним і вони володіють природною
провідністю, що тим самим зменшує виробничі витрати, і також можуть бути змінені
за допомогою звичайних генетичних маніпуляцій, що дозволяє отримати нанодроти з
різними функціональними можливостями [8].
На основі Lactobacillus plantarum створена
нова біонанотехнологія з очищення систем опалення, трубопроводів, котлів та
іншого теплотехнічного обладнання від накипу та інших відкладень. У процесі
роботи теплового обладнання, утворюються солевидкладення, які не вдається
видалити реагентами або механічно (карборунд, цементитом, магнезіти, сідеріти
та інші солі). Ці мінерали нерозчинні у воді, кислотостійких, відмінні
ізолятори, мають високу механічну міцність. Новий спосіб очищення від шламу і
накипу за допомогою біоорганічної рідини безпечний для людини і навколишнього
середовища, дешевий, так як використовуються відходи молокопереробних
підприємств - молочна сироватка [9]. Дана технологія є прикладом побутового
застосування нанотехнологій на основі бактеріальних клітин.
1.1.3
Гриби
Мікроскопічні гриби також є об’єктом
нанотехнологічних досліджень. Дослідникам з Northwestern University (США)
вдалося покрити тонким шаром золота гіфи - тонкі волокна, утворені грибом [10].
Спори гриба Aspergillus niger культивували у присутності частинок золота
розміром 13 нм. Частинки осідали на поверхні гіф. До часток золота були
приєднані фрагменти однолацюгової ДНК, що дозволяло приєднувати інші
мікроскопічні об'єкти, на поверхні яких знаходилися комплементарні фрагменти
ДНК. Аналогічні маніпуляції були пророблені і з іншими видами грибів, які
утворюють гіфи різного розміру. Таким чином можна конструювати досить складні
структури з наночасток. За допомогою грибів можна буде створювати матеріали
заданої наноструктури для застосування в оптиці і електроніці. Дріжджі в роботі
[11] використовують для отримання мікросфер шпінелі Co3O4.
1.2
Отримання сировини методами генної інженерії
Як зазначалося у пункті 1.1, особливістю вірусів
є те, що вони можуть бути адаптовані шляхом спрямованої еволюції,
використовуючи їх вбудовані колокалізації генотипу і фенотипу, що означає
можливість не тільки використовувати вже закладені природою властивості, а і
власноруч програмувати необхідні процеси. Проведено безліч дослідів, що
доводять дивовижні властивості наночастинок, отриманих з використанням
модифікованих вірусних клітин.
наноматеріал мікроорганізм нанотехнологія
В одній з найперших робіт в цьому напрямку за
рахунок генної інженерії на поверхню капсида М13 були поміщені білки, які
специфічо з'єднуються з наночастинками різних матеріалів: ZnS, або CdS, або
CoPt, або FePt. Після закінчення злипання віруси піддавали термічній обробці
при температурі 400-500°С, за якої органічні речовини (ДНК і білки вірусу)
згорали, виходили мінеральні нанопровідники. Цей метод виявився економічно
невигідним і занадто складним [12].
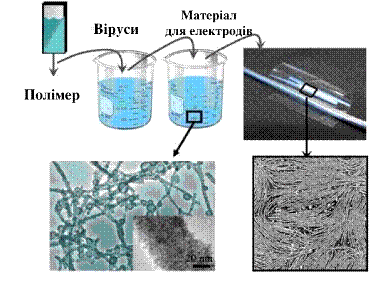
Рисунок 1 - Схема отримання наночастинок оксиду
кобальту
Наприклад, у роботі [13] вищезгаданою
дослідницькою групою А. Белхер використовуються частинки вірусу М13. При
додаванні послідовності нуклеотидів до ДНК вірусу М13 вдалося створити його
модифікацію, яка при розмноженні взаємодіяла з іонами кобальту. На рис.1
наведена схема отримання наночастинок оксиду кобальту з використанням
нанобіотехнології. Пластина з твердого полімеру-електроліту занурювалася в
розчин вірусу М13, після чого опускалася в розчин з іонами кобальту. В
результаті пластина покривалася шаром оксиду кобальту, який є цінним матеріалом
для електродів літій-іонних і літій-полімерних батарей. Віруси були генетично
модифіковані таким чином, щоб на поверхні їх білкової оболонки перебували
пептиди, здатні зв'язувати сульфід цинку або сульфід кадмію, які є
надпровідниковими кристалами [13]. При досить високій концентрації частинок
бактеріофага в розчині вони самостійно організовувалися в структуру на зразок
рідкого кристала, в якій напівпровідникові кристали розташовувалися по одній
лінії. При використанні інших технологій домогтися аналогічного результату не
вдавалося. Даний метод дозволяє організовувати віруси в мініатюрні дроти.
Рідкокристалічна система [14] була використана
для виготовлення високо впорядкованого композитного матеріалу за допомогою
генномодифікованого M13 бактеріофага і нанокристалів сульфіду цинку (ZnS).
Бактеріофаги, які лягли в основу системи самообслуговування, були відібрані за
наявністю специфічного фрагменту розпізнавання ZnS-поверхні кристала.
Бактеріофаги з'єднували з попередниками ZnS і спонтанно утворювався гібридний
плівковий матеріал, який поділявся на нанорівні і в масштабі мікрометра в
приблизно 72-мікрометрові домени, неперевно пов'язані на довжині близько
сантиметра. Крім того, були приготовані суспензії, в яких поведінка ліотропної
рідкої кристалічної фази гібридного матеріалу контролюється концентрацією
розчинника і використанням магнітного поля [15].
1.3
Використання продуктів життєдіяльності мікроорганізмів
Відомо багато мікроорганізмів, які виробляють
неорганічні речовини. Властивості матеріалів, отриманих за допомогою
мікроорганізмів, як правило, перевершують аналогічні синтетично вироблені
матеріали з аналогічним фазовим складом. Отримані структури високо організовані
від молекулярного до нано, мікро і макро масштабів, часто в ієрархічному
порядку, зі складною наноархітектурою, і в кінцевому підсумку складають безліч
матеріалів із широким спектром властивостей. Вони одночасно є ''розумною'',
динамічною, складною, здатною до самовідновлення і багатофункціональною
системою.
Одноклітинні діатомові водорості виробляють
кремнезем - сполуку кремнію з киснем, склад якої відповідає звичайному склу.
Діатомові водорості мають "панцир", до складу якого входить діоксид
кремнію. Їх можна використовувати для отримання наноструктурованих частинок
діоксиду кремнію [16]. На жаль, такий метод синтезу не передбачає отримання
точно однакових наноструктур, і тим більше структури на мікрорівні. Це залежить
від панцира водоростей і від варіацій в їх структурі. Діатомові водорості
зображені на рис.2. Недоліком методу є те, що розмір пір і сама структура
одержуваних матеріалів визначені структурою бактерій. Найчастіше вдається
отримати тільки аморфний діоксид кремнію, так як відпалювання при високій
температурі призводить до втрати макропористої структури [17]. Незважаючи на
це, неушкоджені панцирі діатомових водоростей використовують як фотонні
кристали, оптичні сенсори, які мають зелену фотолюмінесценцію, і також як
темплати для створення наноматеріалів.

