Половые клетки и оплодотворение
Контрольная работа
ПОЛОВЫЕ КЛЕТКИ И ОПЛОДОТВОРЕНИЕ
Содержание
1. Женские половые клетки (яйцеклетки)
. Мужские половые клетки (сперматозоиды)
. Оплодотворение и партеногенез
.1 Общая характеристика процесса оплодотворения
и его биологическое значение
.2 Механизмы оплодотворения
. Партеногенез, гиногенез, андрогенез.
Естественный и искусственный партеногенез
. Генетическое определение пола
Литература
. Женские половые
клетки (яйцеклетки)
Приступая к изучению женских
половых клеток, следует хорошо разобраться в особенностях их организации.
Необходимо обратить внимание на их полярность, на формирование кортикального
слоя и на оболочки яйца: их строение, и значение. Особое значение надо уделить
классификации яйцеклеток, основанной на количестве желтка и его распределении в
цитоплазме (ооплазме) клетки. Это важно, так как строение яйцеклетки имеет
существенное значение для дальнейшего развития, определяя типы дробления,
способы гаструляции и др.
У преобладающего большинства
животных яйцеклетки неподвижны и отличаются однообразием формы (чаще всего
округлые).
Количество выделяемых
организмом яйцеклеток, так же как и размеры их, различны у разных групп
животных: как правило, это самая крупная клетка организма (Рис.1, Рис. 2).
По общим признакам строения
яйцеклетки не отличаются от соматических клеток. Они имеют ядро, цитоплазму и
оболочку Цитоплазма яйцеклетки называется ооплазмой. Ооплазма яйца содержит
разнообразные органеллы, которые встречаются и в соматических клетках. Однако
наряду с общими для всех клеток признаками, яйцеклетки имеют и ряд
особенностей.
Одной из них является наличие
особого поверхностного слоя ооплазмы, называемого кортикальным слоем или
кортексом, в котором содержатся гранулы или альвеолы и сократительные структуры
- микрофиламенты, состоящие из белка актина.

Рис. 1. Фолликул
морской свинки. В центре - яйцеклетка: 1-3 - клетки
фолликулярного эпителия; 4- оболочка; 5 - ядро; 6 - эндоплазматический
ретикулум (по Айзенштадт)
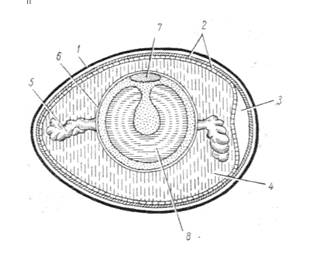
Рис. 2. Схема
строения куриного яйца (из Антипчук) 1- скорлупа; 2-
подскорлуповая пленка; 3 - воздушная камера; 4 - белковая оболочка (белок); 5 -
халазы; 6 - желточная оболочка; 7 - зародышевый диск; 8 - желток.
Кортикальные гранулы обнаружены
в яйцеклетках разных животных, включая яйца млекопитающих. Содержимое
кортикальных гранул в яйцах разных животных сходно по своему химическому
составу - оно богато мукополисахаридами, в них обнаруживается также некоторое
количество структурных белков и ферментов.
Все большее количество данных
свидетельствует о том, что кортикальный слой ответственен за организацию яйца.
Кроме того он играет важную роль при возникновении кортикальной реакции
яйцеклетки в момент оплодотворения и является основой того механизма, который
защищает яйцо от проникновения сверхчисленных спермиев.
Другой особенностью в строении
яйцеклеток является наличие в ооплазме значительных запасов питательных
веществ, обеспечивающих развитие и рост зародыша. Питательные вещества откладываются
в виде желточных включений. Накопление желтка в цитоплазме яйцеклеток
Происходит во время периода
роста ооцита. Яйцеклетки разных животных содержат разное количество желтка. Это
тесно связано с филогенетическим развитием вида животного. Различия в
количестве желтка в яйцеклетках разных животных приводят к большой
вариабельности их размеров. Имеется классификация яйцеклетки, основанная на
количестве желтка и распределении его в ооплазме. Знание классификации
яйцеклеток необходимо, так как с различным содержанием желтка в яйцеклетках и
характером его распределения в ооплазме связаны особенности развития зародыша в
начале эмбриогенеза.
В зависимости от количества
желтка различают: алецитальные, олиголецитальные, мезолецитальные и
полилецитальные яйца (Рис. 3).
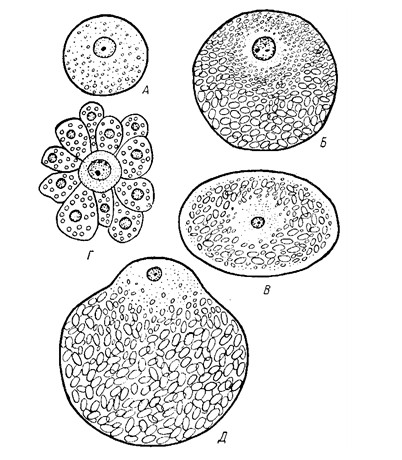
Рис. 3. Типы
яйцевых клеток А - изолецитальная клетка
ланцетника; Б - телолецитальная клетка лягушки; В - центролецитальная клетка
насекомого; Г - алецитальная яйцеклетка плоского червя, окруженная желточными
клетками; Д - телолецитальная яйцеклетка костистых рыб (по Кнорре).
Алецитальные яйца. Яйцеклетки
почти без желтка или с незначительным количеством его. Яйца такого строения
характерны для плацентарных млекопитающих и некоторых беспозвоночных животных.
Олиголецитальные
яйца (oligos -
малочисленный, lecythos
- желток) - с малым количеством желтка, характерны для многих беспозвоночных:
кишечнополостных, червей, иглокожих и моллюсков. Размеры таких яйцеклеток
небольшие (диаметр их около 0,1мм или ненамного больше).
Мезолецитальные
яйца (mezos - средний)
яйца. Яйцеклетки со средним содержанием желтка. Такие яйца встречаются у
амфибий и осетровых рыб. Диаметр яиц амфибий достигает 2-3мм, у лососевых рыб
до 7-9 мм.
