Растительность болот
Введение
Болото представляет собой экосистему,
состоящую из трех основных компонентов: воды, специфической болотной
растительности и торфа. Поэтому болото является предметом внимания нескольких
самостоятельных направлений, таких как, например ботаники и геоботаники,
гидрологии, почвоведения и многих других. Болота выполняют разнообразные
функции, одна из которых заключается в сохранении биологического разнообразия.
Для каждого типа болота характерна своя растительность, которая принимает
участие в образовании разного по химическому составу торфа (Тюремнов, 1976).
Особый интерес для исследований представляют олиготрофные болота, так как они
занимают наибольшие территории Западной Сибири, отличающейся высокой
заболоченностью (Лисс и др., 2001). Специфические особенности болотного
субстрата как почвы создают среду, к которой смогли приспособиться очень не
многие виды растений. Растительность болот - это мощный фактор формирования
биоценоза болот. Растительность болот является источником ценных пищевых,
кормовых и лекарственных ресурсов (Лисс, Астахова, 1982). Сфагновые мхи служат
гигантскими естественными фильтрами, поглощающими токсичные элементы.
Растительный покров служит хорошим индикатором гидрологических особенностей
болот и, прежде всего, указывает на современные условия водно-минерального
питания болот. Некоторые фитоценозы олиготрофного типа могут быть использованы
в качестве индикатора строения и свойств торфяной залежи (Лисс и др., 2001). В
целом, изучение растительности необходимо для организации рационального
использования растений, получения на их основе новых продуктов питания,
лекарств, строительных материалов, а также в целях сохранения генофонда для
работы в сфере селекции. Несмотря на то, что исследование условий
заболачивания, растительного покрова, флоры болот проводится уже более ста лет
остается еще много не решенных вопросов в области изучения растительности болот
и ее классификации (Кузнецов, 2007).
Цель курсовой работы - изучить характерные особенности
олиготрофных болот и биоморфологические признаки растений в связи с
экологическими условиями олиготрофного болота.
Задачи курсовой работы:
. Изучить особенности образования олиготрофных болот.
. Выявить общие характерные черты в строении растений
олиготрофных болот.
3. Познакомиться с основными фитоценозами олиготрофного типа.
олиготрофный болото экологический фитоценоз
1. Олиготрофные болота и их образование
Существуют различные опеределения понятия «болото». Болото -
географический ландшафт, формирующийся под действием совокупности факторов
среды и в частности растительности, что определяет периодическую или постоянную
обводнённость почвенного слоя, чем обуславливается гидрофильность
флористического состава болот, болотный тип почвообразования и процесс
аккумуляции торфа (Пьявченко, 1963). Болото - это участок суши,
характеризующийся избыточным увлажнением, повышенной кислотностью и низкой
плодородностью почв, выходом на поверхность сточных и проточных грунтовых вод,
но без постоянного слоя воды на поверхности (Вомперский, 1994). Типы болот и
аккумулирующиеся в них торфа определяют по эколого-фитоценотическому принципу,
основывающемуся на составе флористического состава болот и присутствию в торфе
растительных остатков:
. Верховые болота или олиготрофные болота, которые образуются
на водоразделах и верхних террасах речных долин;
. Низинные или эвтрофные болота, располагающиеся чаще всего в
долинах рек, озерных котловинах, различных мелких депрессиях всех зон;
. Мезотрофные болота - переходная форма между верховыми и
низинными болотами (Инишева, Моторин, 2000).
Данная классификация является общепринятой. Но Н.И. Пьявченко
(1963) выделяет четвёртый тип - разнотипный (гетеротрофный, комплексный),
состоящий из сочетания пониженных и повышенных участков различного питания.
Другие учёные классифицируют типы болот по растительному покрову (экологический
аспект), по широкому ряду признаков в географическом аспекте (Абрамова и др.,
1974). Отличия этих типов болот друг от друга состоит в особенностях
водно-минерального питания произрастающих на них растений и условиях
заболачивания территорий. Верховые (олиготрофные) болота формируются в условиях
обильного увлажнения и бедного водно-минерального питания. Это питание
осуществляется в основном за счет атмосферных осадков, которые бедны
минеральными веществами. В низинные болота поступают грунтовые и поверхностные
воды, которые в отличие от верховых болот содержат большое количество
питательных элементов, что наделяет такие болота высоким потенциальным
плодородием, переходные болота формируются в условиях смешанного питания:
атмосферными, поверхностно-сточными и частично грунтовыми водами (Абрамова и
др., 1974). Болота образуются разным путем: одни болота появились на месте
ранее существующих водоемов, в процессе заторфовывания водоемов, а другие же
возникли непосредственно на минеральном грунте, то есть в процессе
заболачивания суши. Олиготрофные болота формируются преимущественно на
водоразделах, при заболачивании лесных массивов, местах их вырубок и пожарищ,
зарастании и заторфовывании водоемов (преимущественно непроточных озёр)
(Инишева, 2009). Рассмотрим заболачивание суши (рис. 1.).


Рис. 1. Динамика уровня грунтовых вод по мере нарастания
торфа в различных условиях увлажнения исходного субстрата: 1 - минеральный
горизонт; 2 - поверхность торфяника; 3 - уровень грунтовых вод; А - суходольное
заболачивание, грунтовые воды у поверхности; Б - заболачивание водоемов;
В-суходольное заболачивание, грунтовые воды в толще минерального горизонта
(Лисс, Астахова, 1982)
Из-за почвообразовательного процесса под лесами с
доминированием хвойных пород с большой частотой формируются труднопроницаемые
для воды горизонты, проникновение воды через них практически прекращается,
отчего верхние горизонты почвы переувлажняются, отчего вода застаивается.
Первый признак застойной воды - обильный рост мха кукушкин лен (Polytrichum commune), его заросли хорошо
задерживают дождевую воду, отчего осложняется поступление кислорода в почву,
таким образом формируются анаэробные условия. В таких условия процесс
разложения растительных остатков замедляется, отчего в дальнейшем формируется
масса полуразложившихся, либо неразложившихся остатков частей растений. По мере
увеличения увлажнения зеленых мхи сменяются сфагновыми, происходит ухудшение
роста деревьев, затем смена растительности, и начинается накопление торфа. При
улучшении аэрации лишь в кратковременные периоды пересыхания торфа, тогда
процесс разложения усиливается из-за активизации деятельности микроорганизмов и
беспозвоночных. С геологической точки зрения торф является горючим
полезным ископаемым, является первой стадией метаморфоза отмерших растений,
подвергшихся неполному разложению, в дальнейшем превращается в каменный уголь.
Торф аккумулируется на дне болот при высокой влажности с ограниченным доступом
кислорода (Геологический словарь, 1978). Торфяная залежь - закономерное
вертикальное напластование торфов отдельных видов от поверхности до
минерального дна торфяного месторождения или подстилающих озёрных отложений.
Отличаются от других органических отложений земной коры тем, что процесс
торфообразования идёт в данное время (Тюремнов, 1976). Верхний хорошо
аэрированный слой торфяной залежи носит название «деятельный». На олиготрофных
болотах он имеет мощность 0,5 метра и определяется уровнем грунтовых вод.
Скорость нарастания торфа составляет 1 мм/год в ширину и высоту. В нём
интенсивно идут процессы водо-, газо- и теплообмена, а также накапливается,
впитывается, фильтруется и всасывается корнями растений дождевая вода. (Анучин
и др., 1986; Денисенков, 2000). В основе торфяной залежи олиготрофных болот
лежит низинный торф, который состоит преимущественно из гипновых мхов, либо
осок которые являются «пионерами» заболачивания. После низинного аккумулируется
переходный слой торфа, а затем - олиготрофный. На рисунке 2 показан процесс
формирования болота из озера, которое постепенно зарастает при накоплении
органики на дне. При дальнейшей аккумуляции эта органика увеличивает свою
мощность, образуя сперва торф эвтрофного болота, затем мезотрофного. Конечной
стадии этой аккумуляции является олиготрофное болото. (Инишева, Моторин, 2000).
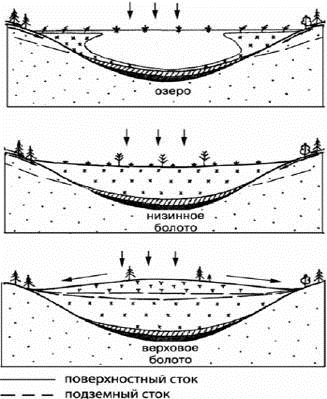
Рис. 2. Стадии развития болота (Инишева, Моторин, 2000)
Сфагновые мхи на болотах образуют слои мохового очёса
мощностью 20-30 см. Торф включает остатки частей сфагнов, состоящих наполовину
из пустых гиалиновых клеток, которые впитывают огромное количество влаги,
удерживая ее длительное время. Когда почвенная вода соприкасается с торфом, то
поднимается вверх, как по губке. В отличие от заболачивания, связанного с
подъемом воды под влиянием внешних причин, объем воды в почвенном слое не
растёт, идёт её вертикальное и горизонтальное распределение. Таким образом,
гравитационная вода переходит в каппилярную. Дренажа (удаления воды из почвы)
при этом нет, однако в силу того, что торф в этом случае представляет собой
системы каппиляров, вода останавливается на границе между ними и поверхность
местообитания становится суше, с большей степенью аэрацией. Из-за этих
процессов сменяются виды гидрофитов на менее влаголюбивые, которые тянут воду
из глубины торфа по системе капилляров. Те же процессы идут при заторфовывании
озёр и больших водоёмов (см. рис. 1.). Вследствие наращивание торфа его верхние
горизонты иссыхают, отчего на них поселяются виды, способные выдерживать долгие
засухи верхнего слоя летом (Лисс, Астахова, 1982). На сегодняшний день понятие
«трофность» не имеет единого согласованного определения и трактуется примерно,
как: «богатство среды», «общая обеспеченность элементами питания» (Аветов,
Шишконакова, 2013). Первые центры заболачивания на территории Западноц Сибири брали
начало в суббореальном периоде среднего голоцена на месте углублений от стока
ледниковых вод, котловин разного происхождения, водоёмах разной степени
трофности. Изначально это были мелкие изолированные болота. В относительно
тёплом и сухом климате уровень вод в мелких озёрах сильно понизился, отчего их
зарастание усилилось, вследствие чего они превратились в болота. В раннем
голоцене после переполнения торфом понижений рельефа началось заболачивание
соседних территорий и слияние отдельных болот в большие болотные системы
(Абрамова, 1974; Тюремнов, 1976). По версии Н.И. Нейштадта (1985) эти процессы
идут и сегодня весьма интенсивно, захватывая ежегодно 92 км², отчего заболачиваются и гибнут леса и луга. Такая динамика
способствует полному заболачиванию и заторфовыванию Западно-Сибирской равнины в
будущем (рис. 3.). Исключение составят только наиболее дренируемые
возвышенности. По версии Ф.З. Глебова (1973) значительное уменьшение ежегодного
прироста площадей болот за последние 2 тысячелетия говорит об угасании
интенсивности процесса болотообразования. Депрессии и понижения
Западно-Сибирской равнины уже заняты болотами. Исходя из этой тенденции через 1
тысячу лет процесс заболачивания закончится, отчего суммарная площадь болот
увеличится только на 2% от сегодняшней.