Рисунок 2 - Панцир діамантових водоростей
Деякі мікроорганізми здатні засвоювати сполуки
металів і потім у процесі біосинтезу накопичувати метал у вигляді структур з
суворою просторовою конфігурацією. У статті [18] описаний біосинтез кристалів
солей срібла бактеріями Pseudomonas stutzeri штаму AG259. Цей вид
бактерій мешкає на родовищах срібних руд. В процесі метаболізму між
цитоплазматичною мембраною і клітинною стінкою бактерій, в основному біля
полюсів клітини, утворюються кристали солей срібла розміром до 200 нм. Було
показано, що бактерії здатні утворювати не менше трьох різних типів кристалів з
чіткою просторовою структурою. Вчені припускають, що, змінюючи умови
культивування бактерій, можна буде синтезувати кристали із заданими
параметрами.
У дослідженні Клаус та ін. Р. Stutzeri
продукує срібло. Не відомо, чи є Ag-вмісні частинки побічним продуктом
випадкового механізму, або організм використовує їх в клітинній активності
багатофункціонально. Можливо, що цей вид бактерій використовує іони срібла, як
частину процесу переносу електронів з його відновленням до елементарного
срібла. Цікаво відзначити, що щільність срібла може привести до збільшення
щільності клітин, в яких він формується, що сприяло б їх осадження в товщі
води. Частинки срібла в розчині, зазвичай утворюють рівноважну, кубооктаедричну
морфологію на основі їх ізоморфної кристалічної структури. У P. stutzeri,
Ag-вмісні кристаліти розташовані в периплазмі, і плоска форма може бути
наслідком обмеженого простору, доступного для пластинчастої морфології. З
іншого боку, форма може бути результатом ферментативної реакції або наявності
шаблонів, які можуть впливати на кінетику росту. Ізоморфні кристаліти з
плоскою, шестигранною або трикутною формами вказують, що плата має найвищу
атомну щільность. Плоский кристаліт з високим співвідношенням сторін
(відношення периметру до товщини), наприклад, 100: 1, буде мати відношення
поверхні до об'єму, на два порядки більше, ніж рівноважне симетричної форми і,
отже, може зростати з більшою швидкістю [18].
У живих істотах зустрічаються наночастинки
магнітних матеріалів, переважно магнетиту (Fe3О4). Такі
структури, зокрема, виявлені в здатних до магнітотаксису бактерій, котрі за допомогою магнітних нанозерен, синтезованих у
процесі біомінералізації, орієнтуються у
магнітному полі Землі [19, 20]. Наприклад, бактерії Magnetospirillum magnetotacticum біомінералізують
залізо в наночастинки магнетиту - магнетосоми розміром біля 50 нм, що
дозволяють організму реагувати на магнітне поле в середовищі існування. Такі
наноструктури мають значний потенціал
використання у нанобіології, біотехнології та медицині завдяки їх вузькому
розподілу за розмірами та природній біодоступності
[21].
Вченими розроблені магнетосоми для чутливого
методу виявлення поверхневого антигену гепатиту В (HBsAg) у сироватці крові
людини із використанням магнітної імунної полімеразної ланцюгової реакції (ПЛР) [20, 21,
22]. Феромагнітні магнетосоми магнетиту, отримані з бактерії Magnetospirillum gryphiswaldense, модифікуються
олігонуклеотидами та антитілами для виявлення HBsAg. До антигену додатково
приєднується кон'югат "ДНК-антитіло" для ампліфікації ПЛР. Для даного методу спостерігається 125-кратне збільшення
чутливості порівняно з імуноферментним аналізом та 25-кратне - порівняно з
порогом виявлення HBsAg з використанням комерційно доступних магнітних кульок. Найбільш ймовірно, що
висока продуктивність магнетосом, у порівнянні зі звичайними магнітними кульками, є результатом їх нанорозміру, монодисперсності та високої намагніченості [23].
2.
Нанобіоелектроніка
2.1
Біокомп’ютери
Біокомп'ютери представляють собою гібрид
інформаційних технологій та біохімії. Дослідники з різних областей науки
(біології, фізики, хімії, генетики, інформатики) намагаються використовувати
реальні біологічні процеси для створення штучних обчислювальних схем. Існує
декілька принципово різних типів біологічних комп'ютерів, заснованих на різних
біологічних процесах: штучні нейронні ланцюги, еволюційне програмування, генні
алгоритми, ДНК-комп'ютери і клітинні комп'ютери. Перші два стали досліджуватися
ще на початку 40-х років, але досі ці дослідження, ні до чого реально
працюючому не привели. Останні три, засновані на методах генної інженерії, мають
набагато більші перспективи, але робота в цих областях почалася тільки 10 років
тому (особливо просунулися в цьому питанні Массачусетський технологічний
інститут, лабораторії Берклі, лабораторії Рокфеллера, а також Техаський
університет).
2.1.1
Елементи ДНК-комп’ютера
В більшості випадків конструктивними елементами
нанопристроїв є або молекули, вилучені з оточуючого середовища, або молекули,
спеціально синтезовані для цих пристроїв, або деякі комбінації того і іншого.
Подальшим логічним розвитком пристроїв з великої кількості незалежних елементів
стала ідея використання структур, що здатні до самоорганізації, оскільки в
рамках цієї моделі прилад конструююється не як набір незалежних молекул, а як
єдиний механізм, в якому молекули об’єднані за допомогою міжнуклеотидних
зв’язків [24].
Стабільні молкули ДНК можуть бути об’єднані в
нановузли, а вони в свою чергу в стабільні констукції - наногратки. Сучасні
дослідження направлені на побудову нанограток зі складною топологічною
структурою і розробку технологій, що дозволяє з однакових нуклеотидних
послідовностей отримувати наногратки з різною топологією. В рамках досліджень,
зв’язаних зі зміною топологічної структури послідовностей ДНК і РНК, виявлені
молекули ДНК топоізомерази, що дозволяють здійснювати такі зміни.
Форми нанограток, що неможливо здійснити без
розриву вже існуючих зв'язків. Топоізомерази - клас ферментів-ізомераз, що
впливають на топологію ДНК. Класифікація ферментів основана на механізмі їх дії
на молекулу ДНК: до І типу відносять ферменти, зо каталізують зміну топологію
ноги стану молекули за допомогою одноланцюгового розриву-зшивання
двохланцюгової ДНК, ІІ типу - здійснює двохланцюговий розрив [25]. В одній
молекулі можуть бути одночасно декілька видів топоізомераз, при чому вони мають
певний рівень специфічності: деякі типи перетворень виконують топоізомерази
певного виду. Наприклад перетворення на рис.3 може бути здійснено
топоізомеразою ІІІ Escherichia coli, але не може бути здійснено
топоізомеразою І Escherichia coli. Дослідження по вивченню ДНК-топоізомерази
представляють інтерес не тільки в задачі конструювання нанограток із згаданими
властивостями, а і як інструмент, що дозволяє здійснювати розрив між
нуклеотидами. Це може бути використано при конструюванні наноприладів на основі
принципу вільної енергії [26].

