Изучение видового состава и структурных особенностей сообществ долгоносиков-хортобионтов города Гомеля
ВВЕДЕНИЕ
Предполагается, что город является не только
местом обитания человеческой популяции, но и предоставляет условия для
существования различных видов животных, растений, грибов, простейших,
прокариот, являющихся неотъемлемыми элементами среды обитания горожанина [7].
Постоянное градостроительство, развитие
промышленности и изменение человеком характерных особенностей биоценозов ведет
к формированию других специфических экосистем. В связи с этим, появляются
различные экологические ниши, изменяются видовое разнообразие, численность,
распределение и динамика беспозвоночных животных.
Город - нестабильная экосистема. В отличие от
природных экосистем
направление развития города определяется
деятельностью человека [3]. Ода из главных особенностей городской экосистемы в
том, что в ней встречаются как условия, характерные для естественных
местообитаний, так и особенные, к которым относятся: более теплый и сухой
микроклимат, обширная кормовая база, а также немалое количество «новых»
свободных экологических ниш для заселения [1].
Городская среда обитания сильно трансформирована
в сравнении с естественными биоценозами и представлена измененными природными
ландшафтами. Примыкающие к городской границе пустыри и поля превращаются в
окультуренные газоны, окраинные леса становятся парками и т.д. В городских
поселениях преобразование ландшафтов достигает своей максимальной величины, что
не может не сказывать на составе и структуре сообществ различных организмов
[6].
Естественные биоценозы Беларуси и Европы изучены
в большей степени, чем фауна городов и прочих мест компактного проживания
человека. Фауна жесткокрылых города Гомеля, который является вторым по величине
городом Беларуси, изучена недостаточно. В связи с недостатком сведений по
видовой структуре сообществ городских экосистем затруднена работа по оценке
степени трансформирования урбоценозов человеком [2].
Долгоносики, или слоники (Curculionidae),- одно
из самых многочисленных по количеству видов семейств жесткокрылых. Они являются
важными компонентами трофических и топических взаимодействий, а также
вредителями сельского и лесного хозяйства [3].
Часто долгоносики своевременно не опознаются и
вызываемые ими повреждения не всегда фиксируются. Поэтому изучение их весьма
важно как с научной, так и с практической точки зрения [4].
Цель исследования - изучение видового состава и
структурных особенностей сообществ долгоносиков-хортобионтов города Гомеля.
Задачи исследования:
Выявление и уточнение видового состава
долгоносиков-хортобионтов, обитающих на участках с разной степенью
антропогенной нагрузки.
Выявить особенности сезонной динамики активности
долгоносиков-хортобионтов.
Составление электронной базы данных.
Анализ таксономической структуры долгоносиков
исследованных территорий.
Анализ экологических особенностей видовой структуры.
Объектом наших исследований являлись
долгоносики-хортобионты, обитающие на травянистой растительности.
Предметом исследований являлись: видовой состав
и структура сообществ долгоносиков-хортобионтов в условиях различной степени
рекреационной нагрузки на исследуемых участках.
Гипотезой в наших исследованиях выступало
положение, что видовой состав и структура сообществ долгоносиков - хортобионтов
неоднородны и зависят от степени антропогенного воздействия на их условия
существования.
Практическая и экономическая значимость работы
. Составленный аннотированный список может быть
использован для дальнейших исследований.
. Полученные данные о видовом составе и динамике
сезонной активности потенциальных вредителей помогут более эффективно выбирать
и использовать методы ограничения их численности.
видовой долгоносик хортобионт
антропогенный
1 .ОБЗОР ЛИТЕРАТУРЫ
.1 Исторический очерк
Наука о насекомых - энтомология (от греческих
слов entomon - насекомое, loqos - наука) - представляет обширную область знания,
тесно связанную со многими сторонами жизни человеческого общества [5].
Интерес к насекомым зародился в глубокой
древности. Человек издавна сталкивался с вредом, наносимым насекомыми, и
использовал полезных насекомых. В ассирийских клинописных табличках и
египетских папирусах 3-го тыс. до н. э. упоминаются опустошительные налеты
саранчи, в древнекитайских рукописях этого же периода имеются указания о
разведении тутового шелкопряда и борьбе с насекомыми - вредителями огородов.
Труды древнегреческого философа Аристотеля (IV в. до н. э.), выделившего среди
«животных без крови» группу «энтома», содержат сводку данных о насекомых.
Однако лишь в XVII в. возникла наука
энтомология. Основы ее заложили труды голландского ученого Я. Сваммердама по
анатомии и развитию пчелы (1669), итальянских ученых М. Мальпиги по анатомии и
развитию шелкопряда (1686), Ф. Буонанни по строению ротовых аппаратов
насекомых, нем. ученого И. Гедарта по типам метаморфоза. В XVII в. были сделаны
первые попытки создать систему насекомых английским ученым Дж. Реем. Выдающийся
шведский естествоиспытатель Карл Линней (1707-1778) в своей «Системе природы»
отвел должное место и насекомым. Предложенные им названия наиболее крупных
отрядов сохранились до настоящего времени. Со времени Линнея начался быстрый
рост систематики, стали появляться работы по фауне и биологии насекомых.
В XVIII в. появляются многотомные сводки знаний
о насекомых: «Мемуары по истории насекомых» (т. 1-6, 1734-1742) французского
ученого Р. Реомюра, открывшего партеногенез у тлей (1737).
В XIX в. свои системы насекомых предложили
английские энтомологи У. Керби, Дж. Уэствуд и Дж. Леббок, выделившие еще ряд
отрядов. Французский энтомолог П. Латрейль (1831) предложил систему в объеме
класса, примерно соответствующую современной. Чарльз Дарвин (1809-1882),
работая над «Происхождением видов», многие положения своего бессмертного труда
иллюстрирует примерами из области энтомологии. С его появлением системы
насекомых стали строиться на филогенетической основе [25].
Развитие энтомологии в XX в. характеризуется
накоплением огромного фактического материала по мировой энтомофауне , широким
размахом работ по систематике, важными открытиями и исследованиями во всех
областях теоретической и прикладной энтомологии. Число известных науке видов насекомых
возросло до 1 млн. (предположительно существуют 2-3 млн. видов), каждый год
открывают сотни новых видов, перестраиваются и совершенствуются системы
насекомых. Выделено около 35-40 отрядов. Традиционные методы систематики
(сравнительно-морфологические и др.) обогащаются более тонкими методами
исследования (электронная микроскопия, кариосистематика) с использованием ЭВМ.
Насекомые стали важным объектом научных
экспериментов (например, дрозофила в генетике, малый мучной хрущак в
популяционной генетике и т. п.) [28].
.2 Становление энтомологии в России и за рубежом
Первые сообщения о повреждениях леса и
деятельности лесничих в области борьбы с вредителями появляются еще в начале
XIX столетия. Большинство этих сообщений было помещено в «Лесном журнале»,
издававшемся с 1837 г. В 1845-1851 гг. вышло двухтомное руководство «О вредных
насекомых», изданное Ученым Комитетом Министерства Государственных Имуществ.
Эта книга была первой сводкой по вредителям леса, если не считать кратких
сведений, сообщаемых в курсе «Лесоохранение, или правила сбережения растущих
лесов», читавшемся в Санкт-Петербургском практическом лесном институте почти с
момента его основания (1811) [12].
Решающую роль в развитии теоретических основ
лесной энтомологии и ее пропаганды сыграла деятельность выдающегося зоолога Н.
А. Холодковского, начавшего в 1885 г. чтение энтомологии в Петербургском лесном
институте [13].
Русская лесоэнтомологическая школа внесла
большой вклад в дело борьбы с вредителями леса. Работами И.Я.Шевырева и
К.Э.Линдемана определилось экологическое направление исследований, тесно
связанное с практикой лесного хозяйства [10].
В.И.Гусевым и М.Н. Римским-Корсаковым был издан
«Определитель повреждений лесных и декоративных деревьев и кустарников
Европейской части СССР», вышедший последним изданием в 1951 г.
Крупным центром в области лесной энтомологии
является в настоящее время Институт леса и древесины Сибирского отделения РАН.
Здесь ведутся широкие биоценологические исследования лесной фауны в связи с
химическими обработками лесов, изучается динамика численности стволовых
вредителей, сибирского шелкопряда и других вредителей тайги [21].
Быстрое развитие прикладной
энтомологии в России вытекало из нужд сельского и лесного хозяйства. К этому
времени насекомые стали наносить очень большой вред зерновым и техническим
культурам, плодоводству и лесному хозяйству. Особенно сильно ощущался вред,
причиняемый насекомыми на юге страны, где приносили страшные опустошения
саранча, хлебный жук, свекловичный долгоносик, непарный шелкопряд <#"868581.files/image001.gif">
Рисунок 1 - Долгоносик
малинно-земляничный (Anthonomus rubi)