Рис. 3. Схема развития торфяного месторождения в голоцене (по
М.И. Нейштадту (1985)); а - древний голоцен (12-10 тысяч лет назад); б
- ранний голоцен (10-8 тысяч лет назад); в-средний голоцен (8-2
тысячи лет назад); г - поздний гооцен и современная поверхность (2
тысячи лет назад и по настоящее время); 1 - современная граница торфяного
месторождения; 2 - границы болот и озёр в голоцене; 3 - торф; 4 - сапропель
(озёра); 5 - песок, суходол (Инишева, Моторин, 2000)
Болото является средой обитания для различных видов растений,
хотя и имеет крайне неблагоприятные условия для их произрастания: застойность и
обилие воды, низкая теплопроводность, дефицит в торфе макро и микроэлементов
минерального питания, постоянное нарастание дернины из мхов-сфагнумов и
образования новых слоёв торфа. К произрастанию на болотах адаптировались самые
неприхотливые виды, имеющие очень малое содержание минеральных солей в
организме, что способствует их существованию в неблагоприятных условиях. Все
растения верховых болот - ацидофилы (произрастают в условиях высокой
кислотности среды) (Анучин и др., 1986; Денисенков, 2000). По причине малой
теплопроводности (особенно мохового малой степени разложения) торфа на верховых
болотах создаётся особый температурны режим. Покров из сфагновых мхов изолирует
нижележащие слои торфа от прогревания, отчего температура корнеобитаемого слоя
всегда ниже на 2-4°С. Летом в полдень верхние органы растений нагреваются до
30-40°С и их интенсивность транспирации увеличивается, а корни из-за более
низких температур плохо поглощают влагу. С наступлением весны торфяная вода
оттаивает медленно и корни растений на большом протяжении времени находятся в
холодном субстрате и плохо функционируют. Растения, произрастающие на холодных
и влажных почвах, но испытывающие недостаток влаги из за низких температур в
корнеобитаемом слое, выделяют в самостоятельную экологическую группу -
психрофиты. В осеннее время вода в торфе долго остывает, отчего в корневом слое
остаётся благоприятная температура для вегетации, когда верхние органы растений
уже не вегетируют. Из-за поздней остановки и раннего начала заморозков болотная
флора имеет недолгий период вегетации. По этой причине на олиготрофных болотах
преобладают раннецветущие виды, проходящие все фазы развития за короткий
вегетативный период. Вследствие этого размножение семенами развито слабо,
доминирует вегетативное. Ещё одна приспособительная особенность растений
верховых болот - вечнозелёность, которая даёт им развиваться в условиях короткой
вегетации. Так как олиготрофные болота бедны доступными соединениями азота,
поэтому некоторые растения приспособились получать минеральные вещества не из
почвы, а из насекомых. Своеобразный фактор верховых болот - это нарастание
покрова из сфагновых мхов и торфа, отчего поверхность болот поднимается от
минерального горизонта и уровня грунтовых вод. В период интенсивного накопления
торфа корни растений практически перестают контактировать с грунтовыми водами,
вследствие питаются дождевыми водами, бедными минеральными солями. Кроме того,
ежегодный прирост сфагна погребает нижние части стеблей и корни многолетних
растений (Анучин и др., 1986; Денисенков, 2000). В условиях олиготрофных болот
произрасатют растения следующих экологических групп:
. Гигрофиты - наземные виды, населяющие биотопы с избыточно
увлажненным субстратом и высокой влажностью воздуха (виды из семейства Hymenophyllaceae,
Cyperaceae) (Якушкина, 1993).
. Оксилофиты - включают обитателей олиготрофных болот,
приспособленных к весьма малым концентрациям доступных форм органических и
неорганических веществ и к периодической недоступности воды, связанной в тканях
сфагновых мхов (Oxycoccus palustris Pers., Carex chordorrhiza L.) (Тимонин, Нотов,
1993).
. Психрофиты - растения влажных и холодных почв в холодных
местообитаниях высокогорий и северных широт. Несмотря на достаточное увлажнение
почвы, они часто испытывают недостаток влаги (или из-за физиологической
сухости, вызванной низкими температурами, или в связи с преобладанием в почве
недоступной влаги, как, например, на торфянистых почвах) (Прокопьев, 2001).
. Эпифиты - приспособлены к существованию на поверхности тела
других растений. Они приурочены главным образом к областям с безморозным
влажным климатом (многие лишайники, древесные мхи). Характерными чертами
биотопа эпифитов являются бедность субстрата доступными для растения
неорганическими и органическими веществами и кратковременность доступности воды
в течение суток (Тимонин, Нотов, 1993).
5. Насекомоядные растения - получают азотистые соединения
непосредственно из животных организмов, главным образом из членистоногих (Drosera rotundifolia, D. anglica) (Тимонин, Нотов, 1993).
2. Биоморфологические особенности растений
олиготрофных болот
Изучением растительности болот и олиготрофных в том числе в
своё время занимались такие исследователи, как Н.И. Пьявченко (1963), Ф.З.
Глебов (1973), С.Н. Тюремнов (1976), О.Л. Лисс с соавт. (1981; 2001), М.С. Боч,
Смагин В.А. (1993), Е.Д. Лапшина с соавт. (1997; 2003), Л.А. Волкова (1998),
Л.Г. Бабешина с соавт. (2010; 2011), а именно в Томской области А.В. Положий
(1985), Е.Я. Мульдияров (1990), Н.Ф. Вылцан с соавт. (1994), С.П. Ефремов, Т.Т.
Ефремова, (2000), А.А. Храмов и В.И. Валуцкий (1977), Ю.А. Львов (1987).
Болотные растения в связи с их требовательностью к элементам
делят на следующие группы:
.олиготрофные растения - (от греч. олигос - большой и трофэ -
пища, питание), которые мало требовательны к содержанию питательных веществ в
субстрате.
. эвтрофные растения - (от греч. эу-хорошо), более требовательные
к содержанию питательных веществ в субстрате.
. мезотрофные растения - занимающие промежуточное положение
между эвтрофными и олиготрофными растениями.
Однако растений, приуроченных только к одному определенному
типу болот по питательности субстрата, почти нет. В растительном покрове болот
выделяют несколько ярусов: древесный, кустарниковый, кустарничковый, травяной и
моховой (или мохово-лишайниковый) (Лапшина, Руденко, 1997).
Растениями-торфообразователями называют растения видов, доминирующих в растительном
покрове болот, имеющими преимущественную роль в отложении торфа. В большинстве
своём флора болот голарктическая. Преимущество имеют евро-сибирские,
евро-азиатские, евро-азиатско-американские виды растений. Меньше растений
европейского, атлантического, аркто-альпийского геоэлементов. В широтных
группах доминируют бореальные виды (больше 80%), остальные группы
гипоарктические, аркто-бореальные, бореально-неморальные, неморальные менее
широко распространены (Денисенков, 2000). Из гипоарктических видов севера
встречаются такие представители, как: подбел, водяника, багульник болотный,
карликовая берёза, жирянка. Из южных видов: осока (Carex panuculata), меч-трава (Cladium mariscus) (Боч, Смагин, 1993).
Чёткого ответа на вопрос о происхождении растительности олиготрофных болот на
сегодняшний день нет, т.к. он довольно сложен и не полностью изучен. Нынешняя
растительность болот бореального пояса Евразии рассматривается, как составная
из различных флорогенетических компонентов. В самих же болотных условиях
образовалось очень малое количество видов, большинство из известных сейчас
растений болот являются «пришлыми» из неболотных ландшафтов. Наиболее древними
растениями болот являются гидрофильные травянистые растения: белокрыльник,
сабельник, вахта, наумбургия. Из древней группы деревьев - чёрная ольха. Данные
растения являются потомками флоры лесных низинных болот третичного периода
(Денисенков, 2000). В эпоху оледенения многие третичные болота подверглись
разрушению, но в периоды освобождения территории от ледникового покрова
(межледниковье) процесс заболачивания вновь активировался, во многом из-за
влажных и тёплых климатических условий. Именно в этот временной отрезок
происходило образование флоры болот. Изначально составляющие флору болот виды
растительности были из различных мест происхождения: горных, лесных,
суходольных, водных, водно-болотных и прочих (Богдановская-Гиенэф, 1972).
Потомками горных растений являются карликовая берёза, берёза приземистая, ива
черниколистная, пухонос, мелкотравье: камнеломка болотная, жирянка. Их предки
произрастали в горных долинах, руслах горных рек, на возвышенных местах с
бедным почвенным покровом. Во время оледенения данные виды спустились на
равнины. После отступления ледника эти растения остались расти на новообразующихся
болотных массивах. На болота также проникало множество видов из других
водоёмов, хотя некоторые из них (хвощ приречный, тростник, некоторые виды ив и
осок, камыш) могут произрастать в прибрежьях водоёмов в настоящее время. Но
некоторые в ходе эволюции полностью приспособились к новой среде обитания.
Примером тому является шейхцерия, ставшая исключительно болотным растением, но
даже на болотах она может обитать в небольших водоёмах (озёрках) (Боч, Смагин,
1993). Современные болотные кустарнички являются потомками тропических и
субтропических вечнозелёных кустарников. В конце третичного периода из-за
общего похолодания кустарники внедрились в листопадные леса, после
распространились по болотам, образовав приспособленные к неблагоприятным
условиям расы (багульник, подбел, болотный мирт, из травянистых - морошка).
Водяника и вереск сперва произрастали на скалах и песчаных пустошах, лишь потом
спустившись в болота. Из лесов речных долин на болотах появилась ольха чёрная и
несколько видов ивы (Salix cirenea и т.д.). В третичный
период на болотах росли гипновые мхи. После отступления ледника мхи начали
активно заселять остаточные водоёмы, активно участвуя в заболачивании
минеральных почв. Сфагновый мох получил широкое распространение в поздние
периоды голоцена. Таким образом флора болот лесной зоны относительна молода,
процесс взаимной адаптации к совместному произрастанию на данный момент не
окончен (Денисенков, 2000).
Деревья и кустарники
Древесные растения - это многолетние вечнозелёные и
листопадные растения, ствол и ветви которых образуют древесину и имеют
многолетние надземные побеги с почками возобновления, находящимися на большой
высоте в кроне. Древесные растения подразделяют на три типа: деревья,
кустарники и кустарнички (Тимонин, 2007). Древесные породы, произрастающие на
верховых болотах не типичны для них. Их семена заносит из сухих местообитаний и
они развиваются на болотах только потому, что могут выносить эти
неблагоприятные условия. Здесь обитают угнетённые формы деревьев: сосна, кедр,
лиственница, береза и др. (Анучин и др., 1986). По мнению Бабешиной Л.Г. (2011)
древесные растения, а именно берёза и сосна на верховых болотах произрастают в
угнетённом состоянии из-за недостатка элементов минерального питания,
недостатка кислорода в корнеобитаемом горизонте и разницы температур наземной и
подземной частями, отчего сокращён вегетационный период и происходит нарушение
транспирации. В неблагоприятных для роста и развития деревьев условиях
понижается интенсивность ростовых процессов, что проявляется в уменьшении
прироста, размерах хвои, изменении некоторых параметров шишек, кроме того, как
показали исследования у хвойных деревьев, растущих на болотах Западной Сибири в
таких условиях происходит изменение морфологии хромосом, а также частот
встречаемости вторичных перетяжек и некоторых аллелей (Седельникова, Муратова,
2000). На сфагновых болотах сосна обыкновенная (Pinus sylvestris) образует или небольшие
деревца или даже кустарники с выраженными следами угнетенного роста. Ее ствол
достигает небольших диаметров, а у некоторых болотных форм главный ствол или
совсем отсутствует или имеет в высоту лишь 75 см. Хвоя сильно укорочена, шишки
встречаются редко. Предельный возраст таких растений 70 лет, причём весь
стволик с ветвями погружен в торф на 40-60 см и над ковром сфагнума поднимаются
лишь концы ветвей. Другие же формы болотной сосны хотя и имеют вид деревца, но
также не высокого: в 1-3 м на моховых глубоких болотах, а на менее глубоких,
растет форма с более стройным стволом высотою в 10-12 м, достигающая возраста
120-140 лет (Волкова, 2000; Тимонин, 2007). Биология болотной сосны в некоторых
отношениях иная, нежели сосны, выросшей на сухих местах. Разные условия
произрастания вносят поправки в обычный облик сосны обыкновенной (рис. 4.).
Этот вид может образовывать формы, приспособленные к различным влияниям
факторов среды. О генетических различиях между разными формами свидетельствуют
и биохимические исследования. Экстремальные условия произрастания на болоте
привели к возникновению в ферментных системах, особенно участвующих в дыхании,
большого разнообразия. Возникновению генетических отличий, видимо, способствует
специфическое направление отбора в разных условиях и обусловленная
существованием на болотах пространственная, фенологическая и, в конечном итоге,
репродуктивная изоляция. В результате формируются популяции сосны,
характеризующиеся особым обликом (Волкова и др., 2008).