Рисунок 3 - Топологічні перетворення ДНК Escherichia
coli за допомогою топоізомерази ІІІ
2.1.2
Запам’ятовуючі пристрої на вірусній основі
Гібридні біонеорганічні наноструктури є
перспективними для застосування в майбутньому наноелектроніки і у сфері
запам’ятовуючих пристроїв. Наноструктуровані віруси є привабливими для
використання в якості шаблонів для розташування квантових точок, щоб отримати
самозібрані будівельні блоки для електронних пристроїв наступного покоління.
Поки тільки декілька видів електронних пристроїв були виготовлені з біомолекул
через відсутність переносу заряду по біомолекулярним сполукам. В роботі [27]
показаний новий ефект електронної пам'яті шляхом включення наночастинок платини
в вірус тютюнової мозаїки. Ефект пам'яті оснований на комутації провідності,
яка призводить до виникнення бістабільних станів з вкл/викл співвідношенням
більше, ніж на три порядки. Механізм цього процесу пов'язаний з захопленням
заряду в наночастинках для зберігання даних і тунелювання в стані високого
рівня провідності.
Наноструктуровані біоматеріали являють собою
ідеальну систему для використання в біологічному розпізнаванні через їх
унікальну вибірковість. Важливі досягнення були зроблені у синтезі біонаноструктур
з нанокристалів, в тому числі вірусів з білковою оболонкою, модифікованих
металами або напівпровідниковими наночастинками. Значні успіхи також в
застосуванні нанокристалічних структур на молекулах ДНК для виявлення різних
фрагментів. Однак, організація упорядкованих неорганічних наночастинок за
допомогою м'яких матеріалів як шаблонів має велике значення для побудови
електронних пристроїв з новим функціями. Такі функції були реалізовані при
забезпеченні біомолекулами процесу шунтування нанорозмірних електричних схем.
Прикладом електронного пристрою, що виготовлений безпосередньо з біоматеріалів,
є зберігання інформації в гібридній системі, що складається з вірусу тютюнової
мозаїки (ВТМ), сполученого з наночастинками платини (ВТМ-Pt). Збільшення
електропровідності в цьому ВТМ-Pt нанокомпозиті змінює його властивості і
робить його достойним кандидатом для застосування в електроніці. Такі системи
показують електричну бістабильність в залежності від станів провідності, що
контролююься напругою. Важливими особливостями цієї бістабільності є
енергонезалежність, низька робоча напруга і велике вкл/викл співвідношення. У
попередній роботі група вчених [28] показала цей тип перемикання в пристроях на
основі металевих наночастинок, пов'язаних з провідним полімером. Така поведінка
перемикання тепер поширюється на біологічні матеріали, такі як ВТМ. Розміри
цього пристрою ВТМ-Pt можуть бути зменшені, і система може бути інтегрована з
використанням методів наноматеріалів.
ВТМ - рослинний вірус з одноланцюговою РНК
позитивної полярності, що складається з 2,130 ідентичних білків оболонки. ВТМ
поміщають в групі IV відповідно до системи класифікації вірусів Балтимора, тому
що геном інкапсульований в білок оболонки ВТМ є позитивною одноцланцюговою РНК.
Позитивно-полярна РНК може виступати в якості вірусної кур'єрної РНК, в якій
сама по собі вірусна РНК є інфекційною (тобто, він може бути безпосередньо
зчитана клітиною-господарем). Жорстка віріона є 300-нм-ою трубкою з 18-нм
зовнішнього і 4-нм внутрішнього діаметрів. Хімічна і фізична стабільність ВТМ
була вивчена в різних умовах, у тому числі температури, рН і т.д., що і
доводить стійкість ВТМ.
Кон'югації наночастинок з ВТМ проводили з
використанням сполук йону платини для безелектролізного зміщення. Іони металів
здатні зв'язуватись зі специфічними функціональними групами на поверхні
вірусного білка. Наночастки платини досить рівномірно розподілені по поверхні
вірусу і мають розміри від 7 до 15 нм, з середнім розміром ~10 нм.
Складна ВТМ-Pt система володіє унікальною поведінкою
перемикання провідності, і, отже, може бути використана у першому електронному
пристрої біопам'яті. Електричний ефект перемикання спостерігається в
наночастках, включених в полімерні/органічні тонкі плівки. Ці плівки
використовують наночастинки для захвату заряду або створення
молекулярно-легованого полімеру, так що провідність плівки може бути
налаштована. Поки електронний транспорт на поверхні біоматеріалів і
наночастинок не був вивчений.
2.1.3
Клітинний біокомп’ютер
Ще одним цікавим напрямком є створення клітинних
комп'ютерів. Для цієї мети ідеально підходять бактерії, в геном яких вдалося б
включити якусь логічну схему, яка могла б активізуватися в присутності певної
речовини. Такі комп'ютери дуже дешеві у виробництві. Їм не потрібна настільки
стерильна атмосфера, як при виробництві напівпровідників. І одного разу
запрограмувавши клітину, можна легко і швидко виростити тисячі клітин з такою ж
програмою [29].
Американські вчені створили трансгенні
мікроорганізми, клітини яких можуть виконувати логічні операції І/АБО. Фахівці
лабораторії Оук-Рідж, штат Теннессі, використовували здатність генів
синтезувати той чи інший білок під впливом певної групи хімічних подразників.
Вчені змінили генетичний код бактерій Pseudomonas putida таким чином, що
їх клітини знайшли здатність виконувати прості логічні операції. Наприклад, при
виконанні операції І в клітку подаються дві речовини (по суті - вхідні
операнди), під впливом яких ген виробляє певний білок. Тепер вчені намагаються
створити на базі цих клітин більш складні логічні елементи, а також подумують
про можливість створення клітини, що виконує паралельно кілька логічних
операцій [30].
Дослідники з Університету Лідса досліджуть
природну здатність бактерій Magnetospirillum magneticum споживати
шматочки заліза атомного розміру і перетворювати їх в біти в крихітних
магнітів, що зберігаються всередині клітини. Магнітні частинки нанометрового
розміру аналогічні тим, які використовуються в сучасних жорстких дисках. Це
відкриття може привести до бактерій, які продукують електронні компоненти на
замовлення.
Бактерії роду Magnetospirillum зазвичай
живуть в неглибоких водоймах з великою кількістю розчинених елементів. За
допомогою спеціального білка, бактерії всмоктують залізо з води і формують
маленькі шматочки магнетиту, один з найбільш високооплачуваних натуральних
магнітних матеріалів. Вони використовують магніти, щоб вирівнятись з магнітним
полем Землі і орієнтуватися в просторі, що в свою чергу допомагає їм знайти
області зі сприятливим вмістом кисню [31].