Рисунок 2 - Chlorophanus sellatus

Рисунок 3 - Mecaspis atternans

Рисунок 4 - Плодожир каштановый
(Curculio elephas)

Рисунок 5 - Phyllobius pyri
Внешнее строение долгоносиков
рассмотрим на примере Tychius karkaralensis.
Головотрубка в 1.18 - 1.34 раза
короче переднеспинки, более или менее равномерно сужена от основания к вершине
или со слабовыемчатыми в основной части (до основания усиков) боками, очень
слабо изогнута, в вершинной части умеренно уплощена. Спинка головотрубки
матовая, в густых мелких точках; вершинная часть слабо блестящая, с рядами
удлинённых точек вдоль краёв и в очень мелких разреженных точках посередине.
Усики прикреплены на расстоянии 0.37 - 0.47 длины головотрубки от её вершины,
довольно тонкие, средней длины. Членики жгутика слабо или умеренно поперечные,
начиная с 3-го или 4-го, иногда 4-й членик длиннее соседних. Булава
удлинённо-яйцевидная. Лоб плоский. Глаза умеренно выпуклые.
Ширина переднеспинки в 1 - 1.24 раза
больше её длины. Бока округлены довольно равномерно, вершина с глубокой
перетяжкой, диск равномерно умеренно выпуклый, в густых небольших точках. Щиток
треугольный.
Надкрылья в 1.1 - 1.33 раза шире
переднеспинки, длина их в 1.55 - 1.63 раза больше ширины. В основной половине
надкрылья почти параллельносторонние, в вершинной - равномерно сужены, на
вершине довольно широко округлены. Диск заметно приподнят вдоль середины, в
средней части не выпуклый в продольном направлении, в основании скошен иногда
довольно круто, вершинный скат пологий, начинается почти от середины диска.
Бороздки отчётливые, узкие, но довольно глубокие. Промежутки в густых довольно
крупных точках, нечётные иногда слабовыпуклые.
Ноги средней длины, бёдра умеренно
утолщены в вершинной части. Зубец на задних бёдрах хорошо развит, острый; на
средних - маленький, в виде зёрнышка, но обычно заметен, а на передних бёдрах
если и выражен, то скрыт приподнятыми чешуйками и незаметен. Проксимальнее
середины передние бёдра несут ещё один небольшой, но обычно отчётливый зубец в
виде складки с округлым верхним краем. Все голени с небольшим изогнутым мукро.
Передние голени с крупным острым зубцом на внутреннем крае немного проксимальнее
середины, средние и задние - умеренно двувыемчатые. Лапки короткие и довольно
узкие, 3-й членик в передних в 1.4 раза шире 2-го, коготковый довольно широкий,
более чем на 2/3 выдаётся за лопасти 3-го членика. Коготки крупные, с крупным
зубцом в основных 2/3. Эдеагус в основной половине слабо, в вершинной -
изогнут, почти параллельносторонний, у вершины коротко сужен и затем почти
прямо срезан, ширина ровного вершинного участка составляет треть ширины
эдеагуса в средней части его длины. Вентральная сторона у вершинного края несёт
с каждой стороны около 10 тонких и длинных волосков, направленных вперёд и
загнутых вершинами к средней линии; ещё по группе волосков, перпендикулярных
поверхности эдеагуса, расположено вдоль краёв десклеротизованного участка
вентральной стороны немного отступя от вершины. Со стороны слабопоперечного
семяизвергательного отверстия вершина несёт 2 коротких волоска, расходящихся
вершинами наружу[30].
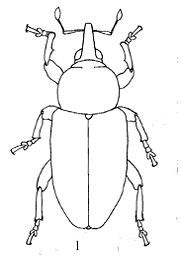
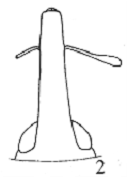

Рисунок 6 - Tychius karkaralensis.
- общий вид самца, 2 - головотрубка,
3 - вершина эдеагуса сверху.
.1.2 Жизненный цикл
Жизненный цикл табачного
долгоносика, как и всех жуков, состоит из четырех стадий - личинка, гусеница, куколка
и взрослое насекомое - и длится от 10 до 12 недель. Поселиться в табаке он
может и на складе плантации, и на табачной фабрике, и в сигарном магазине. В
скрученную сигару отложить личинки из-за плотного покровного листа ему довольно
сложно, но возможно. Если нет других вариантов, он размещает их на беззащитных
срезах сигар. Однако все может начаться еще в сушильных сараях. Табачный
долгоносик терпеливо выжидает, когда закончится период сушки табака, потому что
даже небольшая влажность листа лишает его существование всякой радости. Он
словно специально подгадывает время, когда снимут весь урожай, расплатятся с
работниками и заключат контракты. После чего коварная тварь разворачивает свою
вредительскую деятельность. Ежедневно каждая самка Lasioderma, оказавшаяся в
нужное время в нужном месте, высаживает на сигарное сырье десант, состоящий из
сотни личинок, не видных невооруженным глазом (размер каждой 0.5 мм). О
дальнейшей судьбе «деток» мамаша может теперь не беспокоиться. Куда бы они ни
были заброшены по воле торговца табаком, сытое будущее им обеспечено, а значит,
свое предназначение - плодиться и размножаться - они выполнят[16].
Первая метаморфоза - превращение
личинок в маленьких гусеничек - происходит через 5 - 10 дней, в зависимости от
температуры и влажности. Это самая продолжительная по времени и самая опасная
стадия развития насекомого. Гусеницы начинают активно прогрызать сложные
лабиринты в сигарах или уничтожать еще не обработанные листья. Белые гусеницы
выглядят достаточно симпатично: миниатюрные (длиной 4 мм), с желтой полосочкой
на голове и небольшой каштановой кисточкой на хвостике.
Благоприятная температура для
дальнейшего развития табачного долгоносика +24°C и выше. При температуре +15 -
16°С гусеница не погибает, а словно впадает в летаргический сон до тех пор,
пока за пределами ее «домика» не установится подходящая погода. Спустя 5 - 10
недель после того, как только это произойдет, вокруг гусеницы начинают
образовываться тонкие стенки кокона, и постепенно она превращается в куколку. Одним
табачным жукам необходимо 10 дней, другим - в два раза больше, чтобы приступить
к догрызанию листьев табака и выбраться наружу. Есть и такие, которые могут
ждать удобного момента несколько лет.
Самки рисового долгоносика после
оплодотворения самцами выгрызают в зерне небольшое углубление и откладывают в
него яйцо, сверху запечатывая быстро затвердевающим секретом [15].
Вылупившаяся через несколько дней
личинка обычно прогрызает ход к центру зерновки, затем прогрызает ходы вдоль
бороздки. Поэтому внутри одной зерновки могут развиваться две личинки при
условии, что их ходы будут на противоположных сторонах бороздки. Личинки дл.
2.5 - 3.0 мм, желтовато-белые, безногие. При своём развитии личинка выедает
около половины зерна. Перелиняв четыре раза, личинка превращается в
куколку[10]. Через несколько дней из куколки появляется жук, который, окрепнув,
прогрызает оболочку зерна и выходит наружу. Внутри полупустого зерна остаются
личиночные шкурки и экскременты насекомого. Всё развитие жука от яйца до имаго
занимает при благоприятной температуре 1 - 1.5 месяца. Продолжительность жизни
жуков составляет 3 - 6 месяцев.
Оптимальная температура для жизни
рисового долгоносика лежит в пределах от 27 до 31ºС,
оптимальная относительная влажность воздуха около 70%. При таких условиях самка
может отложить за сутки до 10 яиц. Средняя плодовитость за всю жизнь составляет
около 380 яиц (максимальная плодовитость 570 яиц от одной самки). Требователен
к теплу (оптимальная для развития температура воздуха 27 - 29 °С, критическая -
О (C ), влажности воздуха (оптимальная - 80 - 90%, критическая - 50%), к
влажности зерна (оптимальная - свыше 17%, критическая - ниже 10%). В тёплых
помещениях рисовый долгоносик может иметь от 3-х до 8-ми поколений в год [16].
.2 Распространение
Долгоносики широко распространены по
всему земному шару. Географическое положение и природа Беларуси обеспечивают
разнообразие экологических условий для обитания большого числа видов
долгоносиков, приуроченных как к лесной, так и к степной и лесостепной зонам [19].
.3 Пищевая специализация
По характеру питания почти все
долгоносики относятся к фитофагам и лишь немногие - к фитосапрофагам. В своём
развитии они тесно связаны с травянистыми и древесно-кустарниковыми растениями.
Личинки большинства видов долгоносиков питаются в растительных тканях. Только
немногие кормятся снаружи на листьях, почках и цветках. У значительного числа
видов развитие проходит в почве на корнях растений. Некоторые питаются опадом,
немногие- гниющей древесиной [8].
.4 Практическое значение
В полевых и лесных биоценозах
Белоруссии удельный вес долгоносиков среди других групп жесткокрылых значителен
и роль отдельных видов велика. Их практическое значение определяется в основном
особенностями питания [17]. Долгоносики являются вредителями сельского и
лесного хозяйства. Они повреждают также растительные продукты и техническую
древесину на складах. В условиях нашей страны как вредители бобовых культур
немаловажное значение имеют клубеньковые долгоносики (Sitona), клеверные
семяеды (Apion), фитономусы (Phytonomus) и т. д. ощутимый вред в садах приносят
яблоневый долгоносик- цветоед (Anthonomus pomorum L.), малинно- земляничный
цветоед (A. rubi Hbst.) (рис. 1) и другие. В лесах постоянно вредят большой
сосновый долгоносик (Hylobius abietis L.), желудёвый долгоносик (Curculio
glandium Marsh.) и другие. Особенно сильно повреждают долгоносики растения в
питомниках и культурах [9].
.5 Место и методы проведения
исследования
Исследования проводились стационарно
с мая по август 2010 года на трёх участках, которые находились в районе
микрорайона «Солнечный». Стационары представляли собой:
«Опушка леса». Стационар наименее
подверженный рекреационной нагрузке. Расположен на границе города.
Растительность представлена мятликовым разнотравьем, из травянистых растений
преобладают одуванчик лекарственный - Taraxacum officinale Wigg., клевер
пашенный - Trifolium arvense L., клевер ползучий - T. repens L. Подрост
составляли молодые сосны и берёзы. Почва - супесь тяжёлая.