Полилецитальные
яйца
(poly - много) - яйца с
большим количеством желтка. Они присущи многим рыбам, членистоногим (в том
числе всем насекомым), всем рептилиям, птицам и яйцекладущим млекопитающим. У
акулообразных рыб диаметр яйцеклетки достигает 60-70 мм. У курицы, не включая
белковую и скорлуповую оболочки - больше 30 мм, а у страуса больше 90 мм.
В зависимости от характера
распределения желтка в ооплазме различают: гомолецитальные, или
изолецитальные, телолецитальные и центролецитальные яйца.
В гомолецитальных яйцеклетках
желток распределяется равномерно. Обычно в эти яйца - с малым содержанием яйца
(олиголецитальные). В телолецитальных яйцах желток распределяется относительно
анимально-вегетативной оси неравномерно. В таких яйцеклетках желток
концентрируется в вегетативном (нижнем) полушарии яйца, а в области анимального
(верхнего) полушария его мало или почти нет. Ядро располагается в анимальном
полушарии яйцеклетки. Такое расположение желтка характерно для полилецитальных
яйцеклеток (рыб, рептилий и птиц) и всех мезолецитальных. Центролецитальные яйцеклетки
характеризуются также неравномерным расположением желтка в ооплазме яйца. В
этих яйцеклетках желток сосредоточивается в средней части ооплазмы. По обе
стороны желтка находятся две зоны ооплазмы свободные от желтка. Одна из этих
зон находится на периферии клетки, а другая - около ядра. Эти безжелтковые зоны
ооплазмы яйца связаны между собой ооплазматическими мостиками. Ядро в этих
яйцеклетках занимает центральное положение. Такое расположение желтка в
ооплазме характерно для полилецитальных яиц членистоногих.
Следующей особенностью в
строении яйцеклеток является их полярность. Полярность проявляется в
неравномерном распределении в яйцеклетке ооплазматических включений (желтка,
пигментов) и органоидов. Особенно четко выражена полярность в яйцеклетках
содержащих большое и среднее количество желтка и относящихся к телолецитальному
строению. Верхняя часть яйца бедная желтком или совсем его лишенная, называется
анимальной, а нижняя, где концентрируется желток, называется вегетативной.
Условная линия, соединяющая анимальный полюс яйца с вегетативным, получила
название, анимально-вегетативной оси яйцеклетки. Направление
анимально-вегетативной оси яйцеклетки определяет направление главной оси
зародыша: анимальный ее полюс будет соответствовать переднему, а вегетативный -
заднему отделу тела зародыша.
Полярность яйца также может
определяться местом образования полярных телец, местом нахождения микропилей и
т.д. Все большее количество данных свидетельствует о том, что полярная
организация яйцеклеток обусловлена кортикальным слоем.
У некоторых животных зрелые
яйцеклетки обнаруживают четкую билатеральную симметрию. Билатеральная
организация яйца обусловлена ассимметрично-эксцентричным расположением
некоторых включений и химических веществ в ооплазме. Обычно билатеральная
симметрия яйца вызывается появлением по одну сторону оси базофильного серпа,
богатого РНК, митохондриями и т.д. Эта часть яйца определяет будущую спинную
сторону зародыша. На противоположной стороне яйца в цитоплазме накапливается
жир. Эта часть яйца будет соответствовать вентральной стороне зародыша.
Характерной особенностью
строения яйцеклеток является наличие специальных защитных оболочек,
отсутствующих только у некоторых беспозвоночных, яйцеклетки которых могут
активно передвигаться. Различают несколько видов специальных оболочек:
первичные, вторичные и третичные оболочки.
Первичная (желточная)
оболочка яйцеклетки образуется самой яйцеклеткой. Желточная оболочка
представляет собой тонкую пленку, непосредственно примыкающую к плазматической
мембране ооцита. Примером могут служить желточные оболочки яйцеклеток
беспозвоночных животных (кроме яйцеклеток, которые способны амебоидно
передвигаться).
Вторичная оболочка
яйцеклетки образуется в результате жизнедеятельности фолликулярных клеток. К
ним относится блестящая оболочка (Zona
pellucida) яйцеклеток
млекопитающих (Рис. 3). Считают, что в образовании этой оболочки принимают
участие ооцит и фолликулярные клетки. Примером вторичных оболочек могут также
служить плотные оболочки насекомых.
Третичная оболочка
яйцеклетки образуется из веществ, секретируемых железами выводящих половых
путей. К ним относятся: белковая оболочка и твердая наружная капсула у акуловых
рыб, студенистая оболочка яйцеклеток амфибий, белковая, известковая и
волокнистая оболочки у крокодилов и черепах, а также белковая, подскорлуповая и
скорлуповая оболочки у птиц (Рис. 4).
Функции яйцевых оболочек весьма
разнообразны. В первую очередь они выполняют защитную функцию, предохраняя
яйцеклетку, а в дальнейшем и зародыш от влияния вредных факторов внешней среды.
Через них осуществляется связь зародыша с внешней средой. Дополнительные
оболочки яйцеклеток выполняют трофическую (питательную), поставляющие
органические и неорганические вещества, необходимые для развития нового
организма.
Жизнеспособность женских
половых клеток ограничена во времени. Сразу же на выходе из яичника яйцо
претерпевает определенные изменения, которые можно охарактеризовать как
старение. Для большинства млекопитающих, в том числе у человека, овулировавшее
яйцо должно быть оплодотворено в течение суток. В противном случае оно
становится нежизнеспособным. Яйца почти всех беспозвоночных также, как рыб и
амфибий должны быть оплодотворены не позднее, чем через несколько минут после
их откладки. Продолжительность жизни яиц других животных того же порядка, что и
у человека.
2. Мужские половые
клетки (сперматозоиды)
При изучении мужских половых
клеток следует разобраться в строении жгутиковых (бичевидных) сперматозоидов,
являющихся преобладающей формой сперматозоидов и для сравнения познакомиться с
морфологией безжгутиковых (небичевидных) сперматозоидов. Не обходимо обратить
внимание на такие физиологические особенности, как их подвижность,
продолжительность жизни, зависимости активности от окружающей среды и т.д.
