Перетинчастокрилі півдня України
ДЕРЖАВНИЙ ВИЩИЙ НАВЧАЛЬНИЙ ЗАКЛАД
«ЗАПОРІЗЬКИЙ НАЦІОНАЛЬНИЙ УНІВЕРСИТЕТ»
МІНІСТЕРСТВА ОСВІТИ І НАУКИ УКРАЇНИ
Біологічний факультет
Кафедра загальної та прикладної екології і зоології
Курсова
робота
на
тему: Перетинчастокрилі півдня України
Виконав:
студент 4 курсу, групи 4121-2з
6.040102
Біологія
С.С.
Носко
Керівник
Горбань В.В.
Запоріжжя
- 2015 рік
ВСТУП
Комахи - найбільш численний клас
членистоногих безхребетних. За різними оцінками їх чисельність сягає 3 млн
видів. На сьогоднішній день описано близько 1 млн видів комах, що становить
приблизно 70% від загального числа відомої фауни. Представники класу комах
зустрічаються повсюдно і в більшості екологічних спільнот відіграють істотну
роль [1].
Відомо, що серед контактів людини з
навколишнім живим середовищем, одними з найтісніших, є контакти саме з
комахами. Цей зв’язок у деяких людей більший ніж з домашніми та господарськими
тваринами.
Перитинчастокрилі
(Hymenoptera) - один з найбільших рядів комах
<#"869593.files/image001.gif">
Рисунок 1.1 - Зовнішня будова
медоносної бджоли: 1 - голова; 2 - груди; 3 - черевце; 4 - антени (вусики); 5 -
прості очі; 6 - складне око; 7 - верхня щелепа (мандибула); 8 - хоботок; 9 -
язичок; 10 - нижня щелепа; 11 - переднє крило; 12 - заднє крило; 13 - дихальце;
14 - передня ніжка; 15 - середня ніжка; 16 - задня ніжка; 17-19 - грудні
сегменти; 20 - проподеум; 21-26 - черевні сегменти; 27 - жало
Хітиновий панцир виділяється гіподермою
- одношаровим епітелієм, лежачим на базальній мембрані, зазвичай він
складається з двох шарів: епікутикули і прокутикули. Основа останньої - білки,
пов'язані з азотовмісним полісахаридом хітином. Він дуже міцний і в природі
розкладається нечисленними видами грибів і бактерій, що мають фермент хітинази.
Нижній шар, в свою чергу, ділиться на ендокутикулу (еластична, надає покровам
гнучкість) і екзокутикулу (забезпечує міцність покривів, тому часто
склеротизованими). Верхній шар необхідний для запобігання випаровування вологи
і зсушення організму, він складається з чотирьох шарів і містить велику
кількість ліпідів [14].
Наявність хітинової капсули
перешкоджає постійному зростанню тіла, тому перитинчастокрилы линяють: старий
хітиновий панцир відстає від тіла, а епітелій виділяє новий і поки він не
придбав нормальної міцності, організм швидко росте.
Внутрішня будова
Порожнина тіла має змішану будову:
мезодермальні соміти (первинні спинні сегменти), в яких розвиваються вторинні
(целомічні) порожнини, є тільки у ембріонів, згодом вони розпадаються на окремі
клітини, в результаті вторинна порожнина тіла зливається із залишками
первинної, а всі простори між внутрішніми органами вільно повідомляються один з
одним.
М'язова система утворена пучками
поперечносмугастих волокон. Загальне число м'язів комах досягає 1,5-2 тис.,
Вони здатні скорочуватися до 1000 разів на секунду. Така висока швидкість
досягається завдяки тому, що на один нервовий імпульс м'яз відповідає кількома
скороченнями. Скелетні м'язи кріпляться до хітинових пластинок через особливі
нитки - тонофіламенти, що проходять через клітини епітеліального шару. У ході
ліньки заміщення відбувається таким чином, що м'язи не втрачають зв'язку з
хітиновими пластинками [15].
Травна система починається ротовою порожниною,
утвореної верхньою губою та іншими органами ротового апарату. У задній частині
ротової порожнини відкриваються протоки 1-3 пар слинних залоз (рис.1.2). У
гусениць метеликів слинні залози перетворені в прядильні. Ротова порожнина
з'єднана з трубчастої горлом. Далі травний канал продовжується передньої
кишкою, що складається із стравоходу і зоба. У деяких хижих комах задній кінець
передньої кишки перетворюється на жувальний шлунок, покриті хітином стінки
якого утворюють численні горби і зуби і служать для додатково перетирання їжі.
У середній кишці відбувається остаточне перетравлення і всмоктування їжі, на її
початку можуть впадати пилоричні придатки - випинання кишковика, які збільшують
всмоктувальну поверхню. Епітелій середньої кишки виділяє перітрофічну мембрану,
через яку переносяться ферменти та продукти травлення, крім того, вона захищає
епітеліальні клітини. Неперетравленні залишки їжі виводяться через задню кишку.
Задня кишка відіграє також роль органу видільної системи. На межі середньої і
задньої кишки відходять мальпігієві судини, де всмоктуються продукти обміну у
вигляді солей сечовини, які кристалізуються, всередині судин, а звільняється
при цьому вода всмоктується назад в гемолімфу. Найбільш інтенсивне всмоктування
води йде в задній кишці та її придатках - ректальних залозах. Кристали сечової
кислоти виводяться разом з неперетравленними харчовими рештками.
Травний канал сильно розвинений і
довгий у тих форм, які порівняно довго живуть в стадії дорослої комахи (бджоли,
мурашки, оси), і короткий у форм, що живуть у дорослому стадії недовго
(орехотворки, наїздники). Трахейна система часто забезпечена бульбашками. У
будові нервової системи чудово сильний розвиток так званих стебельчатих тіл або
мозкових звилин надглоткового вузла у тих перетинчастокрилих, які відрізняються
найбільш розвинутим життям (мурах, бджіл, ос); при цьому у різних неподільних
одного виду помічаються такі ж відмінності; так, у самців бджіл (трутнів) ці
органи розвинені менше, ніж у діяльних робітників. Властивий канібалізм.
Основна функція жирового тіла -
запасання поживних речовин, але деякі його клітини накопичують конкреції
сечової кислоти. Яка в даному випадку не виводиться з організму. Похідними
жирового тіла є органи світіння жуків-світлячків Lampyris, світло випускається
в результаті окислення люциферина ферментом люціферази при наявності кисню
[17].
Розташовані з боків серця
перикардіальні клітини (Нефроцитів) можуть поглинати з порожнини тіла сторонні
речовини, наприклад барвник кармін.
