Мхи Сахалина
Содержание
Введение
. Остров Сахалин
.1 К истории
.2 География
.3 Рельеф
.4 Климат
. Общая характеристика
мохообразнных
.1 Происхождение и филогения
.2 Как устроены мохообразные
. Практическая часть
Заключение
Список использованной литературы
Введение
В наше время в мире происходят непрерывные
изменения стратегии и методов, поэтому проблема исследования данной темы
остаётся актуальной.
Ввиду того, что данная темы достаточно
распространённая, следует учесть тот факт, что данную тему уже анализировали
другие авторы в своих работах: учебниках, монографиях, в периодических
изданиях, а также конечно и в интернете. Но, тем не менее, можно отметить, что
при изучении литературы наблюдается недостаточное количество информации по
данной теме.
Научная значимость работы состоит в улучшении и
упорядочивании уже существующей научно-методологической базы по исследуемой
теме - еще одним независимым авторским исследованием.
Цель: изучить новые местонахождения
краснокнижных мхов на Сахалине.
Задачи:
· изучить литературу (статьи,
монографии, диссертации и т.д) по данной теме
· организовать маршрутные походы
· собрать гербарий
· отметить новые точки местонахождения
· обработать материал для публикации
Глава 1. ОСТРОВ САХАЛИН
1.1 К
истории Сахалина
Древняя и средневековая история Сахалина и
Курильских островов полна тайн. Так, сегодня мы не знаем (и вряд ли
когда-нибудь узнаем), когда на наших островах появились первые люди. Позний
неолит. 3000 до н.э.Археологические открытия последних десятилетий позволяют
сказать только, что это произошло в эпоху палеолита. Загадкой остается и
этническая принадлежность населения островов вплоть до появления здесь первых
европейцев и японцев.
А они появились на островах только в XVII
столетии и застали на Курилах и южном Сахалине айнов, на северном Сахалине -
нивхов. Вероятно уже тогда в центральных и северных районах Сахалина жили ульта
(ороки).
Первой европейской экспедицией, оказавшейся у
курильских и сахалинских берегов, стала экспедиция голландского мореплавателя
М.Г.Фриза. Он не только исследовал и нанес на карту юго-восток Сахалина и Южные
Курилы, но и провозгласил Уруп владением Голландии, что, однако, осталось без
каких-либо последствий.
Огромную роль в изучении Сахалина и Курильских
островов сыграли и русские землепроходцы. Сначала - в 1646 г. - экспедиция
В.Д.Пояркова открывает северо-западное побережье Сахалина, а в 1697 г.
В.В.Атласов узнает о существовании Курильских островов. Айн Уже в 10-е гг.
XVIII в. начинается процесс изучения и постепенного присоединения Курильских
островов к Российскому государству. Успехи России в освоении Курил стали
возможны благодаря предприимчивости, мужеству и терпению Д.Я.Анциферова,
И.П.Козыревского, И.М.Евреинова, Ф.Ф.Лужина, М.П.Шпанберга, В.Вальтона,
Д.Я.Шабалина, Г.И.Шелихова и многих других российских
исследователей-землепроходцев.
Одновременно с русскими, которые двигались вдоль
Курил с севера, на Южные Курилы и крайний юг Сахалина начинают проникать
японцы. Уже во второй половине XVIII в. здесь появляются японские фактории и
рыбалки, а с 80-х гг. XVIII в. - начинают работать научные экспедиции. Особую
роль в японских исследованиях сыграли Могами Токунай и Мамия Риндзо.
В конце XVIII в. исследования у берегов Сахалина
вели французская экспедиция под командой Ж.-Ф.Лаперуза и английская - под командой
В.Р.Броутона. С их работой связано появление теории о полуостровном положении
Сахалина. Свой вклад в эту теорию внес и российский мореплаватель
И.Ф.Крузенштерн, который летом 1805 г. безуспешно пытался пройти между
Сахалином и материком.
Точку в споре поставил Г.И.Невельской, который в
1849 г. сумел найти судоходный пролив между островом и материком. За открытиями
Невельского последовало приосединение Сахалина к России. На острове один за
другим возникают русские военные посты и селения.
В 1869-1906 гг. Сахалин был крупнейшей каторгой
в России.
С начала XIX в. Сахалин и Курилы становятся
объектом российско-японского территориального спора. В 1806-1807 гг. на Южном
Сахалине и Итурупе русскими моряками были разгромлены японские поселения.
Ответом на это стал захват японцами на Кунашире русского мореплавателя
В.М.Головнина. На протяжении двух последних столетий русско-японская граница
неоднократно менялась.
Крейсер "Новик"В 1855 г. в
соответствии с Симодским договором граница прошла между островами Уруп и Итуруп,
Сахалин же был оставлен неразделенным. В 1875 г. Россия передала Японии
принадлежавшие ей Северные Курилы, получив взамен все права на Сахалин.
Пограничный камень, установленный на 50-ой
параллели, разделявшей Сахалин на северную и южную части после Портсмутского
мирного договора (август, 1905)В результате поражения России в русско-японской
войне 1904-1905 гг. Японии удалось отторгнуть у нее Южный Сахалин. В 1920-1925
гг. под японской аккупацией находился Северный Сахалин. В последний раз
российско-японская граница претерпела изменения в 1945 г., когда наша страна в
результате победы во второй мировой войне вернула себе Южный Сахалин и
Курильские острова.
1.2 География
Сахалинская область - единственная в стране,
расположенная на 59 островах, включает в себя остров Сахалин с находящимися
вблизи него островами Монерон и Тюлений и две гряды Курильских островов.
Площадь Сахалинской области составляет 87,1 тыс.
км2, из них около 78 тыс. км2 занимает Сахалин, протянувшийся в меридиональном
направлении на 948 км при средней ширине около 100 км. Отделенный от материка
Татарским проливом и проливом Невельского, а с юга от Хоккайдо (Япония)
проливом Лаперуза, омываемый водами теплого Японского и холодного Охотского
морей, из космоса он похож очертаниями на рыбу, «чешуей» которой служат
многочисленные реки и озера.
Курильский архипелаг, включающий более 30
значительных островов (наиболее крупные - Итуруп, Парамушир, Кунашир, Уруп) и
множество мелких островков и скал, протянувшись на 1200 км от о. Хоккайдо
(Япония) до полуострова Камчатка, отделяет Охотское море от Тихого океана.
Глубокие проливы Буссоль и Крузенштерна разделяют их на три группы, образующие
Большую Курильскую гряду: северную (о-ва Шумшу, Парамушир, Атласова,
Анциферова, Маканруши, Онекотан, Экарма, Харимкотан, Шиашкотан, Чиринкотан и
др.), среднюю (о-ва Матуа, Расшуа, Кетой, Симушир и другие) и южную (о-ва
Черные Братья, Броутона, Уруп, Итуруп, Кунашир и др.). В 60 км от последней
расположена отделенная Южно-Курильским проливом Малая Курильская гряда. Она
пролегла на 105 км параллельно Большой Курильской гряде и включает в себя б
островов, наибольший из которых - Шикотан.
Государственная граница между Российской
Федерацией, представленная в этом регионе Сахалинской областью, и Японией
проходит по проливам Лаперуза, Кунаширскому, Измены и Советскому.
Берега Сахалина слабо изрезаны, крупные заливы
имеются только в южной и средней частях острова. Для восточной части характерны
выровненная береговая линия и многочисленные лощины, образующиеся в устьях впадающих
в море рек.
Поверхность Сахалина весьма гориста. Большая
часть его территории - это средневысотные горы, особенностью которых является
их меридиональная ориентировка. Одним из основных горных поднятий острова,
располагающегося в его восточной части, являются Восточно-Сахалинские горы,
протянувшиеся от низовьев р. Тымь до п-ова Терпения, с наивысшей вершиной
Сахалина - горой Лопатина (1б09 м). Они состоят в основном из двух горных
хребтов - Набильского и Центрального. Западная часть острова занята менее
высокими Западно-Сахалинскими горами (наивысшая точка - г. Возвращения, 1325
м), которые протягиваются от мыса Крильон до р. Хунмакта (Камышовый и
Южно-Камышовый хребты и их отроги).
На юге Сахалина расположены Сусунайский и
Тонино-Анивский хребты.
Горные сооружения острова разделяются
низменностями (Тымь-Поронайская, Сусунайская, Муравьевская и др.), часто
заболоченными и прорезанными многочисленными реками.
Северная часть Сахалина занята
Северо-Сахалинской центральной равниной и прибрежными низменностями на западе и
востоке, отделенными от равнины двумя относительно невысокими (до 600 м)
хребтами, состоящими из останцовых гор (Вагис, Даахуриа, Оссой и др.).
Полуостров Шмидта характеризуется двумя
низкогорными (до б23 м) хребтами, разделенными холмистой Пиль-Диановской
низменностью; вдоль западного побережья развита полоса низких морских террас с
дюнами, пересыпями и болотами.
Курильские острова, являющиеся областью активной
вулканической деятельности, в орографическом отношении представляют собой два параллельных
подводных хребта, которые выше уровня океана выражены цепью островов Большой и
Малой Курильских гряд.
1.3 Рельеф
Рельеф острова составлен
средневысотными горами, низкогорьями и низменными равнинами. Южная и
центральная части острова характеризуются горным рельефом и состоят из двух
меридионально ориентированных горных систем - Западно-Сахалинских (до 1327 м
высотой - г. Онор) и Восточно-Сахалинских гор
<https://ru.wikipedia.org/wiki/%D0%92%D0%BE%D1%81%D1%82%D0%BE%D1%87%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%B8%D0%B5_%D0%B3%D0%BE%D1%80%D1%8B>
(до 1609 м высотой - г. Лопатина), разделённых продольной Тымь-Поронайской
низменностью. Север острова (за исключением полуострова Шмидта) представляет
собой пологую холмистую равнину.
Берега острова слабо изрезаны;
крупные заливы - Анива
<https://ru.wikipedia.org/wiki/%D0%90%D0%BD%D0%B8%D0%B2%D0%B0_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>
и Терпения
<https://ru.wikipedia.org/wiki/%D0%A2%D0%B5%D1%80%D0%BF%D0%B5%D0%BD%D0%B8%D1%8F_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>
(широко открыты к югу) находятся соответственно в южной и средней части
острова. В береговой линии выделяются два крупных залива и четыре полуострова.