Мужские половые клетки по
своему строению и физиологическим свойствам значительно отличаются от женских
половых клеток. Сперматозоиды значительно меньше яйцеклеток. Мужские половые
клетки крокодила имеют длину 20 мкм, воробья 200 мкм, морской свинки - 100 мкм,
быка - 65 мкм, а у человека в среднем 50 мкм. Сперматозоиды многочисленнее
яйцеклеток. Их количество у животных измеряется миллионами. Например, у
человека в 1 см3 спермы содержится 60 млн. сперматозоидов. Зрелые
сперматозоиды - это активно подвижные клетки.
Среди мужских половых клеток
различных групп животных выделяют два существенно различающихся между собой
типа сперматозоидов: жгутиковые (бичевидные) и безжгутиковые (небичнвидные).
Жгутиковые сперматозоиды являются преобладающей формой (Рис.4).

Рис.4. Формы
сперматозоидов человека и животных. 1 - человека; 2 -
тритона; 3 - речного рака; 4 - морской свинки; 5 - свиньи; 6 - быка; 7 -
петуха; 8 - ветвистого рака; 9 - десятиного рака; 10 - лошадиной аскариды; 11 -
острицы (по Голиченкову).
Жгутиковые сперматозоиды даже у
животных очень отдаленных видов построены по одной схеме, что, вероятно,
объясняется сходством их функционального назначения во всем животном мире.
В жгутиковом сперматозоиде
различают четыре отдела: головку, шейку, среднюю часть, хвост (жгутик). Все
отделы сперматозоида покрыты снаружи общей плазматической мембраной.
Головка сперматозоида имеет у
разных классов животных различную форму: у тритона - головка имеет форму
вязального крючка, у воробьиных птиц - штопоровидно закрученная, у
млекопитающих - спереди овальная, с боку грушевидная. Большую часть головки
сперматозоида занимает ядро. В цитоплазме передней части головки расположен
акросомный аппарат, который играет важную роль в растворении оболочек яйца. В акросоме
сосредоточены спермолизины - вещества, относящиеся к протеолитическим ферментам
(Рис.5).
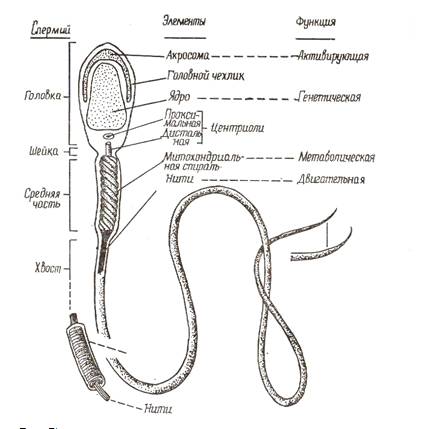
Рис.5. Схема
строения спермия млекопитающего, на которой изображены структуры, выявляемые с
помощью электронного микроскопа и указаны выполняемые ими функции (по
Александровской)
Шейка представляет собой
короткую, более узкую часть сперматозоида. В шейке расположены две центриоли:
проксимальная (передняя), примыкающая к ядру и дистальная (задняя), которая
соединяется с осевой нитью хвоста.
Средняя часть сперматозоида
состоит из осевой нити и окружающей ее цитоплазмы. В цитоплазме находится
большое количество митохондрий, которые расположены друг за другом в виде
спирально закрученной нити. Митохондрии генерируют энергию, необходимую для
движения мужской половой клетки.
Хвостовой отдел (жгутик)
сперматозоида состоит из осевой нити, которая покрыта тонким слоем цитоплазмы.
Осевая нить жгутика представлена 2 центральными и 9 периферическими пара
фибрилл, которые тянутся без существенных изменений вдоль всей длины хвоста
сперматозоида - от шейки почти до кончика (Рис.6).

Рис.6.
Электронно-микроскопическое строение спермия: 1
- головка; 2 - шейка; 3 - осевая нить; 4 - митохондрии; 5 - плазмолемма. ( по
Александровской).
Таким образом, общая
организация сперматозоидов и его отделов приспособлены к выполнению
специфических функций, присущих этой клетке. Такими функциями являются: а)
обеспечение встречи с женской половой клеткой; б) побуждение яйцеклетки к
развитию; в) передача в нее отцовского наследственного материала.
У некоторых животных
сперматозоиды не имеют жгутиков и называются безжгутиковыми (небичевидными).
Эти сперматозоиды имеют самую разнообразную форму: округлую, нитевидную,
двояковогнутую, иногда очень необычного типа (Рис.6).
Жизнеспособность и подвижность
сперматозоидов в воде ограничено во времени. В морской воде они утрачивают
подвижность через несколько часов, а в пресной, как правило, уже через
несколько минут. У животных с внутренним оплодотворением мужские половые клетки
сохраняют свою жизнеспособность несколько дольше: у свиньи - 22 - 30 ч., у овцы
- 30 - 36 ч., у рогатого скота - 25 - 30 ч., в половых путях женщины
продолжительность жизни спермиев колеблется от 2 до 4 суток.
У некоторых видов животных
сперматозоиды сохраняют жизнеспособность в половых путях самки длительное
время. Например, у некоторых летучих мышей осеменение происходит осенью, но на
протяжении всей зимней спячки животных сперматозоиды пребывают в состоянии покоя.
Только на следующую весну происходит оплодотворение. У кур сперматозоиды
сохраняются в течение 3-х недель. У многих насекомых сперматозоиды сохраняются
длительное время. Например, у пчел мужские половые клетки сохраняются в течении
нескольких лет.
По вопросу подвижности
сперматозоидов и их оплодотворяющей способности существовало и еще существует
много неверных представлений. Считали, что оплодотворяющая способность
сперматозоида сохраняется до момента потери им способности к передвижению. К
настоящему времени известно, что подвижность сохранятся гораздо дольше, чем
способность к оплодотворению. Так, сперматозоиды кролика теряют способность к
оплодотворению после 30 часового пребывания в половых путях самки, тогда как
подвижность они могут сохранять в течение более чем двух суток. Сперматозоиды
человека сохраняют способность к оплодотворению в течение 1 - 2 дней, а
подвижность они сохраняют до 4 дней.