Кровоносна система перетинчастокрилих
незамкнута. Поточна по ній рідина називається гемолімфою. Над органами травної
системи знаходяться аорта і серце (рис.1.3). Серце розташоване в черевці і
являє собою трубковидний орган, задній кінець якого сліпо замкнутий. Воно
поділене перегородками на кілька камер (зазвичай 8), кожна забезпечена парою
остій, через які в серце засмоктується Гемолімфа. Передній кінець серця триває
в мускульну аорту. Близько мозку вона закінчується отвором, через яке Гемолімфа
знову потрапляє в порожнину тіла.
Кровообіг відбувається за рахунок
роботи м'язів серця, аорти і парних крилоподібних м'язів, з'єднаних з верхньої
діафрагмою - тонкою перегородкою, яка відокремлює серце від порожнини тіла. При
скороченні м'язів діафрагма опускається, гемолімфа спрямовується в білясерцеву
порожнину (перикардіальний синус) і надходить через остії в серці, а потім в
аорту і знову в порожнину тіла [18].
Основна функція кровоносної системи
- перенесення поживних, регуляторних речовин і продуктів метаболізму, а також
захисна функція, але значення гемолімфи в перенесенні кисню і вуглекислого газу
невелика. Це пов'язано з розвитком трахейної системи.
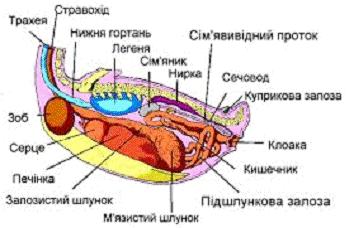
Рисунок 1.2 - Внутрішнє розташування
органів
Дихальна система представлена
сукупністю трахей. Трахея - посудина, внутрішні стінки якої вкриті хітиновим
вистиланням. Вона утворює спіральні потовщення, що перешкоджають спадання
стінок посудини і додають йому еластичність. Трахея багаторазово галузиться, і
кожна її гілочка закінчується кінцевою кліткою з радіальними відростками, які
розходяться. У них проходять кінцеві канальці трахеї, які доставляють кисень до
окремих клітин організму.
На середніх і задніх грудках і на 8
сегментах черевця розташовані дихальця (стигми) - отвори, забезпечені
замикальним апаратом. Через стигми повітря надходить в поперечні канальці, які
з'єднуються в поздовжні трахейні стовбури, а від стовбурів кисень доставляється
до клітин більш тонким трахеям. Замикальні апарати необхідні для зниження втрат
води в процесі дихання [19].
Голова.
Голова, груди і черевце різко
відособлені (крім сидячебрюхих, у яких черевце «щільно сидить» на грудях).
Голова вільна, в основному розширена в поперечному напрямку, гіпогнатна (ротові
органи направлені вниз) або прогнатна (... вперед). Складні очі майже завжди
добре розвинені, між ними звичайно три розташованих у вигляді трикутника
простих очка; зустрічаються також форми, позбавлені очок або зовсім сліпі
(наприклад, робочі мурашки деяких видів). На голові виділяють лоб (frons),
лиштва (кліпеус), тім'я (vertex), потилицю (occiput), щоки (genae), віски
(tempora).
Біля нижньої межі лоба і над
підставами нижніх щелеп розташовуються структури, звані тенторіальними ямками.
Це вп’ячування покривів, які проникають всередину головної капсули і формують
її внутрішній скелет - Тенторі (tentorium). Внутрішній скелет, злиття головних
сегментів і їх хітинових пластинок забезпечують особливу міцність головної
капсули.
Складні фасеткові очі складаються з
окремих очок - омматідіїв, вічко покритий прозорою рогівкою, яка має вигляд
опуклого шестикутника (фасетки) (рис.1.3-1.4). Під рогівкою знаходиться так
званий кришталевий (кристалічний) конус, разом вони утворюють лінзу вічка.
Світло сприймають рецепторні (ретинальні) клітини. Їх чутливі частини -
рабдомери - розташовані в центральній частині вічка і утворюють світлочутливий
елемент - рабдом. По краях лежать пігментні клітини, ізолюючі одне очко від
іншого.
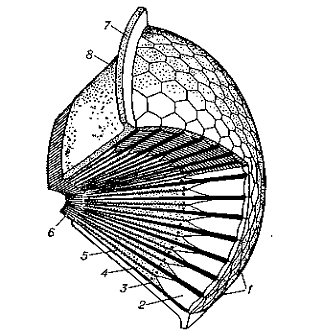
Рисунок 1.3 - Схема будови
аппозиційного фасеткового ока: 1 - рогівкові фасетки; 2 - світлоприломляючий
апарат; 3 - пігментні клітини; 4 - зорові клітини; 5- світлочутливий елемент
омматідія; 6 - аксони зорових клітин, що йдуть в оптичні ганглії; 7 - покриви
голови; 8 - очна капсула
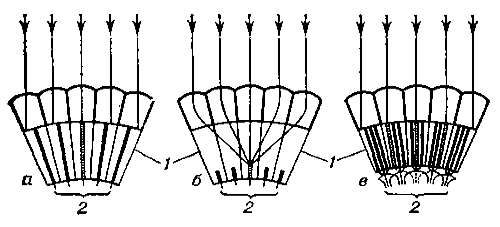
Рисунок 1.4 - Схема виникнення
сітчастого зображення в аппозиціонних (а), оптікосуперпозиціонних (б) і
нейросуперпозиціонних (в) фасеткових очах: 1 - окремі омматідіі з єдиним або
роз'єднаним світлочутливим елементом, складеним рабдомерами; 2 - аксони зорових
клітин. Заштриховані ті світлочутливі елементи, на які потрапляють паралельно
йдуть промені світла (показані стрілками)
Пігментні клітини у денних і нічних
перитинчастокрилих відрізняються за здатністю пігменту переміщатися всередині
клітини. У нічних комах пігмент може накопичуватися у верхній частині клітини,
при цьому промені світла потрапляють на рецепторні клітини сусіднього очка і це
істотно підвищує світлочутливість очей.
Складні очі можуть включати від
декількох сотень до 28 тисяч (у бабки) очок, кожен здатний сприймати вузький
пучок світлових променів. В результаті зображення об'єкта складається з окремих
«точок» (носить мозаїчний характер). Таке зображення виходить розпливчастим і
погано характеризує форму об'єкта. Але перевагою фасеткового ока є дуже широке
поле зору, що дозволяє вловити будь-який рух і вчасно помітити здобич чи ворога
[20].