В рельефе Сахалина выделяют
следующие 11 районов:
. Полуостров Шмидта <https://ru.wikipedia.org/wiki/%D0%9F%D0%BE%D0%BB%D1%83%D0%BE%D1%81%D1%82%D1%80%D0%BE%D0%B2_%D0%A8%D0%BC%D0%B8%D0%B4%D1%82%D0%B0>
(около 1,4 тыс.км²) - гористый
полуостров на крайнем севере острова с крутыми, местами обрывистыми берегами и
двумя меридиональными хребтами - Западным и Восточным
<https://ru.wikipedia.org/wiki/%D0%92%D0%BE%D1%81%D1%82%D0%BE%D1%87%D0%BD%D1%8B%D0%B9_%D1%85%D1%80%D0%B5%D0%B1%D0%B5%D1%82_(%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD)>;
высшая точка - г. Три Брата (623 м); соединён с Северо-Сахалинской равниной
<https://ru.wikipedia.org/wiki/%D0%A1%D0%B5%D0%B2%D0%B5%D1%80%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%B0%D1%8F_%D1%80%D0%B0%D0%B2%D0%BD%D0%B8%D0%BD%D0%B0>
Охинским перешейком, ширина которого в самом узком месте - чуть более 6 км;
. Северо-Сахалинская равнина
<https://ru.wikipedia.org/wiki/%D0%A1%D0%B5%D0%B2%D0%B5%D1%80%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%B0%D1%8F_%D1%80%D0%B0%D0%B2%D0%BD%D0%B8%D0%BD%D0%B0>
(около 28 тыс.км²) - пологохолмистая
территория к югу от полуострова Шмидта
<https://ru.wikipedia.org/wiki/%D0%9F%D0%BE%D0%BB%D1%83%D0%BE%D1%81%D1%82%D1%80%D0%BE%D0%B2_%D0%A8%D0%BC%D0%B8%D0%B4%D1%82%D0%B0>
с широко разветвлённой речной сетью, слабо выраженными водоразделами и отдельными
невысокими горными хребтами, тянется от залива Байкал на севере до слияния рек
Ныш <https://ru.wikipedia.org/wiki/%D0%9D%D1%8B%D1%88> и Тымь
<https://ru.wikipedia.org/wiki/%D0%A2%D1%8B%D0%BC%D1%8C> на юге, высшая
точка - г. Даахуриа (601 м); северо-восточное побережье острова выделяется в
качестве подрайона, для которого характерны крупные лагуны (наиболее крупные -
заливы Пильтун, Чайво, Ныйский, Набильский, Луньский), отделённые от моря
узкими полосами намывных кос, дюны, низкие морские террасы - именно в этом
подрайоне и на прилежащем шельфе Охотского моря находятся основные сахалинские
нефтяные и газовые месторождения;
. Западно-Сахалинские горы
<https://ru.wikipedia.org/wiki/%D0%97%D0%B0%D0%BF%D0%B0%D0%B4%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%B8%D0%B5_%D0%B3%D0%BE%D1%80%D1%8B>
тянутся почти на 630 км от широты с. Хоэ (51º19' с.ш.) на севере до
полуострова Крильон
<https://ru.wikipedia.org/wiki/%D0%9F%D0%BE%D0%BB%D1%83%D0%BE%D1%81%D1%82%D1%80%D0%BE%D0%B2_%D0%9A%D1%80%D0%B8%D0%BB%D1%8C%D0%BE%D0%BD>
на крайнем юге острова; средняя ширина гор - 40-50 км, наибольшая (на широте
мыса Ламанон) - около 70 км; осевую часть образуют Камышовый (севернее
перешейка Поясок) и Южно-Камышовый хребты;
. Тымь-Поронайская
низменность <https://ru.wikipedia.org/w/index.php?title=%D0%A2%D1%8B%D0%BC%D1%8C-%D0%9F%D0%BE%D1%80%D0%BE%D0%BD%D0%B0%D0%B9%D1%81%D0%BA%D0%B0%D1%8F_%D0%BD%D0%B8%D0%B7%D0%BC%D0%B5%D0%BD%D0%BD%D0%BE%D1%81%D1%82%D1%8C&action=edit&redlink=1>
располагается в средней части острова и представляет собой холмисто-увалистую
низменность, простирающуюся примерно на 250 км в меридиональном направлении -
от залива Терпения
<https://ru.wikipedia.org/wiki/%D0%A2%D0%B5%D1%80%D0%BF%D0%B5%D0%BD%D0%B8%D1%8F_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>
на юге до слияния рек Тымь
<https://ru.wikipedia.org/wiki/%D0%A2%D1%8B%D0%BC%D1%8C_(%D1%80%D0%B5%D0%BA%D0%B0)>
и Ныш на севере; максимальной ширины (до 90 км) достигает в устье реки Поронай
<https://ru.wikipedia.org/wiki/%D0%9F%D0%BE%D1%80%D0%BE%D0%BD%D0%B0%D0%B9_(%D1%80%D0%B5%D0%BA%D0%B0)>,
минимальной (6-8 км) - в долине реки Тымь
<https://ru.wikipedia.org/wiki/%D0%A2%D1%8B%D0%BC%D1%8C_(%D1%80%D0%B5%D0%BA%D0%B0)>;
на севере переходит в Набильскую низменность; покрыта мощным чехлом
кайнозойских осадков, сложена осадочными отложениями четвертичного периода:
песчаниками, галечниками; сильно заболоченная южная часть низменности носит
название Поронайской «тундры»;
. Сусунайская низменность
<https://ru.wikipedia.org/w/index.php?title=%D0%A1%D1%83%D1%81%D1%83%D0%BD%D0%B0%D0%B9%D1%81%D0%BA%D0%B0%D1%8F_%D0%BD%D0%B8%D0%B7%D0%BC%D0%B5%D0%BD%D0%BD%D0%BE%D1%81%D1%82%D1%8C&action=edit&redlink=1>
расположена в южной части острова и тянется примерно на 100 км от залива Анива
<https://ru.wikipedia.org/wiki/%D0%90%D0%BD%D0%B8%D0%B2%D0%B0_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>
на юге до реки Найба
<https://ru.wikipedia.org/wiki/%D0%9D%D0%B0%D0%B9%D0%B1%D0%B0_(%D1%80%D0%B5%D0%BA%D0%B0)>
на севере; с запада низменность ограничена Западно-Сахалинскими горами
<https://ru.wikipedia.org/wiki/%D0%97%D0%B0%D0%BF%D0%B0%D0%B4%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%B8%D0%B5_%D0%B3%D0%BE%D1%80%D1%8B>,
с востока - Сусунайским хребтом и Корсаковским плато; в южной части ширина
низменности достигает 20 км, в центре - 6 км, на севере - 10 км; абсолютные
высоты на севере и юге не превышают 20 м над уровнем моря, в центральной части,
на водоразделе бассейнов рек Сусуя и Большой Такой, достигают 60 м; относится к
типу внутренних низменностей и представляет собой тектоническую депрессию,
выполненную большой толщей четвертичных отложений; в пределах Сусунайской
низменности находятся города Южно-Сахалинск
<https://ru.wikipedia.org/wiki/%D0%AE%D0%B6%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA>,
Анива <https://ru.wikipedia.org/wiki/%D0%90%D0%BD%D0%B8%D0%B2%D0%B0_(%D0%B3%D0%BE%D1%80%D0%BE%D0%B4)>,
Долинск
<https://ru.wikipedia.org/wiki/%D0%94%D0%BE%D0%BB%D0%B8%D0%BD%D1%81%D0%BA>
и проживает около половины населения острова;
. Восточно-Сахалинские горы
<https://ru.wikipedia.org/wiki/%D0%92%D0%BE%D1%81%D1%82%D0%BE%D1%87%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%B8%D0%B5_%D0%B3%D0%BE%D1%80%D1%8B>
представлены на севере Лопатинским горным узлом (высшая точка - г. Лопатина,
1609 м) с радиально отходящими от него хребтами; два отрога противоположного
направления представляют собой Набильский хребет
<https://ru.wikipedia.org/w/index.php?title=%D0%9D%D0%B0%D0%B1%D0%B8%D0%BB%D1%8C%D1%81%D0%BA%D0%B8%D0%B9_%D1%85%D1%80%D0%B5%D0%B1%D0%B5%D1%82&action=edit&redlink=1>;
на юге Набильский хребет
<https://ru.wikipedia.org/w/index.php?title=%D0%9D%D0%B0%D0%B1%D0%B8%D0%BB%D1%8C%D1%81%D0%BA%D0%B8%D0%B9_%D1%85%D1%80%D0%B5%D0%B1%D0%B5%D1%82&action=edit&redlink=1>
переходит в Центральный хребет, на севере, резко понижаясь, - в
Северо-Сахалинскую равнину;
. Низменность полуострова
Терпения
<https://ru.wikipedia.org/wiki/%D0%9F%D0%BE%D0%BB%D1%83%D0%BE%D1%81%D1%82%D1%80%D0%BE%D0%B2_%D0%A2%D0%B5%D1%80%D0%BF%D0%B5%D0%BD%D0%B8%D1%8F>
- самый маленький из районов, занимает большую часть полуострова Терпения к
востоку от залива Терпения
<https://ru.wikipedia.org/wiki/%D0%A2%D0%B5%D1%80%D0%BF%D0%B5%D0%BD%D0%B8%D1%8F_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>;
. Сусунайский хребет
<https://ru.wikipedia.org/wiki/%D0%A1%D1%83%D1%81%D1%83%D0%BD%D0%B0%D0%B9%D1%81%D0%BA%D0%B8%D0%B9_%D1%85%D1%80%D0%B5%D0%B1%D0%B5%D1%82>
вытянут с севера на юг на 70 км и имеет ширину 18-120 км; высшие точки - гора
Пушкинская (1047 м) и пик Чехова <https://ru.wikipedia.org/wiki/%D0%9F%D0%B8%D0%BA_%D0%A7%D0%B5%D1%85%D0%BE%D0%B2%D0%B0>
(1045 м); сложен палеозойскими отложениями, у подножия западного макросклона
хребта находится город Южно-Сахалинск
<https://ru.wikipedia.org/wiki/%D0%AE%D0%B6%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA>;
. Корсаковское плато
<https://ru.wikipedia.org/wiki/%D0%9A%D0%BE%D1%80%D1%81%D0%B0%D0%BA%D0%BE%D0%B2%D1%81%D0%BA%D0%BE%D0%B5_%D0%BF%D0%BB%D0%B0%D1%82%D0%BE>
ограничено с запада Сусунайской низменностью <https://ru.wikipedia.org/w/index.php?title=%D0%A1%D1%83%D1%81%D1%83%D0%BD%D0%B0%D0%B9%D1%81%D0%BA%D0%B0%D1%8F_%D0%BD%D0%B8%D0%B7%D0%BC%D0%B5%D0%BD%D0%BD%D0%BE%D1%81%D1%82%D1%8C&action=edit&redlink=1>,
с севера - Сусунайским хребтом <https://ru.wikipedia.org/wiki/%D0%A1%D1%83%D1%81%D1%83%D0%BD%D0%B0%D0%B9%D1%81%D0%BA%D0%B8%D0%B9_%D1%85%D1%80%D0%B5%D0%B1%D0%B5%D1%82>,
с востока - Муравьёвской низменностью
<https://ru.wikipedia.org/w/index.php?title=%D0%9C%D1%83%D1%80%D0%B0%D0%B2%D1%8C%D1%91%D0%B2%D1%81%D0%BA%D0%B0%D1%8F_%D0%BD%D0%B8%D0%B7%D0%BC%D0%B5%D0%BD%D0%BD%D0%BE%D1%81%D1%82%D1%8C&action=edit&redlink=1>,
с юга -заливом Анива
<https://ru.wikipedia.org/wiki/%D0%90%D0%BD%D0%B8%D0%B2%D0%B0_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>,
имеет слабоволнистую поверхность, образованную системой плосковершинных
увалистых гряд, вытянутых в северо-восточном направлении; на южной оконечности
плато на берегу залива Анива
<https://ru.wikipedia.org/wiki/%D0%90%D0%BD%D0%B8%D0%B2%D0%B0_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>
находится город Корсаков
<https://ru.wikipedia.org/wiki/%D0%9A%D0%BE%D1%80%D1%81%D0%B0%D0%BA%D0%BE%D0%B2_(%D0%B3%D0%BE%D1%80%D0%BE%D0%B4)>;
. Муравьёвская низменность
<https://ru.wikipedia.org/w/index.php?title=%D0%9C%D1%83%D1%80%D0%B0%D0%B2%D1%8C%D1%91%D0%B2%D1%81%D0%BA%D0%B0%D1%8F_%D0%BD%D0%B8%D0%B7%D0%BC%D0%B5%D0%BD%D0%BD%D0%BE%D1%81%D1%82%D1%8C&action=edit&redlink=1>
расположена между заливами Анива
<https://ru.wikipedia.org/wiki/%D0%90%D0%BD%D0%B8%D0%B2%D0%B0_(%D0%B7%D0%B0%D0%BB%D0%B8%D0%B2)>
на юге и Мордвинова на севере, имеет увалистый рельеф с плоскими вершинами
увалов; в пределах низменности находится много озёр, в том числе так называемых
«Тёплых озёр», куда любят выезжать на отдых южно-сахалинцы;
. Тонино-Анивский хребет
<https://ru.wikipedia.org/w/index.php?title=%D0%A2%D0%BE%D0%BD%D0%B8%D0%BD%D0%BE-%D0%90%D0%BD%D0%B8%D0%B2%D1%81%D0%BA%D0%B8%D0%B9_%D1%85%D1%80%D0%B5%D0%B1%D0%B5%D1%82&action=edit&redlink=1>
вытянут с севера на юг, от мыса Свободного до мыса Анива, почти на 90 км,
высшая точка - гора Крузенштерна (670 м); сложен меловыми и юрскими
отложениями.
