Semisynthetic and synthetic antibiotics
Anotation
words: Antibiotics, medicine,
E.coli, β-lactam antibiotics,
penicillin,
cephalosporin, macrolides, fluoroquinolone, sulfonamide, tetracycline,
aminoglycoside,
antibiotic resistance, Streptomyces
violatus.
In biotechnology,
biologically active substances describes the beneficial or adverse
effects of a drug <#"821820.files/image001.jpg">
growth
of S. violatus and the production of antibiotic in a starch-nitrate medium were
monitored over a period of 14 days (Fig. 1). The antibiotic production by S.
violatus occurred in a growth-phase dependent manner and the highest antibiotic
yield was obtained in the late exponential phase and the stationary phase,
indicating that it is mainly a product of secondary metabolism. Similar results
were observed for streptomycin production in batch cultures of S. griseus [29]
when grown in a mineral medium and for the production of
candicidin in liquid grown cultures of S. Griseus.The results also showed that
S. violatus produced a blue pigment associated with the antibiotic appearance
in the culture. It was noticed that a direct tight relationship occurred
between the antibiotic production and the intensity of the blue colour formed
in the culture (r=0.95). These results may suggest the production of a
pigmented antibiotic in S. violatus cultures. The production of the
blue-pigmented antibiotic actinorhodin and its physiology are known in S.
coelicolor cultures.
 1. Effect of different incubation periods on the production of
antibiotic by Streptomyces violatus.
1. Effect of different incubation periods on the production of
antibiotic by Streptomyces violatus.
2.2.1
Influence of some cultivation factors on the production of antibioticof
antibiotic production in batch cultures of S. violatus was carried out. This
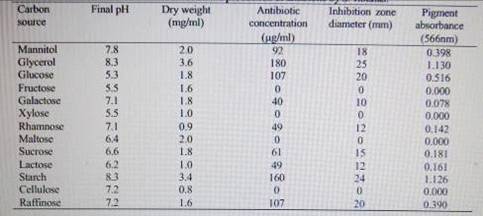
strain was able to grow in all the tested carbon sources (Table 2). However,
maximum antibiotic production was obtained in cultures supplemented with glycerol
as a sole carbon source followed by cultures containing starch. Cultures
containing fructose, maltose, xylose or cellulose did not yield any detectable
amounts of the antibiotic. The results also showed that the increase of
glycerol level in the culture from 10g/l to 12.5 g/l led to 1.32-fold increase
in antibiotic production (Fig 2). The utilisation of glycerol and starch by S.
violatus for growth and production of the antibiotic indicates the presence of
an active uptake system for these substrates. Glycerol was also found to be
used as a sole carbon source by other Streptomyces species.
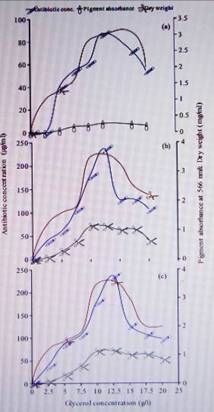 2. Effect of glycerol concentration on the production of
antibiotic byStreptomyces violatus at different incubation periods: a) 4 days,
b) 7 days and c) 10 days.
2. Effect of glycerol concentration on the production of
antibiotic byStreptomyces violatus at different incubation periods: a) 4 days,
b) 7 days and c) 10 days.
2 -
Effect of different carbon sources on the production of antibiotic by S.
violatus.
.2.2
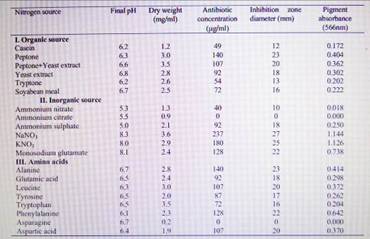
Influence of nitrogen source results revealed that the level of antibiotic
production may be greatly influenced by the nature, type and concentration of
the nitrogen source supplied in the culture medoium (Table 3). Similar
observations have been reported by many investigators. The highest antibiotic
production was obtained in cultures of S. Violatus containing sodium nitrate or
potassium nitrate as a nitrogen source, followed by cultures containing
peptone, alanine, monosodium glutamate or phenylalanine. However, cultures
containing asparagine or ammonium citrate did not yield any antibiotic activity
and showed lowest growth. The results also showed that the concentration of
NaNO3 (Fig. 3) greatly influenced the production of the antibiotic by S.
violatus cultures, while the maximum antibiotic yield was obtained in cultures
suplemented with 2.5 g/l NaNO3. These results are in partial agreement with
those of other investigators. A negative effect of asparagine on the production
of cephamycin C was also observed on cultures of S. cattleya, S. latamdurans
and Cephalosporium acremonium.
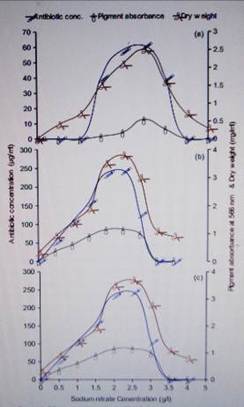
Figure 3. Effect of
sodium nitrate (NaNO 3) concentration on the production of antibiotic by
Streptomyces violatus at different incubation periods: a) 4 days, b) 7 days and
c) 10 days.