Дослідники Лідсу вилучили залізо-продукуючий
білок з бактерій і нанесли його в шаховому порядку на поверхню золота. Золото,
покрите білком, занурюють у розчин зйонами заліза. При нагріванні розчину
залізо збирається на білку за тією ж схемою шаховому порядку, утворюючи багато
крихітних магнітів подібно до бактерій (рис. 4). Цей компонент може бути
основою жорсткому диску комп'ютера [31].
 - Золота основа; b - нанесений залізопродукуючий
білок; с - спродуковане залізо [31].
- Золота основа; b - нанесений залізопродукуючий
білок; с - спродуковане залізо [31].
Рисунок 4 - Збірка компонентів жорстких дисків
комп’ютера
Технологія жорстких дисків не сильно змінилася з
1960 року. Плата покрита крихітними металевими кристалами, що несуть дрібні
магнітні поля, орієнтація яких зберігає біти (1 або 0) даних. Збільшення обсягу
зберігання інформації потребує зменшення розмірів бітів, з нинішніх розмірів
близько до 10 нанометрів для кожного зерна.
Нанорозмірний комп'ютер потребує також дротів
відповідного розміру. В цій сфері значне досягнення зробив Масайоші Танака з
Токійського університету сільського господарства і технологій. Вчені побудували
дроти 40 нм в діаметрі, покриті різними білками зі штучних бактеріальних
клітин. Перша одновимірна (1D) збірка малотоксичних CuInS2/ZnS квантових точок
(КТ), вкладених в ліпідні нанотрубки, сформовані з ліпосом, представлено в його
роботі [32]. КТ-вмісні ліпідні нанотрубки володіють високим співвідношенням
сторін ≈ 500: 1 (≈ 40 нм діаметру і 20 мкм довжини) і стабільні
протягом більш ніж 20 год. Крім того, ця методика поширюється на зібрання
різних видів наночастинок всередині 1D ліпідних нанотрубок, і включає в себе
матеріали, такі як CdSe і Au (рис.5). Інкапсуляція в гідрофобній серцевині
бішару робить ці матеріали високо біосумісними [32].

а) CdSe/ZnS b) Au [32].
Рисунок 5 - Ліпідні нанотрубки із
"захопленими" наночастками
Інші вчені показали, що можна зменшити розмір
магнітних доменів жорсткого диску до атомно-масштабних розмірів. Командою
Університету штату Массачусетс виявлено, що бактерії Geobacter
sulfurreducens можуть виробляти електропровідні пілі (див. розділ 1.2) [8].
2.1.4
Модульні логічні вентилі з бактеріальних геномів
Дослідники з Університету Райса і Медичного
Центру при Університеті Канзасу (США) працюють над створенням особливих
генетичних циклів, створених з частин неспоріднених бактеріальних геномів, які
могли б виконувати складні завдання, керуючи цілими білковими блоками. Складні
генетичні логічні цикли подібні до тих, що використовуються при створенні
звичайних комп'ютерів і електронних пристроїв. У простому циклі, якщо присутні
обидва вихідних параметра, цикл відбувається згідно прописаного порядку.
Наприклад, у дослідженні [33] лондонські вчені продемонстрували, як працюють
біологічні вентилі. В одному експерименті вони показали, як біологічний
логічний вентиль обробляє інформацію тим же способом, що і його електронний
аналог, перемикаючись між станами "включено" або
"вимкнено". Було сконструйовано тип логічних вентилів названих
"І вентиль" з кишкової палички Escherichia coli. Команда
змінила E. Coli модифікувавши ДНК, запрограмувавши її для виконання того
ж самого процесу перемикання станів "включено-виключено", як і в
електронному еквіваленті, при стимуляції хімічними сполуками. Біологічні
вентилі є модульними, це означає, що вони можуть бути з'єднані разом для
отримання різних типів логічних вентилів, даючи можливість в майбутньому
створювати більш складні біологічні процесори. В іншому експерименті,
дослідники створили "НЕ вентиль" і з'єднали його з "І
вентилем" для отримання більш складного "І-НЕ вентиля", використовуючи
систему для ІІІ типу секреції Pseudomonas syringae [33].
Завдяки тому, що генетичні схеми засновані за
типом бінарної логіки, генетичний логічний цикл може ініціювати створення
специфічного білка при наявності двох хімічних сполук, або змусити ДНК клітини
придушити створення цього білка. З розвитком відповідних інструментів для
синтетичної біології прості цикли стало створювати простіше, але для вирішення
складних проблем потрібні комплексні інструменти. Основою найсучаснішого [34]
досліду є генетичний арсенал химерних (тих, що складаються з різних частин)
транскрипційних факторів - білків, контролюючих транскрипцію генів шляхом
зв'язування зі специфічними ділянками ДНК. Химерні транскрипційні фактори
об'єднують в собі генорегулюючу здатність одного транскрипційного фактора і чутливість
іншого. Дослідники продемонстрували, що чотири химери з однаковими
ДНК-зв'язуючими модулями можуть спільно працювати і служити
"воротами" з безліччю вихідних параметрів, як пригнічуючи, так і
знімаючи пригнічення специфічних генів. Дослідники успішно протестували химерні
комбінації в кишковій паличці (Escherichia coli) на здатність до ап - і
даун-регуляції гена, що кодує зелений флуоресцентний білок.
Такі системи дозволять отримувати синтетичні
клітини для вироблення біопалива, відновлення навколишнього середовища або
лікування захворювань людини. Модель взаємодії описаних біологічних вентилів
впритул наближається до моделі побудови нашого мозку, що закладає основи для
розробки штучного інтелекту.
2.2
Мікробні клітинні біосенсори
Серед численних видів біосенсорів, мікробні
цільноклітинні біосенсори мають численні переваги. Культивування
мікроорганізмів дешевше порівняно з культурами клітин вищих організмів. Вони
можуть бути отримані у великих кількостях, піддані більш суворому контролю, ніж
клітини вищих організмів, і легко зберігаються. При цьому, цільноклітинні
біосенсори зазвичай трансформовані рекомбінантними плазмідами і вони мають в
складі цих плазмід промотор, що відповідає на вплив речовиною-подразником
транскрипцією репортерного гена, що знаходиться під контролем цього промотора,
і дають в результаті трансляції репортерний білок, який легко виміряти за
допомогою флуориметра, люмінометра або кольорової реакції. Переваги
використання цільноклітинних біосенсорів пов'язані з їх високою специфічністю,
селективністю і швидкістю дії. Принциповою особливістю їх використання є
здатність оцінити не тільки біодоступність аналіту, а й фізіологічне значення
отриманих даних.
Цільноклітинні бактеріальні біосенсори, що
з'явилися в останні десятиліття, преважно спрямовані на визначення
екотоксикантів в об'єктах довкілля. Вони є незамінними методами екологічних
досліджень за рахунок ряду переваг:
дозволяють визначати тільки біодоступність для
живої клітини компоненти, даючи об'єктивну відповідь щодо токсичної або
мутагенної дії екотоксикантів. Це дозволяє вжити заходи для біоремедіації
навколишнього середовища. Якщо аналіт біодоступний, він потенційно
біодеградуємий.