Рис. 4. Экологические формы сосны (Pinus silvestris), произрастающей на
олиготрофных болотах: 1 - f. pumila, 2 - f. willkommii, 3 - f. litwinowii, 4 - f. uliginosa (Тюремнов, 1976)
Сосну можно найти там, где другие породы не смогут расти.
Такими местами могут служить пески, скалы, меловые выходки. В настоящее время
известно большое количество форм сосны, примерами которых могут служить
«узкокронная» и «ширококронная» сосна, «плакучая» и «змеевидная» сосна, «сосна
в юбке» и многие другие. В целом у сосны обыкновенной выделяют 5 подвидов,
около 40 географических рас, более 100 разновидностей, а также огромное число
экотипов и форм (Волкова, 2000). В зависимости от вида болот и типа
растительных сообществ, сосна обыкновенная имеет различные угнетённые формы
(см. рис. 4.). Данные формы сосны - имеют разную высоту, различные варианты
формы кроны, разную длину и ширину шишек. Лучше всего растёт сосна f.uliginosa. Высота этого дерева
варьирует от 10 до 12 м, с с более-менее прямым стволом и кроной шаровидной
вормы на его верхушке. Эта форма сосны относительно густо произрастает по
окраинам болот, либо на очень хорошо дренируемых участках. Сосна формы
Литвинова (f. litwinowii) представляет собой дерево с высотой от 2 до 4 м,
имеющее яйцевидную крону в верхней половине, либо трети ствола. Произрастает
преимущественно на олиготрофных болотах, с большой частотой встречается на
границах болот этого типа. Сосна формы Вилькома (f. willkommii) является деревом,
высота которого варьирует от 1 до 3 метров и тупоконусовидной кроной. Ветвление
густое, всестороннее; ветви идут от ствола практически под прямым углом,
начинаясь почти от прикорневой шейки. F. willkommii произрастает на участках
с большей степенью обводнённости, чем f. litwinowii, но с лучшим освещением.
Сосна формы «пумила» (f. pumila) по габитусу больше
напоминает чахлый кустарник высотой от 0,75 до 1,5 м. В некоторых случаях
практически полностью погребается в сфагновом покрове, оставляя на поверхности
только верхушки ветвей. Данная форма обитает на центральных участках с сильной
степенью обводнённости (Тюремнов, 1976). Сосна в процессе эволюции приспосабливалась
к тяжёлым (для не болотных растений) условиям. Если в борах на песчаных почвах
она имеет крупный стержневой корень, достигающий большой глубины, то на
торфяниках, в условиях недостаточной аэрации, корневая система её стала
горизонтальной, доходящей до глубины лишь 30-50 см. Чем угнетённая сосна, тем
больший диаметр охватывает её корневая система (30-50 летние растения имеют
радиус корневой системы до 5 м, а окончания мелки корешков распространяются
ближе к поверхности субстрата) (Тюремнов, 1976). Рассмотрим несколько примеров
растений-доминантов из семейства сосновых в древесном ярусе.
Сосна сибирская (Pinus sibirica L.) - распространена на
верховых болотах Западной и Восточной Сибири, а на Урале растет по окраинам
крупных низинных болот в елово-кедровых сограх. Сосна сибирская, по сравнению с
другими видами сосен, лучше приспособлена к росту на верховых болотах, так как
по мере нарастания торфяного пласта она дает придаточные корни в нижней части
ствола, выше корневой шейки. Встречается на глубоких сфагновых болотах Сибири,
но в этих неблагоприятных для неё условиях Pinus sibirica выглядит чахлым
невысоким деревом (Лапшина, 2003).
Ель сибирская (Picea obovata Ledeb.) - представитель
семейства сосновые (Pinaceae), теневыносливое влаголюбивое растение,
плохо переносящее избыток воды, особенно застойной. Поэтому, как правило, она
не растет на верховых болотах и встречается чаще вместе с ольхой на таких
участках низинных болот, где имеется проточная вода. На болотистых и на
умеренно влажных почвах ель способна в самом поверхностном горизонте почвы
образовать придаточные корни, начиная с 3-4-летнего возраста. Но здесь она
имеет угнетенный вид и достигает лишь 8-10 м в высоту.
На территории верховых болот
Западно-Сибирской равнины произрастает лиственница сибирская (Larix sibirica Ledeb.). Её высота здесь
варьирует от 5 до 6 м, а диаметр ствола от 10 до 12 см. Так же, как и сосна
сибирская в ходе эволюции она адаптировалась к условиям постоянного нарастания
торфяной толщи на верховых болотах. Ярким приспособлением является способность
отпускать придаточные корни в нижней части ствола, которые заменяют глубоко
погребённые под постоянно растущим торфяником, отмершие в слое многолетней
мерзлоты первичные корни. На олиготрофных болотах лиственница живёт дольше
сосен - от 300 до 400 лет, а сосны гибнут преждевременно. Из семейства
берёзовых на олиготрофных болотах произрастают несколько древесных видов:
карликовая, приземистая, пушистая. Их различия состоят в высоте стволов и форме
листьев. На верховых болотах Камчатки и Сахалина произрастают кустарниковые
формы берёз: миддендорфа и тощая (Тюремнов, 1976).
Берёза пушистая (Betula pubescens Ehrh.) - растение из
семейства берёзовые (Betulaceae), достигающее при благоприятных условиях
25-30 м в высоту и до 80 см в диаметре. Живёт около 120 лет, реже до более
взрослого возраста. Кора у молодых деревьев коричневато-бурая, а с 8-10 лет
белеет. Хорошо отличается от других деревьев по белой коре; кора почти до
самого основания ствола белая, гладкая, без чёрных трещин в отличие от берёзы
повислой, лишь внизу неглубоко растрескивается в старости. Молодые побеги
густоопушенные, в отличие от берёзы повислой на ветвях нет желёз-бородавочек.
Ветви не поникающие. Крона в молодом возрасте стройная, узкая, с возрастом
становится раскидистой. Листья яйцевидные или ромбически-яйцевидные, 3,5-7 см
длины, 2,5-5 см ширины, короткозаострённые на верхушке, с округлым, реже
сердцевидным или усечённым основанием; края двоякозубчатые. В молодом возрасте
листья густо опушены, затем опушение сохраняется только снизу и на черешках.
Растёт по краям верховых болот в дренируемой полосе и в центре на возвышенности
(Фёдоров, 1980).
Берёза карликовая (Betula nana L.) - растение относящееся
к семейству берёзовые (Betulaceae). Представляет листопадный сильно ветвистый
кустарник с мелкими и округлыми листьями. Её высота 20-70 (до 120) см.
Карликовая береза имеет приподнимающиеся или распростертые побеги, которые на
вид густо бархатистые или пушистые. Кора почти голая, с тёмно-коричневой или
красновато-тёмно-бурой окраской. Листья же округлые, реже округло-овальные,
длиной 5-15 мм, шириной 10-20 мм, у основания округлые или, часто,
широко-клиновидные, с закруглённой верхушкой, широко клиновидным основанием, с
притупленными зубчатыми краями. Сверху листья тёмно-зелёные, глянцевые, снизу
светло-зелёные и рассеянно пушистые; в молодом возрасте клейкие. Произрастает
на верховых болотах северных (тундровых) районов, образуя нередко сплошной
кустарниковый ярус; реже встречается на низинных болотах (Гроздова и др.,
1986). Итак, основными чертами приспособленности деревьев и кустарников в
условиях жизни на олиготрофных болот является, прежде всего, горизонтальная
корневая система, распространяющаяся в моховом слое; низкий рост и диаметр
стволов, что сказывается из-за дефицита минеральных веществ.
Кустарнички
Доминантами в растительном покрове
олиготрофных болот наряду со сфагновыми мхами являются кустарнички. В
растительном покрове торфяных болот кустарнички представлены в основном
семействами: вересковых (Ericaceae) и водяниковых (Empetraceae).
Кустарнички - жизненная форма растений, с несколькими надземных
одревесневающими побегами. Отличаются от кустарников продолжительностью
жизненного цикла побегов (не более 5-10 лет). Кустарнички имеют высоту от 5 до
60 см, собственно из за малых размеров они носят это название. (Быков, 1983).
Кустарнички и травы образуют приспособления, помогающие им адаптироваться к
нарастанию. Кустарнички (в основном) пускают придаточные корни из погруженных в
сфагнум и торф частей (ветвей, стволов, корневищ) (Денисенков, 2000). Для
большинства кустарничков олиготрофных болот характерна ксероморфность,
обусловленная двумя причинами:
1. Большинство кустарничков являются потомками субтропических
и тропических растений, поселившихся на болотах ещё в третичный период;
. Летом поверхность сфагновых болот сильно иссушается и
нагревается до +40°С.
Кустарнички олиготрофных болот подразделяют на вечнозелёные
ксероморфные плосколистные и на вечнозелёные ксероморфные кустарнички с мелкими
листовыми пластинками, свёрнутыми внутрь на нижней стороне. Данные типы
кустарничков сочетают в себе ксероморфные и гидроморфные черты. Так
представители листопадных кустарничков, имеющие листья мезоморфного строения на
болотах имеют низкорослые формы (ива (Salix), голубика (Vaccinium uliginosum)). Надземные и подземные
органы кустарничков олиготрофных болот не имеют, либо имеют очень слабо
выраженную систему межклетников и воздушных полостей, отчего живая часть их
корневой системы распространяется в достаточно хорошо аэрируемых повышениях
микрорельефа, немного выше уровня грунтовых вод (Тюремнов, 1976). То есть,
корневые системы болотных кустарничков являются поверхностными, что типично для
высших растений олиготрофных болот. Характерной чертой кустарничков верховых
болот является приспособленность к бедному минеральному питанию и высокой
кислотности субстрата. Рассмотрим некоторые кустарнички, произрастающие на
олиготрофных болотах. Одним из распространённых семейств олиготрофных болот
является семейство вересковых (Ericaceae), представители которого не имеют
травянистых форм (рис. 5.). Листья у вересковых не имеют прилистников, в
большинстве случаев с очередным листорасположением, с меньшей частотой можно
встретить супротивное листорасположение. Листовые пластинки подразделяются на три
типа: 1) плоские, широкие, кожистые (брусника); 2) заостренные игольчатые; 3)
эрикоидные - мелкие, линейные, с бороздкой на нижней стороне, где сосредоточены
устьица. Интересно, что все виды этих листьев имеют ксероморфное строение,
из-за невозможности потребления грунтовых вод. При произрастании на кислых
почвах и в условиях повышенной влажности (т.е. на верховых болотах) растения
данного семейства образуют очень густые заросли. Примерами являются голубика (Vaccinium uliginosum), клюква (Oxycoccus quadripetalus и Oxycoccus microcarpus), багульник (Ledum palustre) и многие другие
(Еленевский и др., 2006).

Рис. 5. Разновидности вересковых: 1 - багульник болотный (Ledum palustre); 2 - подбел-белолистник
(Andromeda polifolia); 3 - болотный мирт (Chamaedaphne calyculata); 4 - вереск (Calluna vulgaris); 5 - клюква
четырёхлепестная (Oxycoccus quadripetalus) (Тюремнов, 1976)
За длительное время произрастания на кислых и бедных
минеральными веществами почвах олиготрофных болот растения семейства вересковые
(Ericaceae) выработали ряд приспособлений для выживания в этих сложных
условиях. Например вид кассиопея Редовского (Cassiope redowskii) имеет лист с
полузамкнутой полостью, похожий на палец перчатки. У множества видов вересковых
листовые пластинки имеют способность сворачиваться книзу в засушливые периоды
(например у брусники). Эта особенность обусловлена тем, что сильное испарение
воды в засуху снижает напряжение воды в мезофилле на нижней стороне листа,
вследствие чего лист сгибается, либо скручивается книзу, что практически
полностью ограничивает контакт устьиц с окружающей средой. Известно, что у
множества видов вересковых устьица имеются и на верхней стороне листа (у
вереска обыкновенного). Их нет только на боковых сторонах, которые имеют
толстый кутикулярный слой. Следовательно - процесс испарения может
осуществляться даже при полной изоляции устьиц нижней стороны листа при
свёртывании. Если такой свёрнутый лист опустить в воду, то его верхняя сторона
будет смочена, зато снизу останется пузырёк с воздухом, который нельзя удалить
при встряхивании. Из-за данной особенности устьица нижней стороны листа
остаются сухими и могут транспирировать даже под водой (Федоров, 1981). Ещё
одной характерной адаптационной особенностью вересковых для обитания на почвах
верховых болот - симбиоз с грибами, то есть образование микоризы. Практически
все корневые системы вересковых плотно оплетают гифы грибов, которые снабжают
кустарнички питательными веществами из перегноя. Микориза подразделяется на 2
типа: эндомикориза (гифы оплетают корень, не проникая в клетки) и эктомикориза
(некоторые виды простейших грибов обитают внутри клеток корня). В ряде случаев
поражённые грибами корневые волоски видоизменяются в микоматии - клубеньки
грушевидной формы. Примечательно, что семена вересковых не могут прорастать без
вмешательства микоризы. По мнению некоторых учёных есть предположение, что
вересковые живут на кислых почвах только из-за симбиоза с грибами, но эти грибы
не могут расти на щелочных почвах, отчего и вересковые на них не обитают
(Федоров, 1981). Таким образом, можно сделать вывод, что у ряда вересковых
имеются физиологические ответы на изменение условий окружающей среды. На
болотах из семейства Ericaceae встречаются преимущественно следующие виды:
багульник болотный, подбел-белолистник, болотный мирт, вереск, голубика,
вороника и клюква болотная, брусника, черника (Тюремнов, 1976).
Багульник болотный (Ledum palustre L.) (см. рис. 5.)
представляет собой кустарничек из семейства вересковые (Ericaceae), имеющий хорошо
сформированную крону из немногочисленных побегов. Багульник имеет узкие
кожистые, вечнозелёные листья на коротких черешках с рыжими трихомами, нижняя
сторона которых загнута. Листовые пластинки пережившие зиму значительно
оттопырены от стеблей. Цветки собраны в соцветие полузонтик, имеют белую
окраску. Примечательно, что во всех органах багульника имеется большое
количество эфирных масел, отчего имеют сильный запах ледола. Большая часть
ареала багульника приурочена к олиготрофным болотам и заболоченным лесным
массивам. Здесь на сильно обводнённых участках он имеет угнетённую форму,
высотой от 100 до 150 см, в мочажинах обычно не растёт. В процессе
торфообразования багульник имеет незначительную роль (Денисенков, 2000).
Клюква болотная (Oxycoccus quadripetalus Gilib) (см. рис. 5.)
является вечнозелёным многолетним кустарничком из семейства Вересковые (Ericaceae). Кустарничком это
растение считается потому, что её стебель одревесневает и покрывается сверху
корой с защитной пробковой тканью, как и ветви деревьев. Внешне это низкорослое
растение, длина побегов которого доходит до 80 см. Листья клюквы мелкие,
продолговатые по форме. Примечательно, что листовые пластинки с нижней стороны
покрыты воском, что является защитой от высоких температур, исходящих от
мохового слоя в летнее время. К тому же восковой налёт не даёт заливать воде
устьица. Цветки клюквы имеют темно-розовую окраску, находятся они на концах
прошлогодних ветвей. Цветёт с мая по июнь, примечательно, что срок жизни одного
цветка составляет 18 дней. Плоды мелкие и круглые, похожи на шарик, отчего
происходит название растения («oxys» - кислый; «coccus» - шарообразный)
(Губанов, 2004). Помимо воскового налёта клюква имеет ещё одно приспособление
для выживания на олиготрофных болотах - гриб, произрастающий на её корневой
системе. Характерно, что его гифы плотно соединяются с клетками корня, что в
совокупности образует микоризу. Его гифы поглощают из почвы растворы
минеральных веществ и переносят их к корням. Гифы гриба поглощают из почвы
питательные растворы и передают их корням. Эта особенность возникла из-за
отсутсвия собственных корневых волосков (Лисс, Астахова, 1982). Клюква болотная
преимущественно произрастает на олиготрофных болотах, на сплавинах зарастающих
водоёмов, заболоченных и редкостойных лесах, образует заросли. Ягоды используют
в пищу в свежем, либо переработанном виде, к тому же они являются ценным сырьём
для промышленности (из плодов вырабатывают пищевой краситель). Листья
используют вместо чая. Является лекарственным растением и инсектицидом. С
меньшей частотой на олиготрофных болотах Северного полушария встречается Клюква
мелкоплодная (Oxycoccus microcarpus Turcz. Ex Rupr.). Этот вид схож с
клюквой болотной, но все её органы меньше, цветки располагаются на голых цветоножках
(у болотной они опушены). Ягоды также используют в пищу, их свойства такие же,
но из-за мелких размеров они не имеют большого спроса (Губанов и др., 2004).
Черника (Vaccinium myrtillus L.) представитель
семейства вересковые (Ericaceae). Представляет собой кустарничек, высота
которого варьирует от 30 до 40 см (рис. 6.). Побеги зелёные, остроребристые
голые, корневища длинные и ползучие. Цветки одиночные, расположены по два в
пазухах листьев, поникающие. Плод - шаровидная чёрно-синяя ягода, имеющая длину
от 0,5 до 1 см. Цветёт в мае (Харкевич, 1987). Листья тонкие эллиптической,
либо яйцевидной формы. Их длина варьирует от 1 до 3 см, края пильчато-зубчатые
(зубчики на конце с железистой щетинкой). Произрастает в хвойных лесах с
высокой влажностью воздуха и на олиготрофных болотах. Является микотрофом.
Используется в пищу и в медицине. Ягодный сок является хорошим красителем,
верхняя часть растения применяется для дубления кож из-за наличия дубильных
веществ (Губанов и др., 2004).