1.4 Климат
Климат Сахалина - умеренно-муссонный (средняя
температура января от −6ºС
на юге до −24ºС на севере,
августа - от +19ºС до +10ºС
соответственно), морской с продолжительной холодной снежной зимой и средним
тёплым летом. Среднегодовая температура на севере острова (по многолетним
данным) составляет около −1,5ºС,
на юге - +2,2ºС.
На климат влияют следующие факторы:
Географическое положение между 46º
и
54º
с.ш.
обусловливает приход солнечной радиации от 410 кДж/год на севере до 450 кДж/год
на юге.
Зимой погода в значительной степени определяется
Сибирским антициклоном: в это время преобладают северные и северо-западные
ветры, могут стоять крепкие морозы, особенно в центральной части острова с
умеренно континентальным микроклиматом. В то же время с юга могут приходить
зимние циклоны (которых практически нет в материковых районах российского
Дальнего Востока), определяющие сильные и частые бураны. Так, зимой 1970 г. на
область обрушились серия многоснежных циклонов, сопровождавшихся
многочисленными сходами снежных лавин. Ветер достигал ураганной силы (отдельные
порывы - до 50 м/сек), снежный покров в южной части Сахалина превышал норму в
3-4 раза, достигая в отдельных местах 6-8 м. Бураны парализовали работу всех
видов транспорта, морских портов, промышленных предприятий.
Положение между евразиатским материком и Тихим
океаном обуславливает муссонный характер климата. С ним связано влажное и
тёплое, довольно дождливое сахалинское лето. Лето начинается в июне и
заканчивается в сентябре.
Летом контраст между западным и
восточным побережьями острова усиливается за счёт соответственно тёплого
Цусимского течения Японского моря
<https://ru.wikipedia.org/wiki/%D0%AF%D0%BF%D0%BE%D0%BD%D1%81%D0%BA%D0%BE%D0%B5_%D0%BC%D0%BE%D1%80%D0%B5>,
которое достигает юго-западной оконечности Сахалина, и холодного
Восточно-Сахалинского течения
<https://ru.wikipedia.org/wiki/%D0%92%D0%BE%D1%81%D1%82%D0%BE%D1%87%D0%BD%D0%BE-%D1%81%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%BE%D0%B5_%D1%82%D0%B5%D1%87%D0%B5%D0%BD%D0%B8%D0%B5>
Охотского моря
<https://ru.wikipedia.org/wiki/%D0%9E%D1%85%D0%BE%D1%82%D1%81%D0%BA%D0%BE%D0%B5_%D0%BC%D0%BE%D1%80%D0%B5>,
идущего вдоль восточного побережья с севера на юг.
Максимальная температура на Сахалине
(+39ºС)
отмечалась в июле 1977 г. в пос. Пограничное на восточном побережье (Ногликский
<https://ru.wikipedia.org/wiki/%D0%93%D0%BE%D1%80%D0%BE%D0%B4%D1%81%D0%BA%D0%BE%D0%B9_%D0%BE%D0%BA%D1%80%D1%83%D0%B3_%D0%9D%D0%BE%D0%B3%D0%BB%D0%B8%D0%BA%D1%81%D0%BA%D0%B8%D0%B9>
район). Минимальная температура на Сахалине (-50ºС) была
зарегистрирована в январе 1980 г. в пос. Адо-Тымово (Тымовский
<https://ru.wikipedia.org/wiki/%D0%A2%D1%8B%D0%BC%D0%BE%D0%B2%D1%81%D0%BA%D0%B8%D0%B9_%D0%B3%D0%BE%D1%80%D0%BE%D0%B4%D1%81%D0%BA%D0%BE%D0%B9_%D0%BE%D0%BA%D1%80%D1%83%D0%B3_%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA%D0%BE%D0%B9_%D0%BE%D0%B1%D0%BB%D0%B0%D1%81%D1%82%D0%B8>
район). Зарегистрированный температурный минимумЮжно-Сахалинска
<https://ru.wikipedia.org/wiki/%D0%AE%D0%B6%D0%BD%D0%BE-%D0%A1%D0%B0%D1%85%D0%B0%D0%BB%D0%B8%D0%BD%D1%81%D0%BA>
- −36ºС (январь
1961 г.), максимум -
+34,7ºС
(август 1999 г.).
Глава 2.
общая характеристика мохообразных
.1
Происхождение и филогения
Происхождение и филогения любого
отдела растительного царства всегда полны глубочайшего интереса. И чрезвычайно
сложны. Что касается мохообразных, то здесь и научный интерес, и сложности
присутствуют, так сказать, «в двойном размере», если не в тройном.
Сложностей здесь больше потому, что
мохообразные, будучи очень древней группой, сравнительно плохо фоссилизуются,
то есть сохраняются в ископаемом состоянии. Поэтому изучение ископаемых
остатков бриофитов даст для выяснения их происхождения гораздо меньше
материала, чем можно было бы ожидать, учитывая тот факт, что мохообразные -
одни из древнейших наземных растений.
Интерес к происхождению этой группы
растений определяется ее положением в системе растительного царства.
Мохообразные - самые примитивные из ныне существующих высших растений. И хотя
они представляют собою слепую, боковую ветвь эволюции и не являются предками
остальных высших растений, все же они - ближайшие родственники этих предков. К
тому же родственники современные, а не вымершие. Вопрос о происхождении
мохообразных - это также в большей мере и вопрос о происхождении всех высших
растений вообще. Вот почему к нему постоянно обращаются ботаники разных
специальностей, а не одни только бриологи.
Как и почти в любом случае, когда
речь заходит о происхождении таксона самого крупного ранга - отдела, так и в
происхождении бриофитов много неясного, неопределенного, предположительного.
Наиболее древние остатки
мохообразных найдены в девонских отложениях (один вид печеночника) и в
отложениях верхней половины каменноугольного периода. Представлены эти остатки
талломными печеночниками и одним листостебельным мхом. Печеночники похожи на
маршанцию, листостебельный мох - на представителей семейства Polytrichaceae, к которому
относится уже многократно упоминавшийся кукушкин лен. Из этого, конечно, не
следует, что ископаемые остатки бриофитов могут рассматриваться как относящиеся
к роду маршанция и к семейству политриховых. И все же сходство (и родство) этих
древнейших (палеозойских!) организмов и ныне живущих просто поразительно. И
вряд ли можно оспаривать правомерность включения каменноугольных и девонских
мохообразных в состав современных порядков.
В следующем периоде палеозойской эры
- пермском - мохообразные несколько более многочисленны и разнообразны,
особенно листостебельные мхи, которых насчитывается около 20 видов, относящихся
к двум порядкам. При этом представители одного порядка (Protosphagnales) известны
только из пермских отложений. Ни до ни после они не обнаружены.
В отложениях более позднего возраста
количество видов ископаемых бриофитов резко увеличивается. В особенности это
относится к кайнозою. К сегодняшнему дню известно, вероятно, не менее 300 видов
ископаемых мохообразных. Из них на долю печеночников (антоцеротовые в
ископаемом состоянии неизвестны) приходится примерно 50-70 видов, остальное -
листостебельные мхи.
Самое существенное, что вытекает из
анализа ископаемых мохообразных, это следующее. Раньше девона мохообразные
достоверно неизвестны, листостебельные мхи - раньше каменноугольного периода,
листостебельные печеночники - раньше третичного периода. Наиболее древние из
известных ископаемых остатков бриофитов представлены уже довольно далеко
разошедшимися и продвинувшимися формами. И следовательно, возникли
мохообразные, скорее всего, значительно раньше того времени, из которого
известны их первые ископаемые остатки.
Обращает на себя внимание большая
степень близости ископаемых мохообразных к современным. Начиная с третичного
периода, большая их часть отождествляется с ныне живущими видами. Наиболее
отличающиеся от современных формы выделяются в особые роды и семейства и, как
исключение, в особые порядки, точнее в порядок. Ибо известен всего лишь один
порядок, представленный только ископаемыми формами - Protosphagnales.
Из всего этого напрашивается вывод о
большой древности и всего отдела мохообразных и большей части его подразделений
(всех классов, почти всех порядков, основной части семейств, не менее половины
родов).
Как современная бриология
представляет себе происхождение мохообразных?
На этот счет существует много
высказываний, взглядов, гипотез, точек зрения. Попытаемся все это многообразие,
конечно, с известной долей схематизации и упрощения сгруппировать по тому
общему, что их объединяет. Получится две основные группы гипотез. Первая
исходит из предположения, что предками мохообразных были псилофиты (риниофиты).
Вторая - выводит мохообразных из водорослей. Каждая из этих групп гипотез имеет
свои привлекательные стороны, и каждая же - весьма серьезные изъяны.
Рассмотрим каждую из этих групп
гипотез.
Первая группа. Возможные предки
мохообразных - псилофиты. Псилофиты и мохообразные - наиболее родственные,
близкие между собой отделы высших растений. Родство это настолько несомненно и,
можно сказать, велико, что ряд ботаников один из классов мохообразных -
антоцеротовые - даже выделяет из отдела мохообразных в включает в состав отдела
псилофитов. И хотя большинство бриологов эту точку зрения не разделяет, все же
несомненно, что из ничего она возникнуть не могла, что для этого необходимы
серьезные основания.