Рисунок 7 - Стационар «Опушка леса»
«Заливной луг». Стационар покрыт
большим количеством прибрежной растительности. Рекреационная нагрузка
незначительная, преимущественно встречаются рыбаки. Растительность представлена
снытью обыкновенной - Aegophodium padagrara L., лютиком жгучим - Ranunculus
flammula L. и лапчаткой серебристой - Potentila argenthea L. Почва - суглинок
лёгкий.

Рисунок 8 - Стационар «Заливной луг»
«Суходольный луг». Характеризуется
значительной рекреационной нагрузкой, т.к. расположен недалеко от проезжей
части. Из растительность преобладают мятликовые. Почва - супесь легкая.

Рисунок 9 - Стационар «Суходольный
луг»
Сбор материала проводился при помощи
кошения энтомологическим сачком.

Рисунок 10 - кошение сачком
Энтомологический сачок (рис. 11)
состоит из трёх частей: обруч, бязь и рукоятка. Обруч изготавливается из
стальной проволоки диаметром 4 мм. В качестве рукоятки сачка была использована
деревянная палка длиной 1,5 м. [15].

Рисунок 11 - Сачок
Ручной сбор мелких долгоносиков
производился эксгаустером. Эксгаустер (рисунок 12) состоял из полистироловой
баночки (объём около 200 мл), резиновой пробки и двух трубок (длина 30-40 см,
диаметр 4-5 мм). Верх банки обрезан, туда вставлялась пробка. В пробке сделаны
соответственно два отверстия под трубки, одна из которых пластичная. На конец
этой трубки, который находится внутри баночки, был одет фильтр, для того, чтобы
в ротовую полость не попадали инородные тела, т.к. через противоположный конец
производилось засасывание воздуха из банки. Вторая трубка направлялась на
особей, находившихся в сачке. Таким образом, насекомые с током воздуха по
трубочке, как по коридору, попадали в баночку.
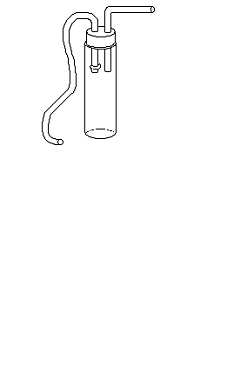
Рисунок 12 - Эксгаустер
Собранные эксгаустером насекомые
помещались в морилку (рисунок 13) для умерщвления. Морилка представляла собой
специальное устройство для умерщвления насекомых. Современные морилки
изготавливаются из стеклянных или пластиковых стойких к растворителю
широкогорлых банок с плотной крышкой. Объём произвольный и зависит от размера и
количества помещаемых в морилку насекомых. На дне располагается либо ватный
тампон, зашитый в ткань, либо кусок поролона таким образом, чтобы он не
вываливался при переворачивании банки [15].
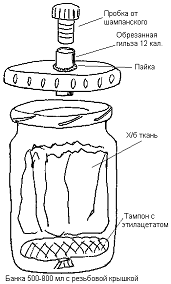
Рисунок 13 - Морилка
Заполнить объём морилки куском белой
х/б или льняной ткани, который прекрасно впитывает избыток влаги, препятствует
повреждению насекомых и не нуждается в частой замене. Морилка заправляется
ядом. С прошлого века пользуются обычным медицинским эфиром или хлороформом.
Насекомые после замаривания этими ядами получаются жёсткими и почти не поддаются
расправлению и монтировке для коллекции. Лучше всего использовать эфиры
уксусной кислоты - этиловый, амиловый, бутиловый. В крышке морилки проделыватся
отверстие, в котором закрепляется отрезок тонкостенной трубки (удобна охотничья
гильза), закрытый пробкой. Через это отверстие в банку помещаются насекомые
[13].
Всего за период исследований изучено
3 стационара. В результате на первом стационаре (Опушка леса) было собрано 120
экземпляров долгоносиков- хортобионтов; на втором (Заливной луг) - 82
экземпляра; на третьем (Суходольный луг) - 87 экземпляров. Всего было собрано
289 насекомых.
В лаборатории умерщвленные насекомые
выкладывались на ватные слои для последующего хранения и определения.
Определение проводилось при помощи общепринятых определительных таблиц [19 -
21].
Структуру сообществ определяли при
помощи ряда индексов разнообразия: индекс шеннона, Симпсона, выравненность по
Пиелу. Статистическая обработка полученных данных производилась с
использованием программы EXEL на персональном компьютере [22].
Для определения доминирования в
сообществе использовали шкалу O. Renkonnen, согласно которой виды, составляющие
более 5% от общего числа особей считаются доминантными; 2 - 5% -
субдоминантными; 1-2% - рецедентными; менее 1% - субрецендентными [23].
3. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ И ИХ
ОБСУЖДЕНИЕ
.1 Аннотированный список видов
долгоносиков фауны города Гомеля
На основе собственных данных был
составлен аннотированный список видов долгоносиков в пределах г. Гомеля,
который включал в себя следующие харатеристики 82 видов: название,
распространенные синонимы, тип ареала, биотопическую приуроченность, зимующую
фазу и срок генерации.
СЕМЕЙСТВО APIONIDAE SCHÖNHERR,
1823
Подсемейство Apioninae
Schöncherr,1823
Род Apion Herbst, 1797
. A. corniculatum (Germar,1817)
Евро-сибиро-центральноазиатский вид. Обычен. Обитают на травянистой
растительности, в том числе и на дроке.
. A. difficile (Herbst,1797)
Западно-центральнопалеарктический вид. Распространен повсеместно. Обитает на
нивянике обыкновенном и ромашке непахучей. Личинка в корневой шейке и корне
этих растений.
. A. fulvipes (Geoffroy, 1785).
Транспалеарктический полизональный вид. В массе. Обитает на клеверах.
. A. loti (Kirby, 1808).
Транспалеарктический полизональны вид.Обычен. Обитает на лядвенце. Личинки питаются
в бобах семенами.Окукливаются тут же.
. A. pomonae (Fabricius, 1798).
Транспалеарктический полизональный вид. Обычен. Обитает на бобовых (горошки,
вика, чечевица). Личинки питаются незрелыми семенами разных видов горошков, а
также чины. Имаго могут повреждать листья плодовых деревьев и кустарников.
. A. trifolii (Linnaeus, 1768)
Транспалеарктический полизональный вид. Обычный вид. Обитает на клеверах.
Личинка - в цветах. При сильном повреждении соцветия деформируются.
. A. violaceum (Kirby, 1808). Транспалеарктический
полизональный вид. Распространен по всей территории. Имаго на клевере,
лядвенце, щавеле. Личинка - на щавеле.ЕМЕЙСТВО CURCULIONIDAE LATREILLE, 1802
Подсемейство Otiorhynchinae Schöncherr,1826
Триба Otiorhynchini Schöncherr,1826
Род Otiorhynchus Germar, 1824
. O. ovatus (Linnaeus, 1758).
Евро-сибиро-среднеазиатский вид. Обычен и распространен повсеместно. Зимуют
имаго в подстилке (рис 14).