Рис. 6. Черника (Vaccinium myrtillus L.) (Губанов и др., 2004)
Брусника (Vaccinium vitisidaea L.) вечнозелёный
кустарничек из семейства вересковые (Ericaceae). Стебель ползучий,
веточки поднимаются на высоту от 2,5 до 25 см, побеги округлые в сечении,
веточки беловато-волосистые. Листья на коротких опушённых черешках, зимующие,
кожистые жёсткие, верхняя сторона блестящая. Форма листа эллиптическая либо
обратнояйцевидная, имеют завёрнутые края. Длина листовой пластинки разнится от
0,5 до 3 см длиной, ширина от 0,3 до 1,2 см шириной. Верхняя сторона имеет
тёмно-зелёную окраску, нижняя бледная с тёмно-бурыми рассеянными желёзками.
Цветки располагаются на коротких опушённых красноватых цветоножках, собраны в
густой поникающей кисти из 2 или 8 цветков на концах прошлогодних веточек
собраны короткой, но густой поникающей 2-8-цветковой кистью, обладают слабым,
но приятным запахом. Ягода тёмно-красного цвета, форма её близка к шаровидной.
Цветёт с мая по июнь (Флора СССР, 1952; Положий и др., 1985; Вылцан, 1994;
Денисенков, 2000). Произрастает брусника в сосновых борах, на кислых почвах
заболоченных березников, либо еловых массивах, на грядах и по краям
олиготрофных болот формируют густой сплошной покров. Используется в пищу, в
дублении кож, в медицине и ветеринарии, как кормовое растение для медведей и
кормовой дичи (Губанов и др., 2004).
Болотный мирт (Chamaedaphne calyculata (L.) Moenh) - вечнозеленый
кустарничек (рис. 7.) из семейства вересковые (Ericaceae) с выраженым
разветвлением.
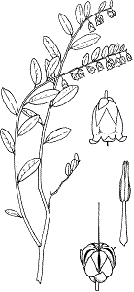
Рис. 7. Болотный мирт (Chamaedaphne calyculata (L.) Moench) (Губанов и
др., 2004)
Высота кустарничка варьирует от 40 до 100 см. Корневая
система поверхностная и представляет собой совокупность придаточных корней
нескольких порядков, произрастающие на погружённых в сфагнум стволиках. Молодые
побеги чешуйчато-пушистые. Листья располагаются на коротких черешках, кожистые,
их форма варьирует от продолговато-ланцетной до продолговато-овальной. С обеих
сторон листовой пластинки имеется покрытие в виде мелких беловатых или бурых
чешуек. Верхняя сторона листа имеет грязно-зелёный, а нижняя - ржаво-зелёный
цвет. Цветки поникающие, имеют белую окраску, собраны в облиственные однобокие
кисти на концах побегов. Обитает преимущественно на олиготрофных болотах и
светлых заболоченных лесных массивах. На бедных кислых почвах формирует густые
заросли. Является микотрофом. Растение ядовито, может вызвать отравление
мелкого рогатого скота. Мёд с цветков болотного мирта ядовит для человека и
носит название - «пьяный мёд». (Губанов и др., 2004).
Подбел-белолистник (Andromeda polifolia L.) - растение,
представляющее вечнозелёный кустарничек из семейства вересковые (Ericaceae), длина стеблей которого
достигает около 30 см (рис. 8.). Стебли тонкие, имеют красновато-буроватую
окраску, облиственны длинными и узкими кожистыми листовыми пластинками.
Примечательно, что на зиму цвет листьев не изменяется. Края листовых пластинок
завёрнуты вниз, нижняя их поверхность голубовато-белой окраски из-за воскового
налёта. Из-за увеличения высоты нарастающего сфагнового «ковра» корневая
система образует ползучие, хорошо ветвящиеся придаточные корни,
распространяющиеся в верхнем слое «ковра». Интенсивное размножение корневищами
сделало данный кустарничек одним из устойчивых составляющих верховых болот.
Цветки имеют ярко-розовую окраску, собранные в верхушечные соцветия. Плод -
почти шаровидная коробочка, длина которой колеблется от 3 до 5 см. Цветёт с мая
по июнь, плодоношение происходит с июля по август. Произрастает на обводнённых,
хорошо дренируемых участках олиготрофных болот (Тюремнов, 1976; Минина, 2002;
Губанов и др., 2004).

Рис. 8. Подбел-белолистник (Andromeda polifolia)
(Инишева, 2009)
Вереск обыкновенный (Calluna vulgaris (L.) Hull) - вечнозелёный
ветвистый кустарник (рис. 9.) из семейства вересковые (Ericaceae), имеющий высоту от 20
до 80 см. Кора имеет красновато-бурую окраску. Листья эрикоидные, с сильно
завёрнутыми вниз краями, располагаются в черепитчатом порядке, имеют линейную
форму и трёхгранны в сечении, при основании глубоко стреловидной формы. Цветки
собираются однобоких кистях, переходящих в несущие листья веточки. Чашечка
четырёхраздельная, блестящая, плёнчатая, как и венчик розовая, иногда белая.
Плод - четырёхстворчатая пушистая коробочка, с небольшим количеством семян,
открывающаяся по перегородкам. Произрастает на олиготрофных болотах, местами
образует густые заросли - верещатники. Используют как медонос, веточный корм
для скота, в местах с мягким климатом используется, как корм для овец,
декоративное растение (Губанов и др., 2004).

Рис. 9. Вереск (Calluna vulgaris (L.) Hull) (Губанов др., 2004)
Голубика
обыкновенная (Vaccinium uliginosum L.) - многолетний, листопадный,
сильно ветвистый кустарник, имеющий высоту от 30 до 120 см. Имеет придаточные
корни, постоянно формирующиеся на погребённых стволиках. Побеги тёмно-серые или
коричневато-бурые, цилиндрическими. Стебли прямостоячие, молодые ветки зелёные.
Листья плотные, эллиптические, жесткие, с цельными, немного загнутыми вниз
краями, на коротких или опушенных черешках. Сверху листья светло-зелёные или
тёмно-зелёные, снизу сизые или сизоватые, покрытые голубоватым восковым
налетом, с длиной до 3 см. Осенью листья краснеют, на зиму отпадают. Цветы
голубики кувшинчато-колокольчатые, бледно-розоватого или беловатого цвета со
слабым приятным ароматом, по 2-3, редко одиночные, расположены на коротких
поникающих цветоножках в пазухах листьев. Цветки расположены на ветвях так, что
отверстие венчика направлено вниз. По краю отверстия располагаются 4-5 мелких
зубчиков, представляющих собой концы лепестков. Издавна использовалось, как
источник красящих веществ, как лекарственное и пищевое растение (Минаева,
1997).
Травянистые растения
Травянистые растения верховых болот в своей основе являются
многолетними и в большинстве случаев гигрофиты. Органы растений сфагновых болот
приспособлены к переувлажнённой среде и имеют довольно развитую систему
воздушных полостей. Корневая система этих растений имеет вид длинных корневищ с
симподиальным ветвлением. К общим чертам, характерным для данных растений,
относится преимущественно вегетативное размножение и положение перезимовывающих
органов с почками возобновления почти у всех под поверхностью болота. Такие
растения, как росянка, пушица, вахта - образуют вертикальные и наклонные
корневища, ежегодно вынося узлы кущения на поверхность (и это всё при приросте
сфагнового покрова и торфа) (Денисенков, 2000). Среди травянистых растений
болот, в том числе и олиготрофных встречаются следующие растения:
Осока - принадлежит классу однодольных,
среди растений-торфообразователей большое значение имеет богатое видами семейство
осоковых (Cyperaceae) (рис. 10.). Это многолетники с характерным стеблем
без узлов. К этому семейству принадлежат роды: осок (Carex), очеретников
(Rhynchospora), пушиц (Eriophorum), пухоносов (Trichophorum),
камышей (Scirpus). Стебель осоки обычно трехгранный или с режущим краем.
Большинство видов осок нуждаются в хорошем минеральном питании, отчего довольно
часто встречаются на эвтрофных (низинных) болотах, но есть виды, менее
требовательные к минеральному питанию, поселяющиеся на мезотрофных (переходных)
и олиготрофных (верховых) болотах. Это такие виды осок, как: осока топяная (Carex limosa L.), осока плетевидная (Carex chordorrhiza Ehrh. ex L. f), осока
вздутая (Carex rostrate Stokes), осока двудомная (Carex dioica L.), осока миддендорфа (Carex middendorfii F. Schmid) и др. На
олиготрофных болотах на кочках из Sphagnum fuscum нередко встречаются C.
pauciflora Lightf., C. globularis L. (Западная Сибирь) и C.
middendorfii (о. Сахалин и Камчатка) (Федоров, 1982; Инишева, 2009).
Семейство осоковых играет очень большую роль, так как многие виды являются
эдификаторами в сильно увлажнённых фитоценозах олиготрофных болот (Еленевский,
2006).

Рис. 10. Разновидности осок: 1 - вздутая (C. rostrata Stokes); 2 - топяная (C. limosa L.); 3 - волосистоплодная (C.
lasiocarpa); 4 - сближенная (C. appropinguata Schumach); 5 - дернистая
(C. caespitosa L.) (Тюремнов С.Н., 1976)
Осока топяная (Carex limosa L.) - многолетнее
травянистое растение (см. рис. 10.). Имеет ползучие корневища. Основания
побегов с цельными красновато-бурыми чешуевидными и листовыми влагалищами.
Листья узкие, с желобками, сероватые 0,5 - 1,5 мм шириной, чуть короче стеблей.
Стебли в верхней части слегка шероховатые или гладкие. Их высота колеблется от
20 до 50 см. Верхний колосок тычиночный, остальные (1-3) - пестичные или иногда
андрогинные, немного расставленные, на тонких длинных ножках, поникающие,
эллиптические. Используется как кормовое растение (Малышев, Пешкова, 1984).
Осока шаровидная (Carex globularis L.) представляет собой
многолетнее травянистое растение с расходящимися, либо сближенными стеблями,
высота которых варьирует от 15 до 40 см, ширина около 0,5 мм (рис. 11.). Стебли
ортотропные, прямые, на ощупь шероховатые, в базальной части имеют
чешуеобразные листья пурпурного окраса, на плёнчатой части которых имеется
опушение.

Рис. 11. Осока шаровидная (Carex globularis L.) (Губанов и др., 2002)
Соцветие имеет от 2 до 4, редко 5 колосков, примечательно что
верхний из них является тычиночным. Его длина составляет около 12 см, окраска
светло-коричневая. Пестичные колоски сидячие, слегка оттопыренные, имеют форму
шара или эллипса, длиной около 1 см. Кроющие чешуйки на пестичных цветках имеют
яйцевидную форму, красно-бурой окраски с зелёной полоской по средине, их края
перепончатые. Произрастает по окраинам олиготрофных болот, заболоченных моховых
сосняках, формирует густые заросли, является доминантом в травяном сообществе
(Губанов и др., 2002).
Пушица влагалищная (Eryophorum vaginatum L.) - представитель
семейства осоковые (Cyperaceae), образует плотные кочки на болотах.
Листья темно-зеленые, поникающие. Прикорневые листья узкие, трехгранные,
стеблевые - вздутые, трубчатые, с темными пленчатыми верхушками. Стебли высотой
30-100 см, многочисленные, прямые, треугольные, с одиночными направленными
вверх колосками. Цветущие колоски желтые. Плодоносящие колоски широко
обратнояйцевидные, ширина пуховки равна ее длине 4-4,5 см, щетинки белые,
блестящие. Используется как кормовое и лекарственное растение, является одним
из основных торфообразователей, составляя главную массу «пушицевого торфа»
(Ефремов, Ефремова, 2000).
Очеретник белый (Rhynchospora alba (L.) Vahl.) - многолетнее
травянистое растение, являющееся представителем семейства осоковые (Cyperaceae)
(рис. 12.). Имеет стебли высотой 10-40 см и 0,2-1 см шириной, трёхгранные, в
верху по граням шероховатые. Длина листовых пластинок варьирует от 3 до 13 см,
а ширина от 0,2 до 1,5 см. Листья сложены вдоль, имеют цельные, трубчатые
влагалища. Корневище имеет короткие побеги и образует редкий дёрн. Соцветие
верхушечное, пучковидное до 1 см длиной. Колоски имеют длину 3,5-5. Плод длиной
1,5-2 см, шириной 1 мм, плосковыпуклый, к верху заострён в носик 1 мм длиной.
Растёт на верховых болотах и торфяных грядах, массово активно заселяет
выгоревшие торфяники. В грядово-озерном комплексе Rhynchospora alba
образует сплошной пояс вокруг озер. Занесено во многие Красные книги областей
России (Губанов и др., 2002).