Одно из наиболее ярких свидетельств
родства псилофитов и мохообразных - замечательное сходство в строении спорангия
псилофитов и коробочки мохообразных. У части псилофитов и у некоторых
мохообразных (у антоцеротовых, сфагнов и андреевых) спорангий и коробочка
(которая тоже является спорангием, просто в отношении мохообразных термин «спорангий»
не особенно употребителен) устроены очень сходно. Напомним это строение.
Бокаловидная в схеме коробочка заполнена клетками двух родов: материнскими
клетками спор (впоследствии спорами), образующими споровый мешок, и стерильными
клетками - колонкой. Колонка представляет нечто вроде столбика-стерженька,
поднимающегося со дна коробочки. Споровый мешок накрывает ее сверху наподобие
колокола или шапки.
Такое сходство, конечно, не
случайно. И проходить мимо него нельзя. Другое дело - какие выводы делать. Можно
ли на основании этого сходства (понятно, с учетом и других признаков) считать,
что одна группа растений возникла из другой, а именно мохообразные из
псилофитов? В принципе, конечно, можно. По нельзя забывать и того, что чисто
теоретически можно допустить, по меньшей мере, три варианта объяснения этого
сходства: мохообразные возникли из псилофитов, псилофиты - из мохообразных, обе
группы растений имеют независимое происхождение от общих предков. И какой из
этих вариантов ближе к истине, сказать (обоснованно доказательно!) очень
нелегко.
Большой группой бриологов и вообще
ботаников, занимающихся этим вопросом, был взят на вооружение первый вариант. И
главное основание для такого подхода - до недавнего времени были псилофиты,
известные из гораздо более древних отложений, чем те, в которых обнаружены
самые древние мохообразные. Значит, вроде бы псилофиты возникли гораздо раньше
мохообразных, явившись первыми наземными растениями. И взгляд на мохообразных
как на потомков псилофитов в этом случае вполне понятен.
Но действительно ли псилофиты старше
мохообразных? Еще недавно самые ранние бриофиты датировались карбоном. Сейчас
они известны уже из девона. И мы довольно уверенно можем говорить о
приблизительно одновременном появлении на «свет» и псилофитов и бриофитов, по
крайней мере талломных печеночников.
А если так, то очень трудно одну
группу растений выводить из другой.
Есть еще некоторые моменты,
затрудняющие попытки рассматривать псилофиты в качестве возможных предков
мохообразных. Мы знаем, что псилофиты были сравнительно крупными растениями с
довольно сложным и высоко организованным спорофитом, способным к
самостоятельной жизни. Считать мохообразные потомками псилофитов, значит,
допускать возможность (вернее, необходимость) значительной деградации спорофита
и утерю им способности к самостоятельному существованию. Не очень понятны
причины такой деградации. Почему растения с более развитым спорофитом
(псилофиты) вымерли, а их потомки с сильно деградировавшим спорофитом
(мохообразные) сохранились, выжили, несмотря на то, что в целом у наземных
растений спорофитная линия эволюции оказалась единственно плодотворной и
перспективной. Все наземные высшие растения (кроме мохообразных) осуществили
именно спорофитную линию эволюции.
Вторая группа гипотез. Возможные предки
мохообразных - водоросли. Здесь проблема возникновения многоклеточных
гаметангиев приобретает первостепенное значение (у водорослей их нет, или они
устроены очень своеобразно и не могут рассматриваться в качестве исходных для
возникновения архегония и антеридия). Она в центре внимания исследователей,
видящих в водорослях возможных предков мохообразных. Еще в начале XX века
английский ботаник Б. Дэвис нарисовал картину предположительного происхождения
архегониев и антеридиев из многокамерных гаметангиев того типа, что наблюдается
у бурых водорослей. Основной путь превращения многокамерного гаметангия бурых
водорослей в многоклеточные гаметангии мохообразных - частичная стерилизация
материнских клеток гамет.
Многокамерный гаметангий бурых
водорослей - карпогон - представляет собой овальное или веретеновидное тело,
сидящее на ножке. Все клетки карпогона фертильны. Все - в самом прямом и точном
смысле, то есть, включая и те, что образуют наружный слой - стенку карпогона.
Все они являются материнскими клетками гамет. Располагаются эти клетки в
карпогоне не хаотично, а правильными рядами-этажами, направленными
перпендикулярно длине карпогона. Мужские и женские карпогоны устроены одинаково
и различаются в основном размерами - собственными и производимых ими гамет.
Последние снабжены каждая двумя жгутиками и могут двигаться самостоятельно (и
мужские и женские).
Представим, что под влиянием
наземного образа жизни у выходящих на сушу водорослей произошла частичная
стерилизация материнских клеток гамет. Наиболее вероятна потеря фертильности
клетками наружного слоя карпогона, превращение их в оболочку, предохраняющую
заключенные внутри нее материнские клетки гамет. В этом случае мы получим
антеридий и ничто другое. Кстати, в молодом возрасте материнские клетки антерозоидов
располагаются в антеридии точь-в-точь, как и материнские клетки гамет в
карпогоне, - правильными рядами-этажами, ориентированными перпендикулярно длине
антеридия. А если допустить, что стерильными стали все клетки, кроме одной
(яйцеклетки), то мы получим архегоний в чистом виде.
И все же надо сказать, что в идее Б.
Дэвиса о происхождении архегониев и антеридиев путем частичной стерилизации
фертильных клеток карпогона или органа, близкого к нему по строению и функциям,
очень много подкупающего. Чрезвычайно заманчиво выглядит эта идея.
Руководствуясь ею, можно сравнительно легко вывести архегонии и антеридии из
органа типа карпогона бурых водорослей. Вывести, пусть и чисто теоретически, но
все-таки без лишнего нагромождения недоказанных допущений и умозрительных
предположений, оставаясь, так сказать, на твердой земле фактов. И это уже
немало.
Трудность подкарауливает с другой
стороны. Два момента существенно осложняют гипотезы водорослевого происхождения
мохообразных, в том числе и нарисованную выше картину возможного происхождения
многоклеточных гаметангиев,
Первый момент заключается в том, что
гаметангии типа бегло охарактеризованного карпогона бурых водорослей есть
только у бурых водорослей. У зеленых ничего похожего нет. Ни у современных, ни
у ископаемых. И даже, если у них встречаются иногда многоклеточные гаметангии
(у харовых и у Chaetonema), то
рассматривать эти органы как возможные морфологические предшественники
архегониев и антеридиев очень трудно. Между тем и мохообразные, и все остальные
архегониатные растения - это растения зеленые. И пигментный состав, и вся
физиология, и вся биохимия у них совсем не те, что у бурых водорослей.
Второй осложняющий момент.
Мохообразные хотя и стоят ближе всех остальных современных растений к
водорослям, все же очень далеки от них. И выводить мохообразные непосредственно
из водорослей - все равно зеленых или бурых, современных или ископаемых,
реальных или полностью гипотетических (выдуманных) - крайне трудно. Возникает
необходимость в промежуточном звене - каких-то полуводорослях-полумохообразных.
А где их взять?
Вот и выясняется, что ни та ни
другая группа гипотез не решают вопроса о происхождении мохообразных достаточно
убедительно. Гипотезы псилофитного происхождения привлекают больше сторонников
и более распространены, чем водорослевые. Однако это не является следствием
большей убедительности или доказанности их. Предпочтение какой-либо гипотезы
сегодня не вывод из непреложных фактов, а личная точка зрения исследователя, не
столько вытекающая из фактов, сколько обусловленная не всегда понятными
«поворотами мысли». К тому же в некоторых из последних вариантов, исходящих из
предположения, что мохообразные произошли от каких-то сравнительно просто
устроенных, первичных гипотетических псилофитов, эти гипотезы в какой-то мере
смыкаются с водорослевыми гипотезами. Ведь эти гипотетические первичные
псилофиты в сущности и есть то промежуточное звено между водорослями и
мохообразными, те полуводоросли-полумохообразные, о которых говорилось выше.
Таким образом, утверждать, что
вопрос о происхождении мохообразных решен сколько-нибудь удовлетворительно,
будет, пожалуй, рановато. Слишком много еще неясного, неопределенного, в том
числе и в вопросах филогении этой группы. Да и трудно было бы ожидать иного:
происхождение и филогения тесно между собою связаны, и нерешенность одной
проблемы сразу же ставит массу вопросов в другой.
Почти без преувеличений можно
сказать, что в филогении мохообразных нет пока убедительных решений основных
вопросов. Так, мы не знаем, в какой последовательности появлялись классы и
подклассы мохообразных. Более или менее единодушны бриологи в признании за
печеночниками большего возраста, чем за листостебельными мхами. Предположим,
это - верная точка зрения. Но и она - скорее предположение, чем вывод из фактов.
Совершенно неясным остается время появления антоцеротовых. Мы уже упомянули,
что в ископаемом состоянии они неизвестны. Правильнее будет сказать, что
неизвестны макроостатки. Споры обнаружены в отложениях третичного возраста. А
ведь по многим признакам, например наличию хлоропласта с пиреноидом, именно
антоцеротовые наиболее близко стоят к водорослям. Какова последовательность
появления порядков, каковы их родственные связи? Ответа, четкого,
доказательного, нет.
В попытках выйти из этих сложностей
родилась точка зрения, исходящая из предположения, что мохообразные
представляют собою полифилетическую по происхождению (возникшую от разных
предков) группу, что одни предки их дали талломные формы, а другие -
листостебельные. Те и другие, следовательно, развивались сами по себе, не
превращаясь одна в другую. Бесспорно, такой подход снимает массу вопросов,
касающихся истории мохообразных. Но возникают другие, и вопрос о
непосредственных предках мохообразных почти совсем теряет ясность.
В оправдание бриологам надо сказать,
что ответов на перечисленные вопросы нет не по их вине. Дело в объективных,
очень больших трудностях. Ведь мохообразные представлены рядом далеко
разошедшихся, часто очень слабо между собой связанных, мало родственных линий
развития. Эти линии - циклы форм различного ранга от рода до класса, как бы
замкнуты внутри себя, не связаны переходами друг с другом. Часть линий развития
представлена крайне олиготипными или даже монотипными таксонами, нередко
довольно высокого ранга. Например, монотипный порядок Schistostegales с одним
видом Schistostega
pennata,
олиготипные порядки Takakiales с одним родом и двумя видами, Tetraphidales с двумя
родами и тремя видами, Phyllothalliales с одним родом и двумя видами
и т. п.
Такой характер родственных связей
крайне затрудняет установление степени родства между отдельными группами
мохообразных и построение системы бриофитов. Он свидетельствует об очень
большой древности мохообразных, о вымирании многих промежуточных звеньев,
когда-то связывавших ныне далеко друг от друга отстоящие группы.