Рисунк 14 - Otiorhynchus ovatus
[24].
. O. raucus (Fabricius, 1777).
Евро-сибиро-среднеазиатский вид. Обычен и широко распространен в лесах, садах и
парках.
. O. tristis (Scopoli, 1763).
Евро-ленский вид. Встречается в лесах, садах, на полях и огородах на
розоцветных, березе, ольхе, иве, осине, грабе, дубе, бересклете. Имаго зимует в
подстилке.
Триба Trachyploeini Lacordaire, 1863
Род Trachyphloeus Germar, 1817
. T. bifoveolatus (Beck, 1817).
Евро-кавказский вид. Обычен и распространен повсеместно. Обитает в лесах,
садах, на лугах и газонах в подстилке на сухих песчаных местах под камнями и на
травянистой растительности. Размножается партеногенетически.
Триба Phyllobiini Schoncherr, 1826
Род Phyllobius Germar, 1824
. Ph. arborator (Herbst, 1797).
Европейский вид. Массовый вид, распространен по всей территории. Предпочитает
мезофитные биотопы, питается листьями деревьев и кустарников.
. Ph. argentatus (Linnaeus, 1758).
Европейский вид. Один из самых распространенных и вредоносных видов в Беларуси.
Дендрофил, населяет гигрофитные биотопы. Кроме лиственных пород могут вредить
сосне и ели.
. Ph. brevis (Gylenhal, 1834).
Транспалеарктический вид. Широко распространённый вид. Часто встречается в
массе, особенно в Полесье. Характерен для возвышенных, открытых и освещённых
мест. Многояден. Питается манжеткой, вредит садовой и дикорастущей землянике.
. Ph. maculatus (Tournier, 1880).
Евро-байкальский вид. Обитает на ивах.
. Ph. maculicornis (Germar, 1824).
Евро-ленский вид. Обычен, дендрофил.
. Ph. oblongus (Linnaeus, 1758).
Трансевразиатский температно-южносибирскаий вид. Один из самых распространенных
видов в Беларуси. Вредитель плодово-ягодных культур - обгрызает почки.
. Ph. pomaceus (Gyllenhal, 1834).
Евробайкальский вид. Обычен и широко распространен. Предпочитает увлажненные
биотопы. Полифаг.
. Ph. pyri (Linnaeus, 1758). Евро-сибиро-центральноазиатский
вид. Обычен, распространен по всей территории. Встречается на деревьях и
кустарниках.
. Ph. scutellaris (Redtenbacher,
1849). Евро-байкальский вид. Очень редок. Встречается на сырых и заливных
лугах.
. Ph. virideaeris (Laicharting,
1781). Западно-центрально-палеаркти-ческий вид. Редок. Имаго вредит листьям
различных деревьев и кустарников, а также травянистых растений. Встречается во
фруктовых садах.
Триба Polydrosini Schoncherr,1823
Род Polydrusus Germar,1817
. P. cervinus (Linnaeus, 1758).
Трансевразиатский бореомонтанный вид. Обитает в лесах, парках, садах на
древесно-кустарниковой раститель-ности. Полифаг. Жуки вредят молодым лиственным
деревьям, объедая кору и почки. Личинка живет как инквилин в ходах клещика
Eriophyes.
. P. pilosus (Gredler, 1866).
Трансевразиатский борео-монтанный вид. Обычен. Повреждает преимущественно
хвойные, но питается также на березе. Буке, грабе, ольхе, лещине, рябине,
боярышнике.
. P. undatus (Fabricius, 1781).
Евро-байкальский вид. Обычен, встречается повсеместно. Обитает в лесах, садах,
на дикой травянистой растительности. Многояден.
Триба Brachyderini Schöncherr,1826
Род Sciaphilus Schöncherr,1823
25. S. asperatus (Bonsdorff, 1785).
Евро-кавказский вид. Обычен, распространен повсеместно. Обитает в тенистых и
влажных местах в полстилке и мху. Питается листьями земляники, малины, ежевики,
тополя. Размножается партеногенетически.
Род Brachysomus Schöncherr,1823
26. B. echinatus (Bonsdorff, 1785).
Трансевразиатский температно-южносибирский вид. Нередок, распространен по всей
территории. Размножается партеногенетически, ведет приземных образ жизни.
Зимуют имаго в подстилке.
Триба Strophosomini Gistel, 1856
Род Strophosoma Billberg, 1820
. S. albolineatus (Seidlitz, 1867).
Западно-палеарктический вид. Обычен на заливных лугах. Долгоносик биологически
связан со щавелями, листьями которого питается.
. S. capitatum (DeGeer, 1775).
Трансевразиатский температный вид. Обычен, широко распространен по всей
территории. Обитает на древесных породах, наносит существенный вред сосновым
питомникам и культурам. Личинка многоядна, развивается в почве на корнях
различных растений, в том числе на вереске.
Триба Tanymecini Lacordaire, 1863
Род Chlorophanus Sahlberg, 1823
. C. viridis (Linnaeus, 1785).
Евро-сибиро-среднеазиатский вид. Обитает в лесах, садах, лугах и полях. Вредит
землянике, и питается как на травянистых, так и на древесных растениях.
Род Tanymecus Germar, 1817
. T. palliatus (Fabricius, 1787).
Евро-казахстанский вид. Обитает на полях, лугах, садах, лесах и парках.
Многояден. Приносит ущерб сельскому и лесному хозяйству, а также садоводству.
Подсемейство Sitoninae Gistel,1856
Род Sitona Germar,1817
. S. hispidulus (Fabricius, 1777).
Трансевразиатский суббореальный вид. Обитает главным образом в полевых
биотопах, но может встречаться и в садах, парках, полезащитных полосах. Вредит
различным клеверам, люцерне и другим бобовым. Может повреждать хвою молодых
сосны и ели.
. S. lepidus (Gyllenhal, 1834).
Транспалеарктический полизональный вид. Отмечается в различных биотопах: леса,
луга, торфяники, обочины дорог, парки, сады, подстилка. Серьезно вредит
клеверам, повреждает также горох, люцерну, люпин и другие бобовые. Личинки
могут развиваться на люцерне, эспарцете, лядвенце, ракитнике, дроке.
. S. lineatus (Linnaeus, 1858).
Транспалеарктический полизональный вид. На лугах, полях, обочинах дорог, в
лесах, садах. Отмечен на свекле. Повреждает всходы различных лиственных и
хвойных деревьев. Личинки питаются на корнях клевера, лядвенце, люцерны и
других бобовых, а также желтой акации.
. S. macularius (Marsham, 1802).
Циркумтемператный вид. Встречается на различных бобовых. Вредит. Иногда может
встречаться на свекле.
. S. puncticollis (Stephens,
1831).Западно-центральнопалеарктический вид. Обитает в основном на клеверах
(особенно красном), редко - на других бобовых. В Минске - на всех стационарах с
луговой растительностью.
Подсемейство Erirhininae Schonherr,
1825
Триба Rhytirrhini Lacordaire, 1863
Род Gronops Schonherr, 1823
. G. sulcatus (Bohemian, 1842).
Южноевропейско-кавказский вид. Редок. Встречается местами. В хорошо
обогреваемых биотопах.
Триба Phytonomini Gistel, 1856
Род Hypera Germar, 1817
. H. meles (Fabricus, 1792).
Западно-центральнопалеарктический вид. Массовый и широко распространенный вид.
Серьезный вредитель клевера. Личинки объедают почки и соцветия.
. H. nigrirostris (Fabricius, 1775).
Западнопалеарктический вид. Обычный и широко распространенный вид в фауне
Беларуси. Обитает на клеверах, которыми и питается.
Триба Erirhinini Schönherr,
1825
Род Dorytomus Germar, 1817
. D. longimanus (Forster, 1771).
Западно-центральнопалеарктический вид. Личинка - на мужских соцветиях тополей,
ивы, ольх, иногда - почках. Имаго повреждают листья.
. D. melanophthalmus (Paykull,
1792). Западно-центральнопалеаркти-ческий вид. Имаго повреждает листья, а
личинка - сережки различных видов.
. D. nebulosus (Gyllenhal, 1836).
Евро-сибиро-центральноазиатский вид. Долгоносик биологически связан с тополями.
. D. salicinus (Gyllenhal, 1829).
Евро-ленский вид. Долгоносик развивается на ивах около воды.
. D. tremulae (Fabricius, 1787).
Евро-байкальский вид. Развивается на осине, пирамидальном тополе и козьей иве.
Род Pseudostyphlus Tournier, 1874
. P. pillumus (Gyllenhal, 1836).
Европейский вид. Нередок. Обитает на ромашке и тысячелистнике.
Подсемейство Cioninae Schönherr,
1825
Род Cionus Clairville et
Schellenberg, 1798
. C. hortulanus (Geoffroy, 1785).
Западно-центральнопалеаркти-ческий вид. Обычен и распространен на всей
территории. Обитает на Scrophularia nodosa, S. aquatica, Verbascum nigrum, V.
phlomoides. Личинка эктофаг питается на листьях и цветках указанных растений.
Подсемейство Tychiinae Thomson, 1859
Триба Tychiini Thomson, 1859
Род Tychius Germar, 1817
. T. aureoles (Kiesenwetter, 1851).
Рыжий люцерновый семяед. На территории Беларуси встречается редко.
. T. crassirostris (Kirsch, 1871).
Западнопалеарктический вид. Донниковый листовой галловый долгоносик.
Распространение: очень редок. Встречается на юге Полесья. Личинка долгоносика
живёт в галлах на листьях донника и люцерны.
. T. haematopus (Gyllenhal, 1836).
Транспалеарктический вид. Золотистый донниковый семяед. Обычный и повсюду
распространённый вид нашей фауны. Долгоносик живёт на доннике и люцерне.
Личинка развивается в бобах донника. Жук повреждает венчики, пестики и
тычиночные трубки, вызывая опадения бутонов.
. T. meliloti (Stephens, 1831).
Трансевразиатский вид. Серый донниковый семяед. Попадается не часто. Долгоносик
биологически связан с донниками. Личинка развивается в галлах на листьях.
. T. picirostris (Fabricius, 1787). Циркумтемператный
вид. Обитает на клевере и прочих бобовых. Вредитель бобовых культур. Личинки
развиваются в плодах.
. T. quinquepunctatus (Linnaeus,
1758). Трансевразиатский вид. Пятиточечный долгоносик. Обычен. Распространён
повсюду. Долгоносик является вредителем гороха, вики, фасоли, чечевицы, чины,
клевера, люцерны и других бобовых. Личинки питаются семенами в бобах.
Триба Ellescini Thomson, 1859
Род Ellescus Dejean, 1821
. E. scanicus (Paykull, 1792).
Западнопалеарктический вид. Обитает на иве, осине, тополе. Личинки развиваются
в соцветиях, имаго - на мужских цветках.
Подсемейство Acalyptinae Thomson,
1859
Род Acalyptus Schonherr, 1833
. A. carpini (Fabricius, 1792).
Трансевразиатский температно-южно-сибирский вид. Редок. Развивается на
узколистных ивах.
Подсемейство Curculioninae
Latreille, 1802
Триба Anthonomini Thomson, 1859
Род Anthonomus Germar, 1817
. A. phyllocola (Herbst, 1795).
Евро-каваказский вид. Вредит. Питается хвойными, в частности сосной, может
встречаться на боярышнике и в почках ели.
. A. pomorum (Linnaeus, 1758).
Транспалеарктический полизонально-южносибирский вид. Обитает на косточковых.
Личинка развивается в косточках черемухи, вишни, черешни, сливы, персика. Имаго
питается цветами и листьями. Сильно вредит.
. A. rubi (Herbst, 1795).
Западно-центральнопалеарктический вид. Обитает на розоцветных. Является
вредителем малины и земляники. Весной имаго питается пыльцой сложноцветных.
Род Furcipes Desbrochers des Loges,
1868
. F. rectirostris (Linnaeus, 1758).
Транспалеарктический полизональ-но-южносибирский вид. Обычен. Вредит
косточковым. Личинка развивается в косточках черемухи, сливы, вишни, черешни,
персика. Имаго питается цветами и листьями.
Триба Curculionini Latreille, 1802
Род Curculio Linnaeus, 1758
. C. crux Fabricius, 1777. Западно-палеарктический
вид. Обитает преимущественно на ивах, произрастающих в сырых и затененных
местах, по берегам рек, в лесах, садах. Личинки развиваются в галлах на листьях
ивы.
Подсемейство Rhynchaeninae Thomson,
1859
Род Rhynchaenius Clairville et Schellenberg,
1798
. Rh. alni (Linnaeus, 1758).
Евро-кавказский вид. Редок. Обитает на ильмовых. Скелетирует листья березы,
ольхи, ильма. Личинка при питании делает широкие мины по краю листьев.
. Rh. populicola Silferberg, 1977.
Евро-сибиро-центральноазиатский вид. Обитает на ивах и тополях. Имаго при
питании скелетирует листья, а личинка проделывает в них мины.
. Rh. salicis (Linnaeus, 1758).
Транспалеарктический полизонально-южносибирский вид. Обитает на ивах, а также
осине и тополе. Личинки делают на листьях широкие мины, похожие на пятна.
. Rh. stigma (Germar, 1821).
Транспалеарктический полизонально-южносибирский вид. Обитает на ивах, березе,
ольхе и осине. Личинка развивается в минах на листьях ив.
Подсемейство Gymnetrinae Thomson,
1859
Триба Gymnetrini Thomson, 1859
Род Gymnetron Schonherr, 1825
. G. melanarium (Germar, 1821).
Западноевропейский вид. Обитает в лесах на Veronica teucrium.
. G. pascuorum (Gyllenhal, 1813).
Западнопалеарктический вид. Широко распространен на влажных лугах, где обитает
на подорожниках. Личинка развивается в семенной коробочке Plantago lanceolata,
в которой питается незрелыми семенами.
Подсемейство Anoplinae Bedel, 1884
Род Anoplus Germar, 1820
. A. plantaris (Naezen, 1794).
Европейский вид. Обитает на березе и ольхе. Имаго повреждает почки и молодые
листья. Личинка минирует листья.
Подсемейство Magdalinae LeConte,
1876(1856)
Род Magdalis Germar, 1817
. M. ruficornis (Linnaeus, 1758).
Евро-байкальский вид. Обычен. Вредит плодовым деревьям и кустарникам, а также
розам. Имаго питается листьями, а личинки - лубом внутри ветвей.
Подсемейство Molytinae Schonherr,
1823
Триба Hylobiini Kirby, 1837
Род Hylobius Germar, 1817
. H. abietis (Linnaeus, 1758).
Трансевразиатский бореальный вид. Обычный и широко распространенный вид нашей
фауны. Обитает в лесах. Известен как опасный вредитель хвойных деревьев,
особенно сосны и ели. Жуки приносят большой вред лесным культурам. Личинки
развиваются в корнях сосновых и еловых пней, иногда на корнях ослабленных
деревьев, а также в кучах сосновой и еловой коры.
Триба Pissodini Gistel, 1856
Род Pissodes Germar, 1817
. P. pini (Linnaeus, 1758).
Евро-казахстанский вид. Заселяет в основном стволы старых сосен, реже елей и
лиственницы, предпочитая при этом больные и ослабленные деревья.
Подсемейство Baridinae
Schoncherr,1839
Род Baris Germar, 1817
. B. artemisiae (Herbst, 1795).
Евро-сибиро-центральноазиатский вид. Обитает на полынях. В наших сборах
присутствовал на стационарах «Степянка-2», «Автозавод» и «Уручье».
Подсемейство Ceutorhynchinae Gistel,
1856
Триба Phytobiini Gistel, 1856
Род Phytobius Dejean, 1835
. P. quadrituberculatus (Fabricius,
1787). Транспалеарктический полизонально-южносибирский вид. Обитает на горце
Polygonum persicaria.
Род Rhinoncus Schoncherr,1826
. Rh. pericarpius (Linnaeus, 1758).
Циркумтемператный вид. Обитает на гречихе и щавелях, является вредителем ревеня
и огородного щавеля. Личинка развивается в галлах на корнях.
Триба Ceutorhynchini Gistel, 1856
Род Tapinotus Schönherr,
1826
72. T. sellatus (Fabricius, 1794).
Евро-сибиро-центральноазиатский вид. Редок. Обитает у воды на вербейнике
(Lysimarhia vulgaris).
Род Coeliodes Schönherr,
1826
73. C. nigritarsus Hartmann, 1895.
Европейский вид. Обитает на березе и иве. Нередок.
Род Ceutorhynchus Germar, 1824
. C. assimilis (Paykull, 1792).
Западно-центральнопалеарктический вид. Обитает в травостое леса, луга и сада,
на полях бобовых. Местами долгоносик сильно вредит культурам озимого рапса и
прочих крестоцветных.
. C. cochleariae (Gyllenhal, 1813).
Европейский вид. Обитает на крестоцветных.
. C. contractus (Marsham, 1802).
Евро-кавказский вид. Долгоносик вредит различным крестоцветным культурам.
Личинка питается в листьях капусты, репы.
. C. erysimi (Fabricius, 1787).
Транспалеарктический полизональный вид. Долгоносик живет по берегам рек на
различных крестоцветных. Личинка развивается в стеблях пастушьей сумки.
. C. punctiger (Sahlberg, 1835).
Европейский вид. Имаго обитает на лугах на одуванчиках, иногда - бодяке
полевом. На личинке паразитирует наездник Bracon maculiger.
. C. rapae (Gyllenhal, 1837).
Западно-центральнопалеарктический вид. Имаго биологически связаны с
крестоцветными. Личинка развивается в корнях. Местами вредит культурным
крестоцветным.
Триба Ceutorhynchini Gistel, 1856
Род Ceutorynchidius Jacquelin du
Val, 1854
. C. rhenanus (Schultze, 1895).
Евро-кавказский вид. Долгоносик развивается в стручках диких крестоцветных.
Род Trichosirocalus Colonelli, 1979
. T. troglodytes (Fabricius, 1787).
Западно-центральнопалеаркти-ческий вид. Обитает на подорожнике, встречается на
сырых местах.
Род Nedynus
Schoenherr, 1825 Cidnorhinus Thomson, 1859
82. N. quadrimaculatus (Linnaeus,
1758). Евроленский вид. Обитает на травянистой растительности, преимущественно
на крапиве, в сырых лесах, на лугах около рек.
По литературным данным на территории
Гомельской области встречается 284 вида долгоносиков, что составляет 65%.
Наиболее богатое видами подсемейство Otiorhynchinae, триба Phyllobiini.
Варьирование различных видов на
территории Гомельской области зависит от различных факторов: пищевая база (так
как большинство долгоносиков питаются различными частями растений),
биотопическая приуроченность (каждому виду характерен свой биотоп), климат и
микроклимат (погодные условия, температура, влажность), сезонная ритмичность
(погодные условия во время размножения), физико-химические факторы в разных
средах. Конечно, все факторы в совокупности оказывают огромное влияние на
распространения и встречаемость видов семейства слоников, но решающим фактором
является пищевая база, так как именно наличие корма влияет на плодовитость. Чем
больше корма, тем больше потомков. Это объясняет такое распределение видов.
.2 Видовой состав и параметры
биоразнообразия хортобионтных долгоносиков исследованных территорий
За время исследований на трех
стационарах нами было отловлено 294 экземпляров долгоносиков травостоя.
Наибольшей численностью представителей семейства Curculionidae характеризовался
стационар «Опушка леса» (таблица). Ниже численность божьих коровок была
отмечена на стационаре «Суходольный луг» (таблица), а наименьшее количество
экземпляров - на стационаре «Заливной луг» (таблица).
Таблица - Видовой состав и структура
доминирования (%) божьих коровок, обитавших на исследованных территориях.
|
Вид
|
Стационары
|
|
Заливной
луг
|
Суходольный
луг
|
Опушка
леса
|
|
1
|
2
|
3
|
4
|
|
Apion
corniculatum Germar,1817
|
0
|
0
|
0,82
|
|
Apion
difficile (Herbst,1797)
|
0
|
2,22
|
1,65
|
|
Apion
fulvipes (Geoffroy,1785)
|
19,28
|
24,44
|
14,88
|
|
Apion
loti (Kirby,1808)
|
0
|
0
|
0,82
|
|
Apion
pomonae (Fabricius,1798)
|
1,20
|
0
|
0
|
|
Apion
trifolii (Linnaeus, 1768)
|
2,40
|
0
|
2,47
|
|
Apion
violaceum (Kirby,1808)
|
1,20
|
0
|
1,65
|
|
Chlorophanus
gibbosus (Paykull,1793)
|
1,20
|
2,22
|
1,65
|
|
Chlorophanus
viridis (Linnaeus,1785)
|
0
|
2,22
|
0,82
|
|
Cionus
tuberculosus (Scopoli,1763)
|
0
|
1,11
|
0
|
|
Hylobius
abietis (Linnaeus,1758)
|
0
|
0
|
0,82
|
|
Hypera
meles (Fabricus,1792)
|
1,20
|
1,11
|
2,48
|
|
Hypera
nigrirostris (Fabricius,1775)
|
0
|
0
|
1,65
|
|
Lixus
punctirostris (Boheman,1843)
|
0
|
0
|
0,82
|
|
Otiorhynchus
porcatus (Herbst,1795)
|
0
|
0
|
0,82
|
|
Phyllobius
argentatus (Linnaeus,1758)
|
1,20
|
1,11
|
0
|
|
Phyllobius
brevis (Gyllenhal,1834)
|
1,20
|
0
|
0
|
|
Phyllobius
pyri (Linnaeus,1758)
|
1,20
|
0
|
0
|
|
Phyllobius
viridicollis (Fabricius,1792)
|
0
|
3,33
|
0
|
|
Pleurocleonus
sollicitus (Gyllenhall in
|
0
|
0
|
|
Sitona
cylindricollis (Fabraeus,1840)
|
0
|
0
|
1,65
|
|
Sitona
lineatus (Linnaeus,1758)
|
1,20
|
1,11
|
0
|
|
Sitona
macularius (Marsham,1802)
|
3,61
|
5,55
|
0
|
|
Sitona
sulcifrons (Thunberg,1798)
|
0
|
0
|
0,82
|
|
Strophosoma
albolineatus (Seidlitz,1867)
|
0
|
0
|
0,82
|
|
Strophosoma
capitatum (Degeer,1775)
|
18,07
|
21,11
|
36,36
|
|
Tychius
lineatulus (Stephens,1831)
|
42,17
|
34,44
|
28,10
|
|
Всего
экземпляров
|
83
|
90
|
121
|
|
Всего
видов
|
14
|
12
|
19
|
|
Индекс
Шеннона
|
0,73
|
0,76
|
0,82
|
|
Индекс
Симпсона
|
0,25
|
0,23
|
0,24
|
|
Выравненность
по Пиелу
|
0,28
|
0,31
|
0,28
|
Подобный характер имело также распределение по видовому
богатству. Так, наибольшее число видов было зафиксировано на стационаре «Опушка
леса» - 19 видов (таблица 1). Практически одинаковое число видов (14 и 12) было
отмечено на стационарах «Заливной луг» и «Суходольный луг» соответственно.
Таким образом, как по видовому богатству, так и по численности долгоносики
преобладали на стационаре «Опушка леса». Это в определенной степени можно
объяснить тем, что основная масса встреченных видов - гигрофилы, в связи с чем
засушливые условия суходольного луга не являются оптимальными для обитания
большинства собранных видов долгоносиков-хортобионтов..
Видовая структура долгоносиков на всех трех
исследованных стационарах варьирует в значительной степени. Так,
видами-доминантами, которые преобладали на всех трех изученных участках были
Tychius lineatulus, Strophosoma capitatum и Apion fulvipes. При этом
численность их была хоть и высокой, но не равнозначной для всех стационаров.
Первый из рассматриваемых видов (Tychius lineatulus) сокращал свое
относительное обилие при смене влажности на биотопах - от 42,17до 28,10% на
заливном лугу и опушке леса (таблица). Второй общий доминант (Strophosoma
capitatum) , имел среднее относительное обилие на стационаре «Суходольный луг»
- всего 21,11%. На опушке леса его численность возросла незначительно и
составила уже 36,36%. На самом влажном из исследованных участков (стационар
«Заливной луг») ее численность была самой низкой (практически в два раза ниже,
чем на суходольном лугу) - 18,07%. Третий доминант Apion fulvipes имел наибольшее
относительное обилие на суходольном лугу - 24,44% и практически в два раза
снизил своё относительное обилие на стационаре «Опушка леса» -14,88%. Это можно
объяснить тем, что стационар «Опушка леса» затенён, что в некоторой степени
препятствует нормальному развитию данного вида.
.3 Биотопический преферендум божьих коровок в
исследованных сообществах
.3.1 Стационар «Опушка леса»
В результате проведенных исследований на данном
стационаре было собрано 121 экземпляр долгоносиков, относящихся к 19 видам. Характеризуя
биотопическую приуроченность долгоносиков на стационаре «Опушка леса», можно
сказать, что по видовому богатству эвритопные виды не уступали лесным видам
(рисунок 15), что и следовало ожидать.
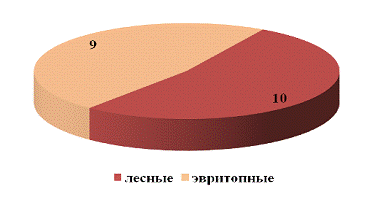
Рисунок 15 - Биотопический преферендум
долгоносиков-хортобионтов на стационаре «Опушка леса» (виды)
Следует отметить, что луговые виды не были нами
отмечены. Это объясняется переувлажнением данного биотопа.