3. Оплодотворение и
партеногенез
При изучении оплодотворения
необходимо сосредоточить внимание
на общей характеристике и
биологическом значении этого процесса, познакомиться с процессами осеменения,
которые предшествую процессу оплодотворения.
Хорошо разобраться в механизмах
взаимодействия половых клеток. Особое внимание уделить дистантному. Контактному
взаимодействию между половыми клетками и событиями, которые происходят в яйце
после проникновения сперматозоида. Показать роль в этих процессах гамонов,
акросомной и кортикальной реакции. На примере иглокожих уметь описать
последовательные стадии акросомной и кортикальной реакции. Обратить внимание на
механизмы защиты яйца от проникновения сверхчисленных спермиев у мономпермных
животных. Знать способы проникновения сперматозоида в яйцо у разных групп
животных.
При изучении партеногенеза,
гиногенеза и анрогенеза сосредоточить внимание на распространении этих способов
в животном мире, на факторах, побуждающих яйцеклетку к партеногенетическому
развитию, а также на механизмах, приводящих к диплоидизации яйцеклетки.
Познакомиться с опытом по искусственному партеногенезу и их теоретическим и
прикладным значением.
3.1 Общая
характеристика процесса оплодотворения и его биологическое значение
Каково биологическое значение
процесса оплодотворения?
Во-первых, при оплодотворении
происходит восстановление диплоидного набора хромосом, характерного для данного
вида животных.
Во - вторых, при оплодотворении
происходит активация яйцеклетки. Как это надо понимать? До оплодотворения
яйцеклетка в отношении обмена веществ находится в состоянии депрессии.
Метаболические процессы протекают на низком уровне. Кроме того, у большинства
животных яйцеклетка не завершает своего развития. Например, у млекопитающих
развитие яйцеклетки останавливается на стадии метафазы второго деления
созревания, у асцидий, некоторых моллюсков - на стадии метафазы первого деления
созревания, у аскариды - до первого созревания. Только у морского ежа
яйцеклетка успевает полностью завершить оогенез. Вот такие яйцеклетки,
содержащие в себе все, что необходимо для развития зародыша и находящиеся в
покоящемся состоянии ожидают толчка извне, для того, чтобы выйти из
анабитического состояния и стать на путь развития. Таким толчком и является
оплодотворение.
В - третьих, объединение
отцовских и материнских генов, приводит к созданию генетического многообразия
организмов, что служит материалом для естественного отбора и эволюции вида.
Оплодотворению предшествует
осеменение. Под осеменением понимают процесс выделения семенной жидкости во
внешнюю среду, либо в половые пути самки. В связи с этим различают осеменение
наружное и внутреннее.
У большинства животных,
обитающих в воде (многие беспозвоночные, большинство рыб, бесхвостые амфибии)
осеменение наружное. При наружном осеменении яйцеклетки и сперматозоиды
выбрасываются в воду, где и происходит оплодотворение.
У всех животных, размножающихся
на суше, а не в почве осеменение
Внутреннее. Такой способ
характерен и некоторым животным, обитающим в воде, например акуловым рыбам. При
внутреннем осеменении самец при помощи совокупительного органа вводит сперму
непосредственно в половые пути самки, где и происходит оплодотворение. Кроме
наружного и внутреннего осеменения встречается наружно- внутреннее осеменение.
Этот способ осеменения распространен среди членистоногих, обитающих в почве и
аналогичных субстратах, а также среди некоторых позвоночных (тритонов,
саламандр и др.).
У некоторых пиявок наблюдается
кожное осеменение, при котором сперматозоиды прикрепляются к гиподерме тела и
проникают в матку.
3.2 Механизмы
оплодотворения
Процесс оплодотворения у
животных можно разделить на три фазы. Первая фаза характеризуется сближением
сперматозоида с яйцеклеткой до их контакта. В эту фазу осуществляются
дистантные взаимодействия между половыми клетками. Вторая фаза начинается с
того, что сперматозоид прикрепляется к поверхности яйцеклетки. В это время
наблюдаются контактные взаимодействия между половыми клетками. Третья фаза
процесса оплодотворения начинается после проникновения сперматозоида в яйцо и
завершается объединением ядер мужской и женской половых клеток. Эта фаза
характеризует взаимодействие внутри яйца.
Дистантные
взаимодействия между половыми клетками
Дистантные взаимодействия
обеспечиваются рядом неспецифических факторов, среди которых особое место
принадлежит химическим веществам, которые вырабатываются половыми клетками.
Известно, что половые клетки выделяют гамоны или гормоны гамет. Гамоны, которые
вырабатываются яйцеклетками, называют гиногамонами, а спрематозоидами -
андрогамонами. Женские половые клетки выделяют две группы гамонов: гиногамоны I
и гиногамоны II,
оказывающие влияние на физиологию мужских половых клеток. Сперматозоиды
вырабатывают андрогамоны I
и II.
Некоторые из этих химических
веществ направлены на повышение вероятности встречи сперматозоида с
яйцеклеткой. Известно, что движение сперматозоида к яйцу осуществляется через
посредство хемотаксиса - движение сперматозоидов по градиенту концентрации
некоторых химических веществ, выделяемых яйцеклеткой. Хемотаксис достоверно
показан для многих групп животных, особенно беспозвоночных: моллюсков,
иглокожих и полухордовых. Хемотактические факторы выделены из яйцеклеток
морских ежей: у одних видов - это пептид, состоящий из десяти аминокислот, и
назван сперактом, у других видов - пептид состоит из четырнадцати аминокислот
и, получил название резакт. При добавлении экстрактов этих веществ в морскую
воду, сперматозоиды соответствующего вида начинают двигаться вверх по градиенту
их концентрации.
В движении сперматозоидов
млекопитающих по верхним отделам яйцевода существенное значение имеет явление
реотаксиса - способность двигаться против встречного течения жидкости яйцевода.