Рис. 12. Очеретник белый (Rhynchospora alba (L.) Vahl.) (Губанов и др., 2002)
Осоковые, обитающие во влажных условиях
водоёмов выработали в ходе эволюции характерное для семейства адаптивное
высокоспециализированное образование - закрытый мешочек, где находится женский
цветок - пестик, а в дальнейшем - плод. Мешочек является видоизменённым
прилистником боковой, в сильной степени редуцированной оси, на которой
располагается 1 женский цветок. Эти мешочки защищают завязи и зреющие плоды от
неблагоприятных условий, которые присущи олиготрофным болотам. Помимо этого они
участвуют в распространении семян осоковых на большие пространства. Плавучесть
мешочков в условиях водоёмов (олиготрофных болот) обусловлена наличием в них
воздуха. Помимо мешочков осоковые имеют приспособление в виде волосков на
плодах. Если рассматривать на примере пушицы влагалищной (Eriophorum vaginatum), то эти волоски имеют
белую окраску, хотя у других видов они рыжие. Данные волоски развиваются на
месте околоцветника, в процессе созревания плода они значительно удлиняются.
Значение этих волосков заключается в приспособлении размножения посредством
анемохории и в фиксации семян на голом влажном субстрате, чему способствует их
гигроскопичность (Еленевский, 2006).
Также на олиготрофных болотах произрастают
растения семейства вахтовые (Menyanthaceae). Типичным
представителем является вахта трёхлистная (Menyanthes trifoliata L.) -
многолетнее травянистое растение с толстым и длинным ползучим корневищем (рис.
13.). Имеет высоту от 15 до 40 см. Листья в длину до 30 см, имеют длинные
черешки, с влагалищами, крупные, с глубоко разделённой на 3 части листовой
пластинкой, отходят от корневища. Цветонос безлистный, выходит из пазух
листьев, до 30-60 см высотой. Цветки бледно-розовые, собраны в продолговатой
кисти на верхушке цветоносного стебля: лепестки сросшиеся, на внутренней
стороне бахромчато-реснитчатые. Плод - одногнёздные яйцевидные коробочки с 6-8
семенами. Используют при лечении сердцебиения, малярии, проблем пищеварения,
геморроидальных кровотечений, желчекаменной болезни, туберкулеза легких, кашля,
как слабительное и глистогонное средство.
Пищевая промышленность использует листья в
пивоварении (Губанов и др., 2004).

Рис. 13. Вахта трёхлистная (Menyanthes trifoliata L.)
(цветок и плод) (Губанов и др., 2004)
Наумбургия кистецветная (Naumburgia thyrirsiflora Rchb.) один из
представителей семейства вахтовые (Menyanthaceae). Травянистое растение
высотой до 65-100 см. Имеет толстые хорошо разветвленные корни, с мутовками и
коричневыми и беловатыми столонами. Стебли простые, в нижней части с буроватыми
чешуевидными равномерно распределёнными листьями. Верхние листовые пластинки
имеют длину от 4 до15 см и ширину от 0,5 до 4 см, сидячие либо
стеблеобъемлющие. Листьям присуща ланцетная форма с заострением и цельными
завёрнутыми краями. Цветки жёлтой окраски и малых размеров, собраны в соцветие
кисть, на цветоносах, выходящих из пазух листьев. Плод - раскрывающаяся
створками коробочка округлой формы. Является декоративным растением (Харкевич,
1987).
Шейхцерия болотная (Scheuchzeria palustris L.) - многолетнее
травянистое растение из семейства шейхцериевые (Scheuchzeriáceae), с корневищами длиной 50 см, являющимися
ползучими побегами, обитающими в моховой толще (рис. 14.). Корневищная внешняя
кора имеет под эпидермисом три слоя клеток с утолщёнными оболочками, внутренняя
кора имеет объёмные воздухоносные полости, сообщающиеся между собой. В моховом
слое корневища соединяют более трёх надземных побегов. Корневище шейхцерии за
один вегетационный период увеличиваются до длины около 30 см. В связи с
нарастанием торфяного слоя корневище отмирает в отдалённых от надземных побегов
участках. Окраска надземных побегов варьирует от места произрастания и
влажности: жёлто-зелёная на сухом сфагновом покрове, тёмно-зелёная в сильно
увлажнённых участках. Надземные побеги не имеют опушения, имеют высоту от 10 до
30 см. Листья очередные, влагалищные, язычковые, имеют форму полуцилиндрической
пластинки, в основании листового обилие трихом. Соцветие кисть имеет от одного
до восьми цветков. Является одним из основных растений торфообразователей на
олиготрофных и мезотрофных болотах (Минаева, 1997; Губанов и др., 2002).

Рис. 14. Шейхцерия болотная (Scheuchzeria palustris)
(Губанов и др., 2002)
На олиготрофных болотах встречаются насекомоядные растения,
т.к. субстрат этих болот беден азотом в доступной для растений форме,
минеральными и органическими питательными веществами. Наиболее типичными
являются представители семейства росянковые (Droseraceae): росянка круглолистная
(Drosera rotundifolia L.) и росянка английская (Drosera anglica Huds.). Росянка круглолистная
(Drosera rotundifolia L.) - многолетнее розеточное насекомоядное травянистое
растение (рис. 15.). Листья у основания в прикорневой розетке имеют практически
округлую форму, около 2 см шириной, черешковые, длиной от 1 до 4 см, имеют
красный оттенок. На верхней стороне и по краям имеются
железистые-волоски-эмергенты на длинных черешках. Они выделяют капли клейкой
жидкости к которым прилипают насекомые и мелкие беспозвоночные животные. Под
действием выделяемых листьями капель с ферментами попавшие в капли они
перевариваются и усваиваются росянкой. Цветки мелкие, белые, собраны в завитки,
которыми заканчиваются безлистные цветоносы имеющие длину 25 см. Плод
удлинённо-овальная коробочка с мелкими веретеновидными светло-бурыми семенами.
Произрастает на верховых болотах, на открытом торфе и сыром песке. В медицине
препараты из росянки применяются как отхаркивающее при заболеваниях дыхательных
путей (хронический бронхит, ларингит, коклюш) (Губанов и др., 2003).

Рис. 15. Росянка круглолистная (Drosera rotundifolia L.) и её лист (Губанов и
др., 2003)
Росянка английская (Drosera anglica Huds.) - травянистое
насекомоядное многолетнее растение с тонким корнем. Имеет прикорневую розетку
листьев. Листья направленны вверх под углом, черешковые, имеют длину около 9
см, имеют ширину от 2 до 5 мм, линейно-клиновидной либо
линейно-обратнояйцевидной формы с затупленным концом. Листовые пластинки на
дорсальной стороне имеют железистые волоски, на которые прилипают насекомые и
мелкие позвоночные, служащие пищей для растения. Их длина увеличивается от
центра к краю пластинки. Росянка английская обычно имеет 2-3 одиночных стебля,
высотой 10-25 см. Цветки находятся на безлиственных цветоносах и почти в 2 раза
превышают листья по высоте. Цветки белые, мелкие, собраны в завитки
Плод-коробочка. Семена около 1 мм длиной, темные, продолговатой формы.
Произрастает на олиготрофных болотах, на обнажениях торфяной залежи, либо на
торфяном ковре. В медицине используется так же, как росянка круглолистная.
(Губанов и др., 2003).
Мхи
Мхи являются низкорослыми растениями высотой от 10 до 20 см,
имеющие один, либо несколько стеблей со спиральным листорасположением. Мхи
являются доминантами в своем ярусе на олиготрофных болотах. Мхи не имеют
корней, альтернативой им служат волосоподобные многоклеточные образования -
ризоиды. На болотах обычно имеются мхи двух порядков: зеленые (бурые) мхи (Bryales)
и сфагновые (белые, торфяные) мхи (Sphagnales). Бриевые (зеленые) мхи (Bryales)
произрастают на минеральных почвах и торфяных болотах, где они формируют
сплошной моховой дёрн в виде зеленых или бурых «ковров» или «подушек». Данные
мхи нуждаются в минеральном питании, по этой причине они произрастают в
основном на эвтрофных болотах. Но могут встречаться на верховых болотах в
местах с хорошим дренированием (Игнатов, Игнатова, 2003). При образовании
торфяных болот зелёные мхи играют роль доминанта в растительном покрове, а
также в сложении торфяной залежи, отчего в основании залежи обнаруживаются слои
гипнового торфа. Одними из многочисленных бриевых мхов на олиготрофных болотах
являются Амблистегиевые (Amblystegiaceae). Более
распространены водные и болотные мхи самого крупного рода Амблистегиевых -
Дрепанокладуса (Drepanocladus) (Фёдоров, 1978).
Дрепанокладус плавающий (Drepanocladus fluitans, Warnstorfia fluitans (Hedw.) Loeske) - данный мох
имеет лежачий либо плавающий, редко восходящий стебель, длиной от 20 до 30 см,
образует мягкие, рыхлые дерновинки буровато-зеленой или бурой окраски. Их
листья прямые и серповидно-согнутые, длина их достигает до 4-6 мм. В основании
они ланцетные или яйцевидно-ланцетные, постепенно вытянутые в узкий и длинный,
почти шиловидный кончик. Произрастает в мочажинах и топях переходных и верховых
сфагновых болот, по берегам озер и небольших рек, в канавах (Абрамов, Волкова,
1998).
Политрихум сжатый или Кукушкин лён (Polytrichum strictum
Brid.) (Polytrichum juniperinum var. gracilius Wahlenb.) - мох, имеющий
густые дерновинки, довольно высокие, от сизовато-зелёной до буроватой окраски
(рис. 16.). Стебель тонкий, покрыт до листьев чехлом густого и плотного,
желтовато-белого или буроватого ризоидного войлока. Его листья
узко-ланцетно-шиловидные, до 5-7 мм длиной, заострены, имеют широкое желтоватое
влагалищное основание, с широко загнутыми, беловатыми, пленчатыми, цельными
краями. Жилка выступает красно-бурым пильчатым острием, вверху на спинке с
зубчиками (Лаврова, Лебедева, 1979).
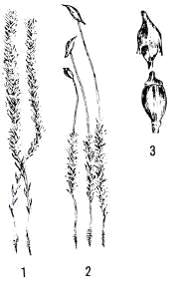
Рис. 16. Политрихум сжатый или Кукушкин лён (Polytrichum strictum
Brid. (Polytrichum juniperinum var. gracilius Wahlenb.) 1 - мужское растение; 2 - женское растение со спорогонами; 3 -
коробочки (Лаврова, Лебедева, 1979)
Сфагновые (торфяные, белые) мхи (Sphagnidae) (рис. 17). Данные мхи
являются растениями-доминантами и эдификаторами на олиготрофных болотах.
Эдификатор (aedificator - строитель) - вид растений с выраженными средообразующими
свойствами, таким образом они определяют строение и видовой состав фитоценоза.
Являются типичными ацидофилами и требуют повышенной кислотности субстрата,
некоторые виды даже при малой концентрации подвижных соединений кальция
(Ефремов, Ефремова, 2000; Инишева, 2009).