Рисунок 16 - Биотопический преферендум
долгоносиков-хортобионтов на стационаре «Опушка леса» (особи)
Многочисленное разнообразие лесных видов
характеризуется присутствием деревьев, которые и формируют стационар «Опушка
леса», т.е. растительность данной территории создала оптимальные условия для
относительного преобладания лесных видов. Можно предположить, что относительное
обилие эвритопных видов связано с расположенным недалеко от места исследования
суходольного луга (рисунок 15).
На стационаре «Опушка леса» по относительному
обилию преобладали эвритопные (90%) и лесные (10%) виды. Обилие эвритопных
видов связано с тем, что подобные виды могут обитать в разных природных
условиях и на разных стационарах. Лесные виды встречаются в незначительном
количестве (относительное обилие - 10%) (рисунок 16). Это может быть связано с
конкуренцией со стороны эвритопных видов.
.3.2 Стационар «Заливной луг»
В результате исследований в период с мая по
сентябрь на данном стационаре было собрано 83 экземпляра долгоносиков 14 видов.
Характеризуя биотопическую приуроченность
долгоносиков на заливном лугу, можно сказать, что по видовому богатству
преоблодания не наблюдалось. Однако лесные виды не были встречены (рисунок 17).
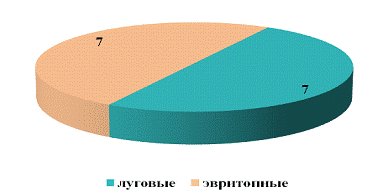
Рисунок 17 - Биотопический преферендум долгоносиков-хортобионтов
на стационаре «Заливной луг» (виды)
Но, несмотря на это, относительное обилие особей
эвритопного вида, значительно больше, чем особей лугового вида, и составило 86%
(рисунок 18).
Такое распределение видов по биотопам может
зависеть от ряда факторов: пищевая база(большинство видов долгоносиков питаются
луговыми растениями и листьями деревьев), биотопическая приуроченность (каждому
виду характерен свой биотоп), климат и микроклимат (погодные условия,
температура, влажность), сезонная ритмичность (погодные условия во время
размножения), физико-химические факторы в разных средах.