Вусики (антени, або сяжки),
розвинені в дуже різною мірою і мають від 3 до 60 члеників; вони бувають прямі
або колінчаті, причому розрізняють стеблинку (скапус) і прикріпленний до нього
під кутом джгутик (flagellum). Між ними іноді виділяють поворотний членик, або
педіцеллюс.
Ротові органи, або ж нижня губа
(labium) і нижні щелепи (максілли) змінені в органи прийняття рідкої їжі
(органи лизання, хлебтання), утворюючи єдиний лабіо-максилярний комплекс.
Сильно рзвинуті верхні щелепи (мандібули, або жвали) у деяких слугують для
подрібнення їжі, а у більшості - знаряддями при будівництві житла, прогризанні
ходів у дереві (прогризали і свинцеві листи), приготування їжі для личинок,
вбивання здобичі, тягання будівельних і поживних матеріалів і т. п. На нижніх
щелепах і нижній губі розташовані нижньощелепні (максилярні) щупики
(6-членників) і нижньогубні (лабіальні) щупики (4-членникові). Лаку ротовий
апарат бджіл та джмелів характеризується подовженням обох пар нижніх щелеп і
утворенням ними хоботка, який служить для всмоктування нектару. Хоботок може
складатися в два коліна і притискатися до голови верхніми щелепами або навпаки,
сильно висуватися вперед і проникати в квітки з глибокими віночками. Верхня
губа і верхні щелепи практично не змінюються, щелепи використовуються для збору
та розмелювання твердого квіткового пилку, побудови гнізд [21].
Мозок робочих бджіл Apis містить
близько 850 тисяч нейронів і займає 0,35% - 1,02% від загального об'єму тіла. У
мурах роду Formica мозок займає 0,57% об'єму тіла. У одного з найменших
наїзників Megaphragma mymaripenne (його довжина 200 мкм) в мозку тільки 4600
нейронів, з яких лише 5% містять хромосом.
Центральна нервова система перетинчастокрилих
складається з надглоткового і підглоткового нервових вузлів (гангліїв), а також
гангліїв черевного нервового ланцюжка. Надглотковий нервовий вузол
функціонально відповідає головному мозку. Він ділиться на три частини. Попереду
розташовується протоцеребрум, який постачає нервами очі і має добре помітні
оптичні частки. З протоцеребрумом пов'язані грибоподібні тіла, які є вищими
асоціативними центрами. Вони найбільш розвинені у суспільних комах зі складною
поведінкою, таких як мурашки, бджоли, терміти, причому у робочих мурашок
грибоподібні тіла розвинені сильніше, ніж у самців і цариць того ж виду. За
передньою частиною головного мозку знаходиться дейтоцеребрум, инервуючий
вусики. Крихітний трітоцеребрум відповідає за головну капсулу і верхню губу
[22].
Підглотковий нервовий вузол регулює
роботу верхніх і нижніх щелеп, він пов'язаний з головним мозком (надгорлянковим
ганглієм). Черевний нервовий ланцюжок починається трьома великими грудними
нервовими вузлами; далі розташовуються черевні нервові вузли, число яких може
доходити до 11, але частіше вони об'єднуються. У більшості перетинчастокрилих є
8 черевних гангліїв, і останній з них несе сліди злиття. Нерідко зливаються
навіть грудні нервові вузли. у деяких комах цей процес призводить до утворення
єдиної ганглієвої маси в грудях, а в черевці залишаються і відходять тільки від
об'єднаної структури нерви.
Майже всі відділи центральної
нервової системи містять нейросекреторні клітини. Синтезовані ними гормони
надходять по аксонах в розташовані над кишковиком залози внутрішньої секреції -
кардіальні і прилеглі тіла, а далі - в гемолімфу. Нейросекрет регулює
діяльність всіх залоз внутрішньої секреції [23].
Органи чуття перетинчастокрилих.
Основою більшості органів чуття є
так звані сенсілли. Сенсілли складаються з однієї або декількох чутливих
клітин, від кожної відходить центральний відросток до нервової системи і
периферічний, який триває в видозміненний джгутик, він відрізняється
відсутністю в системі мікротрубочок двох центральних одиниць. Зверху сенсілли
покриті хітиновим шаром. Таку будову мають механорецепторні сенсілли. При зміні
положення волоска під впливом будь-якого фактора в чутливій клітині виникає
збудження, яке передається по центральному відростку в нервову клітину.
Трохи інакше влаштовані так звані
хордотональні сенсілли, їх периферичний відросток і джгутик оточені щільною
оболонкою, а зверху зафіксовані хітинзированні поверхнею шапкові клітини.
Найчастіше такі сенсілли прилягають до внутрішньої поверхні хітинових пластинок
в місцях з'єднання і служать для сприйняття низькочастотних вібрацій. Частина
хордотональних сенсил занурена в порожнину тіла на тонких мембранних трубках,
натягнутих між двома ділянками хітинових пластинок. Швидше за все, вони
фіксують зміни внутрішнього тиску і механічної напруги [24].
Нюхові сенсілли, розташовані на
вусиках і щелепних щупиках, відрізняються пористістю хітинового шару. Через
пори пахучі речовини потрапляють всередину сенсілли і контактують з
рецепторними клітинами.
Кожна смакова сенсілла містить
чутливі клітини, що реагують на різні смакові подразники, і одну
механорецепторну клітку. Хітиновий шар смакової сенсілли теж пронизаний порами.
Органами слуху служать тімпанальні
органи. Тут є дві поздовжні щілини, що ведуть у барабанні порожнини. Їх тонкі
внутрішні стінки відіграють роль барабанних перетинок і відокремлені один від
одного двома трахейними стволами. Останні, можливо, виконують функцію
резонаторів. До барабанних перетинок і трахей прилягають три групи
хордотональних сенсил. Комахи здатні сприймати звукові хвилі від 10 (інфразвук)
до 45 000 Гц (ультразвук) [25].
Груди.
Членики грудей щільно з'єднані між
собою; передня грудка (проторакс) дуже мала; середня грудка (мезоторакс)
розвинена найсильніше; задня грудка (метаторакс) мало розвинена, і з неї ззаду
щільно зростається невеликий перший членик черевця, утворюючи проподеум. Спинна
частина грудей утворена передньою спинкою (пронотума), середньою спинкою
(мезонотум) і задньою спинкою (метанотум). Найбільш складно влаштована середня
грудка (до неї прикріплюються передні крила), що складається з мезонотума (в
якій виділяють щит, або скутум, і щитики, або скутеллюм), мезопостнотума,
аксілл (два склерита з боків від щитики), мезоплевр (мезепімер і мезепістерни).