Рис. 17. Сфагновые мхи: 1 - сфагнум магелланский (Sphagnum magellanicum); 2 - сфагнум бурый (S. fuscum); 3 - сфагнум ленский (S. lenense); 4 - сфагнум Онгстрёма
(S. aongstroemii); 5 - сфагнум балтийский (S. Balticum); 6 - сфагнум дубравный
(S. Nemoreum); 7 - сфагнум оттопыренный (S. squarrosum); 8 - сфагнум береговой
(S. riparium); 9 - сфагнум прорезной (S. perfoliatum) (Инишева, 2009)
Сфагновые мхи имеют наименьшую потребность в минеральном
питании. Они используют минеральные вещества, которые поступают из атмосферы на
моховой «ковёр». Эти мхи имеют довольно высокую степень влагоёмкости, благодаря
которой некоторые сфагны могут поглощать воды в 20-30 раз больше своего веса в
сухом состоянии. Высокая влагоёмкость обусловлена тем, что ткани сфагновых мхов
состоят в основном из мертвых гиалиновых клеток, среди которых находятся мелкие
живые хлорофиллоносные клетки. Из мертвых гиалиновых клеток состоит и наружный
покров (гиалодермис) стеблей и ветвей. Сплошные «ковры» состоят из
переплетённого множества стеблей растений, благодаря чему сфагновые мхи
понижают испарение воды и удерживают её в себе продолжительное время. При
повышении температуры гиалиновые клетки теряют воду и становятся пустыми, но
при этом мох не погибает. При первом же даже небольшом повышении влаги
гиалиновые с лёгкостью вновь заполняются водой. Из-за способности мхов
накапливать значительное количество влаги «ковры» имеют способность большую
роль в формировании гидротермического режима олиготрофных болот. Род Sphagnum подразделяется на 5
секций, виды которых морфрологически и анатомически различаются между собой: Sphagnum, Squarosa, Subsecunda, Cuspidata, Acutivolia. (Инишева, 2009).
Сфагновые мхи верхового типа требуют повышенной кислотности субстрата. Так как
сфагновый мох корней не имеет, минеральное питание в виде бедных солями водных
растворов воспринимается всей поверхностью растения через гиалиновые клетки.
Поселяясь на заболачивающихся местах, сфагнум уже сам, в условиях достаточно
влажного климата, ускоряет образование болота в этом месте (Лапшина, 2003). По
трофности сфагновые мхи делят на 4 экологические группы:
Ортоолиготрофофиты - мхи, обитающие на критически бедных
минеральными элементами субстратах, как исключение их можно наблюдать на более
богатых минеральным питанием почвах. Данные сфагны получают только атмосферное
водное питание в типичной среде обитания, имеют низкую степень зольности - от 1
до 3%. Зольность - это отношение массы твердогонеорганического остатка,
образующегося после сгорания топлива в определенных условиях, к массе сухого
вещества в топливе (ГОСТ Р 54224-2010). Примеры мхов-ортоолиготрофофитов: Sphagnum angustifolium, S. balticum, S. fallax, S. fuscum, S. girgensochnii.
Мезоолиготрофофиты - данные мхи растут на бедных субстратах,
но больше требовательны к минеральному питанию, чем ортоолиготрофофиты. Примеры
мхов-мезоолиготрофофитов: S. centrale, S. jensenii, S. russowii.
Мезотрофофиты - мхи, имеющие умеренную потребность к
концентрации минерального питания в субстрате. Их зольность составляет 3-7%.
Примеры мхов-мезотрофофитов: S. aongstroemii, S. fimbriatum, S. flexsuosum, S. lenense, S. majus, S. palustre, S. riparium, S. wulfianum.
Мезоэутрофофиты - данным мхам для роста и развития необходимо
значительное количество минерального питания в субстрате. Их зольность
колеблется в пределах 7-10%. Примеры мхов-мезоэутрофофитов: S. cappilifolium, S. compactum, S. contortum, S. magellanicum, S. obtusum, S. papillosum, S. planthuphyllum, S. rubellum, S. squarossum, S. subsecundum, S. teres, S. warnstrofii (Бабешина, Зверев,
2010).
Различные виды сфагнов произрастают в различных условиях
обитания: мочажинах, топях, грядах. Виды сфагновых мхов, растущих на мочажинах:
Sphagnum balticum (Russ.) Russ.ex C. Jens, Sphagnum lindbergii Schimp. ex Lindb, Sphagnum jensenii Lindb, Sphagnum cuspidatum Ehrh. ex Hoffm., Sphagnum obtusum
Warnst. К сфагнам растущим в
рямах относятся следующие виды: Sphagnum angustifolium (Russ. ex Russ.) C. Jens, Sphagnum fuscum (Schimp.) Klinggr, Sphagnum magellanicum Brid. Сфагновые мхи
произрастающие в топях: Spagnum compactum DC. in Lam. et DC, Sphagnum fallax (Klinggr.). На грядах
преимущественно произрастает Sphagnum fuscum (Shimp) Klinggr.
(Фёдоров, 1978). Остановимся подробнее на некоторых видах сфагновых мхов,
произрастающих на олиготрофных болотах.
Сфагнум бурый (Sphagnum fuscum (Shimp) Klinggr.) (см. рис. 17.) - это
растение обладает высокой засухоустойчивостью, жизнеспособно даже в длительные
и периодические засухи. Морфологически выглядит, как густая подушковидная
дернина из множества зеленоватых или ржаво-бурых дерновинок. Является типичным
обитателем олиготрофных болот, произрастает на открытых местах с хорошей
освещённостью. Формирует плотные ковры или гряды. Двудомный, спорофиты редки
(Игнатов, Игнатова, 2003).
Сфагнум балтийский (Sphagnum balticum (Russ) J. lens.)
(см. рис. 17.) - обитает на верховых болотах в обводнённых мочажинах возле
поверхности и на поверхности сплошным «ковром». Образует рыхлый дёрн из мягких
дерновинок желтоватой либо бледно-желто-зеленой окраски. Стеблевой гиалодермис
состоит из 2-3 либо 5 слоёв. Стеблевые листья по форме напоминают равнобедренный
треугольник, имеют длину от 0,9 до 1,2 мм и ширину 0,5-0,8 мм. Засохшие листья
имеют серповидно-согнутую форму, их длина составляет 1-1,7 мм, а ширина 0,3-0,6
мм. Наружная поверхность листа в верхней части имеет водоносные клетки с
мелкими порами, на верхних и нижних концах клеток и в боковых углах, а на
внутренней стороне - крупные округлые поры в углах и в средней части клеточных
стенок (Лаврова, Лебедева, 1979).
Сфагнум узколистный (Sphagnum angustifollium (Russ.) C. Jens (= S. recurvum subsp. angustifollium Russ.) - имеет дерновинки
зелёной, буроватой или желтоватой окраски. Стеблевой гиалодермис плохо заметен.
Склеродермис зеленоватой или желтоватой окраски. Листья стебля имеют длину от
0,4 до 0,8 мм, ширина практически равна длине (иногда бывает чуть шире). Форма
листа варьирует от треугольно-языковидной до языковидной, может присутствовать
небольшое закругление. Верхушка листа имеет небольшую бахромчатость. Веточные
листья имеют узко-ланцетную форму, плавно заужены к концу. Данный мох произрастает
на таких ландшафтах, как кочки, гряды, мочажины (открытые и заросшие лесом),
олиготрофные и комплексные болота, берега зарастающих водоёмов, заболоченные
хвойные и смешанные лесные массивы (Абрамов, Волкова, 1998).
Сфагнум остроконечный. (Sphagnum cuspidatum Ehrh. ex Hoffm.) - данный вид сфагнов
имеет большой высоты дерновинки, рыхлые по структуре, слегка жёсткие на ощупь,
имеющие зеленую, желто-зеленую, либо слегка бурую окраску. Стеблевой
гиалодермис состоит 2-3 слоёв, с хорошо выраженным отграничением. Окраска
склеродермиса желтовато-зеленая. Длина стеблевых листьев варьирует от 1-1,5 до
2 мм, ширина от 0,5 до 0,7 мм. Форма стеблевой листовой пластинки имеет
очертания равнобедренного треугольника, вогнута внутрь, верхушка зубчатая.
Веточные листья имеют длину от 2 до 4 мм. Встречаются несколько вариантов их
формы: от продолговато-ланцетных до шиловидных. Обычно на верху листа имеются
цельные, либо зубчатые загнутые края. Водоносные клетки на дорсальной
поверхности веточного листа имеют мелкие поры, с меньшей частотой в углах
клеточных стенок, местами с боковыми угловыми кольчатыми порами. Растёт на
комплексных олиготрофных болотах среди глубоких мочажин (является эдификатором
мочажинных комплексов), на берегах ламб (зарастающих лесных озёр) и прочих углублениях
типа ям и канавок в торфе (Абрамов, Волкова, 1998).
Сфагнум магелланский (Sphagnum magellanicum Brid. (S. media Limpr.) (см. рис. 17.) -
многолетний мох имеющий окраску дерновинок от красноватой до
пурпурно-фиолетовой, иногда встречаются желтоватая или сизо-зелёная окраска.
Стеблевой склеродермис имеет окраску от красноватой до пурпурной, изредка
светло-зелёную. Форма стеблевых листьев языковидно-лопаточковидная, к верхушке
имеется расширение с порами водоносных клеток, длина варьирует от от 0,8 до 2
мм, ширина от 0,5 до 0,8 мм. Произрастает на олиготрофных и мезотрофных болотах
в хорошо дренируемой части среди древесной растительности, на
грядово-мочажинных фитоценозах, мочажинах, по берегам зарастающих водоёмах, в
заболоченных смешанных и хвойных лесных массивах (Лисс и др., 2001; Мульдияров,
1990).
Сфагнум береговой (Sphagnum riparium Aongstr.) (рис. 18.) -
один из мхов с крупными размерами. Данный мох имеет стебельки зелёной окраски и
практически неограниченным ростом. Имеет по 4-5 веточек в пучке, 2-3 из них
отходят от стебля под углом. Характерно что стебельки ломаются со своеобразным
треском. Апекс на концах стеблей имеет крупные размеры. Стеблевой гиалодермис
развит неравномерно, имеет 2-3 слоя. Склеродермис имеет бледно, либо
жёлто-зелёную окраску. Стеблевые листья имеют языковидную либо
треугольно-языковидную форму с разрезом по средине (из-за разрушения стенок
водоносных клеток), что является видовым признаком. Их длина разнится от 1,2 до
1,4 мм, ширина до 1 мм. Веточные листья до 2,2 мм длиной яйцевидно-ланцетной
формы сухие, слегка волнистые, имеют отогнутые назад верхушки. Произрастает по
обводнённым окраинам болот различных типов и мочажинам, по берегам водоёмов,
сплавинах, в канавах и небольших искусственных углублениях почвы. На олиготрофных
болотах этот мох может формировать густые дернины, которые из-за сильной
обводнённости практически невозможно преодолеть. (Мульдияров, 1990).