Рисунок 18 - Биотопический преферендум
долгоносиков-хортобионтов на стационаре «Заливной луг» (особи)
По относительному обилию эвритопный вид занимает
86% от общей массы зафиксированных долгоносиков. Лесные виды представлены
незначительным количеством и составили всего лишь 14% (рисунок 18).
Так как лесные виды не характерны для данного
стационара, то они и не были нами встречены.
В заключение можно сказать, что по видовому
богатству преобладания между эвритопными и луговыми видами не наблюдалось. Что
же касается относительного обилия, то преобладающими видами выступили
эвритопные, что может быть связано опять же с благоприятными условиями
обитания.
.3.3 Стационар «Суходольный луг»
В результате исследований в период с мая по
сентябрь на данном стационаре было собрано 90 экземпляра
долгоносиков-хортобионтов, относящихся к 12 видам.
Характеризуя биотопическую приуроченность
долгоносиков на суходольном лугу, можно сказать, что по видовому богатству
преобладали эвритопные виды. К этой категории были отнесены 5 видов
долгоносиков (Apion difficile, Apion fulvipes, Chlorophanus gibbosus,
Chlorophanus viridis, Hypera meles, Strophosoma capitatum, Tychius lineatulus).
На два вида им уступают луговые виды, их
количество составило 5 видов (Apion pomonae, Phyllobius argentatus, Phyllobius
viridicollis, Sitona lineatus, Sitona macularius ) (рисунок 19).
Такое незначительное колебание в количестве
встреченных видов может быть связанно с наиболее оптимальными условиями
обитания чем на стационарах «Опушка леса» и «Заливной луг».