И все это вместе очень затрудняет
прояснение филогении мохообразных. К многочисленным трудностям надо добавить
еще одну, сугубо бриологическую. В жизненном цикле мохообразных присутствуют
две ядерных фазы - гаметофаза и спорофаза. И хотя одна без другой (именно
спорофаза без гаметофазы, но не наоборот) жить не может, эволюция той и другой
может идти и независимо, сама по себе, с разной скоростью, да подчас еще и в
разных направлениях! Случаи, когда мы имеем относительно развитый, продвинутый
гаметофит и примитивный или деградировавший спорофит, или прямо
противоположные, с развитым спорофитом и примитивным или редуцированным
гаметофитом нередки среди мохообразных. Примером первого может служить порядок Polytrichales, у
представителей его высокоразвитый гаметофит с проводящей системой наиболее
сложного для мохообразных типа и вместе с тем довольно примитивно устроенный
спорофит с мелкими и малоподвижными зубцами перистома. Аналогичный пример
показывают и сфагны: сложнейше устроенный гаметофит и весьма примитивный, во
всяком случае, явно отставший от него в развитии спорофит. Противоположный
пример - представители порядка Buxbaumiales: у них
очень сложно устроенный крупный спорофит и мелкий, деградировавший (или
примитивный?) гаметофит. Сложно и разнообразно устроены спорофиты у
представителей семейства Splachnaceae при
относительной простоте и однообразии в строении гаметофита.
Но всем этим трудности еще не
исчерпываются. Добавляет их мозаичность в распределении признаков: какой-нибудь
таксон, по всей сумме признаков, несомненно, принадлежащий определенному
классу, вдруг обнаруживает признаки, свойственные другому классу, причем не
всегда самому близкому. Так, колоколообразное строение спорового мешка
наблюдается у антоцеротовых, а затем у сфагнов и андреевых мхов, но отсутствует
у печеночников. Между тем в целом печеночники, несомненно, гораздо ближе к
антоцеротовым, чем сфагновые и андреевые. А в свою очередь сфагны и андреевые,
при всем их несомненном своеобразии, все же гораздо ближе к бриевым мхам,
нежели к печеночникам и тем более к антоцеротовым. Посредством створок
вскрываются коробочки у антоцеротовых, у многих печеночников и у андреевых. По
всем остальным признакам и общей их сумме андреевые мхи ближе к другим
подклассам листостебельных мхов, чем к печеночникам и антоцеротовым.
Однако бриологи не падают духом. И
продолжают пробиваться сквозь многочисленные трудности к познанию происхождения
и филогении мохообразных.
2.2 Как
устроены мохообразные
Мохообразные - небольшие многолетние
или реже однолетние растения, талломные (слоевищные) или листостебельные,
размножающиеся спорами. Многолетние мохообразные - вечнозеленые: таллом на зиму
не отмирает, листья не опадают, и то и другое после периода зимнего покоя
весной возобновляет жизнедеятельность. Период же зимнего покоя короче, чем
можно было бы думать. Установлено, что многие мхи активно фотосинтезируют под
снежным покровом.
Зеленое тело мохообразных, как
говорилось выше, гаметофит или гаметофаза. Оно может быть талломным или листостебельным.
Таллом - это просто лепешка или лента шириной в несколько миллиметров
(максимально до 5 см у южноамериканско-новозеландского печеночника Monoclea
forsteri)
и длиной в несколько сантиметров, чаще не более 4-6. Максимальную для талломных
печеночников длину - до 20 см - имеет все та же Monoclea
forsteri.
Таллом дихотомически ветвится, что особенно хорошо заметно, когда он имеет
форму неширокой ленты. Он всегда стелется и более или менее прижат к субстрату.
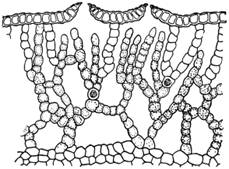
Рис. 1. Поперечный срез слоевища
печеночника, относящегося к подклассу Маршанциевых, - Mannia
brachypoda.
Видны воздушные камеры, ассимиляционные нити (не достигающие верхнего слоя
клеток клеточные ряды) и продушины в верхнем слое клеток слоевища. Пример
довольно сложно устроенного слоевища.
Внутренне строение таллома довольно
разнообразно. У одних видов он большей частью состоит из вполне однородных
клеток. У других же печеночников из подкласса маршанциевых таллом устроен
сложно (рис. 1). Верхняя стенка его образована одним слоем прозрачных клеток. В
стенке имеются особые отверстия - продушины или норы, называемые иногда
устьицами (но ничего общего с настоящими устьицами не имеющие). Эти продушины
ведут в разделенные вертикальными перегородками и заполненные воздухом полости
(камеры). Со дна их поднимаются нити, состоящие из хлорофиллоносных клеток. Эти
нити называются обычно ассимиляционными или ассимиляторами.
Листостебельное растение
мохообразного имеет ряд отличий от остальных листостебельных высших растений.
У всех без исключения бриофитов
листья всегда сидячие, не имеют черешка. Пластинка листа прикрепляется
непосредственно к стеблю. Она может располагаться перпендикулярно стеблю
(подавляющее большинство листостебельных мхов), косо (большинство листостебельных
печеночников) и продольно (только один вид из класса листостебельных мхов - Schistostega
pennata,
относящийся к отдельному монотипному порядку Schistostegales).
Лист устроен довольно разнообразно.
Чаще всего он представляет собой однослойную (из одного слоя клеток) пластинку
с жилкой или без нее. Форма - от линейной до округлой или даже
поперечно-широкой (с шириной, превышающей длину). У листостебельных мхов
пластинка листа всегда цельная, у печеночников часто в той или иной степени
раздельная. Нередко лист продольно складчатый или поперечно-волнистый.
Поверхность листьев не всегда
гладкая. Во многих случаях она «всхолмлена» выступающими частями клеток. В
особенности разнообразны по форме, размерам, количеству (на одну клетку)
выступы клеточной оболочки - папиллы.
Располагаются листья по стеблю
большей частью спирально, и стебель чаще всесторонне облиственный.
У листостебельных печеночников
(стебель у них обычно не бывает прямостоячим, в большинстве случаев он вытянут
вдоль субстрата) листья располагаются двумя четко выраженными рядами, идущими
по бокам стебля. У большинства видов есть, кроме того, и третий ряд листьев,
расположенный внизу, на брюшной (обращенной к субстрату) стороне стебля. По
форме и размерам эти брюшные листья обычно отличаются от спинных и носят особое
название - амфигастрии.
У всех без исключения мохообразных
отсутствуют корни. Прикрепление к субстрату (отчасти также поглощение влаги из
почвенных растворов) осуществляется с помощью ризоидов. Они представляют собою
нити из одного ряда клеток, разделенных косыми перегородками. Это у
листостебельных мхов. У печеночников и антоцеротовых ризоиды всего лишь очень
сильно вытянутые в длину одиночные клетки.
На гаметофите образуются гаметангии
- антеридии и архегонии. И те и другие в принципе устроены одинаково у всех
архегониатных растений. Антеридий - удлиненное, несколько мешковидное, у
печеночников чаще округлое тело. Он имеет стенку из одного слоя стерильных
клеток. Все внутреннее пространство его заполнено материнскими клетками
антерозоидов - мужских гамет. При созревании антеридия на его удаленном от
места прикрепления конце возникает отверстие, и снабженные каждый двумя
жгутиками антерозоиды выходят наружу. Они способны самостоятельно двигаться в
водной среде.
Архегоний - чаще узкоколбовидное или
бутылковидное тело с узкой и длинной верхней частью (шейкой) и расширенной
нижней (брюшком). Шейка имеет стенку из одного слоя клеток, брюшко - из
нескольких. В брюшке располагается неподвижная яйцеклетка - женская гамета.
В результате оплодотворения
(возможного только при наличии воды, только в ней могут двигаться антерозоиды)
возникает зигота, дающая начало спорофиту или спорофазе. Наиболее часто,
впрочем, его называют третьим именем - спорогоном. Спорофит - спороносный орган
и состоит из трех частей: стопы, ножки и коробочки. Стопа - нижняя часть
спорогона, с ее помощью последний внедряется в ткань гаметофита, прикрепляется
к нему механически и получает от него питательные вещества и воду. Ножка - это
стержень (у листостебельных мхов довольно прочный), на верхушке которого сидит
коробочка. Коробочка - собственно и есть спорангий, орган, в котором образуются
споры. У листостебельных мхов она представляет собою чаще бокаловидное
образование с широким отверстием наверху (рис. 2), закрытым особым щитком с
весьма обыденным названием - крышечка. Представьте себе туристский котелок с
суженным основанием, закрытый крышкой. Это и будет коробочка наиболее
распространенного у листостебельных мхов типа. Только для полноты картины
котелок надо еще прикрыть сверху шапкой. Она будет изображать колпачок -
специальное образование, закрывающее сверху коробочку. Форма и размеры колпачка
очень разнообразны (рис. 3). Наиболее часто колпачок напоминает капюшон,
разрезанный с одной стороны. В этом случае он называется клобуковидным. Но
нередко колпачок имеет шапочковидную или колоколовидную форму. Обычно он голый,
но довольно часто покрыт (редкими или густыми) волосками. В особенности
характерно наличие густого покрова из волосков для представителей порядка, к
которому относится хорошо известный мох - кукушкин лен, или политрихум
обыкновенный (Polytrichum
commune).
По этому признаку и порядок и род получили название Polytrichum (в переводе
с латинского языка означает многовласник).
В молодом возрасте коробочка, как
правило, целиком или почти целиком закрыта колпачком, в зрелом - лишь частично.
Роль и функции колпачка ясны - он защищает молодой спорогон и созревающие в нем
споры от неблагоприятных воздействий внешней среды, главным образом резких
колебаний температуры и влажности.

Рис. 2. Открытая коробочка
листостебельного мха. Хорошо виден простой перистом по краю устья коробочки.
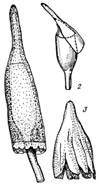
Рис. 3. Различные типы колпачков.
- колокольчатый представителя
порядка Encalyptales; 2 -
клобуковидный;3 - шапочковидный.
Удивляет не функциональное
назначение колпачка, а его происхождение. Если весь спорогон - стопа, ножка,
коробочка с крышечкой - возникли из зиготы, то колпачок - из стенки архегония.
И, значит, он - не спорофит, а часть гаметофита! Странно, не правда ли? Но еще
более странен колпачок некоторых печеночников - он возникнет не только из
стенки архегония, но и из участков стебля или слоевища (таллома), частично
срастающихся со стопкой архегония.
В отличие от крышки, закрывающей
котелок, крышечка на созревающей коробочке листостебельных мхов лежит не
свободно, а как бы прирастает к краю коробочки и составляет с нею одно целое.
Между нижним краем крышечки и верхним краем коробочки (он называется устьем)
располагаются один или несколько рядов особым образом устроенных
гигроскопических клеток. Это колечко, клетки которого имеют резко утолщенные с
одной стороны стенки. При созревании коробочки происходит ее частичное
обезвоживание, подсыхание. Клетки колечка теряют воду. В них создается
напряжение, так как, подсыхая, они уменьшаются в объеме, и неутолщенные части
стенок начинают втягиваться внутрь. Когда напряжение и силы сцепления между
клетками колечка и краями крышечки и коробочки становятся почти равными, тогда достаточно
очень небольшого усилия и произойдет разрыв колечка. Оно распадается на
отдельные клетки или куски из нескольких клеток. Крышечка при этом отпадает,
отделяется от коробочки, и созревшие споры начинают высыпаться наружу. Усилие,
которое разорвет колечко и обеспечит открывание коробочки, могут создать,
например, легкое дуновение ветерка или насекомое, севшее на коробочку.