дають в руки екологів та інших дослідників
експресний, дешеввй і доступний засіб попередньої оцінки дії екотоксикантів на
живу клітину.
використання генетично модифікованих
мікроорганізмів, здатних існувати в навколишньому середовищі і піддавати
екотоксиканти деградації, дає екологам перспективи оцінки віддалених наслідків
впливу аналіту на організм людини і тварин.
дають можливість оцінити канцерогенний потенціал
пестицидів, антибіотиків і хімічних сполук на живий організм.
представляють засіб наукового вивчення
транскрипціонної регуляції різних ферментних систем, що беруть участь у
відповіді на екотоксиканти.
Безумовно, застосування біосенсорів в наукових
дослідженнях також розширюється у зв'язку з можливістю їх використання при
вивченні мутагенезу, генотоксичної дії пестицидів, металів, органічних сполук,
маркерів вірусної інфекції та канцерогенезу. Застосування в біосенсорах
флуорисцентих репортерних білків (зеленого флюорисцентного білка і його
варіантів з унікальними флуоресцентними властивостями, люциферази,
галактозидази) дозволило їх використання в цитології, цитохімії, як
мультизабарвлюючі барвники, в дослідженнях на тваринах і рослинах in situ,
вивчати введення генів репортерних білків в геном вищих тварин. Крім того,
використання цільноклітинних біосенсорів дозволило розширити сферу застосування
засобів реєстрації сигналів з клітки за допомогою приладових методів:
світловолоконної оптики, поверхневого плазмонного резонансу, кондуктометрії,
різних середовищ (гелі, плівки). З'явилися можливості сорбції клітин на
поверхні сенсорів волоконно-оптичного характеру, пористих скляних кульок,
діалізних мембранах, плівках, гелях, що дозволяє підвищити точність визначення
аналіту [35, 36].
2.2.1
Біосенсори на основі генетично модифікованих бактеріальних клітин
Генетично модифіковані цільноклітинні біосенсори
реагують на наявність аналіту шляхом індукції гена, пов'язаного з чутливим до
аналіту промотором шляхом створення гібридного (фьюз) гена. Зазвичай такі
гібридні гени існують в генетично трансформованих бактеріальних клітинах у
складі плазмід, якими трансформують клітини. Структурна частина генетичної
конструкції кодує вимірюваний аналітичним способом репортерний білок. Промотор,
що зливається зі структурним геном, звичайно являє собою регуляторний
компонент, селективний та індукований аналітом.
Біодоступний аналіт проходить через мембрану
клітини і взаємодіє з промотором гена-репортера безпосередньо або шляхом
інактивації його репрессора. Індукований ген транскрибується. Трансляція мРНК
репортерного гена призводить до синтезу білка, який легко реєструється.
Транскрипційні конструкції, створені шляхом злиття фьюз гена-репортера і
промотора є потужним знаряддям для вивчення експресії бактеріальних генів в
природному середовищі і для детекції певних сполук у навколишньому середовищі.
Хоча транскрипційні конструкції, створені для цієї мети, часто дуже слабо
експресуються, і їх індукція може відбуватися протягом тривалого часу, а
екотоксикант може надійти до біосенсорів в обмежений проміжок часу,
необхідність визначати експресію гена-репортера, існує [37].
Для того, щоб синтез репортерного білка відбувся,
необхідно створити генно-інженерними методами плазміду з клонованим в неї
"геном-репортером" під управління промотора, який відповідає на
досліджуваний аналіт. Кілька умов необхідно, щоб використовувати таку
біосенсорну конструкцію. По-перше, генетична конструкція, що відповідає за
синтез репортерного білка, повинна бути відносно простою, по-друге,
специфічність чутливого елемента (промотора) по відношенню до аналіту -
високою, і, нарешті, в клітині повинні бути відсутніми подібні протеїни або
субстрати, які можуть перешкодити визначенню даного репортерного білка.
Зазвичай промотори і енхансери розташовують у цис-положенні по відношенню до
структурного цільового гену. Дослідження на клітинних культурах і трансгенних
тваринах дозволили відібрати репортерні гени і транскрипційні елементи,
відповідальні за тканинно-специфічну експресію генів, зокрема тих, які можуть
брати участь у відповіді на захворювання. Було показано, що вплив на деякі гени
необхідний для лікування спадкових, вірусних, бактеріальних, запальних,
серцево-судинних і ракових захворювань.
Роботи на клітинних культурах і трансгенних
тваринах і рослинах дозволили відібрати репортерного гени, відповідальні за
базальний і тканинно-специфічний рівень синтезу білків клітини. Ідентифікація
деяких репортерних генів (люціферази світляків і зеленого флуорисцентного
білка), які можуть використовуватися як неінвазивні маркери генетичної
експресії, дозволяє в режимі реального часу (наприклад, за допомогою
конфокальної спектороскопіі) стежити за експресією репортерних і регулюючих
метаболізм клітини білків. Так, можна оцінити вплив різних транскрипційних
факторів і медіаторних механізмів клітини для лікування ряду захворювань,
включаючи рак, вірусні, запальні інфекції, серцево-судинні захворювання.
Цільноклітинні біосенсори і репортерні гени в
даний час широко використовуються для виявлення різних за хімічним складом і
структурою сполук. Так, наприклад, є біосенсори, за допомогою яких можна
оцінити наявність в середовищі металів. Можна визначити цілий ряд органічних сполук,
пестицидів, мутагенів, генотоксикантів.
У результаті досліджень останнього десятиліття
були відібрані репортерного білки, найбільш зручні для реєстрації. В даний час
в біосенсорах використовуються вісім унікальних репортерних білків: зелений
флуоресціюючий білок (GFP) і його мутантні похідні (наприклад, CFP -
ціан-флюорисцентний білок, YFP - жовтий флююорисцетний білок), β - галактозидаза, хлорамфеніколацетілтрансфераза,
бактеріальна люціферази і люціферази світляків, акворін, уропорфіроген III метилтрансфераза,
червоний флуоресціюючий білок коралів (DsRed).
Для прикладу розглянмо коло проблем, що виникає у
зв'язку з ракетно-космічною діяльністю. Найбільшу небезпеку для рослинного і
тваринного світу являє падіння перших ступенів ракет-носіїв, в паливних
магістралях яких знаходиться досить велика кількість палива й окислювача.
Основною речовиною в якості пального в рідинних ракетних двигунах ракет-носіїв
є несиметричний диметилгидразин (1,1-ДМГ), що відноситься до першого класу
токсичності і має загальнотоксичну і місцево-подразнюючу дію, а також виявляє
канцерогенні властивості. Проте порівняно небагато відомо про мутагенну і
генотоксичну дію 1,1-ДМГ, що може бути одним з ключових ланок дії на ДНК людини
і тварин. Особливу небезпеку становить накопичення продуктів трансформації
1,1-ДМГ, таких як нітрозодіметіламін (НДМА), тетраметілтетразен (ТМТ),
1-метил-1Н-1,2,4-триазол, 1,2,4,5-тетразин, 2, 3,5,6-тетрагідро-1,4-диметил та
інші.