На мезонотумі можуть бути 2 бороздки, що сходяться ззаду (нотаулі), а на
скутумі - парапсидальні бороздки [26].
Черевце
Черевце вельми різної форми,
складається з 6-8 члеників, не рахуючи членика, щільно з'єднаного з задньою
грудкою, і видозмінених члеників, втягнутих у задній кінець черевця і несучих у
самок жало або свердло, у самців - органи злягання. До грудей черевце
прикріплюється або широкою основою, або більш-менш звуженим і подовженим
стебельком. Придатки заднього кінця черевця (жала і свердла або яйцеклад)
складаються з пари щетинок, або стилет, і жолобоватої частини, що складається з
двох окремих або більш-менш злитих пластинок. При підставі жала, яке в
спокійному стані завжди втягнуто всередину черевця, знаходиться парна отруйна
залоза з резервуаром, в якому накопичується отрута. Іноді (саме у багатьох
мурах) жала немає, є лише отруйна залоза; в цьому випадку комаха робить рану
щелепами і, підгинаючи вперед черевце, вприскує в неї отруту. Яйцеклад, або
свердле, можуть представляти велику різноманітність в будові. Призначення їх -
ввести яйце в рослину або тварину, в яких личинці належить жити; дивлячись по
тому, наскільки доступне місце, куди яйце відкладається, і яйцеклад може бути
коротше або довше, причому короткий яйцеклад в спокої втягнуті, довгі вільні
[27].
Статева система
На VIII-IX сегментах черевця
розташовуються статеві органи комах. Яйцеклад самок утворений 4-6 стулками -
виростами яйцекладних пластинок, які є видозмінною черевних ніг. Стулки, рухомі
відносно один одного, проштовхують яйце по яйцекладу, цей же механізм використовується
для заривання яєць в субстрат: самка проробляє хід, в який потім відкладає
яйця. Деякі комахи відкладають яйця в деревину (рогохвости, пилильщики). Їх
яйцекладні стулки забезпечені реберцями та зубцями. У наїзників, що відкладають
яйця в личинок інших комах, дуже довгий яйцеклад.
Яйцеклад вищих перетинчастокрилих
перетворюються на отруйне жало. У бджоли на стулках знаходяться дрібні щербини,
які допомагають фіксувати жало в шкірі жертви. Через ці зазубрини бджола не
може вийняти жало і гине.
Копулятивний (совокупний) орган
самців, на відміну від яйцеклада самок, які не гомологічні кінцівкам і
складається з підстави (фаллобази) і трубчастого едеагуса, який служить для
сім'явиверження [28].
Статеві залози самок - яєчники -
парні освіти. Яєчник складається з декількох яйцевих трубок, що мають загальний
вивідний канал - яйцевід. У сліпому кінці яйцевої труби - зачатковість клітини,
далі труба розширюється і формує ряд яйцевих камер. У них розташовані великі
ооцити (жіночі статеві клітини), оточені фолікулярними клітинами (необхідні для
харчування та накопичення жовтка, для захисту і регулюванню розвитку ооцита).
Разом вони утворюють фолікул - структурну і функціональну одиницю яєчника. У
міру дозрівання фолікули потрапляють в яйцепровід, а в дальньому кінці яйцевої
труби з зачаткових клітин утворюються нові. Яйцепроводи зливаються в піхву,
поєднанну з підрядними залозами, мускульної злягальною сумкою і сім'яприймача,
в якому сперматозоїди можуть жити протягом досить довгого часу [29].
Ноги
Ноги перетинчастокрилих ходильні
(зустрічаються копальні і хапальні модифікації у деяких ос), мають прості (у
Apocrita) або подвійні (у Symphyta, і Apocrita, крім Aculeata) вертлуги
(trochanter), 5-члениковиє лапки (у Chalcidoidea іноді 4- і 3-члениковиє ). На
вершинах гомілок знаходяться одна або дві рухливі шпори (формула шпор 2-2-2 у
Symphyta, але у багатьох Apocrita - 1-2-2, а у деяких мурашок 1-0-0). Шпора
передніх гомілок модифікована і з оточуючими густими волосками утворює
очищуючий апарат для вусиків. Частина груп мають особливості в будові (кошичок
з волосків на задніх ногах бджіл), пристосовані для збирання квіткового пилку.
Кінцівки кріпляться між черевним і
бічним хітиновими пластинками (склеритами) і складаються з широкого основного
членика - тазика (стегна; coxa), вертлуга, стегна, гомілки і лапки. Тазик
приєднується до тіла за допомогою двох зчленувань (виростків) і може рухатися
вперед і вгору або назад і вниз. Зчленування між вертлугом і стегном дозволяє
здійснювати рухи в різних напрямках, а колінне зчленування між стегном і
гомілкою потрібно, щоб наблизити кінцівку до тіла. Лапка складається з
декількох члеників (зазвичай з п'яти) і закінчується 1-2 кігтиками. З їх
допомогою комаха міцно закріплюється на субстраті. Іноді на вершині гомілки
розташовуються великі щетинки, звані шпорами. Вони беруть участь в русі
кінцівки і служать одним з визначальних ознак в деяких родинах [30].
Крила
Крила завжди перетинчасті, прозорі
або пофарбовані, з рідкісною мережею жилок, іноді зовсім без них; перша пара
крил завжди довше задньої; при польоті - передній край заднього крила за
допомогою особливих гачків причіпляється до заднього краю переднього, так що
обидва складають одну літальну поверхню (рис.1.5). У спокої крила складаються
горизонтально на спині, але у справжніх ос (сімействоVespidae) передні крила
складаються при цьому вдвічі по довжині і лежать з боків тіла. Як і в інших
загонах комах, серед перетинчастокрилих трапляються і безкрилі форми: такі всі
робочі мурашки, самки ос-німкень, самці деяких мурах (напр. Anergates), самки
деяких орехотворок, наїзників (Chalcidoidea, Ichneumonoidea) і ос (Bethylidae,
Dryinidae, Embolemidae, Tiphiidae та інші). Рідше зустрічається зворотний
приклад, коли крила є тільки у самки, - наприклад, у наїздників з сімейства
Agaonidae.