Высыпающиеся споры служат для
бесполого размножения. Возникают они из особых клеток, называющихся
материнскими. При образовании спор происходит редукционное деление, и они имеют
уже гаплоидный набор хромосом. Споровый мешок, или археспорий, из которого
образуются споры, не заполняет обычно всего внутреннего пространства коробочки.
У очень многих видов листостебельных мхов и у антоцеротовых в центре коробочки
располагается столбик-стерженек из стерильных клеток. Называется этот стерженек
колонкой. Основная функция колонки - обеспечение споровой массы влагой и
питательными веществами. Споровый мешок либо прикрывает колонку сверху наподобие
шапки или колокола (у антоцеротовых, у сфагнов и у андреевых мхов), либо
окружает ее в виде цилиндра, открытого и снизу и сверху (у бриевых мхов).
Отсутствует колонка у печеночников.
Стенка коробочки чаще всего
многослойная, но у некоторых видов печеночников она на всем протяжении или
только в верхней части однослойная. В стенке коробочки нередко встречаются
устьица.
У очень многих видов мхов,
относящихся к подклассу бриевых класса листостебельных (и только у них!),
коробочка снабжена чрезвычайно оригинальным образованием, не имеющим аналогов
ни в какой другой группе растений, - перистомом. Это - своеобразный частокол
одного или двух (расположенных концентрически - один внутри другого) рядов
зубцов, обрамляющих устье коробочки. При закрытой коробочке перистом находится
под крышечкой, внутри ее.
Вариантов строения перистома
множество. Так и хочется сказать: бесконечное множество. Но это хоть и не
бесконечное, но все же множество, которое укладывается в два основных типа:
нечленистый перистом и членистый. Соответственно этому выделяются две группы
порядков нечленистозубые и членистозубые. Внутри каждого типа есть перистом
простой - из одного ряда зубцов и двойной, образованный двумя рядами.
Рассмотрим каждый из перистомов.
Примеры представлены на рис. 4 и 5.
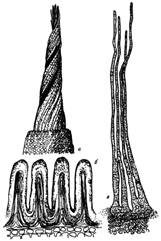
а - спирально завитой перистом Barbula
fallax,
представителя порядка Pottiales; б - нечленистый перистом
представителя порядка Polytrichales, в - почти нитевидный членистый перистом
Ptychomitrium
longisetum.
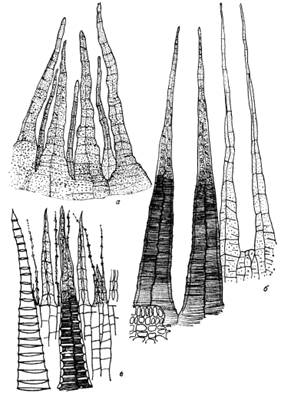
Рис. 5. Различные типы двойного
перистома.
а - перистом Lescuraea
striata
(отростки внутреннего перистома заметно короче зубцов наружного, реснички
отсутствуют); б - перистом Meteoriopsis reclinata. Оба
перистома примерно одинаковой длины, основная перепонка низкая, реснички
отсутствуют; в - перистом Rhynchostegium pallidifolium. Перистомы
одинаковой длины, внутренний на высокой основной перепонке, реснички развиты, с
узловатыми утолщениями. Хорошо заметно различие в строении наружной и
внутренней сторон зубцов наружного перистома (крайний левый - светлый - зубец
наружного перистома обращен к зрителю внутренней стороной, зубец, расположенный
правее, с поперечной штриховатостью - наружной)
Нечленистый перистом чаще всего
простой, однорядный. Образуется из нескольких концентрически расположенных
слоев специальных клеток. Зубцы у него обычно массивные, «толстые», имеющие в
радиальном (по отношению к устью коробочки) сечении треугольную или языковидную
форму. Состоят такие зубцы из целых мертвых клеток, которые у представителей
порядка Polytrichales вытянуты во
всю длину зубцов. В других случаях длин клетки меньше длины зубца. Но и при
этом зубцы не имеют поперечных перегородок.
Нечленистый перистом встречается у
очень небольшого числа мхов, относящихся к трем порядкам. Один из них только
что назван. Два других - это Tetraphidales и Buxbaumiales.
Членистый перистом свойствен
большинству мхов (тех, у которых есть перистом). Чаще он бывает двойным, то
есть состоит из двух перистомов - наружного и внутреннего. Образуется из двух
или трех концентрически расположенных рядов специальных клеток. Наружный
перистом - частокол сравнительно узких (почти линейных по форме) зубцов,
несколько расширенных внизу и суживающихся вверху. Каждый зубец отчленен от
основания, или же все они в основании между собой сливаются.
Зубцы наружного перистома
поперечно-членистые. Каждый состоит из серии низких долей-члеников, разделенных
поперечными по отношению к длине зубца перегородками. Получается нечто вроде
стопки книг, положенных друг на друга таким образом, что каждая верхняя чуть
меньше нижней. При рассматривании в профиль зубец с наружной стороны выглядит
ровным, края всех члеников подогнаны один к другому и образуют почти прямую
линию. С внутренней же стороны зубца картина совершенно иная. Здесь край зубца
выглядит так, словно на большой членик положен маленький и так далее. Линия
получается волнистая.
Но это лишь очень грубая схема
радиального сечения зубца наружного перистома. Членики-доли не занимают всей
толщины зубца, а разделены вертикальной перегородкой (ориентированной
фронтально) на два. И, таким образом, зубец состоит из двух слоев члеников -
наружного и внутреннего. Размеры тех и других не совпадают.
Перейдём к внутреннему перистому. О
его строении легче всего получить представление, если взять бумажную ленту и по
всей ее длине сделать узкотреугольные вырезы примерно на две трети ширины
ленты. На ленте возникают зубчики. После этого ленту надо сложить «гармошкой» и
соединить концы. Вот примерно так выглядит внутренний перистом, нечто вроде
гофрированной короны. Это килевато-складчатая пластинка (она называется
основной перепонкой), усаженная наверху отростками, имеющими более или менее
ланцетную форму. Часто отростки имеют отверстия, расположенные вдоль средней
линии. Между отростками нередко находятся реснички - тонюсенькие ниточки,
обычно как бы с узелочками (небольшими утолщениями, похожими на узелочки) или
же с особыми выростами, придающими ресничке перистый облик.
Мы рассмотрели наиболее типичное
строение двойного перистома. Перейдем к простому.
Очень часто зубцы простого перистома
похожи на зубцы наружного: такие же сравнительно плотные, «толстые», состоящие
из двух слоев клеток, поперечно-членистые и ланцетовидные по форме. Но в
отличие от зубцов наружного перистома зубцы простого нередко расщеплены сверху
на две-три узкие доли, иногда до основания зубца.
Во многих случаях строение простого
перистома очень своеобразно, и сходство с наружным перистомом в этих случаях
малозаметно. Любопытнейшим образом устроен перистом представителей семейства Pottiaceae, в
частности видов родов Tortula и Barbula. У них
зубцы перистома очень узкие, нитевидные и не прямые, как у большинства мхов, а
в один-два раза спирально закрученные (см. рис. 4), а у Tortula
pulvinata
даже в четыре раза. Сидят эти, зубцы на особой трубке (цилиндре) (бриологи
именуют ее тубусом), несколько напоминающей основную перепонку внутреннего
перистома, но в отличие от нее нескладчатой. Эта трубка у видов рода Tortula не является
аналогом основной перепонки двойного перистома. Равным образом и вообще простой
перистом не является аналогом, за редким исключением, наружного или внутреннего
перистома. Но в тех редких случаях, когда у некоторых видов редуцирован либо
наружный перистом, либо внутренний, тогда перистом, ставший простым, аналогичен
нередуцированному. И, как правило, очень похож на него.
Для полноты хотя бы и очень краткой
характеристики перистома необходимо добавить, что поверхность его зубцов (простого
или наружного перистома в случае, если он двойной), отростков, ресничек,
основной перепонки - словом, всех частей очень редко бывает совершенно ровной,
гладкой. Наоборот, почти всегда это сплошные бугры и впадины, выпуклости и
вмятины, очень разнообразные по форме и размерам, но все микроскопически
мелкие. Некоторые из этих «шероховатостей» видны лишь в сканирующий электронный
микроскоп. Заметим попутно, что использование сканирующего микроскопа в
бриологии показало наличие у мохообразных очень сложных и чрезвычайно
разнообразных поверхностных структур не только перистома, но и листьев и спор.
Многие из этих структур имеют, как выясняется, определенное значение с
систематической точки зрения.
Функции перистома немногочисленны,
но чрезвычайно ответственны. Он регулирует рассеивание (высыпание из коробочки)
спор и защищает находящиеся внутри уже открытой коробочки споры от намокания.
Каким образом это осуществляется? С
помощью движения зубцов перистома. Для этого, как и всюду в растительном мире,
где нужны движения, используется гигроскопичность. Перистом необыкновенно
гигроскопичен. Во влажную погоду зубцы его сгибаются внутрь и закрывают устье
коробочки, не давая спорам высыпаться и не пропуская влагу. Чем выше влажность,
тем плотнее смыкаются зубцы перистома. В сухую же погоду они, наоборот,
отгибаются наружу, открывая устье. Споры могут высыпаться.
Но это только схема. На самом деле
все гораздо сложнее и интереснее. Благодаря своему строению зубцы перистома
необычайно гибки и подвижны. Часто они не просто сгибаются внутрь и отгибаются
наружу. Бывают положения, когда нижняя часть зубца еще отогнута наружу, а в
середине его уже наблюдается резкий перегиб, и верхушка склоняется внутрь.
Получается так, что зубцы наружного перистома как бы вонзаются, втыкаются во
внутренний перистом. Неровности обоих перистомов, особенно отверстия отростков
внутреннего, в которые часто попадают зубцы наружного, приводят к тому, что
перистомы цепляются друг за друга. При движении зубцов наружного перистома
взад-вперед внутренний перистом встряхивается и частично разрушается.
Встряхивается и вся коробочка. Все это способствует быстрому высыпанию спор.
Работает, как мы видим, лишь
наружный перистом, внутренний малогигроскопичен, а значит, и малоподвижен. Его
роль в регулировании рассеивания спор всегда чисто пассивная: он лишь
препятствует их быстрому высыпанию. В тех случаях, когда надо поторапливаться,
используя благоприятные условия, наружный перистом начинает «наводить порядок»,
активно разрушая внутренний.
Добавим, что способностью к
гигроскопичным движениям обладает у многих видов и ножка, на которой сидит
коробочка, особенно, если она более или менее длинная (3-5 см). При уменьшении
влажности ножка начинает скручиваться по спирали, а при увеличении -
раскручиваться.
Степень гигроскопичности перистома
некоторых видов мхов фантастически велика, Длинные и тонкие, почти нитевидные
зубцы простого перистома мха Tayloria tenius (он растет
во многих районах Советского Союза) настолько чувствительны, что реагируют даже
на изменение влажности воздуха, вызываемое дыханием человека.