После того, как сперматозоид
пройдет сквозь защитные оболочки яйца и вступит в контакт с его плазматической
мембраной, начинаются контактные взаимодействия между половыми клетками,
которые приведут к проникновению сперматозоида в цитоплазму яйца.
Контактные
взаимодействия между половыми клетками
Контакт сперматозоида с
мембраной яйцеклетки приводит к активации половых клеток. Реакция активации
связана со сложными морфологическими, биохимическими и физико-химическими
изменениями в половых клетках. Активация мужской половой клетки, в первую
очередь связана с акросомной реакцией, а женской - с кортикальной реакцией.
Акросомная реакция характеризуется
быстрыми изменениями в акросомном аппарате головки сперматозоида,
сопровождающимися высвобождением заключенных в ней спермолизинов и
выбрасыванием акросомной нити в сторону поверхности яйца.
Рассмотрим общую схему
акросомной реакции у представителей разных групп морских беспозвоночных -
иглокожих, кольчатых червей, двустворчатых моллюсков, кишечно-дышащих и др.
На вершине головки
сперматозоида, плазматическая мембрана и, прилежащая к ней часть мембраны акросомного
пузырька, растворяются (лизируются). Свободные края обеих мембран сливаются
между собой в единую мембрану. Из обнажившейся акросомы выходят спермолизины в
окружающую среду и приводят к растворению яйцевых оболочек в месте контакта со
сперматозоидом. После этого внутренняя мембрана акросмного аппарата
выпячивается наружу и образует вырост в виде трубочки ( акросомная нить).
Акросомная нить удлиняется, проходит через разрыхленную область дополнительных
яйцевых оболочек и вступает в контакт, с плазматической мембраной яйцеклетки. В
области контакта акросомной нити с поверхностью яйца плазматические мембраны
сливаются и содержимое акросомной трубочки (нити) соединяется с цитоплазмой
яйцеклетки. В результате слияния мембран образуется цитоплазматический мостик.
Чуть позже по цитоплазматическому мостику в цитоплазму яйца перейдут ядро и
центриоль сперматозоида. Акросомная реакция завершается встраиванием мембраны
сперматозоида в мембрану яйцеклетки. С этого момента сперматозоид и яйцеклетка
являются уже единой клеткой (Рис.7, 8, 9.).
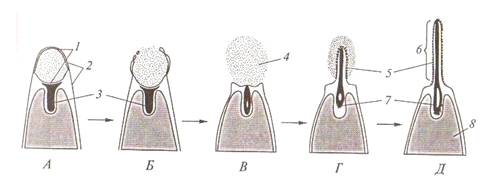
Рис.7. Акросомная
реакция сперматозоида: А - В - слияние наружной
мембраны акросомы и мембранысперматозоида. Излияние содержимого акросомного
пузырька; 1 - мембрана акросомы; 2 - мембрана сперматозоида; 3 - глобулярный
актин; 4 - ферменты акросомы; Г - Д - полимеризация актина и образование
акросомного выроста; 5 - биндин; 6 - вырост акросомы; 7 - актиновые
микрофиломенты; 8 - ядро сперматозоида. (по Голиченкову)
При общем сходстве акросомной
реакции, у этих животных между ними имеются и определенные различия. Так, у
иглокожих в отличие у червей и моллюсков в акросомном аппарате не содержатся
литические ферменты. У большинства изученных животных образуется одна
акросомная нить, а у некоторых червей - несколько таких нитей.
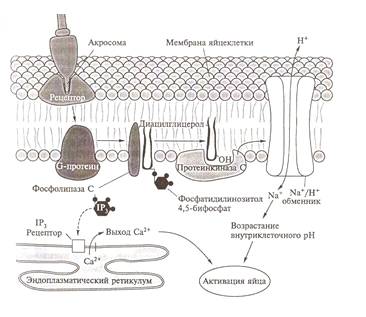
Рис.8.
Последовательность акросомной реакции у морского ежа. (по
Голиченкову)
При оплодотворении у
позвоночных животных также происходит акросомная реакция. У низших позвоночных
(миноги, и осетровые рыбы), она во многом сходна с акрсомной реакцией спермиев
беспозвоночных животных.

Рис.9. Схема
процессов, происходящих при взаимодействии мембран яйцеклетки и сперматозоида в
ходе оплодотворения (по Гилберт).
У акуловых рыб, рептилий и
птиц, яйца которых одеты плотными оболочками, соединение гамет происходит
раньше, чем эти оболочки сформируются. У этих животных акросома продолжает
выполнять свою первоначальную роль и, хорошо развита.
Акросомная реакция у
млекопитающих отличается от такой реакции у ьеспозвоночных и низших
позвоночных. В спермии млекопитающих акросомная реакция протекает без
образования акросомного выроста, Приблизившись к поверхности яйца, спермий
сливается с его плазматической мембраной боковой поверхностью головки.
У насекомых и высших рыб
соединение половых клеток происходит после того, как полностью образуются
плотные дополнительные яйцевые оболочки. В этих случаях сперматозоид проникает
в яйцо через микропиллярные каналы и соединение гамет происходит без участия
акросомы.
Активация яйца.
Кортикальная реакция. После того, как мужская половая
клетка прикрепится к поверхности яйца и ее акросомная нить вступит в контакт с
поверхностью ооплазмы, происходит активация яйцеклетки. Активация яйца связана
со сложными изменениями самых разных сторон его деятельности. Наиболее ярким
внешним проявлением активации являются изменения поверхностного слоя ооплазмы,
получившие название кортикальной реакции (Рис. 10).
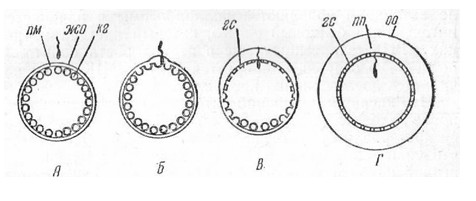
Рис.10.