Фізико-хімічні методи, що існують для вимірювання
рівня гідразинів в організмі людини і навколишньому середовищі, а також
дозволяють оцінити мутагенний ефект цих сполук вимагають заскладного обладнання
і не придатні для рутинного аналізу. Крім того, 1,1-ДМГ вступає в реакції
комплексоутворення з органічними сполуками, білками, ДНК і його визначення
ускладнюється [37].
Постановка досліджень з визначення гідразину та
продуктів його трансформації можлива тільки при розробці біологічних методів
визначення компонентів ракетного палива, що забезпечують проведення
експерименту в лабораторних і польових дослідженнях. Тому метод оцінки
генотоксичних і токсичних властивостей 1,1-диметилгидразину на основі
спеціальної біосенсорної технології із залученням останніх досягнень генної
інженерії та відповідних апаратурних засобів знайшов широке застосування в
екологічних дослідженнях. Одним з підходів у вирішенні цього завдання може
служити розробка застосування рекомбінантних репортерних генів, що кодують
біолюмінесцентні білки.
2.2.2
Люмінесцентні біосенсори
Бактеріальні люмінісцентні біосенсори є поширеним
інструментом оцінки якості питних, поверхневих, грунтових і стічних вод. В
основу їх практичного використання покладено аналіз активності люмінесцентної
системи, що перебуває на перетині основних енергетичних потоків мікробної
клітини і тому інтегрально відповідає на всю сукупність присутніх у середовищі
поллютантів [38]. Біолюмінісцентний аналіз не дозволяє оцінити природу
забруднення, але дає можливість отримати комплексне уявлення про ступінь його
біологічної небезпеки, яка характеризується поняттям "біотоксичність"
При вивченні ефекту основних солей, що входять до
складу мінеральних вод, в дослідженнях [39, 40] продемонстровані особливості
реагування бактеріальних біосенсорів в присутності різних катіонів та аніонів.
При цьому для катіонів показаний двофазний (індукція/інгібування) дозозалежний
ефект, в рамках якого за здатністю до придушення світіння вони формували ряд Са2+
> Na+ > Mg2+ > К+. Встановлено, що
абсолютні значення концентрацій солей, що викликають стимуляцію або пригнічення
світіння, залежать від екологічних особливостей використовуваних люмінісцуючих
мікроорганізмів і виявляються вищими для природного морського штаму Photobacterium
phosphoreum порівняно з рекомбінантним штамом Escherichia coli з
клонованим опероном Photobacterium leiognathi ("Еколюм-9"). На
цьому тлі ефекти аніонів, що також впливають на інтенсивність бактеріальної
біолюмінесценції різноспрямовані дозозалежні ефекти, визначаються їх хімічною
природою і реалізуються через взаємодію з ферментною системою генерації
світіння (галогеніди), вплив на розчинність солей (сульфати) або зміна рівня рН
середовища (карбонати і гідрокарбонати) [39].
Отримані результати дозволили обґрунтувати
підходи до проведення біотестування мінеральних вод з використанням
люминесцирующих мікроорганізмів, що виключають вплив їх нормального
компонентного складу на результати дослідження. Запропонована пробопідготовка
включає послідовну дегазацію, диференційовану (залежну від вихідного сольового
складу) мінералізацію і нормалізацію рН досліджуваних вод, що дозволяє
відновити рівень світіння бактеріальних біосенсорів із збереженням їх
чутливості до дії хімічних полютантів.
На даній основі розроблено та впроваджено
методичні рекомендації "Методика експресного визначення токсичності питних
бутильованих мінеральних вод за допомогою люмінесцентних бактеріальних
біосенсорів" (затверджені 27.08.2007 р), призначені для використання
фахівцями "Центру гігієни і епідеміології" і
"Роспотребнадзора", що є реальним прикладом практичного використання
даного виду біосенсорів.
2.2.3
Системи ферментів мікроорганізмів в якості промоторів
У системі біосенсорного моніторингу особливе
місце займає контроль забруднення навколишнього середовища. Хоча існуючі методи
фізико-хімічного аналізу дозволяють надійно і з високою точністю визначати
якісний і кількісний склад цілого ряду екотоксикантів, вони не завжди можуть
оцінити їх біодоступність і небезпеку для живих організмів. Відомо, що в
грунтових екосистемах рухливість, біодоступність і, отже, токсичність хімічних
сполук істотно залежить від форм їх знаходження і типу зв'язку з грунтом. При
цьому в ряді випадків, аналіз екотоксикантів ускладнений необхідністю очищення
екстрактів з грунтів перед інструментальним визначенням досліджуваних
компонентів, що є досить трудомістким процесом, часто призводить до помилок в
результатах аналізу. У зв'язку з цим, велика увага приділяється експресним
методам аналізу, орієнтованим на швидкий попередній контроль небезпечного рівня
забруднення або превалюючого токсичного ефекту. Поставлені завдання можуть бути
досягнуті, насамперед, шляхом поєднання фізико-хімічних методів і сучасної
біосенсорной технології.
Як правило, іони металів потрапляють в клітину
неспецифічним шляхом, в результаті дифузії по осмотичного градієнту,
створюваному мембраною клітини. Однак ряд металів входить в клітку більш
повільно, специфічно, через певні транспортні канали і з витратою енергії АТФ.
Щоб вижити в навколишньому середовищі, мікроорганізми виробили механізми
стійкості до токсичного впливу металів. Так, може відбуватися зв'язування
металів в цитоплазмі клітини з білками - металотіонеїнами, зв'язування іонів
металів на зовнішній стороні мембрани клітини, відновлення іонів металів, щоб
привести їх у менш токсичну форму, активний транспорт іонів металів з клітини.
Можна модифікувати іони металів, щоб перевести їх в нерозчинну або менш
токсичну форму.
Механізми стійкості мікроорганізмів до дії
металів визначаються опероном, присутніми на плазмідах, транспозонах або
хромосомах клітини. У багатьох випадках, такі оперони кодують ферменти, що
забезпечують стійкість до близьких по біохімічному характеру дії металів.
Попадання іонів металів в клітини індукує синтез ферментів, необхідних для
забезпечення резистентності клітин до токсичної дії. Спираючись на існування механізмів
резистентності, були розроблені численні специфічні тест-системи для оцінки
вмісту іонів у навколишньому середовищі, наприклад, ртуті, хрому, кадмію, міді,
алюмінію, сурми, нікелю, миш'яку.
Органічні сполуки є головними джерелами
забруднення довкілля і вони стали основною мішенню для розвитку цільноклітинних
біосенсорів. Багато бактеріальних видів здатні використовувати органічні
сполуки як джерело вуглецю. Отже, ці мікроорганізми мають ферментні системи, що
дозволяють метаболізувати органічні сполуки, і вони наразі використовуються для
ліквідації наслідків забруднення навколишнього середовища нафтою та іншими
вуглеводнями. Біодоступність екотоксикантів, чутливість і селективність
розроблюваних тест-систем, механізми регуляції, активності та експресії
необхідних для деструкції екотоксикантів ферментів, є основою розроблюваних
тест-систем. Такі системи повинні включати промотор з системи ферментів,
регулюючої розщеплення органіки, і репортерний білок. Створені з цією метою
клітинні біосенсори виявилися експресними і набули широкої популярності.