Рисунок 19 - Биотопический преферендум
долгоносиков-хортобионтов на стационаре «Суходольный луг» (виды)
На стационаре «Суходольный луг» по видовому
богатству преобладал эвритопный вид - 88%, это связано с тем, что именно эти
виды неприхотливы к окружающим условиям и могут обитать на разных биотопах. В
значительном количестве от общей массы видовое богатство луговых видов на
данном стационаре не велико и составляет всего 12% от общей массы встреченных
видов (рисунок 20).
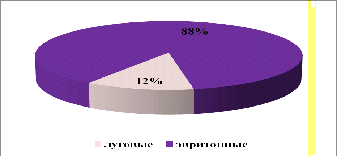
Рисунок 20 - Биотопический преферендум
долгоносиков-хортобионтов на стационаре «Суходольный луг» (особи)
Хотя данный стационар и является естественным
местом обитания луговых видов, такое видовое распределение по биотопам может
зависеть от ряда факторов: пищевая база, биотопическая приуроченность, климат и
микроклимат, сезонная ритмичность, физико-химические факторы в разных средах.
.4 Гигропреферендум долгоносиков-хортобионтов в
исследованных сообществах
.4.1 Стационар «Опушка леса»
На опушке леса большую роль по видовому составу
играли гигрофильные виды - 18 (рисунок 21).
В гораздо меньшей степени представлены мезофилы
(их всего 1 вид - Chlorophanus viridis). Мы можем предложить, что для данного
стационара гигрофилы являлись нехарактерными видами.

Рисунок 21 - Гигропреферендум
долгоносиков-хортобионтов на стационаре «Опушка леса» (виды)
Исходя из результатов представленных на рисунке
21 можно наблюдать, что наиболее высокое видовое богатство характерно для
гигрофильных видов (18 видов из 19). Это виды предпочитающие места с нормальным
увлажнением.
На стационаре «Опушка леса», как и на
стационарах «Суходольный луг» и «Заливной луг», преобладали гегрофилы - 99%,
так как гегрофилы предпочитают места с повышеным увлажнением. В незначительном
количестве встречались мезофилы - 1% (Chlorophanus viridis).

Рисунок 22 - Гигропреферендум
долгоносиков-хортобионтов на стационаре «Опушка леса» (особи)
Можно предположить, что это связано с тем, что
стационар «Опушка леса» место более увлажненное, чем стационар «Суходольный
луг».
.4.2 Стационар «Суходольный луг»
На суходольном лугу по видовому составу между
гигрофилами и мезофилами преобладания не наблюдалось (рисунок 23). Лесные виды
и вовсе не были встречены, так как они данного стационара являются
нехарактерными видами.

Рисунок 23 - Гигропреферендум
долгоносиков-хортобионтов на стационаре «Суходольный луг» (виды)
На стационаре «Суходольный луг» в большом
изобилии встречались гигрофилы - 86% (рисунок 24), что является не характерным,
так как гигрофилы предпочитают места с повышенной влажностью.

Рисунок 24 - Гигропреферендум
долгоносиков-хортобилнтов на стационаре «Суходольный луг» (особи)
Мезофилы, по относительному обилию, были
встречены в незначительном количестве, всего 14%, хотя излюбленными местами
мезофилов являются места с нормальным увлажнением.
.4.3 Стационар «Заливной луг»
На заливном лугу большую роль по видовому
составу играли гигрофильные виды - 9 (рисунок 25). Это явилось ожидаемым
результатом по той причине, что гигрофилы предпочитают места с повышенной
влажностью, которым и является заливной луг.