Случаи активных и быстрых движений в
растительном мире крайне редки. Наибольшей известностью пользуются два вида -
знаменитейшая мимоза стыдливая и, пожалуй, не менее знаменитая венерина
мухоловка. Листья мимозы при прикосновении очень быстро меняют свое положение в
пространстве. Почти молниеносно, теряя тургор, они складываются. Почти
мгновенно складываются и половинки листа венериной мухоловки, захлопываясь
подобно капкану, так что оказавшееся на верхней стороне листа насекомое не
успевает улететь. Но чтобы вернуться листьям этих растений в прежнее положение,
нужно время. Мимозе, во всяком случае, больше минуты, венериной мухоловке - еще
больше. Возвращаются листья в исходное положение медленно.
В обоих случаях быстрое движение
управляется изменением электрического потенциала клеток листа. Только на
гигроскопической основе происходит моментальное скручивание створок плода
недотроги, разбрасывающих при этом семена, растрескивание створок «стручков»
бобовых и т. д.
Однако во всех этих (и других
подобных) случаях быстрое движение происходит только один раз. Зубцы же
перистома тайлории работают непрерывно, пока действуют необходимые условия. Их
движения и быстры, и многократны. Впрочем, это относится не только к тайлории.
Подобной гигроскопичностью обладает немалое число видов. Нам не удалось найти в
литературе примеры других растений (не мхов), способных, подобно мхам,
производить на гигроскопической основе столь же быстрые и притом многократно
повторяющиеся движения. Возможно, таких растений и нет. Тогда мхи с их
сверхчувствительными и сверхподвижными зубцами перистома уникальны.
Так много места уделено
характеристике перистома, конечно, не случайно. Напомним, что перистом есть
только у мохообразных, точнее, в подклассе бриевых мхов. Не найдено ничего
похожего и у ископаемых растений. Перистом - это то, что в наибольшей степени
отличает бриевые мхи от остальных мохообразных и вообще от всех растений.
Однако не все виды бриевых мхов
используют для рассеивания и защиты спор такой сложный аппарат, как перистом.
Многие обходятся и без него. Внимательное изучение таких бесперистомных мхов
показывает, что в большинстве случаев коробочки их сравнительно короткие и
широкие, с широким устьем, по форме похожие на чайную чашку. А иногда такие
широкие и короткие, что их приходится сравнивать уже не с чашкой, а с глубокой
тарелкой. Из этих открытых сосудов споры могут высыпаться легко и быстро, даже
если отверстие коробочки расположено сверху. А чаще всего так и бывает. При такой
форме коробочки споры из нее могут быть выдуты малейшим движением воздуха,
сверхлегким ветерком. Поэтому широко открытая коробочка освобождается от спор
очень быстро, почти сразу же вслед за отделением крышечки. Оно происходит
всегда в сухую погоду.
Очень оригинальный способ вскрытия
коробочки и рассеивания спор избрали виды рода сфагнум (Sphagnum). Здесь
действует сжатый воздух, и недаром механизм вскрытия коробочки сфагнов обычно
сравнивают с механизмом действия воздушного ружья.
А происходит все следующим образом
(рис. 6). Созревающая коробочка сфагнов имеет форму, близкую к форме шара.
Почти все внутреннее пространство этого шара заполнено колонкой, которая
представляет собой бесформенную массу, занимающую примерно две трети полости
коробочки во всю ее ширину. Сверху на колонке лежит споровый мешок, состоящий
сначала из материнских клеток спор, а затем и из самих спор. По мере созревания
коробочки колонка усыхает и резко уменьшается в объеме. Пространство, которое
она занимала раньше, постепенно заполняется воздухом, поступающим снаружи через
устьица. Споровый мешок остается на прежнем месте - в верхней части коробочки.
Только лежит он теперь не на колонке, а на воздушной подушке. Затем устьица
закрываются, а коробочка постепенно начинает усыхать. Продольные стенки клеток,
образующих стенку коробочки, плотнее поперечных, и потому усыхание имеет
односторонний характер. Длина коробочки (в случае шара о длине говорить трудно,
будем считать длиной расстояние от нижней точки коробочки до ее устья) при этом
усыхании почти не меняется, в то время как ширина значительно уменьшается. И
шар постепенно превращается в цилиндр. Внутренний объем коробочки уменьшается.
Воздух внутри, не имея возможности выйти наружу (устьица закрыты, стенка
коробочки воздухонепроницаема), сжимается. Давление воздуха на стенки коробочки
может достигать 3-5 атмосфер! Он давит и на споровый мешок с уже созревшими
спорами. Тот в свою очередь давит на крышечку. И вот однажды это давление
оказывается больше, чем силы сцепления, удерживающие крышечку на месте. С
легким щелчком крышечка отрывается и взлетает вверх на 12-15 сантиметров.
Вместе с нею на ту же высоту взлетают выброшенные «зарядом» споры. Ни дать ни
взять - выстрел воздушного ружья. Происходит это чаще всего в солнечный день,
при наличии хотя бы небольшого ветерка, подхватывающего и разносящего легкие
споры.
краснокнижный мох
сахалин размножение
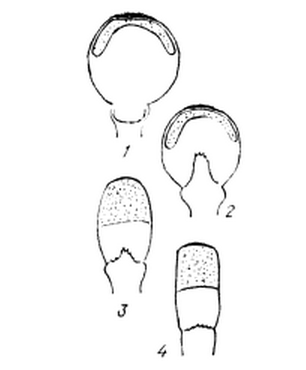
Рис. 6. Схема подготовки к вскрытию
коробочек сфагнов.
- вертикальный разрез через
созревшую коробочку, вверху располагается колоколообразный споровый мешок, вся
полость коробочки занята колонкой;2 - дальнейшая стадия созревания: колонка
усыхает и приобретает вид небольшого столбика в нижней части коробочки, бо́льшая часть
полости коробочки заполнена воздухом; 3, 4 - созревшая коробочка превращается
из шара в цилиндр, полость коробочки уменьшается, воздух в ней сжимается.
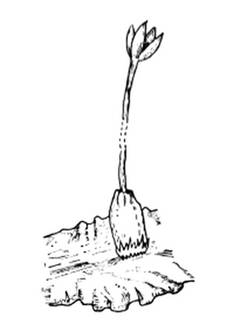
Рис. 7. Так выглядит вскрывшаяся
коробочка многих печеночников.
Этот способ вскрытия коробочки и
рассеивания спор у сфагновых мхов уникален. Впервые наблюдал его и описал еще в
конце прошлого века русский ботаник С. Г. Навашин. Впоследствии он обессмертил
свое имя открытием двойного оплодотворения у покрытосеменных. В начале своей
деятельности С. Г. Навашин занимался изучением мохообразных.
У антоцеротовых и печеночников
вскрытие коробочки осуществляется совсем не так, как у листостебельных мхов. У
них коробочка (в подавляющем большинстве случаев) не имеет крышечки и
вскрывается растрескиванием на две или чаще на четыре продольные створки. В
верхней части они отгибаются наружу, и тогда вскрывшаяся коробочка начинает
напоминать двух- или четырехлепестковый цветок (рис. 7). Иногда вскрытие
осуществляется посредством одной боковой продольной щели (Бардунов, 1984).
При таком способе вскрытия коробочки
споры довольно легко и быстро высыпаются наружу. Для ускорения и регулировки
этого процесса среди спор располагаются специальные стерильные клетки -
элатеры. Они очень тонкие и длинные, собственно, это нити, чрезвычайно
гигроскопичные. Своими движениями элатеры взрыхляют споровую массу, облегчают
высыпание спор через щели между створками коробочки. Чаще элатеры разбросаны
среди спор, иногда собраны пучками и одним концом прикреплены к внутренней
стороне коробочки.
Своеобразно устроен спорогон у
антоцеротовых. Он вытянут и имеет почти игольчатую форму. Растрескивается двумя
или четырьмя створками. А главное - спорогон нарастает снизу в течение всей
своей жизни. Это наблюдается только у антоцеротовых.
У андреевых мхов коробочка тоже
растрескивается на четыре створки. Но они остаются соединенными и внизу и
вверху.
У некоторых видов (это относится и к
листостебельным мхам, и к печеночникам) коробочка вскрывается посредством
неправильных разрывов стенки. Вероятно, это или наиболее примитивный способ
вскрытия коробочки, или, во всяком случае, один из них.
Спора, заботе о которой растением
уделяется так много внимания, сил и средств, - это клетка с гаплоидным набором
хромосом. Попав в благоприятные условия, она прорастает. Однако не сразу в
листостебельное или талломное растение, которое мы потом назовем мхом,
печеночником или антоцеротовым. Вначале возникает протонема - чаще это
разветвленная нить из одного ряда клеток, чрезвычайно похожая на зеленые
нитчатые водоросли. Но в отличие от водорослей, имеющих прямые перегородки
между клетками нити, перегородки у протонемы косые. На нитях протонемы
возникают почки, из которых и возникает взрослое растение мохообразного. Иногда
протонема бывает не нитчатой, а пластинчатой (образованной одним или изредка
несколькими слоями клеток) или отсутствует вовсе.
Вместе с тем размножение с помощью
спор в большинстве групп мохообразных не является главным. Об этом мы уже
упоминали.
Для того чтобы возник спорофит,
необходимо оплодотворение. Для большей части мохообразных это почти
непреодолимая трудность. Дело в том, что для этого необходима вода, ибо только
в ней и способны передвигаться антерозоиды. Но ведь мохообразные - растения
сухопутные. И если архегонии и антеридии не находятся рядом, а разделены
пространством хотя бы в несколько сантиметров (а это бывает сплошь и рядом у
двудомных и однодомных видов, но с раздельным расположением архегониев и
антеридиев на разных участках побега), дождевой воды может и не хватать для
того, чтобы осуществилось оплодотворение. А ведь у двудомных видов архегонии и
антеридии разделены, как правило, расстоянием не в несколько сантиметров, а в
десятки, сотни и даже тысячи (нередко многие тысячи) метров. Так, мужские
растения одной из разновидностей Tortula laevipila известны
только в Европе, а женские - только в Северной Америке! Вот и попробуй добейся
оплодотворения! Нужно ли говорить, что спорогоны у этой разновидности не
найдены и, по всей вероятности, отсутствуют в природе.
К вегетативному размножению способен
не только гаметофит, но и спорофит. В эксперименте удавалось из кусочков ножки
спорогона у целого ряда видов листостебельных мхов получать сначала протонему,
а из нее и развитое листостебельное растение. Этот способ размножения
называется апоспорией. Надо сказать, что некоторые виды отличаются баснословной
способностью к апоспории: у одного мха, например, удалось прорастить 113
кусочков ножки спорогона из 133! Полученное таким способом листостебельное
растение - гаметофит - имеет двойной по отношению к исходному, родительскому
гаметофиту набор хромосом. Ведь возник-то он из спорофита. Насколько широко
распространена апоспория в природе, пока не совсем ясно. Но можно предположить,
что она является одним из путей возникновения полиплоидии у мохообразных, во всяком
случае у представителей класса листостебельных мхов. У печеночников спорофит
очень нежный и быстро отмирающий. Апоспория если и имеет место, то, несомненно,
в гораздо меньших масштабах. Как знать, может быть, именно с этим и связано
слабое развитие полиплоидии в классе печеночников.