Рис. 18. Сфагнум береговой (Sphagnum riparium Aongstr.) [#"869925.files/image020.jpg">
Рис. 19. Кладония оленья (Cladonia rangiferina (L.) F.H. Wigg.) (Голубкова, 1966)
Кладония звёздчатая (Cladonia stellaris (Opiz) Pouzar & Vezda) - лишайник с
горизонтальным накипным долгосрочным таломом. Габитус ортотропный,
кустисто-разветвленный. Ветвление плотное, изотомическое либо политомическое;
поверхность паутинистая или слегка «войлочная». Апотеции имеют коричневый цвет
(встречаются с редкой частотой), Пикнидии с красным содержимым находятся на
концах апикальных веточек. Подеции формируют хорошо выраженные кругловатые,
куполовидные кустики, высота которых варьирует от 5 до 10, иногда от 20 до 30
см. Их окраска разнится от беловато-серой до желтовато-серой, иногда бывает
беловато-желтоватой, в нижней части более темная. Ареал обитания этого вида
лишайника довольно разнообразен. Кладония звёздчатая произрастает обычно на
торфе и песке в открытых, хорошо просвечивемых Солнцем местах, на валежнике и
гнилых пнях, сухих участках сосновых зеленомошных боров, лесах, на олиготрофных
болотах. Используется для получения усниновой кислоты и как средство по борьбе
с микробами (Мучник и др., 2011; Sokolov и др., 2012).
Кладония роговидная (Cladonia cornuta (L.) Hoffm., Schaer) - данный лишайник имеет
горизонтальное первичное слоевище из мелких, волнисто-рассечённых по краям
чешуек длиной от 3 до 8 мм, которые исчезают с возрастом. Базальные чешуйки
продолговатой, неправильно-выемчатой формы, приподнимающиеся, с
серовато-зеленоватой или оливково-зеленоватой окраской. Подеции образуют
сцифовидно-разветвленные, слаборазветвлённые и неразветвленные формации. Высота
подециев колеблется от 1 до 10, иногда 16 см. Внешне они простые, реже
встречаются расходящиеся на 2-3 ответвления в верхней части, имеющие мучнистый
налёт из соредий. Коровый слой в виде бугорков находящийся в средней и нижней
частях подециев, имеет коричневато-зеленоватую окраску. Апотеции формируются на
концах подециев Соредии сероватые, серовато-коричневатые, развиваются лишь в
верхней части подециев. Данный лишайник обитает преимущественно на сухих грядах
олиготрофных болот, гнилой древесине, пнях, валежнике, повышениях у основания
стволов деревьев, на песке, сограх, темнохвойных и смешанных лесных массивах,
сухих участках сосновых боров, мшистых валунах и скалах в различных
сообществах. Является кормом для северных оленей (Голубкова, 1966; Мучник и
др., 2011).
Эверния сливовая (Evernia prunastri (L.) Ach) - данный лишайник имеет
дорсивентральный, плосколопастной таллом (рис. 20.). На дорсальной поверхности
имеются беловатые соредии. Лопасти дорсальной стороны имеют окраску различных
оттенков: светло-зелёная, беловато-зелёная, серовато-зелёная, жёлто-зелёная.
Типичный обитатель верховых болот, произрастает на древесном субстрате.
Применяется в парфюмерной промышленности [#"869925.files/image021.jpg">
Рис. 20. Эверния сливовая (Evernia prunastri (L.) Ach)
[#"869925.files/image022.jpg">
Рис. 21. Вульпицида (Vulpicida pinastri (Scop.) J.-E. Mattsson et M.J. Lai.) [http://macroid.ru/showphoto.php? photo=47738]
Таким образом, видовой состав флоры олиготрофных болот не так
богат, как в других природно-территориальных комплексов из-за условий верховых
болот, но численность растительности (кроме верхнего яруса) достаточно велика.
3. Фитоценозы олиготрофных болот
Растительность - совокупность фитоценозов Земли в целом, либо её
отдельных регионов. Является основным компонентом биосферы. Растительность
подразделяется на водную, мезофитную, ксеро- и галофитную, древесную,
кустарниковую, травянистую и др. Растительность играет главную роль в первичном
синтезе органического вещества и играет большую роль в круговороте веществ
(Дедю, 1989). С греческого слово «фитоценоз» переводится, как состоящее из двух
корней: phyton - растение и koinos - общий. Фитоценозы относят к
растительным сообществам, суммарную численность флоры, приуроченных к
относительно однородной площади (Анучин и др., 1986). Термин «фитоценоз»
применим, как к растительному покрову, в общем к лесному болоту, так и к общей
сумме растений-торфообразователей на отдельном микрорельефе: кочки и мочажины
олиготрофных болот. В большинстве случаев фитоценозы более сложно устроены и
имеют в своем составе несколько растительных ярусов. Данное многообразие
фитоценозов можно классифицировать в определенную систему. Исходя из трофности
среды - это организация из групп растительности, мало требовательных
(олиготрофных), среднетребовательных (мезотрофных) и требовательных (эвтрофных)
к элементам минерального питания (Инишева Л.И., 2009). Также фитоценозом можно
назвать флористический состав болота в общем или совокупность
растений-торфообразователей, произрастающих отдельно на кочках и мочажинах
олиготрофных болот (Тюремнов, 1976). Растительный покров, особенно олиготрофных
болот, пространственно неоднороден, в нем чётко выделяются территориальные
комплексы, представляющие собой закономерные сочетания фитоценозов (Инишева и
др., 2000).
После определения ассоциаций их объединяют в группы, а группы в
формации. Формации - ассоциации с одним общим растением-доминантом. Формации
объединяют в группы и классы формаций, а уже их в типы растительности, что
обозначает объединение флористических сообществ с общими биологическими и
экологическими особенностями растительности (Александрова, 1969). Доминирование
сфагновых мхов на верховых болотах является их олиготрофность и высокая
концентрация органических кислот. Эти факторы длительно поддерживают
постоянство гидрохимических условий, отчего формируется постоянство
растительного покрова современных фитоценозов. Постоянная высокая обводнённость
и высокая кислотность среды, формированию которой способствуют доминирующие
сфагновые мхи, создают условия для высокой степени подвижности ионов,
попадающих из атмосферы или в результате водотока. Таким образом, из-за стока
верховых болот, эти ионы со временем сбрасываются. Это явление сохраняет
олиготрофность среды в центрах болотных систем, а в периферийных и пониженных
участках, которые находятся между повышениями - формирует условия для
эвтрофизации. Отчего в данных понижениях формируются мезотрофные либо эвтрофные
топи. Из-за данных факторов большие выпуклые системы болот обладают свойством
химического самоочищения от загрязнений различного генезиса. В мелких и плоских
болотах изменение минерализации приводит к большей или меньшей смене
флористического состава (Лисс и др., 2001).
Исходя из изложенного флористический состав болот (единичные
фитоценозы либо их комплексы) создаёт возможность использовать его, как
индикатор строения торфяной залежи, её глубины, если, вместе с составом и структурой
фитоценозов, при этом учитывая такие факторы, как:
. географическое и геоморфологическое положение болотных систем;
. величину и местонахождение фитоценоза-индикатора в пределах
нужной болотной системы;
. динамический тип фитоценоза-индикатора: сформированный или
формирующийся (Лисс и др., 2001).
Отдельно взятые фитоценозы являются индикаторами типов водного
питания болота..К примеру, флористический состав болот, питающихся грунтовыми
водами, образовавшийся в зоне тайги на притеррасных пойменных участков,
отличается доминированием сосново-елово-кедрово-берёзово-осоковыми (травяными)
фитоценозами, которые называются «сограми». На центральных участках пойм данной
зоны признаками преобладания грунтового питания являются ивовые,
ивово-осоковые, осоковые фитоценозы. Примечательно, что индикаторами
атмосферного питания являются любые фитоценозы и их комплексы, относящиеся к
олиготрофному типу. Фитоценозы сосново-кустарничково-сфагнового типа (рямы) и
фускум-фитоценозы в северной половине подзоны средней тайги и в северной тайге
являются индикаторами преимущественно фускум-залежи (Лисс и др., 2001).
Постоянно принимаются попытки слияния разных методических школ и синтезирование
единой классификации, что в реальности невозможно достигнуть. В настоящее время
многие исследователи отмечают, что любые классификации имеют право на
существование, так как каждая из них является достаточно условной и создается
для решения определенных задач. Проблема выведения единой классификации
растительных сообществ болот определяется большим объёмом флористических форм в
их составе и значительными разницами в размерах видов эдификаторных сообществ -
древесных растений больших размеров до мелкой мшистой растительности. Этот
фактор является одним из главных. Именно он обусловливает варьирование размеров
площадей болотных сообществ, это вариация от нескольких квадратных метров (мхи)
до тысяч километров квадратных (древесные растения). Характерно, что при этом
сохраняются главные свойства растительных сообществ: продолжительная устойчивость
и способность к регенерации системы (Кузнецов, 2007). Основываясь на сказанном
выше, можно прийти к выводу, что единая классификация болотных фитоценозов,
включающая все важные ландшафтные составляющие вместе (растительность, торфяные
залежи, ландшафтное положение водно-минерального питания) будет груба и
некорректна. В основу классификаций болотных сообществ положен комплекс
динамических, экологических и фитоценотических признаков (Александрова, 1969).
Существуют различные классификации болотных фитоценозов, разработанные в XX веке.
Первая обзорная классификация отмечала детально особенности
флористического состава болот бывшего СССР и носила название
эколого-фитоценотической. Её автором был Цинзерлинг Ю.Д. в 1938 году. В её
основание он положил принципы, предложенные Шенниковым А.П. (1935,
1938). Она охватывает 9 типов растительности, которые в свою очередь
разделены на группы формаций и формации, а они в свою очередь на группы
ассоциаций и ассоциации. Данные синтаксоны выделяют по признакам растительности
с учетом флористического состава фитоценозов, количественной разницы между
видами, соотношением видов доминантов в каждой из ярусов (Миркин и др., 2000;
Вебер и др., 2005). На олиготрофных болотах встречаются разнообразные
фитоценозы, состоящие из девяти типов растительности: лесной, кустарниковый,
кустарничковый, травяной, гидрофильно-моховой, психрофильно-моховой,
печёночный, лишайниковый, водорослевый. Из них три: гидрофильно-моховой,
психрофильно-моховой и печёночный на сегодняшний день объединяют в моховой тип,
так как они сформированы одной жизненной формой. Подтипами являются такие
фитоценозы, как: сосново-кустарничковые, сосново-пушициевый, сосново-сфагновый,
пушициевый, шейцхериево-сфагновый и ангустифолиум, фускум, магелланикум,
грядово-мочажинный, грядово-озёрный, мозаично-сфагновый, озёрно-денудационный.
Помимо типов и подтипов выделяются группы фитоценозов: древесная,
древесно-травянистая, древесно-моховая, травяная, травяно-моховая, моховая
(Тюремнов, 1976; Денисенков, 2000).
Вторая классификация, разработанная в Центральной Европе
(школа Браун-Бланке) была эколого-флористическая классификация, основанная на
применении всего флористического состава фитоценозов и групп типичных
фитоценотических видов. В данной классификации при выделении иерархии
синтаксонов (классы, порядки, союзы, ассоциации) в разной степени учитываются
также физиономические признаки сообществ и экологические свойства фитоценозов.
Есть чёткая система синтаксонов и руководство по их выделению и описанию
(Миркин и др., 2000; Вебер и др., 2005).
Топологические (тополого-экологические) классификации
основаны на рассредоточении фитоценозов и ассоциаций в сети координат. Осями
этих систем служат условия среды: влажность и минеральное питание субстрата
(почвы) мест произрастания. Система координат может быть многомерной, например,
с учетом степени интенсивности движения проточных вод фитоценоза (Воробьев,
1953). Среди фитоценозов олиготрофных типов и их комплексов надёжными
индикаторами строения и глубины торфяной залежи являются:
сосново-кустарничково-сфагновые, сосново-пушицево-сфагновые,
сосново-осоково-сфагновые, грядово-мочажинные, озерково-грядово-мочажинные,
озерково-денудационные (Лисс и др., 2001). В классификации торфяно-болотных
фитоценозов растительный покров подразделяется на крупные единицы, которые
отражены в единицах классификации торфов, которые исходят из пластообразующих
видов торфа (Пьявченко, 1963). Данная классификация - эколого-топологическая,
получила развитие в работе Тюремнова С.Н. 1976 года, где он разделяет классы
формаций относят в ранг типов болотной растительности, следовательно
растительность болот слагается тремя типами: эфтрофным, мезотрофным,
олиготрофным. Вторая ступень этой классификации - подтип, который выделяют по
степени увлажнения (слабо-, средне- и сильно увлажнённые). Последней ступенью
классификации являются «болотные фитоценозы», которые в таксономическом
отношении соответствуют группам ассоциаций, а иногда и более крупных таксонов
(формациям). Рассмотрим некоторые из фитоценозов, наиболее часто встречающиеся
на олиготрофных болотах.
Сосново-кустарничково-сфагновые фитоценозы. В болотоведении
сибирские олиготрофные болота с сосново-кустарничково-сфагновыми группами
формаций принято называть рямами Рямы подразделяются в зависимости от высоты
сосны (Лисс и др., 2001; Инишева и др., 2003). Сосново-кустарничково-сфагновые
сообщества - характеризуются прежде всего разреженным древесным ярусом. Средняя
сомкнутость крон в данном фитоценозе варьирует от 0,2 до 0,3 м. Древесный ярус
преимущественно составляет сосна обыкновенная (Pinus sylvestris), а
именно её болотные формы: f. uliginosa, f. litwinowii,
f. willkommii), с меньшей периодичностью встречается береза пушистая (Betula
pubescens). Данный ярус характеризуется резко колеблющимися высотами деревьев:
от 3 до 12 м. Толщина стволов здешних деревьев варьирует от 10 до 18 см.
Травяно-кустарничковый ярус хорошо развит и покрывает от 40 до 60 (65)% площади
фитоценоза. Высоты данного яруса колеблются в пределах 25-50 см. Травянистых
растений в этом ярусе мало, но в отдельных фитоценозах доминируют осока
шаровидная (Carex globularis) или пушица (Eriophorum vaginаtum), политрихум сжатый (Polytrichum
strictum) (Храмов, Валуцкий, 1977). Сфагновый покров в моховом ярусе развит
хорошо, местами он формирует почти сплошной «ковёр». Покрытие мохового яруса в
данном фитоценозе соотношение колеблется от 30 до 90%. Моховой покров имеет
мелкоконтурную мозаичность с размытыми краями пятен. В этом фитоценозе средняя
численность видового разнообразия, где на 100 м²
приходится
15-16 видов. (Лисс, Березина, 1981).
Доминирующим видом в моховом покрове на повышениях является Sphagnum fuscum (95%), на межкочкарных
понижениях встречаются Sph. angustifolium и Sph. magellanicum (Головацкая, 2009).
Для рассматриваемых сообществ средняя видовая насыщенность