Цільноклітинні бактеріальні генетично
модифіковані біосенсори, являють собою експресний, недорогий і чутливий засіб
для вивчення екологічного стану навколишнього середовища. Сучасні тест-системи
дозволяють проводити визначення зазвичай без екстракції ферментних і
репортерних білків з клітки, що дає вагомі переваги для роботи в польових
умовах на "живий" системі. В даний час застосування цільноклітинних
біосенсорів розширюється в не тільки в галузі біотехнології, але і в медицині
при діагностиці, фармакології, харчовій та хімічній промисловості, в наукових
дослідженнях при вивченні регуляції транскрипції генів, у функціональній
геноміки, мікробної екології in situ. За допомогою цільноклітинних
біосенсорів можна оцінити токсичність металів, загальну токсичність і
генотоксичность, мутагенний і канцерогенний потенціал антибіотиків, багатьох
лікарських і знову синтезованих органічних і неорганічних сполук. Можна вивчати
метаболізм цілих організмів і мікроорганізмів, долю препаратів, що
застосовуються для захисту рослин від шкідників, пестицидів і фунгіцидів.
Модифікації промоторів мікроорганізмів і інші генетичні маніпуляції, дозволяють
збільшити чутливість визначення аналітів. Хоча слід зазначити, що застосування
природних, чи не модифікованих промоторів, дозволяє оцінити вплив
екотоксікантов найбільш адекватно [37, 41].
Висновок
У ході виконання курсової роботи були розглянуті
нанотехнології з використанням мікроорганізмів та продуктів їх життєдіяльності.
Можна зробити певні висновки про значення даних технологій, рівень їх розвитку
на даний момент та перспективи подальшого дослідження.
Визначили, що мікроорганізми в біонанотехнології
використовуються переважно в двох напрямках: виготовлення наноматеріалів і
здійснення певних логічних операцій в якості нанокомп’ютерів, наносенсорів і їх
складових. Використання живих об’єктів мікросвіту зумовлене рядом переваг перед
класичною нанотехнологією.
Властивості матеріалів, отриманих за допомогою
мікроорганізмів, перевершують аналогічні синтетично вироблені матеріали:
отримані структури високо організовані у нано-, мікро- і макромасштабах, часто
в ієрархічному порядку, зі складною наноархітектурою, і в кінцевому підсумку
складають безліч матеріалів із широким спектром властивостей. Вони одночасно є
''розумною'', динамічною, складною, здатною до самовідновлення і
багатофункціональною системою. На відміну від штучних засобів продукування
наноматеріалів, використання мікроорганізмів позбавляє проблеми енергетичного
забезпечення процесу, а велика швидкість розмноження організмів збільшує його
продуктивність.
Біологічні нанокомп'ютери дуже дешеві у
виробництві, не потребують настільки стерильної атмосфери, як при виробництві
напівпровідників. Особливою перевагою біонаномашин є біосумісність, можливість
активно брати участь в біологічних сигнальних каскадах. Вони створені для
роботи в специфічному середовищі і позбавляють проблеми пристосування механізму
до жорстких біохімічних і біофізичних умов. Переваги використання
цільноклітинних біосенсорів пов'язані з їх високою специфічністю, селективністю
і швидкістю дії. Принциповою особливістю їх використання є здатність оцінити не
тільки біодоступність аналіту, а й фізіологічне значення отриманих даних.
"Лідерами” серед об’єктів мікробіології в нанотехнології
є вірус тютюнової мозаїки, бактеріофаг М13 та E. coli.
Проте вищезазначені переваги здебільшого
компенсуються занадто складним технологічним процесом, неможливістю
використання за межами лабораторії, економічною невигідністю і обмеженістю набору
процесів. З огляду на опрацьовані результати досліджень можемо зробити
висновок, що біонанотехнологія з використанням мікроорганізмів потребує
подальшого розвитку для розробки нових методів і оптимізації існуючих.
Список
літератури
1. Fischlechner M. Donath E. Viruses as building blocks for
materials and devices // Angew. Chem. Int. Ed. Engl. - 2007. - Т.46, №18. -
С.3184-3193.
. E. Dujardin, C. Peet, G. Stubbs Organization of metallic
nanoparticles using tobacco mosaic virus templates // Nano Lett. - 2003. - Т.3,
№3. - С.413-417.
. M. Knez, A. M. Bittner, F. Boes Biotemplate synthesis of 3-nm
nickel and cobalt nanowires // Nano Lett. - 2003. - Т.3, №8. - С.1079 - 1082.
. M13 Virus-Enabled Synthesis of Titanium Dioxide Nanowires for
Tunable Mesoporous Semiconducting Networks / Po-Yen Chen, Xiangnan Dang,
Matthew T., Courchesne N. D., Jifa Qi, Md Nasim Hyder, Belcher A. M., Hammond
P. T. // Chem. Mater., - 2015. - T.27, №5. - С.1531-1540.
5. UIC
News Center [Електронний ресурс]: Tiny bio-robot is a germ suited-up with
graphene quantum dots // Режим доступу: http://news.
uic.edu/tiny-bio-robot-is-a-germ-suited-up-with-graphene-quantum-dots
<http://news.uic.edu/tiny-bio-robot-is-a-germ-suited-up-with-graphene-quantum-dots>.
.
S-layers: principles and applications / U. B. Sleytr, B. Schuster, E. Egelseer,
D. Pum // FEMS Microbiology Reviews. - 2014. - Т.38,№5. - С.823-864. 2009. -
T.168, №1. - 207-216.
.
Absorption, steady-state fluorescence, fluorescence lifetime, and 2D
self-assembly properties of engineered fluorescent S-layer fusion proteins of
Geobacillus stearothermophilus / Kainz B., Steiner K., Moller M., Pum D., Schäffer C., Sleytr U. B., Toca-Herrera J. L. // Biomacromolecules. -
Т.11. - С. 207-214.
.
Geobacter sulfurreducens conductive pili as biological nanowires and uses
thereof [Текст]: патент UMA 05-09/Derek R. Lovley; власник патенту University
of Massachusetts, Amherst. - US Patent №7,498,155; опубл.3.03.09, - 6 с.
. Способ
очистки теплотехнического оборудования от накипи [Текст]: патент / Богомолов
В.Н., Звенигородский В. П.; власник патенту Богомолов В.Н. - РФ № 2213922;
опубл.10.10.2003, - 2с.
.
Sarikaya М. Biomimetics: Materials fabrication through biology. PNAS. - 1999. -
Т.96, №25. - С.14183-14185.
.
Synthesis of yeast-assisted Co3O4 hollow microspheres-A
novel biotemplating technique / Li Yang, W. Guan, Bo Bai, Qing Xu, Yun Xiang //
Journal of Alloys and Compounds. - 2010. - Т.504, №L. - C.10-13.
.