Кортикальная реакция в яйце морского ежа А-приближение
спермия к яйцу; Б-Г-последовательные стадии кортикальной реакции; показаны
волна выделения содержимого кортикальных гранул, распространяющаяся от места
проникновения спермия, отделение оболочки и образование перивителлинового
пространства, формирование гиалтнового слоя; гс-гиалиновый слой; жо-желточная о
болочка кг-кортикальная гранула; оо-оболочка оплодотворения пм-плазматическая
мембрана; пп-перивителлиновое пространство, заполненное перивителлиновой
жидкостью (по Гинзбург).
Рассмотрим последовательные
стадии кортикальной реакции на примере наиболее полно, изученных яйцеклеток
морского ежа. Кортикальная реакция начинается с того, что мембрана,
ограничивающая каждую кортикальную гранулу, слипается с плазматической
мембраной яйца. В этом месте гранулы открываются, и их содержимое изливается в
желточную оболочку. Процесс секреции содержимого кортикальных гранул начинается
от места прикрепления сперматозоида и волнообразно распространяется во все
стороны до тех пор, пока не охватит всю поверхность яйца. Часть выделенного
содержимого кортикальных гранул оводняется и растворяется, образуя
перивителлиновую жидкость, которая оттесняет желточную оболочку от плазмолеммы
яйца, приводя к увеличению объема перивителлинового пространства. Другая часть
содержимого кортикальных гранул сливается с желточной оболочкой, которая при
этом утолщается и преобразуется в оболочку оплодотворения. Часть кортикальных
гранул, не участвующих в образовании оболочки оплодотворения, превращаются в
плотный слой, называемый гиалиновым слоем, расположенным над плазматической
мембраной. После того, как сформируется оболочка оплодотворения, другие
сперматозоиды лишаются возможности проникнуть в ооплазму яйца.
В последние годы был изучен
химический состав содержимого кортикальных гранул. Показано, что содержимое
кортикальных гранул содержит следующие вещества: а) протеолитический фермент
(актеллиновая деламиназа), разрывающий связи между клеточной оболочкой и
плазматической мембраной яйца; б) протеолетический фермент (сперм-рецепторная
гидролаза), который освобождает осевшую на желточной оболочке сперму; в)
гликопротеид, втягивающий воду в пространство между желточной оболочкой и
плазматической мембраной, вызывая их расслоение; г) фактор, способствующий
образованию оболочки оплодотворения; д) структурный белок гиалин, участвующий в
образовании гиалинового слоя.
Каково биологическое значение
кортикальной реакции?
Во-первых, кортикальная реакция
является тем механизмом, который защищает яйцо от проникновения сверхчисленных
сперматозоидов.
Во-вторых, образующаяся в
результате кортикальной реакции перивителлиновая жидкость, служит специфической
средой, в которой протекает развитие зародыша.
При активации яйца наблюдаются
и другие изменения самых разных сторон его деятельности.
Во-первых, снижается тормоз,
который блокировал мейоз и, ядерные преобразования продолжаются с той самой
стадии, на которой они остановились к моменту выхода яйца из яичника.
Во-вторых, наблюдается серия
биохимических изменений, сопровождаемых усилением углеводного обмена,
повышением синтеза липидов и белков.
В-третьих, резко возрастает
проницаемость клеточной мембраны для ионов натрия и калия.
События,
происходящие в яйце после проникновения сперматозоида
После того, как плазматическая
мембрана акросомной нити спермия сливается с плазматической мембраной яйца,
спермий утрачивает свою подвижность и его вовлечение внутрь яйца происходит
благодаря действию сил, исходящих из активированного яйца. Обычно сперматозоид
втягивается в ооплазму вместе с хвостом, но иногда хвостовой отдел
отбрасывается. Однако и в тех случаях, когда жгутик проникает в яйцо, он
отбрасывается и рассасывается.
Высоко-конденсированное ядро
сперматозоида начинает набухать, хроматин разрыхляется и ядро превращается в
своеобразную структуру, называемым мужским пронуклеусом.
Аналогичные изменения
происходят и в ядре яйцеклетки, в результате чего образуется женский
пронуклеус. В период формирования пронуклеусов, вдоль хромосом происходит
репликация ДНК. В дальнейшем пронуклеусы начинают перемещаться к центру
яйцеклетки. Ядерные оболочки, окружающие каждого из пронуклеусов разрушаются,
пронуклеусы сближаются и происходит кариогамия. Кариогамия - это последняя
стадия оплодотворения. При объединении пронуклеусов образуется ядро с
диплоидным набором хромосом. Затем хромосомы занимают экваториальное положение,
и наступает первое деление зиготы.
Ооплазматическая
сегрегация. После проникновения сперматозоида
начинаются интенсивные перемещения цитоплазмы яйцеклетки (ооплазмы). При этом
происходит расслоение, отмешивание различных составных частей ооплазмы, что
обозначается как ооплазматическая сегрегация. В ходе этого процесса намечаются
основные элементы пространственной организации зародыша. Поэтому данный этап
развития называют также проморфогенезом: имеется в виду, что в это время как бы
расставляются вехи для будущих морфогенетических процессов.
Моно- и полиспермия
Проникновение в яйцеклетку
одного сперматозоида, получило название, физиологической моноспермии.
Моноспермия присуща всем группам животных с наружным осеменением и многим
животным с внутренним осеменением (тем, которые подобно, млекопитающим имеют
яйцеклетки небольшого размера).
У других животных, например, у
некоторых членистоногих (насекомые), моллюсков (класс брюхоногих), хордовых
(акулообразные рыбы, хвостатые амфибии, рептилии и птицы) в яйцеклетку
проникает большое количество сперматозоидов. Такое явление получило название
физиологической полиспермии. Однако и в этом случае с ядром яйцеклетки
соединяется только ядро одного сперматозоида, тогда как остальные разрушаются
(рис.11).
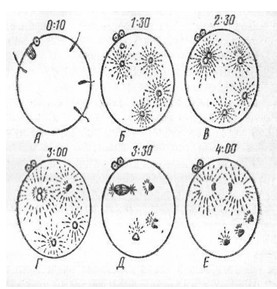
Рис. 11.