Жилки пронизують все крило комахи
(особливо багато їх у переднього краю крила), забезпечуючи необхідну жорсткість
конструкції і її аеродинамічні властивості. Перша, найбільш товста жилка
називається костальною. Нижче неї розташована субкостальна жилка. Далі йдуть
розгалужені радіальна і медійна жилки. Їх відгалуження нумеруються і
позначаються R1, R2, R3, ..., M1, M2, M3 ... Вони утворюють сектор радіуса і
медіальний сектор. У задній частині крила проходять кубітальні, анальні і
югінальні жилки. Крім того, у місця впадання субкості в косту може перебувати
щільна кутикулярна освіта - Крилове вічко, який знімає одну з шкідливих
вібрацій при польоті. Руху крил обмежують численні дрібні хитинові пластинки,
розташовані в області сочленованній мембрани між спинною і бічної платівкою
[31].
Рисунок 1.5 - Схема середньої спинки
і прикріплення крила: 1 - передня і задня фрагм; 2 - предщит; 3 - щит; 4 -
щитики; 5 - заспінка; 6 - парапсідальний шов; 7 - У-подібний шов; 8 - аксилярні
пластинки; 9 - югальна область
До деяких з них кріпляться м'язи,
службовці для зміни кута нахилу крил або їх складання вздовж тіла. М'язи, що
забезпечують рухи крил у польоті, зазвичай є м'язами непрямої дії: їх
скорочення викликає деформацію стінок так званого стерно-плейрального комплексу
(черевна і бічні хитинові пластинки зливаються, утворюючи капсулу у вигляді
чаші, яка додає сегменту додаткову міцність), розпластування або вигинання
спинної пластинки , і відповідно, рух крил вгору або вниз.
1.4
Представники видів перетинчастокрилих півдня України
Найбільш типові представники
сімейств пильщиків, вершників, дорожніх та громадських ос, а також ройових,
бджолиних та мурах:
Пильщик сосновий (Diprion pini). Має
опукле тіло завдовжки 7-10мм. Вусики 26-ти членникові. Характеризуються
вираженим статевим диморфізмом: самець менше, має в основному чорно-коричневе
або чорне забарвлення, вусики сильно гребінчасті. Тіло самки трохи крупніше,
більш потужне з пиловидними вусиками. Переважає світло-жовте забарвлення з декількома
темними плямами на грудях і на серединних сегментах черевця, яке на кінці має
пиловидний яйцеклад [32]. Ложногусениці світло-жовті або жовто-зелені, мають
три пари темних грудних ніг, вісім пар коротких помилкових ніг. Голова
порівняно невелика, коричнева. Довжина дорослої ложногусеници 26мм.
Алома-переможниця (Alomya
debellator) (рис.1.6). Середньої величини паразитичні комахи. Предня грудка без
виразних поздовжніх потовщень на боках. Середня спинка з двома чіткими
поздовжніми жолобками. Тіло струнке, чорного кольору. Голова кубічна волосиста.
Черевце стебельчате, дрібно і густо точкове. У самця вусики чорні, черевце
цілком або тільки 2-4 перших членика жовті. Передні стегна жовто-бурі з чорною
підставою. Середні стегна чорні з жовтою вершиною. У самки вусики
червоно-жовті, зазвичай з чорною вершиною. Черевце червоно-буре, його стеблинка
6-7-й членики чорні. Ноги жовто-червоні, задні тазики, вертлуги і стегна чорні.
Довжина тіла 12-14мм.

Рисунок 1.6 - Alomya debellator
Оса дорожня червонобрюха (Anoplius
viaticus) (рис.1.7). Осовидні перетинчастокрилі з дуже довгими ногами і
вусиками. Тримаються біля поверхні грунту, полюють на павуків, яких паралізують
для вигодовування своїх личинок. Вершини гомілок задніх ніг зстирчать в сторони
шипами, тіло чорне, лише підстава черевця знизу, а у самки і вершини стегон
червоні. Довжина тіла 7-9 мм.
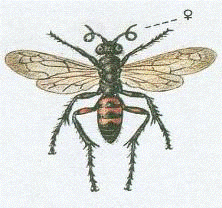
Рисунок 1.7 - Anoplius viaticus
Шершень (Vespa crabro) (рис.1.8).
Великі добре літаючі комахи, додають в спокої передні крила поздовжньо вдвічі.
Гомілки середніх ніг двома шпорами, кігтики прості. Живуть сім'ями, полюють на
комах. Черевце конічне, з прямо усіченою підставою. Голова за очима сильно
витягнута назад, задні оченята ближче один до одного, ніж до краю потилиці.
Забарвлення червоно-буре, груди чорні з жовтими плямами, черевце з чорними
смугами. Довжина матки від 25 до 35мм, робочі особини і самці коротші.

Рисунок 1.8 - Vespa crabro
Оса лісова (Dolichovespula saxonic)
(рис.1.9). Голова не витягнута, задні оченята віддалені від краю потилиці не
далі, ніж один від одного. Щоки довгі, довші 2-го членика вусиків, очі
відсунуті від жвал. Наличник з чорною точкою (самки і робочі) або суцільно
жовтий (самця). Довжина тіла 15-20мм.

Рисунок 1.9 - Dolichovespula saxonic
Піскорій піщаний (Ammophila
sabulosa) (рис.1.10). Крила з трьома кубітальними осередками. Черевце ясно
стебельчате. Стеблинка черевця тонка і складається лише з стергіта першого
черевного кільця. Радіальний осередок на вершині закруглений. Ноги довгі. Тіло
велике. Обидві поворотні жилки впадають у другу кубітальний осередок. Гомілки в
шипиках. Передні лапки у самки з гребенем. Стеблинка черевця довша іншої
частини черевця. Чорний, стеблинка черевця і два наступних черевних кільця
червоні. Третя кубітальна комірка стебельчата. Довжина тіла 15-20мм. Паралізує
гусениць нічних метеликів.

Рисунок 1.10 - Ammophila sabulosa
Бджола медоносна (Apis mellifera)
(рис.1.11). Передні крила з трьома кубітальними осередками. Задні гомілки без
шпор. Радіальний осередок майже досягає вершини крила. У жіночої особини голова
кругла, очі сходяться на тімені, вічка зрушені на лоб, черевце коротке, коротше
складених крил, тупо закруглене на кінці. У матки голова більш-менш трикутна,
очі на тімені розсунуті, між ними розташовані вічка, черевце більш-менш
витягнуте, загострене на кінці, збірний апарат на задніх ногах і черевці.

Рисунок 1.11 - Apis mellifera
Джміль садовий (Bombus hortorum)
(рисунок 1.12). Відрізняється від земляного джмеля тим, що у самок кінець
черевця в білих волосках. Голова попереду витягнута, щоки довгі, удвічі довше
своєї ширини на підставі. Довжина тіла 15-24мм.