Virus-based toolkit for the directed synthesis of magnetic and semiconducting
nanowires / Mao C., Solis D.J., Reis B.D., Kottmann S.T., Sweeney R.Y.,
Hayhurst A., Georgiou G., Iverson B., Belcher A.M. // Science. - 2004. - Т.213,
№7. - С.5655.
.
Virus-enabled synthesis and assembly of nanowires for lithium ion battery
electrodes / Nam K.T <http://www.ncbi.nlm.nih.gov/pubmed/?term=Nam%20KT%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Kim D.W
<http://www.ncbi.nlm.nih.gov/pubmed/?term=Kim%20DW%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Yoo P.J <http://www.ncbi.nlm.nih.gov/pubmed/?term=Yoo%20PJ%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Chiang C.Y
<http://www.ncbi.nlm.nih.gov/pubmed/?term=Chiang%20CY%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Meethong N <http://www.ncbi.nlm.nih.gov/pubmed/?term=Meethong%20N%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Hammond P.T
<http://www.ncbi.nlm.nih.gov/pubmed/?term=Hammond%20PT%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Chiang Y.M
<http://www.ncbi.nlm.nih.gov/pubmed/?term=Chiang%20YM%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.,
Belcher A.M
<http://www.ncbi.nlm.nih.gov/pubmed/?term=Belcher%20AM%5BAuthor%5D&cauthor=true&cauthor_uid=16601154>.
// Science. - 2006. - T.885, №8. - С.5775.
.
Ordering of quantum dots using genetically engineered viruses / Lee S.W., Mao
C., Flynn C.E., Belcher A.M. // Science. - 2002. - Т.296, №5. - С.5569.
.
Галаганова Е.Н. Методы получения наноматериалов из микроорганизмов и их
применение // Электронный журнал: Молодежный научно-технический вестник. -
2013. - Т.68.
.
Беззубов С.И., Воробьева Н.А., Ефимов А.А. Мать - природа в помощь // Нанометр.
- 2010. - T.54, №2. - С.121-134.
.
Two-stage photobioreactor process for the metabolic insertion of nanostructured
germanium into the silica microstructure of the diatom Pinnularia sp. / C.
Jeffryes, T. Gutu, Jun Jiao, G. L. Rorrer // Materials Science and Engineering.
- 2008. - Т.28. - С.107-118.
.
Silver-based crystalline nanoparticles, microbially fabricated / T. Klaus, R.
Joerger, E. Olsson, C. - G. Granqvist // PNAS. - 1999. - Т. 1986, №77. -
С.5331-5345.
.
Guimaraez A.P. Principles of nanomagnetism / Guimaraez A.P. - Berlin,
Heidelberg: Springer, 2009. - 221с.
.
Нанобіологія, біоміметика та природні наноструктури / Чекман І.С., Сімонов
I.В., Нагорна О.О., Шаторна В.Ф. // Нанонаука, нанобіологія, нанофармація - К.:
Полишграф плюс, 2012. - С.160-173.
.
Controlled cobalt doping of magnetosomes in vivo / S. Staniland, W. Williams,
N. Telling // Nat. Nanotechnol. - 2008. - Т.3, №3. - С.158-162.
.
Niemeyer C. M. Immuno-PCR: high sensitivity detection of proteins by nucleic
acid amplification / C. M. Niemeyer, M. Adler, R. Wacker // Trends Biotechnol.
- 2005. - Т.23, №4. - С. 208-216.
. Seeman
N. C. Nucleic Aacid Junctions and Lattices // Journal of Theoretical Biology. -
2003. - T.99. - C.237-247.
.
Бронштейн И.Б. Молекулярная биология. - 1999. - Т.23. - С.1553-1557.
.
Topological transformations of synthetic DNA knots // Biochemistry. - 1995. -
T.34. - C.673-682.
. Digital
memory device based on tobacco mosaic virus conjugated with nanoparticles / R.
J. Tseng, C. Tsai, Liping Ma, J. Ouyang, C. S. Ozkan, Yang Yang // Nature
Nanotechnology. - 2006. - T.1. - C.72 - 77.
. Tseng
R. J. Polyaniline nanofiber/gold nanoparticle nonvolatile memory // Nano Lett.
- 2005. - Т.5. - С.1077-1080.
.
Третьяков Ю.Д. Нанотехнология. Азбука для всех // М.: Физматлит. - 2009. -
368с.
.
Silva-Rocha R., Lorenzo V. Mining logic gates in prokaryotic transcriptional
regulation networks // FEBS letters. - 2008. - T.582, №8. - С.1237-1244.
.
Biotemplated Magnetic Nanoparticle Arrays / J. M. Galloway, J. P. Bramble, A.
E. Rawlings, G. Burnell, S.D. Evans, S.S. Staniland // Small7 - T.8, №2. - С.
204-208.
.
Fabrication of Quantum Dots Embedded Lipid Tubules by Membrane Tubulation
Protein / Tanaka M., Critchley K., Matsunaga T., Evans S., Staniland S. //
Small. - 2012. - T.8, №10. - С.1590-1595.
.
Engineering modular and orthogonal genetic logic gates for robust digital-like
synthetic biology / B. Wang, R. Kitney, N. Joly, M. Buck // Nature
Communications. - T.508, №2. - 2011. - С.1516-1537
.
Modular, Multi-Input Transcriptional Logic Gating with Orthogonal LacI/GalR
Family Chimeras / D. L. Shis, F. Hussain, S. Meinhardt, L. Swint-Kruse, M. R.
Bennett // Synth. Biol. - T.9, №3. - 2014. - С.645-651
. Горленко
В.А., Кутузова Н.М., Пятунина С.К. Научные основы биотехнологии. Нанотехнологии
в биологии. - М.: Наука, 2013. - 264 c
. G. K.
Knopf, A. S. Bassi Smart Biosensor Technology. - CRC Press, 2006. - 664с.
.
Лавриненко И.А., Лавриненко В.А. Бактериальные биосенсоры и их применение в
экологическом мониторинге. - Новосибирск, 2009. - 132с.
. Каримов
И.Ф. Использование люминесцирующих бактерий при оценке фагоцитарный активности
нейтрофилов: Автореф. дис. канд. биол. наук: 03.00.07; 14.00.36. - Пермь, 2009.
- 17с.
. Алешина
Е.С. Использование люминесцирующих микроорганизмов для биотестирования
минеральных вод: Дис. канд. биол. наук: 03.00.07. - Оренбург, 2008. - 182с.
.
Использование бактериальной биолюминесценции для тестирования токсичности воды
при ее очистке от ПАВ / Кацев А.М., Стародуб Н.Ф., Левковец И.А., Гончарук
В.В., Клименко Н.А., Вакуленко В.Ф. // Химия и технология воды - 2003. - Т.25,
№ 6 - С.594 - 602.
.
Левковець І.А. Розробка інструментальних аналітичних біотестів та вивчення
їхніх основних характеристик при визначенні токсичності об’єктів довкілля:
Автореф. дис. на здобуття наук. ст. канд. біол. наук: 03.00.20. - Київ, 2005. -
18с.