Вегетативное размножение
осуществляется в основном не случайно отламывающимися кусочками гаметофита или
спорофита, а с помощью специально устроенных органов, которые возникают на
гаметофите (ни спорофите они не известны). Какое-то время эти органы могут
существовать и будучи отделенными от материнского растения, в состоянии покоя,
и в таком виде способны транспортироваться на значительные расстояния ветром
или водой. Словом, ведут себя как споры или семена. При попадании в благоприятные
условия они прорастают и дают сначала протонему (в отличие от «обыкновенной»
протонемы, возникающей из споры, эта протонема называется вторичной), а потом
листостебельный побег или слоевище. Реже они прорастают непосредственно во
взрослое растение, не образуя предварительно протонемы. Что же это за органы?
Один из наиболее простых случаев -
легко опадающие выводковые листья.
Выше мы охарактеризовали
мохообразные как растения вечнозеленые. Это так и есть. Но у некоторых видов
листья частично опадают в течение всего вегетационного периода, и не под
влиянием неблагоприятных условий, а по достижении определенной степени
зрелости, как опадают созревшие семена. В простейших случаях опадающие листья
решительно ничем от неопадающих, прочносидящих не отличаются. Так, например,
происходит у широко распространенного вида Dicranodontium
denudatum.
Очень часто, однако, заметны существенные отличия от них и в форме и в
размерах. Иногда такие специализированные листья собраны на особых веточках или
группируются в определенных местах, чаще на верхушке стебля. Примером может
служить Brothera
leana.
Иногда опадает не весь лист, а
только его часть. Лист в таких случаях распадается на несколько кусков. В
отдельных случаях вегетативному размножению служит уже не вообще кусочек листа,
а лишь кусочек жилки, обычно ломкой и в верхней части сильно расширенной.
Именно так действует, например, арктоальпийский мох Barbula
johansenii,
изредка встречающийся в Советском Союзе.
Широко распространены выводковые
тела. Это многоклеточные образования, состоящие из нескольких клеток или
нескольких десятков клеток (рис. 8). Размеры, понятно, микроскопические. По
форме и способу образования эти тела чрезвычайно разнообразны. Они бывают
линейными или удлиненными, округлыми, овальными, грушевидными и так далее.
Могут возникать непосредственно на пластинке листа, на его жилке, в пазухе или
на стебле. Они могут быть одиночными или в виде больших групп концентрироваться
на специальных разветвленных нитях-подставках, приуроченных к определенным участкам.
Одним из вариантов выводковых тел являются выводковые нити, состоящие из
нескольких (обычно не более 7-9) клеток, вытянутых в ряд.

Рис. 8. Различные органы
вегетативного размножения листостебельных мхов.
- выводковые почки, образующиеся в
верхней части стебля; 2 - отпавшая выводковая почка; 3, 4 и 5 - выводковые
тельца различной формы; 6 - ризоиды на верхушке листа.
Вегетативному размножению служат
также специальным образом устроенные мелкооблиственные веточки. В определенный
момент они легко выпадают из гнезда, в котором сидят, разламываются на
несколько частей. Веточки, называемые флагеллами, обычно собраны пучками на
верхушках стеблей.
Есть еще выводковые почки. Они так
же, как и выводковые тела, имеют микроскопические или чаще почти
микроскопические размеры, но представляют собой именно почки, содержащие
зачатки листочков и стебли. Созревшие почки легко опадают (см. рис. 8).
Очень важный и уникальный для высших
растений способ вегетативного размножения у антоцеротовых. Они имеют на
вооружении апланоспоры.
Есть еще один способ вегетативного
размножения - с помощью ризоидов, возникающих на листьях (чаще близ верхушек)
некоторых видов мхов (см. рис. 8). Внешне эти ризоиды ничем от «настоящих»
ризоидов, находящихся в основании стебля или создающих на стебле войлок, не
отличаются. Только возникают в неожиданном месте. Образуются они из особых
клеток верхней части листа, заметно отличающихся от остальных и формой и
размерами. Обламываясь, эти ризоиды, как и остальные органы вегетативного
размножения, дают начало новому растению.
Мы вкратце рассмотрели не все,
конечно, но многие виды и способы вегетативного размножения. Для ряда видов
вегетативное размножение столь постоянно, что это даже находит отражение в
определительных таблицах. Немалое число видов настолько «увлекается»
вегетативным размножением, что не расстается с ним, даже если создаются
благоприятные условия для полового размножения. И тогда на таком растении можно
видеть сразу и спорогоны, и какие-либо органы вегетативного размножения.
Глава 3.
Практическая часть
В ходе бриологической экспедиции на
остров Сахалин летом 2014 года нами были отмечены новые местонахождения редких
видов листостебельных мхов, включенных в Красные книги России (2008) и
Сахалинской области (2005). Это Bryoxiphium savatieri (Husn.)
Mitt., Tetrodontium repandum (Funck in Sturm) Schwaegr., Dicranum nipponense
Besch.
Bryoxiphium savatieri найден на
северо-западном побережье о. Сахалин, на вертикальной поверхности и в
расщелинах большого валуна в елово-пихтовом лесу на крутом северо-западном
склоне г. Вагис в среднем течении р. Уанги, (h = 323 м над
ур. м., N
52°21´42.5", E 142°05´49.9", 17.07.2014, собр.
Тубанова Д.Я., Тумурова О.Д., # S14010).
В Красную книгу РФ (2008) этот вид
включен в категорию 3д и имеет статус «редкого вида с ограниченным ареалом,
часть которого находится на территории России».
На территории России вид
распространен на Дальнем Востоке: в Камчатском крае (8), в Еврейском АО (1), в
Приморском крае (4). В Сахалинской области имеется по одному местонахождению
вида на Курильских островах: Онекотан, Парамушир, Шумшу, Итуруп, Кунашир,
Шикотан; и на острове Сахалин известно два местонахождения вида: между рр.
Агнево и Найнай (у р. Большая Ахзнгы и р. Малу). Вне России - Япония, Корея,
Китай, Филиппины (Красная книга РФ, 2008).
Ранее B. savatieri был внесен в
Красные книги СССР (1984) и РСФСР (1988). Вид включен в Красные книги
Сахалинской области (2005), Еврейского АО (2006) и Приморского края (2008).
Tetrodontium repandum обнаружен
нами на северо-западном побережье о. Сахалин, на почве в основании большого
валуна в елово-пихтовом лесу на крутом северо-западном склоне г. Вагис в
среднем течении р. Уанги, (h = 323 м над ур. м., N
52°21´42.5", E 142°05´49.9", 17.07.2014, собр. Тубанова
Д.Я., Тумурова О.Д., # S14010).
T. repandum в Красной
книге РФ (2008) присвоена категория 3б и статус «редкий вид, имеющий
дизъюнктивный ареал и низкую численность популяций на всем его протяжении».
На территории России вид известен из
двух отдаленных территорий: две точки в Мурманской области, а также 4
местонахождения в Хабаровском крае. В Красной книге РФ (2008) было пропущено
еще одно местонахождение в России - это дальневосточное местонахождение вида на
вулкане Тятя (о. Кунашир, Сахалинской области) на старом лавовом массиве на
поверхностях скал (Сherdantseva, Ignatov, Ignatova, 2006). Вне
России - Европа, Япония, Северная Америка, Грузия (Красная книга РФ, 2008).
Помимо Красной книги РФ, вид включен
в Красную книгу Мурманской области (2003) со статусом «Бионадзор».
Таким образом, T. repandum необходимо
включить в новое издание Красной книги Сахалинской области.
Новое местонахождение Dicranum nipponense - это
восточное побережье о. Сахалин, устье р. Анна, крутой склон южной экспозиции ~
40˚ с выходами горных пород, покрытых мхом, на правом берегу реки, на
скалах с мелкоземом в ельнике аянском с березой (h = 160 м над
ур. м., N
47°09´27.9", E 143°00´57.9", 24.07.2014, собр.
Тубанова Д.Я., Доржиева Л.Х., # S14022).
Dicranum nipponense входит в
список краснокнижных видов Сахалинской области (2005) под статусом V(2),
уязвимый вид. Этот вид имеет распространение в России только на Дальнем
Востоке: в Приморском крае (в заповедниках Уссурийский, Лазовский и «Кедровая
Падь») и в Сахалинской области (в окр. г. Южно-Сахалинск, в Анивском районе на
о. Сахалине и на о. Кунашире) (Отнюкова, 2001; Cherdantseva et al., 2006).
Вид встречается в Японии, Китае, Корее и на о. Тайвань.
Заключение
В ходе бриологической экспедиции
были найдены новые местонахождения трех краснокнижных мхов, ранее не указанных
бриологами для Сахалина. Это Bryoxiphium savatieri (Husn.)
Mitt., Tetrodontium repandum (Funck in Sturm) Schwaegr., Dicranum nipponense
Besch.
Список
использованной литературы
1. Бардунов
Л.В. Древнейшие на суше.-Новосибирск: Наука, 1984
. Красная
книга Еврейской автономной области. Редкие и находящиеся под угрозой
исчезновения виды растений и грибов/ Отв. ред. Т. А. Рубцова; науч. ред. С. Д.
Шлотгауэр.- Новосибирск: АРТА, 2006.- 247 с.: ил.
. Красная
книга Мурманской области / Правительство Мурм. обл., Упр. природ, ресурсов и
охраны окружающей среды МПР России по Мурм. обл.; [Андреева В. Н. и др.;
Худож.: А. М. Макаров]. - Мурманск: Кн. изд-во, 2003. - 400 с: ил.
. Красная
книга Приморского края: Растения. Редкие и находящиеся под угрозой исчезновения
виды растений и грибов. Владивосток: АВК «Апельсин», 2008.- 688 с. Ил.
. Красная
книга Российской Федерации (растения и грибы).- М.: КМК, 2008.- 855 с.
. Красная
книга РСФСР. Растения.- М.: Изд-во, 1988.- 591 с.
. Красная
книга Сахалинской области: Растения/ Отв. редактор проф. д.б.н. В.М. Еремин.-
Южно-Сахалинск: Сахалинское кн. изд-во, 2005.- 348 с.
. Красная
книга СССР.- М.,1984.- Т. 2.- 480 с.
. Сherdantseva
V.Ya., Gorobets K.V., Harpel J., Ignatov M.S., Ignatova E.A., Teleganova V.V.
New moss records from Sakhalinskaya Province. 1. Sakhalin // Arctoa (2006) 15:
249-271
10. Сherdantseva
V.Ya., Ignatov M.S., Ignatova E.A. New moss records from Sakhalin Province. 2.
Kunashir. // Arctoa (2006) 15: 249-271
11. Otnyukova
T. N. Notes on Dicranum (Dicranaceae, Musci) in Russia.1. Dicranum nipponense
found in Far East // Arctoa (2001) 10: 157-160
12. Беликович
А.В., Галанин А.В., Афонина О.М., Макарова И.И. Публикуется по тексту:
Беликович А.В., Галанин А.В., Афонина О.М., Макарова И.И. Растительный мир
особо охраняемых территорий Чукотки . Владивосток: БСИ ДВО РАН, 2006. 260 с.
. http://www.sakhalin.ru/