У самця голова довга, щиток завжди в
світлих волосках.

Рисунок 1.12 - Bombus hortorum
Мураха чорний (Lasius niger)
(рис.1.13). Дрібні громадські комахи; робочі безкрилі, самки крилаті тільки на
час літа, самці крилаті, але живуть недовго. На голові виділяються лобові
валики, лобова борозенка і лицьовой щит. Черевце стебельчате, стеблинка
складається з одного членика. Самки і робітники не мають жала. Жвали з
зазубреним жувальним краєм, трикутні. Вусики прикріплені біля початку лобових
валиків поблизу лицьового щита. 2-5-й членики джгутика не довше інших члеників.
Забарвлення матове або слабо-блискуче, буре або чорно-буре. Довжина тіла 3-5мм
(робітники і самці), 7-8мм (самки). Живуть у землі або на пнях [33].

Рисунок 1.13 - Lasius niger
1.5
Характеристика району дослідження
Фізико-географічний район
досліджуваних нами комах - південь України. На всій цій території досить м’який
клімат, рання весна, пізня осінь, степовий ландшафт переважає, різноманітність
тваринного та рослинного (більше 4 тис. вищіх видів) життя, є усі переваги
розповсюдження та розвитку загону, особливо для суспільних комах. Доступність
широкого кола рослин для бджолиних, завдяки їх здатності до швидкого
переміщення в просторі шляхом активного польоту, багатий видовий склад, висока
чисельність популяцій, біологічні пристосування для збору пилку і нектару,
зробило їх основними учасниками процесу розмноження квіткових рослин. У той же час
для бджолиних квітки рослин є основним джерелом їжі [34].
Для півдня України характерна
трав'яна
<#"869593.files/image014.gif">
Рисунок 2.1 - Ентомологічний
сачок
Жалоносні перетинчастокрилі
можуть бути зібрані також у місцях їх гніздування, особливо в тих випадках,
коли вони утворюють колонії. При цьому вони можуть бути спіймані разом з
видобутком (її бажано наколоти при монтуванні на ту ж шпильку разом з осою)
[39] .
Місцем збору мурах є гніздо.
Гнізда бувають різних типів: підземні, надземні, в дуплах, під каменями.
Гніздо, якщо воно не у вигляді купи, зазвичай знаходять за одиничними мурашкам,
що несе видобуток в нору. Знаходять їх також по скупченості окремих мурашок, по
нагромадженню деревної муки, на пнях, нарешті, під камінням [40].
Мурах, залежно від розмірів,
потрібно ловити пінцетом, невеликим пензлем, змоченим в спирті, а також ексгаустером
(рис.2.2).
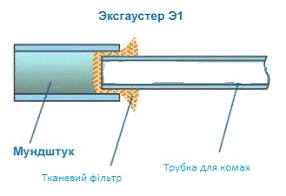
Рисунок 2.2 - Ексгаустер
Деякі види перетинчастокрилих
(наїзники, іноді оси) активно летять на світло, і для їх збору можуть бути
використані світлопастки.
Гнізда комах відшукують днем,
простежуючи за напрямком польоту окремих особин, і маркують невеликими світлими
щитами для кращого відшукання їх у вечірній час. За кількістю вилітають в
денний час з житла особин оцінюють загальну кількість комах в гнізді. Установку
пастки проводять між 22-24 год, коли дорослі особини повністю концентруються в
гнізді і температура навколишнього середовища невисока. Пастка (рис.2.3) являє
собою конструкцію, яка містить корпус у вигляді зігнутої конічної труби, на
одному кінці якої закріплено підстава, що має занурювані в грунт штирі і
поролоновий ущільнювач, а на іншому кінці - кріпильний кільце. Корпус
забезпечений вузлом нагнітання повітря, який за допомогою трубки з'єднаний з
порожниною корпусу [42]. До кріпильному кільцю за допомогою відповідного
кріпильного кільця прикріплений накопичувач комах, виконаний з
повітропроникного матеріалу, який натягнутий на кільцях, що мають стійки.
Пастка працює таким чином. У зібраному вигляді підставу закріплюють за
допомогою штирів над отвором льотка сім'ї комах. При природньому ранковому
вильоті з льотка комахи потрапляють у пастку і збираються роєм у верхній
частині накопичувача. При необхідності повного вилучення комах з гнізда вузлом
нагнітання повітря створюють спрямовану струмінь повітря в отвір льотка, що
підвищує інтенсивність вильоту комах з гніздовий камери в накопичувач [43].
Після збору комах накопичувач пастки перев'язують біля основи або швидко
закривають кришкою і від’єднують від корпусу. При необхідності до укріпленого
вже на льотці гніздовій камері корпусу можна приєднати новий порожній
накопичувач. Після закінчення збору накопичувач з комахами транспортується за
допомогою переносної ручки в лабораторію. Накопичувач добре вентилюється, що
дозволяє зберегти комах живими протягом декількох діб без застосування
спеціального пристрою. При необхідності наркотизації комах весь накопичувач або
частину його поміщають в ємність з наркотичним засобом. Пристрій простий у
виготовленні та експлуатації і володіє високою ефективністю вилову (від 500 до
1800 особин за один прийом залежно від розміру гнізда) [44].
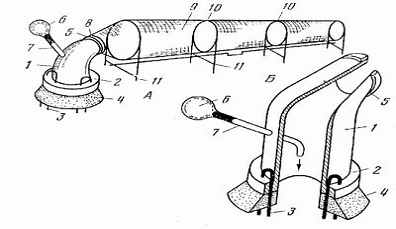
Рисунок 2.3 - Пристрій для
вилову комах: А - загальний вигляд, Б - корпус пастки (розріз); 1 - корпус, 2 -
підстава, 3 - штирі, 4 - ущільнювач, 5 кріпильне кільце корпусу, 6 - вузол
нагнітання повітря, 7 - трубка, 8 - кріпильне кільце накопичувача, 9 -
накопичувач, 10 - кільця, 11 - стійки.
Коли грунт прогріється, і стають
припустимими земляні роботи можна зайнятися виготовленням штучних гніздівель
для перетинчастокрилих (бджіл і ос). Найбільш простий спосіб залучення жищих у
грунті одиночних бджіл - спорудження нір на крутих схилах ярів, канав тощо,
позбавлених рослинності. Для цього шматком дроту товщиною до 1 см слід
продавлювати отвори під невеликим кутом до горизонтальної площини так, щоб
дощова вода не могла потрапляти в майбутні нори. Глибина нір може бути різною
(20-50 см) [45].