Полиспермия у тритона. А-проникновение спермиев в яйцо
на стадии метафазы II
деления созревания; Б-синхронные изменения семенных ядер, образование семенных
звезд; В-женское ядро соединяется с одним из семенных ядер; Г - Е-синкарион
вступает в митоз, сверхчисленные семенные ядра оттесняются в вегетативное
полушарие и дегенерируют. Цифры над изображением яиц - время после проникновения
спермиев при температуре 23о (по Гинзбург).
При физиологической моноспермии
имеются особые механизмы защиты яйца от полиспермии. Первый механизм связан с
изменением мембранного потенциала. Установлено, что в яйцеклетке лягушки, через
несколько секунд, после контакта со сперматозоидом заряд мембраны изменяется от
-28 до 8 мв и остается положительным в течение 20 мин. Такие же изменения
мембранного потенциала были обнаружены в яйцеклетках морского ежа. Оказалось,
что положительный заряд мембраны препятствует полиспермии. Другой широко
распространенный механизм защиты яйца от проникновения сверхчисленных
сперматозоидов связан с образованием оболочки оплодотворения и перивителлиновой
жидкости.
4. Партеногенез,
гиногенез, андрогенез. Естественный и искусственный партеногенез
Яйцеклетки некоторых животных
могут развиваться во взрослую особь и без оплодотворения. Такое развитие
называется естественным партеногенезом или девственным развитием Оно
встречается среди насекомых, низших ракообразных, коловраток, моллюсков и
других животных. У многих из них партеногенез чередуется с половым
размножением. Так, в течение лета коловратки откладывают яйца (летние яйца), из
которых развиваются без оплодотворения только женские особи. Последнее
поколение партеногенетических яиц дает и самцов и самок, откладывающих уже
оплодотворенные яйца.
Пчелы, осы, муравьи,
откладывают яйца, из которых часть оплодотворенные, а часть -
неоплодотворенные. Из оплодотворенных яиц развиваются самки, а из
неоплодотворенных - самцы. Такое своеобразное явление получило название
факультативного партеногенеза.
Некоторые животные, обычно
развивающиеся из неоплодотворенных яиц, начинают размножаться уже на личиночной
стадии. Это яаление называют педогенезом. Оно встречается у некоторых мух,
сосальщиков, ракообразных и других.
Партеногенетическое развитие
широко распространено не только среди беспозвоночных, но и у позвоночных
животных (рыбы, птицы, пресмыкающиеся). Так, у рыб партеногенетические яйца
дают нормальных особей, но только самок. Из неоплодотворенных яиц развиваются
нормальные особи мужского пола. Скальные ящерицы, содержащиеся в лабораторных
условиях отдельно от самцов, в течение двух лет откладывали яйца, из которых
развивались только самки. У других видов позвоночных естественный партеногенез
неизвестен.
Наблюдаемые явления
партеногенетического развития изучены с генетической и эмбриологической стороны
еще недостаточно. Известно, что у пчел и ос, развивающихся партеногенетически,
соматические клетки содержат гаплоидный набор хромосом (гаплоидный
партеногенез). В большинстве случаев после партеногенетической активации
яйцеклетки вней восстанавливается диплоидный набор хромосом. Существует два
основных способа диплоидизации: амейотический и мейотический. При аймеотическом
партеногенезе выпадает процесс редукции числа хромосом. При мейотическом
партеногенезе редукция числа хромосом происходит, но диплоидный набор
восстанавливается путем объединения хромосомных наборов обоих ядер,
получившихся в результате митотического деления гаплоидного ядра яйцеклетки.
Таким образом, в соматических клетках зародыша сохраняется диплоидный набор
хромосом.
С открытием естественного
партеногенеза были предприняты попытки искусственно вызвать развитие яиц.
Первые опыты по искусственному партеногенезу были проведены А.А. Тихомировым в
1886 году на тутовом шелкопряде. Исследователем было обнаружено, что
неоплодотворенные яйца тутового шелкопряда начинают развиваться после обработки
их концентрированной серной кислотой, при изменении температуры или после механического
повреждения, вызываемого потиранием яиц щеткой.
Обширные исследования были
проведены Ж. Лебом в 1913, 1916 гг. Он показал, что если яйца морского ежа
поместить в масляную, уксусную или в валериановую кислоту на несколько минут, а
затем последовательно перенести в подщелоченную воду и нормальную воду, то
наступала активация яйцеклетки.
Большой интерес вызвали опыты
Е. Батайона в 1910 г. Исследователь вызвал партеногенетическое развитие лягушек
и жаб уколом иглы.
Ж. Принкусу в 1956 г. удалось
добиться партеногенетического развития яиц млекопитающих (кроликов). Известны
многие попытки вызвать партеногенетическое развитие яйцеклеток других
млекопитающих. Однако достоверных данных о возможности партеногенетического
развития у млекопитающих на более поздних стадиях, чем бластоциста, пока нет.
Из изложенного выше видно. Что
партеногенетическое развитие яиц можно вызвать воздействием на них
разнообразными механическими, физическими и химическими агентами. Действия
данных факторов приводит к одним общим изменениям, подобным тем, которые
происходят в яйцеклетке при оплодотворении.
Опыты по искусственному
партеногенезу не только способствуют выяснению механизма этого процесса, но и
приобретают большое практическое значение. Б.Л. Астауров в 40-х годах разработал
промышленный способ стимулирования развития неоплодотворенных яиц тутового
шелкопряда при действии высоких температур. Этот способ позволяет получать в
большом количестве партеногенетических бабочек женского пола и тем самым
значительно увеличить производство коконов. Позднее З.А. Струнников разработал
метод низкотемпературной активации яиц. Партеногенетическое потомство,
полученное этим способом, состоит из одних самцов, которые вырабатывают больше
шелка, чем самки. Проблема партеногенеза ждет новых исследователей, так как
представляет большой теоретический и практический интерес.
Гиногенез и
андрогенез
Гиногенез - явление сходное с
партеногенезом и отличается от него тем, что при гиногенезе сперматозоиды
участвуют как активаторы развития яйцеклетки, не внося свой генетический
материал в яйцеклетку. Например, яйца серебряного карася могут быть
стимулированы спермой сазана, плотвы, обыкновенного карася. Развитие зародыша
осуществляется только за счет женского ядра. В популяциях гиногенетических
животных встречаются только особи женского пола. Гиногенез обнаружен у круглых
червей, серебряного карася и некоторых других животных. Гиногенетическое
развитие можно вызвать искусственно, воздействуя на сперматозоиды
рентгеновскими лучами, температурой и другими агентами.