Рисунок 25 - Гигропреферендум
долгоносиков-хортобионтов на стационаре «Заливной луг» (виды)
В незначительно меньшей степени представлены
мезофилы (их всего 1 вид).
Исходя из данных представленных на рисунке 26
можно наблюдать, что широко представлены гигрофильные виды, их относительное
обилие составило 88%. Обилие особей мезофильного вида составило 12%.
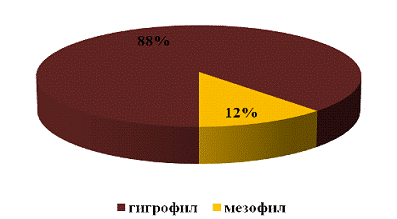
Рисунок 26 - Гигропреферендум
долгоносиков-хортобионтов на стационаре «Заливной луг» (особи)
Причиной такого распределения особей является
то, что мезофильные виды предпочитают места с нормальным увлажнением, но не
зависимо от этого распространяются повсеместно.
В заключении можно сказать, что и по видовому
богатству и по относительному обилию преобладали виды, предпочитающие места с
повышенным увлажнением (гигрофилы). Мезофилы встречались в незначительном
количестве, так как предпочитают места с более нормальным увлажнением, чем
рассмотренные стационары.
ЗАКЛЮЧЕНИЕ
В ходе проведённого исследования нами можно
сделать ряд выводов:
. Был уточнен видовой состав
долгоносиков-хортобионтов, обитающих в открытых урбоценозах г. Гомеля. Всего
было собрано 294 экземпляра исследуемых долгоносиков 45 видов.
. На основе изучения видового состава на
стационаре «Опушка леса» отловлено наибольшее количество особей (121) . Ниже
численность долгоносиков была отмечена на стационаре «Суходольный луг» - 90
особей, а наименьшее количество экземпляров было характерно для стационара
«Заливной луг» - всего 83 особи.
Подобный характер имело также распределение по
видовому богатству. Так, наибольшее число видов было зафиксировано на
стационаре «Опушка леса» - 19 видов. Практически одинаковое число видов (14 и
12) было отмечено на стационарах «Заливной луг» и «Суходольный луг»
соответственно. Таким образом, как по видовому богатству, так и по численности
долгоносики-хортобионты преобладали на стационаре «Опушка леса».
Также нами было выявлено, что
видами-доминантами, которые встречались на всех трех изученных участках были
Tychius lineatulus, Strophosoma capitatum и Apion fulvipes .
. Составлена электронная база данных,
представленная в виде аннотированного списка. Который включил 82 вида,
встречающихся на территории Гомельской области.
. На основе изучения биотопичесского
преферендума долгоносиков в исследованных сообществах было выявлено, что по
видовому богатству и по относительному обилию на всех трёх биотопах преобладали
эвритопные виды.
. На основе гигропреферендума долгоносиков в
исследованных сообществах можно прийти к выводу, что на всех трех стационарах
преобладали гигрофилы.
Таким образом, сообщества
долгоносиков-хортобионтов открытых урбоценозов г. Гомеля сложены
преимущественно эвритопными, реже - луговыми видами, предпочитающие повышенные
условия увлажнения.
В связи с этим, считаем, что поставленные задачи
реализованы, а цель исследования достигнута.
СПИСОК ЛИТЕРАТУРЫ
Александрович
О. П., Лопатин И. К. Каталог жесткокрылых. Минск, 1996.
Арнольди
Л. В. Жесткокрылые, или жуки - Coleoptera. Животный мир СССР., т. 4. М. - Л. ,
1953. - С. 434- 485.
Брамсон
К. Л. Вредные насекомые и меры борьбы с ними. Ч.1. Екатеринослав, 1894.
Воронцова
А. И. "Лесная энтомология. Учебник для вузов" //Изд. 3-е, перераб.
М., "Высш. школа", 1975. - С.- 368 ( с изменениями).
Воронцова
А.И.,Семенкова И.Г. Лесозащита: Учебник для техникумов. - 2-е изд.,исп. и доп.
- М.: лесн.пром-сть, 1980 г. - 328 с.
Добровольский
Б. В. Распространение вредных насекомых. М., 1959ёанiciянi Т. Р. Матэрыялы па
бiялогii жукоý - апiёнаý
у
Беларусi. // Весцi. ААНБ ССР, Сер. бiял. навук., №4, 1961а. - С. 113 - 122.
Иоаннисиани
Т. Г. Жуки- долгоносики (Coleoptera, Curculionidae) Белоруссии. Мн., «Наука и
техника», 1972. - С. 5- 7.
Иоаннисиани
Т. Г. Эколого- географические особенности распространения некоторых видоа
жуков- долгоносиков (Coleoptera, Curculionidae) в Белоруссии. Фауна и экология
животных Белоруссии. Минск, 1969. - С. 178- 190.
Исаев
А. Ю. , Грачёв В. Г. Энтомологическое обозрение, LXXIII, 2, 1994. - С. 318-
320.
Краткие
определительные таблицы жуков - вредителей запасов (Для работников карантина
растений).- Москва, 1968.
Крушев
Л.Т. Биологические методы защиты леса от вредителей. - Минск: Лесная
промышленность, 1973. - 159 с.
Крыжановский
О.Л. Состав и распространение энтомофаун земного шара. - М.: Т-во научных
изданий КМК, 2002. - 237 с.
Лаврова
Н. К. Виды слоников, встречающиеся на культурных и дикорастущих бобовых
растений в Беларуси. // 2 зоологическая конференция БССР, Минск, октябрь 1962
Мазинг
В.В. Экосистема города, её особенности и возможности оптимизации / В.В. Мазинг
// Экологические проблемы городских систем. - Мн., 1984. - С. 181-191.
Молодова,
Л.П. Структура фауны жесткокрылых-герпетобионтов в биотопах Гомеля // Вестник
Белор. гос. ун-та. Сер. 2, Химия. Биология, География. - 1990. - № 3. - С.
39-42.
Молодова,
Л.П. Структура фауны жесткокрылых-герпетобионтов в различных биотопах г. Гомеля
/ Л.П. Молодова, Т.Р. Ряхова // Биоиндикация в городах и пригородных зонах: сб.
науч. ст. / Рос. АН. Ин-т эволюц. морфологии и экологии животных им. А.Н.
Северцова; редкол.: Д.А. Криволуцкий (отв. ред.) [и др.]. - М.: Наука, 1993. -
С. 79-83.
Околув
Ч., Савицкий Б.П., Дьяченко Н.Г. Фоновые виды насекомых Беловежской пущи //
Животный мир Белорусского полесья, охрана и рациональное использование: Тез.
докл. обл. науч.-практ. конф. УНПО "Фауна Полесья". Октябрь 1982 г.
-Гомель: Гомельский ГУ. -1982. -С. 52-60.
Определитель
насекомых Дальнего Востока СССР . Т. III. Жесткокрылые, или жуки. Ч. 1.- Л.;
Наука. 1989.- 572 с.
Песенко
Ю. А. Принципы и методы количественного анализа в фаунистических исследованиях.
- М.: Наука, 1982 - 284 с.
Под
ред. Бей- Биенко Г. Я. Определитель насекомых европейской части СССР в пяти
томах. Т. 2.- М.- Л.; Наука, 1965
Соколов
Е.А. Вредители запасов, их карантинное значение и меры борьбы / Под общей
редакцией и с предисловием М.И. Маслова. - Оренбург: Печатный дом «Димур»,
2004.
Справочник
- определитель карантинных и других опасных вредителей сырья, продуктов запаса
и посевного материала / Сост. Я.Б. Мордкович, Е.А. Соколов; Под ред. В.В.
Поповича. - М.: Колос, 1999
Сушко
Г. Г. Эколого- фаунистическая характеристика жесткокрылых (Coleoptera)
естественных и антропогеннонарушенных верховых болот Белорусского Поозерья. //
Ученые записки УО «ВГУ им. П. М. Машерова». Т. 3.- Естественные науки.
Биология, 2004. - 276 - 280.
Танский
В.И. Экологические основы вредоносности насекомых. - М.: Агропромиздат, 1988. -
127 с.
Тер-
Минасян М. Е. Обзор видов рода Curculio L. фауны СССР и определённых территорий
// Энтомологическое обозрение.- 1956.- С. 168- 173.