Дуже привабливі для
перетинчастокрилих гніздів'я, зроблені з очеретяних трубок (рис.2.4) довжиною
близько 15 см, зібраних в блоки по 10-20 штук, причому неодмінна умова
успішного заселення - наявність внутрішньої перемички (отвір не повинний бути
наскрізним) [46].

Рисунок 2.4 - Трубки з очерету
Для замаренування перетинчастокрилих
можна використовувати хлороформ і ефір. У польових умовах до монтування
перетинчастокрилих краще розкладати на ватяні шари і зберігати на них. Не
рекомендується використовувати для зберігання спирт, так як він може попсувати
матеріал і ускладнює монтування [47].
Найкраще монтувати свіжоспійманих
комах. При монтуванні з ватних шарів перетинчастокрилі, як правило, повинні
бути попередньо проведені через ексикатор. Великі перетинчастокрилі (бджоли,
оси, деякі наїзники) монтуються безпосередньо на шпильку. Прокол шпилькою слід
робити через середину передньої спинки [48].
Дрібні, головним чином паразитичні
перетинчастокрилі і мурашки наклеюються на пластинки з тонкого щільного
картону. Зручно один кінець пластинки загострити і наклеїти комаха на цей
кінець клеєм бажано на правий бік ногами в сторону шпильки. У такому положенні
ноги комахи легше зберегти неушкодженими (вони можуть мати важливі ознаки для
визначення).
ВИСНОВКИ
Біологічне різноманіття Hymenoptera,
багатство загону видами і їх велика кількість в найрізноманітніших біоценозах і
агроценозах роблять їх досить помітними на тлі господарської діяльності людини.
Рослиноїдні перетинчастокрилі - пилильщики, рогохвости, сім’яїд, які завдають
шкоди сільському і лісовому господарству. В першу чергу слід назвати такі види,
як звичайний і чорний хлібні пильщики, ріпаковий пильщик, вишневий пильщик.
Істотну шкоду в лісі наносять соснові пильщики, що об'їдають хвою. Технічними
шкідниками деревини є рогохвости.
Однак серед перетинчастокрилих
значно більше корисних видів, ніж шкідливих. Всі знають домашню бджолу, яка дає
нам не тільки мед, віск та інші продукти бджільництва, а й запилює культурні
рослини. Основними запилювачами рослин є джмелі і дикі поодинокі бджоли.
Обпилювачами можуть бути також оси і деякі наїзники, яких можна часто зустріти
харчуючимися на квітках. Але основну позитивну роль наїзники і оси грають в
якості ентомофагів.
Корисна діяльність ос як ентомофагів
вивчена ще недостатньо, краще за інших відомі тифії, що знищують личинок
хрущів. Безсумнівно, що і багато інших ос корисні в якості регуляторів
чисельності шкідливих комах. Найважливіша роль у цьому належить паразитичним
перетинчастокрилим - наїзникам [49].
Перетинчастокрилі відрізняються від
інших загонів комах цілим переліком різноманітностей: анатомією, розмноженням,
життєдіяльностю, розвитком і багато іншим. Наприклад у них повне перетворення
навідмінно від клопів (Hemiptera) та богомолів (Mantoptera), у яких неповне. У
паразитичних і жалючих перетинчастокрилих, на відміну від сидячебрюхих, перший
сегмент черевця при підставі звужений (черевце стебельчате) і з'єднаний з
грудьми рухливим зчленуванням. Таке зчленування забезпечує більшу рухливість
черевця, і багато з цих комах здатні підгинати кінець черевця до голови.
Доцільність цього стає зрозумілою, якщо врахувати, що відкладання яєць в тіло
живих комах вимагає великої рухливості кінця черевця, на якому поміщається
яйцеклад. У Hymenoptera не багато видів хижаків, богомоли ж усі. Клопи не мають
зобика та жилкування крил, їх нервова система значно простіша ніж у
перетинчастокрилих. У богомолів присутня складна статева поведінка, при якій
самець намагається привабити самку не як здобич, а як партнер. У жуків
(Coleoptera) сильно відрізняються крила, передня пара яких перетворилася у
тверді склеротизовані надкрила. Також у жуків сильно виражений статевий
диморфізм, а у Hymenoptera іноді зовсім немає [50].
Найбільші сімейства: Ichneumonidae
(більше 20 000 видів), Formicidae (більше 12 000 видів), Braconidae (більше 11
000 видів), Crabronidae (близько 7000 видів), Apidae (близько 5000 видів),
Tenthredinidae (близько 5000 видів) , Pompilidae (близько 4900 видів), Vespidae
(близько 4500 видів), Mutillidae (близько 4300 видів), Eulophidae (понад 4200
видів). Найбільші пологи включають по 1000 і більше видів: бджоли Lasioglossum
(1750 видів), Megachile (1560), Andrena (1500), мурашки Camponotus (1500) [51].
Використання сучасних методів
дослідження, що дозволяють отримувати незалежні дані для вирішення питань
таксономії і філогенії перетинчастокрилих. Одним з таких методів є хромосомний
аналіз, з успіхом застосовується для вивчення багатьох груп тварин, у тому
числі комах Зокрема, дослідженням хромосом комах та інших безхребетних успішно
займаються вітчизняні фахівці.
ПЕРЕЛІК ПОСИЛАНЬ
1.
Акимушкин И. И. Интересная биология / И. И. Акимушкин. - Смоленск : Русич,
1999.
.
Бей-Биенко Г.Я. Общая ентомология / Г. Я. Бей-Биенко - М.: Высшая Школа, 1966,
1971, 1980.
.
Горностаев Г.Н. Насекомые / Г. Н. Горностаев ; СССР. - М.: Мысль, 1970.
.
Догель В. А. Зоология беспозвоночных: Учебник для университетов / В. А. Догель
; под ред. Полянского Ю. И. ; 7-е изд., перераб. и доп. - М. : Высшая школа,
1981.
.
Зенкевич Л. А. Жизнь животных. Т. 3. Беспозвоночные / Л. А. Зенкевич. - М. :
Просвещение, 1969.
.
Захваткин Ю. А. Курс общей энтомологии / Ю. А. Захваткин. - М. : Агропромиздат,
1986.
.
Козлов А. Т. Структура и механизмы поведения беспозвоночных / А. Т Козлов., Н.
А. Тушмалова - В. : Квадрат, 1995.
.
Панов Е. Н. Общение в мире животных / Е. Н. Панов. - М. : Знание, 1970.
.
Плавильщиков М. М. Определитель насекомых / М. М. Плавильщиков. - М. :
Просвещение, 1957.