Андрогенез - явление
противоположное гиногенезу. При андрогенезе развитие осуществляется за счет
ядер сперматозоидов и цитоплазмы яйцеклетки. Андрогенез наблюдается в тех
случаях, когда ядро яйцеклетки погибает, а в яйцеклетку проникает несколько спермиев.
Ядра двух сперматозоидов объединяются, и образуется ядро с диплоидным набором
хромосом. Естественный андрогенез обнаружен у тутового шелкопряда и
паразитической осы. При андрогенезе образуются только самцы. Андрогенез может
быть вызван и искусственно. Особую известность приобрели исследования Б.Л.
Астаурова (1957) по искусственному андрогенезу у тутового шелкопряда, имеющие
как теоретическое, так и практическое значение. Эти способом можно получить
популяцию самцов тутового шелкопряда, коконы которых содержат на 25-30% шелка
больше, чем коконы самок. Для получения андрогенетических особей, яйца тутового
шелкопряда облучались большой дозой рентгеновских лучей, при этих условиях ядро
разрушалось, а ооплазма не утрачивала жизнеспособности. После этого
осуществляли искусственное осеменение. Два из многих, проникших в клетку
сперматозоида сливались и давали начало ядру с диплоидным набором хромосом.
Такое ядро и обеспечивало нормальное развитие.
Пол зародыша обычно обусловлен
генотипически и зависит от сочетания половых хромосом в оплодотворенном яйце.
При помощи половых хромосом регулируется численное равенство в природе.
Для обозначения половых
хромосом у большинства млекопитающих, некоторых амфибий(леопардовая лягушка) и
насекомых (дрозофила) используют латинские буквы Х и У. Половые хромосомы самки
обозначают ХХ (знак ♀), а половые хромосомы самца ХУ (знак ♂). В
этом случае самцы продуцируют два типа сперматозоидов Х - или У - хромосомой. У
самок же образуется один тип яиц с Х-хромосомой. Особи, образующие один тип
половых клеток называются гомогаметными, а те особи, которые образуют два типа
половых клеток - гетерогаметными, т.е. самки были гомогаметны, а самце -
гетерогаметны.
Если яйцо будет оплодотворено
сперматозоидом с Х-хромосомой, из него разовьется самка, содержащая
ХХ-хромосомы. Если же яйцо будет оплодотворено сперматозоидом с У-хромосомой,
то из него образуется самец, содержащий ХУ-хромосомы. Таким образом, у
перечисленных выше животных У-хромосома содержит гены, которые определяют
мужской пол.
У некоторых животных (бабочки,
рыбы, птицы) гетерогеметен женский пол. В этом случае для обозначения половых
хромосом используют другие буква - Z
и W. Половые хромосомы
самки обозначают буквами ZW,
а половые хромосомы самца - ZZ.
У этих животных самки продуцируют два типа яйцеклеток с Z
или W-хромосомой., а
самцы - один тип сперматозоидов с Z
- хромосомой.
В этих механизмах нет ничего
принципиально различного, за исключением того, что гетерогаметны в одном случае
самцы, а в другом самки.
Полученные в последние годы
экспериментальные данные показали, что гены, ответственные за определение пола,
располагаются не только в половых хромосомах, но и в аутосомах. Тл обычно
является результатом взаимодействия нескольких пар генов. Пол является
полигенным признаком.
Следует также отметить, что
рассмотренный выше хромосомный механизм определения пола фиксирует лишь
первичные признаки пола - строение половых жлез, в то время как вторичные
половые признаки при определенных условиях могут претерпевать обратное
развитие, превращаясь в признаки противоположного пола. Решающую роль в этом
играют половые гормоны.
Литература
яйцеклетка
сперматозоид партеногенез гиногенез
Биология. Пособие для поступающих в вузы / А.Г.
Мустафин, Ф.К. Лагнуев, Н.Г. Быстренина и др., под ред. В.Н. Ярыгина. - М.:
Высшая школа, 2005. - 492 с.
Биология. Справочник студента / А.А. Каменский,
А.И. Ким, Л.Л. Великанов, О.Д. Лопина, С.А. Баландин, М.А. Валовая, Г.А.
Беляков. - М.: Физиологическое общество «СЛОВО», 2007. - 640 с.
Биология. Справочник школьника и студента / Под
ред. З. Брема, И. Мейнке. - М.: Дрофа, 2004. - 400 с.
Вахненко Д.В., Гарнизоненко Т.С., Колесников
С.И. Биология с основами экологии. Учебник для вузов / Д.В. Вахненко, Т.С.
Гарнизоненко, С.И. Колесников. - Ростов-на-Дону: Феникс, 2008. - 448 с.
Грин Н., Стаут У., Тейлор Д. Биология / Н. Грин,
У. Стаут, Д. Тейлор, под ред. Р. Сопера. - М.: Мир, 2004. - Т. 1. - 368 с.
Грин Н., Стаут У., Тейлор Д. Биология / Н. Грин,
У. Стаут, Д. Тейлор, под ред. Р. Сопера. - М.: Мир, 2004. - Т. 2. - 325 с.
Грин Н., Стаут У., Тейлор Д. Биология / Н. Грин,
У. Стаут, Д. Тейлор, под ред. Р. Сопера. - М.: Мир, 2004. - Т. 3. - 325 с.
Кемп П., Армс К. Введение в биологию / П. Кемп,
К. Армс. - М.: Мир, 2000. - 671 с.
Лысов П.К., Акифьев А.П., Добротина Н.А.
Биология с основами экологии: Учебник/ П.К.Лысов, А.П.Акифьев, Н.А.Добротина-
М.: Высшая школа., 2007.- 655 с.
Пехов А.П. Биология с основами экологии. Учебное
пособие для вузов с грифом МО / А.П. Пехов. - СПб.: Изд-во «Лань», 2008. - 672
с