составляет 16-17 видов на 100м2, включая цветковые растения, мхи и
лишайники. Доминируют бореальные виды: Pinus sylvestris, Ledum palustre,
Chamaedaphne calyculata, Sphagnum fuscum, Sphagnum angustifolium, Polytrichum
strictum, Pleurozium schreberi. При содоминировании в моховом покрове
зеленых мхов выделены сосново-плевроциево - политрихумово-кустарничковые
сообщества (Тюремнов С.Н., 1957).
Пушициевые фитоценозы располагаются на хорошо дренируемых
окраинах олиготрофных болот в районах европейской части России, местами
развиваются небольшими участками пушициевые кочкарники Плотные кочки пушицы, имеющие
высоту от 20 до 30 см и диаметром до 30 см занимают примерно 50% площади
фитоценоза. Торфяная залеж между кочками сильно уплотнена на поверхности,
отчего талые и дождевые воды застаиваются в этих понижениях долгое время. После
высыхания воды они зарастают сильно разреженным моховым «ковром» из Sphagnum magellanicum и Sphagnum angustifolium, которые несильно
прикрывают почву, иногда торфяная залеж остаётся непокрытой мхами. Данные
участки безлесны. Кустарнички вересковых здесь встречаются редко (Пьявченко,
1963).
Фускум-фитоценозы имеют повышенную влажность субстрата
(92-93%). Sphagnum fuscum образует большие дерновины, которые покрывают болота объёмными
плоскими подушками, имеющие высоту до 30 см и имеют большую влагоёмкость. Из-за
повышенной влажности и плотности мохового «ковра» не создаются благоприятные
условия для произрастания древесного яруса, отчего здесь произрастают
угнетённые формы сосны: f.pumila, f. Willkommii, высота которых
варьирует от 1 до 3 м. Кустарничковый покров развит слабо, состоит
преимущственно из голубики (Vaccinium uliginosum) рее водяники (Empetrum
nigrum), подбела (Andromeda polifolia), клюквы четырёхлепестной (Oxucoccus
quadripetalus). Более чаще встречается клюква мелкоплодная (Oxucoccus
microcarpus). В травяном ярусе доминантом является пушица (Eriophorum
vaginatum), встречается морошка (Rubus chamaemorus). Почти 100% мохового
покрова составляет Sphagnum fuscum с малой примесью Sphagnum
angustifolium и Sphagnum magellanicum у основания кочек. Данные
фитоценозы находятся на восточном макросклоне Урала и Западной Сибири
(Тюремнов, 1976).
Выводы
. Олиготрофные болота - это тип болот, водно-минеральное
питание которых осуществляется за счет атмосферных осадков. Олиготрофные болота
формируются обычно на водоразделах и верхних террасах речных долин при
заболачивании лесов, лесных вырубок и пожарищ, а также при зарастании и
заторфовывании водоемов (преимущественно непроточных озёр). Наиболее активно
процесс образования болот протекал 10-12 тысяч лет назад в голоценовый период.
. Олиготрофные болота характеризуются следующими
экологическими условиями для произрастания растений: сильная обводненность и
связанные с этим слабая аэрация, низкая теплопроводность, крайне низкое
содержание элементов минерального питания, повышенная кислотность, постоянное
нарастание дернины из мхов-сфагнумов.
3. Выявлено, что растения олиготрофных болот имеют ряд
биоморфологических особенностей, связанных с их местом обитания. Все растения
олиготрофных болот - ацидофилы. Общими чертами растений олиготрофных болот
является: вечнозелёность листовых пластин в совокупности с опушением, либо
восковым налётом, ксероморфность, постоянный рост вверх горизонтальной корневой
системы в «деятельном» слое мохового «ковра» и постоянном отмирании её нижних
погребённых участков. Вследствие дефицита кислорода в корневых системах
болотной растительности (особенно у трав) хорошо развились системы
воздухоносных ходов и полостей, которые получают кислород через диффузию от
наземных органов. Так, например сфагновые мхи являются эдификаторами
олиготрофных болот. Ткани листьев содержат мертвые гиалиновые клетки, способные
поглощать воду в 20-30 раз больше собственного веса.
. Установлено, что в настоящее время единой классификации
растительности болот нет. Согласно эколого-фитоценотической классификации
выделяют 9 типов, 12 подтипов, 6 групп фитоценозов. На олиготрофных болотах
флористическое разнообразие значительно беднее, чем на эвтрофных. Отличительной
особенностью олиготрофных болот является обилие и видовое разнообразие сфагновых
мхов.
Литература
1. Абрамов И.И., Волкова Л.А. Определитель
листостебельных мхов Карелии // Бриологический журнал. - Т.7. - Прилож/1. изд.:
KMK Scientific Press Ltd, 1998. 390 с.
2. Абрамова Т.Г., Боч М.С., Галкина Е.А.
Типы болот СССР и принципы их классификации. Л.: Наука, 1974. 260 с.
. Аветов Н.А., Шишконакова Е.А. Бюллетень
Почвенного института им. В.В. Докучаева. 2013. Вып. 71 36 УДК 631.427.12
Понятие трофности в связи с антропогенной эвтрофикацией верховых болот
Ханты-Мансийского Приобья, 2013. с. 36-49
. Александрова В.Д. Классификация
растительности - Изд-во. Л.:Наука, 1969 г. 275 с.
. Анучин Н.А., Атрохин В.Г., Виноградов
В.Н. / Лесная энциклопедия: В 2-х т., т. 2/ Гл.ред. Воробьев Г.И.; Ред.кол.:
Анучин Н.А., Атрохин В.Г., Виноградов В.Н. и др. - М.: Сов. энциклопедия, 1986.
631
c.
. Арсланова Х.А., Голубчина М.Н.,
Искандерова А.Д. и др, Геологический словарь: в 2-х томах/; под ред. К.Н.
Паффенгольца. - 2-е изд., испр. - М.: Недра, 1978. 400 с.
. Бабешина Л.Г. Сфагновые мхи
Западно-Сибирской равнины: морфология, анатомия, экология и применение в
медицине: Автореферат диссертации на соискание ученой степени доктора
биологических наук. ГБОУ ВПО «Сибирский государственный медицинский
университет» Министерства здравоохранения и социального развития Российской
федерации. Томск, 2011. 38 с.
. Бабешина Л.Г., Зверев А.А. Оценка
условий местообитаний сфагновых мхов Западно-Сибирской равнины: фактор
трофности // Вестник Томского Государственного Университета №338, Сентябрь
2010. с. 188-194.
. Баркман Н. Современные представления о
непрерывности и дискретности растительного покрова и природе растительных
сообществ в фитосоциологической школе Браун-Бланке // Ботанический журнал,
1989, т. 74 №11. 37 с.
. Богдановская-Гиенэф И.Д. О болотах
плейстоцена // Ботанический журнал, 1972, №6, с. 615-621.
. Боч М.С., Смагин В.А. Флора и
растительность болот Северо-Запада России и принципы их охраны. СПб., 1993. 233
с.
. Быков Б.А. Введение в фитоценологию.
Алма-Ата: Наука, 1983 с. 215
13. Вебер Х.Э., Моравец Я., Терийя Ж.-П. 2005.
Международный кодекс фитосоциогической номенклатуры. З-е изд. // Растительность
России. №7. СПб. С. 3-39.
. Волкова Е.М., Вислогузова [Зацаринная]
Д.В., Барбашов Е.Р. О разнообразии болот Белевского района Тульской области //
Исследования природы Тульской области и сопредельных территорий: cб. науч. тр.
Вып. 1. Тула, 2008. С. 149-162
15. Волкова Е.М., Формы сосны на болотах
Тульский государственный педагогический университет им. Л.Н. Толстого // -
Издательский дом «Первое сентября», Журнал N29 (564), 2000
. Вомперский, С.Э. Биосферное значение
болот в углеродном цикле // Природа. - 1994. - №7. - с. 44-50.
. Воробьев Д.В. Типы лесов европейской
части СССР. Киев, 1953. 448 с.
. Вылцан Н.Ф. Определитель растений
Томской области. - Томск: Изд-во Том. Ун-та, 1994. С. 103
19. Глебов Ф.З. (ред.) Комплексная оценка болот
и заболоченных лесов в связи с их мелиорацией - Новосибирск.: Наука, 1973. 238
с.
20. Головацкая Е.А. Биологическая
продуктивность олиготрофных и эвтрофных болот южнотаежной подзоны Западной
Сибири / Journal
of Siberian Federal University. Biology 1. 2009 г. С. 38-53
21. Голубкова Н.С. Определитель лишайников
средней полосы европейской части СССР. М.-Л.: Наука, 1966. 257 с.
. Гроздова Н.Б., Некрасов В.И.,
Глоба-Михайленко Д.А. Деревья, кустарники и лианы / изд.: Лесная
промышленность, 1986. 353 с.
. Губанов И.А., Киселёва К.В., Новиков
В.С., Тихомиров В.Н. Иллюстрированный определитель растений Средней России. \
М.: Т-во научных изданий КМК, Ин-т технологических исследований Т. 1., 2002.
526 с.
. Губанов И.А., Киселёва К.В., Новиков
В.С., Тихомиров В.Н. Иллюстрированный определитель растений Средней России. \
М.: Т-во научных изданий КМК, Ин-т технологических исследований Т. 2., 2003 г.
665 с.
. Губанов И.А., Киселёва К.В., Новиков В.С.,
Тихомиров В.Н. Иллюстрированный определитель растений Средней России. \ М.:
Т-во научных изданий КМК, Ин-т технологических исследований Т. 3, 2004. 520 с.
26. Дедю И.И. Экологический энциклопедический
словарь. Кишинев: Главная редакция Молдавской советской энциклопедии, 1989. 406
с.
. Денисенков В.П. Основы болотоведения:
Учеб. Пособие. - СПб.: Изд-во С.-Петерб. Ун-та, 2000. 224 с.
28. Еленевский, А.Г., Соловьёва М.П.,
Тихомиров В.Н. Ботаника. Систематика высших, или наземных, растений: 4-е изд.,
испр. - М.: Академия, 2006. 456 с.
. Ефремов, С.П., Ефремова Т.Т. Строение и
продуктивность сообществ сфагновых мхов на болотах Западной Сибири [Текст] /
С.П. Ефремов, Т.Т. Ефремова // Сиб. экологический журн., 2000, №5. с. 615 - 626
. Жизнь растений в 6 томах, главн. ред.
член-корреспондент АНСССР, профессор А.А. Федоров под ред. проф. М.Н.
Голлербаха., - М; Просвещение, 1997. 504 с.
. Игнатов М.С., Игнатова Е.А. Флора мхов
средней части европейской России. Sphagnaceae - Hedwigiaceae. М., 2003. Т. 1.
608 с.
. Инишева Л.И. Болотоведение: учебник для
вузов /Л.И. Инишева; ГОУ ВПО «Том. гос. пед. университет». - Томск:
Издательство Томского государственного педагогического университета, 2009. 210
с.
33. Инишева Л.И., Моторин А.С. Происхождение
торфяных болот и их многофункциональная роль. Томск, изд-во ЦНТИ, 2000. 59 с.
. Козловский Е.А. Горная энциклопедия. -
М.: Советская энциклопедия. Под редакцией Е.А. Козловского. 1984-1991. 436 c.
35. Красная книга Воронежской области: в 2 т.
Т. 1: Растения. Лишайники. Грибы / Правительство Воронеж. обл.; Упр. по экол. и
природопользованию Воронеж. обл.; Воронеж. гос. ун-т.; [науч. ред. В.А.
Агафонов]. - Воронеж: МОДЭК, 2011. 472 c.
. Красная книга Хабаровского края: Редкие
и находящиеся под угрозой исчезновения виды растений и животных: официальное
издание /Министерство природных ресурсов Хабаровского края, Институт водных и
экологических проблем ДВО РАН. - Хабаровск: Издательский дом «Приамурские
ведомости», 2008. 632 с.
. Кузнецов О.Л. Основные методы
классификации растительности болот // Актуальные проблемы геоботаники. III Всероссийская
школа-конференция. Лекции. Петразаводск: КрНЦ РАН, 2007. с. 241-269.
. Лаврова К.Г., Лебедева Н.В., Лантратова
А.С. и др. / Мхи-папоротники - Петрозаводск: Карелия, 1979. 81 с.
. Лапшина Е.Д. Флора болот юго-востока
Западной Сибири. Томск: Изд - во Том. ун - та, 2003. 296 с.
. Лапшина Е.Д., Руденко В.В. Лишайники
олиготрофных болот лесной зоны Западной Сибири // Состояние и перспективы
развития Гербариев Сибири: Тез. докл. Томск, 1997. 83-84 с.
. Лесные и болотные фитоценозы Восточного
Васюганья (структура и биологическая продуктивность) / А.А. Храмов, В.И.
Валуцкий; АН СССР, Сиб. отд-ние, Центр. сиб. ботан. сад; Отв. ред. А.В.
Куминова. Новосибирск: Наука, 1977. 221 с.
. Лисс О.Л., Абрамова Л.И., Аветов Н.А.,
Березина Н.А., Инишева Л.И., Курнишкова Т.В., Слука 3. А., Толпышева Т. 10.,
Шведчикова Н.К. Болотные системы Западной Сибири и их природоохранное значение.
Под ред. д. б. н. профессора В.Б. Куваева. - Тула: Гриф и К°, 2001. 584 с.
43. Лисс О.Л., Березина Н.А. Болота
Западно-Сибирской равнины. - М.: Изд-во Московского ун-та, 1981. 206 с.
44. Лисс О.Л., Астахова В.Г. Лесные
болота.-М.: Лесн. Пром-сть, 1982. 128 с.
45. Малышев Л.И., Пешкова Г.А. Особенности и
генезис флоры Сибири (Предбайкалье и Забайкалье). Новосибирск. 1984. 364 с.
. Минаева Т.Ю. Биологическая флора
Московской области // Вып. 13. 1997. 30-49 c.
47. Минина З.Б. «Болото как экосистема»
Тульский государственный педагогический университет им. Л.Н. Толстого // -
Издательский дом «Первое сентября», Журнал N18 (649), 2002. 9 c.
. Миркин Б.М., Наумова Л.Г., Соломещ А.И.
Современная наука о растительности. - М.: Логос, 2002. 264 с.
. Мульдияров Е.Я. Определитель
листостебельных мхов Томской области. Томск: ТГУ, 1990. 208 с.
. Мучник Е.Э., Инсарова И.Д., Казакова
М.В. Учебный определитель лишайников Средней России. Рязань, Ряз. гос. ун-т,
2011. 360 с.
. Нейштадт М.И. Болотообразовательные
процессы в голоцене // Известия АН СССР, серия географическая. - 1985. - В. 1.
- С. 39-47.
. Окснер А.Н. / Лихенология // Большая
советская энциклопедия: в 30 т. / гл. ред. А.М. Прохоров. Т.14. 3-е изд. - М.:
Советская энциклопедия, 1973 г. 623 c.
. Положий А.В., Ревушкин А.С., Баранова
В.В. Определитель растений юга Томской области: Учебное пособие - Томск: Изд-во
Том. Ун-та, 1985. 244 с.
. Прокопьев Е.П. Экология растений (особи,
виды, экогруппы,
. жизненные формы): Учебник для
биологических факультетов вузов. -
. Томск: Томск. Гос. ун-т, 2001. 328 с.
. Пьявченко Н.И. Лесное болотоведение.
Основные вопросы М.: АН СССР, 1963. 192 с.
. Седельникова Т.С., Муратова Е.Н.,
Ефремов С.П. Сибирский ботанический журнал Выпуск №1 / том 2 / 2000. 73-80 с.
. Сосудистые растения советского Дальнего
Востока. Т. 5. / Отв. ред. С.С. Харкевич. - С.-П.: Наука, 1991. 390 с.
60. Топливо твердое из бытовых отходов.
Определение зольности. ГОСТ Р 54224-2010. М.: Стандартинформ, 2011. 8 с.
. Тимонин А.К. Высшие растения: учебник для
студентов высших учебных заведений. - М.: Академия, 2007. с. 352.
. Тимонин А.К., Нотов А.А. 1993. Большой
практикум по экологической анатомии покрытосеменных растений. Тверь: Изд-во
Твер. ун-та, Ч. 1. 23 с.
64. Тюремнов С.Н. Торфяные месторождения //
М.: Недра, 3-е издание, 1976. 488 с.
. Федоров А.А. Жизнь растений. В 6-ти т.
Т. 4. Мхи. Плауны. Голосеменные растения. / Под ред. И.В. Грушвицкого и С.Г.
Жилина. Гл. ред. чл.-кор. АН СССР, проф. А.А. Федоров. М.: Просвещение, 1978.
447 с.
. Федоров А.А. Жизнь растений. В 6-ти т. /
Т.5. Ч. 1. Цветковые растения / Под ред. А.Л. Тахтаджяна. Москва: Просвещение,
1980. 430 с.
. Фёдоров А.А. (ред.), Тахтаджян А.Л.
Жизнь растений. Том 5, часть 2. Цветковые. Москва: «Просвещение», 1981. 512 с.
. Фёдоров А.А. Жизнь растений: В 6-ти т.
/Гл. ред. А.Л. Тахтаджян. Т.
. 6. Цветковые растения/ Под ред. А.Л.
Тахтаджяна. Москва: Просвещение,
. 1982. 543 с.
. Флора СССР // ред. Комарова В.Л.
Ленинград: Изд-во АН СССР, Т. 18, 1952. 477 с.
. Цинзерлинг Ю.Д. Растительность болот //
Растительность СССР. Москва-Ленинград. Т. 1, 1938. 355 с.
. Шенников А.П. Луговая растительность
СССР // Растительность СССР. Москва; Ленинград. Т. 1, 1940. с. 622
. Шенников А.П. Принципы ботанической
классификации лугов / А.П. Шенников // Сов. ботаника. Вып. №5, 1935. 35-49 с.
. Якушкина, Н.И. Физиология растений
[Текст]: учеб. пособие для биол. спец. высш. пед. учеб. заведений / 2е изд.,
перераб. Москва: Просвещение, 1993. 351 с.
76. Sokolov, D.N.; Luzina, O.A.; Salakhutdinov,
N.F. Usnic acid: preparation, structure, properties and chemical
transformations. - Russian Chemical Reviews. - 2012. - 81. c. 747-768.