.
Негробов А. П. Определитель семейств насекомых / А. П. Негробов., Ю. И.
Черненко. - В. : Издательство ВГУ, 1990.
.
Красная Книга Украины (Животный мир) / под ред. М. М. Щербака. - Киев:
Украинская Энциклопедия, 1994.
.
Плавильщиков Н. Н. Определитель насекомых: Краткий определитель наиболее
распространенных насекомых европейской части России / Н. Н. Плавильщиков. - М.
: Топикал., 1994 - 544 с.
.
Зенкевича Л. А. Жизнь животных. В 7т. Т.3. Беспозвоночные. / под ред. Л. А.
Зенкевича - М. : Просвещение, 1969. - 375 с.
.
Фасулати К. К. Полевое изучение наземных беспозвоночных / К. К. Фасулати. - М.
: Высшая школа, 1961. - 304 с.
.
Мамаев М. М. Определитель насекомых европейской части СССР / М. М. Мамаев., Л.
Н. Медведев., Ф. Н. Правдин. - М. : Просвещение, 1976. - 303 с.
.
Мамаев М. М. Определитель насекомых европейской части СССР. Т.3.
Перепончатокрылые. Первая часть / М. М. Мамаев., Л. Н. Медведев., Ф. Н. Правдин
; под общ. ред. Г. С. Медведева. - Л. : Наука, 1978. - 584 с.
.
Богданов-Катька М. М. Руководство к практическим занятиям по общей ентомологи /
М. М. Богданов-Катька. - М. ; Л.: ОГИЗ-Сел.
.
Брайен М. Общественные насекомые. Экология и поведение / М. Брайен. М. : Мир,
1986. - 398 с. хозгиз, 1947. - 356 с.
.
Коппел Х. Биологическое подавление вредных насекомых / Х. Коппел., Дж. Мертинс.
- М. : Мир, 1980. - 427 с.
.
Мазохин-Поршняков Г. А. Зрение насекомых / Г. А. Мазохин-Поршняков. М. : Наука,
1965. - 263 с.
.
Зеликман А. П. Практикум по зоологии беспозвоночных / А. П. Зеликман. М. :
Высшая школа, 1969. - 335 с.
.
Жерихин В. В. Насекомые в экосистемах суши: Историческое развитие класса
насекомых / В. В. Жерихин. - М. : Наука, 1980. - 189 -223 с.
.
Яхонтов В. В. Экология насекомых / В. В. Яхонтов. - М. : Высшая школа, 1969. -
488 с.
.
Штейнхаус Е. Патология насекомых / Е. Штейнхаус. - М. : Изд-во иност.лит.,
1952. - 840 с.
.
Чернышев В. Б. Суточные ритмы активности насекомых / В. Б. Чернышев. - М. :
Изд-во МГУ, 1984. - 216 с.
.
Холодковский Н. А. Курс энтомологии теоретической и прикладной ; изд.4-е.,
Т.1-3 / Н. А. Холодковский. - М. ; Л., 1927-1931.
.
Тельман П.М. Методы лесоентомологических обследований / П. М. Тельман., А. А.
Катаев. - Л., 1964.
.
Тамарина Н. А. Основы технической ентмологии / Н. А. Тамарина. - М. : изд-во
МГУ, 1990. - 202 с.
.
Рафес П. Н. Роль и значение растительноядных насекомых в лесу / П. Н. Рафес. -
М. : Наука, 1968.
.
Прингл Дж. Полет насекомых / Дж. Прингл. - М. : Изд-во иностр. лит-ры, 1963. -
179 с.
.
Падий М. М. Краткий определитель вредителей леса / М. М. Падий. - Л. : Лесная
промышленность, 1979. - 238 с.
.
Падий М. М. Перепончатокрылые и двукрылые / М. М. Падий. - Л. : Лесная
промышленность, 1981. - 221 с.
.
Захваткин Ю. А. Эмбриология насекомых / Ю. А. Захваткин. - М. : Высшая школа,
1975. - 328 с.
.
Джекобсон М. Половые феромоны насекомых / М. Джекобсон. - М. : Мир, 1976. - 391
с.
.
Тищенко В. П. Физиология фотопериодизма насекомых / В. П. Тищенко. М. : Тр.
ПЭО, 1977. - Т.59 - 152 с.
.
Ушатинская Р. С. Скрытая жизнь и анабиоз / Р. С. Ушатинская. - М. : Наука,
1990. - 180 с.
.
Шванвич Б. Н. Курс общей энтомологии / Б. Н. Шванвич. - М. ; Л. : Сов. Наука,
1949. - 900 с.
.
Андриянова Н. С. Экология насекомых / Н. С. Андриянова. - М. : изд-во МГУ,
1970. - 158 с.
.
Викторов А. Г. Проблемы динамики численности насекомых на примере вредной
черепашки / А. Г. Викторов. - М. : 1967.
.
Гилмур Д. Метаболизм насекомых / Д. Гилмур. - М. : Мир, 1968. - 229 с
.
Еськов Е. К. Поведение медоносных пчел / Е.К. Еськов. - М. : Колос, 1981. - 184
с
.
Гринфельд Е. К. Происхождение и развитие Антофилия у насекомых / Е. К.
Гринфельд. - Л. : Изд-во ЛГУ, 1978.
.
Зенкевича Л. А. Жизнь животных : Беспозвоночные / Л. А. Зенкевича ; под ред. Л.
А. Зенкевича. - М. : Просвещение, 1969. - 563 с.
.
Мамаев Б. М. Гравитационная теория происхождения насекомых / Б. М. Мамаев. - М.
: Ентомол. Осмотрев., 1975. - Т.54, вып. 3. 499-506 с.
.
Приставко В. П. Принципы и методы экспериментальной энтомологии / В. П.
Приставко. - Минск, 1979. - 133 с.
.
Тышлер В. Сельскохозяйственная екология / В. Тышлер. - М.: Колос, 1971. - 433
с.
.
Сукачев В. Н. Основы лесной биогеоценологии / В. Н. Сукачев. - М. : 1964.
.
Бей-Биенко Г. Я. Практикум по сельскохозяйственной энтомологии / Г. Я.
Бей-Биенко., М. К. Асатур. - Л. : Колос, 1968. - 360 с.
.
Воронцов А. И. Лесная ентомологія / А. И. Воронцов. - М. : Высшая школа, 1975.
- 368 с.
.
Брайен М. Общественные насекомые. Экология и поведение / М. Брайен. М. : Мир,
1986. - 398 с.