3 - Effect of different
nitrogen sources on the production of antibiotic by S. violatus.
2.2.3 Influence of
potassium phosphate and magnesium sulphate salts is a major factor in the
synthesis of a wide range of antibiotics. However, an excessive amount of
inorganic phosphate suppresses the production of antibiotics such as
tetracycline, actinomycin and candicidin (Kishimoto et al. 1996). The results
of the present work (Fig 4) showed that KH2PO4 was not favourable for the
production of antibiotic by S. violatus, while K2HPO4 at a concentration of
1g/l yieldes an inhibition zone of 22 mm, equivalent to an antibiotic
concentration of 128 µg/ml. It was also observed that addition of a mixture of
both phosphate salts (KH2PO4 and K2HPO4) showed the most positive effect on the
production of antibiotic by S. violatus. The antibiotic concentration reached
its maximum value (245µg/ml) when using a phosphate salt mixture of 1g/l,
showing a 1.9-fold and 6.1-fold increase when compared to the highest values
obtained when K2HPO4 and KH2PO4 were individually supplied to the medium,
respectively. These results are in agreement with those reported by other
investigators. The results also showed that addition of 0.5g/l magnesium sulphate
to the culture medium was optimal for the production of a maximum yield of
antibiotic by S. violatus (Fig 5). At this MgSO4.7H2O concentration, the
antibiotic yield was 4.2-fold than that in cultures devoid of magnesium
sulphate. The importance of magnesium sulphate for antibiotic production by
other Streptomyces species has been reported by several investigators . The
effects of magnesium availability are presumably due to requirements of this cation
for protein synthesis, and its depletion may restrict enzyme synthesis and
activity.
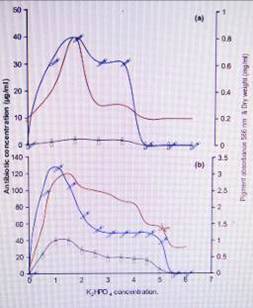 4.
Effect of different (a) KH2PO4 and (b) K2HPO4 concentrations on the production
of antibiotic by Streptomyces violatus.
4.
Effect of different (a) KH2PO4 and (b) K2HPO4 concentrations on the production
of antibiotic by Streptomyces violatus.
results also showed that
addition of 0.5g/l magnesium sulphate to the culture medium was optimal for the
production of a maximum yield of antibiotic by S. violatus (Fig 5). At this
MgSO4.7H2O concentration, the antibiotic yield was 4.2-fold than that in
cultures devoid of magnesium sulphate. The importance of magnesium sulphate for
antibiotic production by other Streptomyces species has been reported by
several investigators. The effects of magnesium availability are presumably due
to requirements of this cation for protein synthesis, and its depletion may
restrict enzyme synthesis and activity (Aasen et al. 1992; mNatsume et al.
1994).
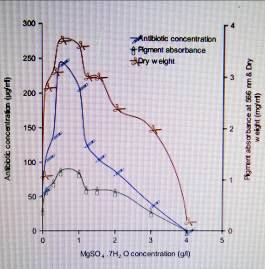 5.
Effect of (MgSO4 .7H2O) concentration on the production of antibiotic by
Streptomyces violatus.
5.
Effect of (MgSO4 .7H2O) concentration on the production of antibiotic by
Streptomyces violatus.
.2.4 Influence of trace
elements results given in Table 4 showed that iron and manganese could play an
important role in the promotion of antibiotic production, the highest dry
weight (3.8 mg/ml) was also recorded for manganese. A slight increase in the
antibiotic concentration was recorded for Cu, whereas Zn addition lowered the
antibiotic concentration compared to the control. The highest antibiotic
concentration was achieved in the presence of all elements in the culture
medium, yielding a 2.1-fold increase compared to the control reported on the
importance of ferrous ions for the growth and antibiotic production by
Streptoverticillium rimofaciens. Mansour et al.[30]showed that manganese ions enhanced growth and granaticin
production in S. violaceolatus.
Table 4 - The role of
trace elements on the production of antibiotic by S. violatus.
3. Protection of workers and life safety
In modern conditions of development
of production of a problem in the field of industrial and ecological safety
tend to an aggravation. Relevance of a problem of safety of the person and
environment is especially sharply shown directly at the enterprises when
carrying out technological processes. On trebitel of medicines are interested in
receiving qualitative and safe preparations. The workers who are carrying out
technological process have to have optimum working conditions.
The main gas emissions in the
atmosphere of the enterprises for production of antibiotics containing harmful
substances include, except air emissions of all-exchange and local ventilation,
technological air emissions at biosynthesis of antibiotics, emissions of boiler
and some other auxiliary productions. Various ways of cleaning provide catching
about 60% of the harmful substances departing from all sources of
pollution.harmful substances consist generally of carbon monoxide (77,4%),
sulphurous gas (15,2%) and nitrogen oxides (7,4%).of organic solvents making
24,3% of total amount of the thrown-out substances (tab. 3) belong to liquid
and gaseous products, specific to production of antibiotics.Besides, at air
emissions there is a number of impurity of vapors of various substances making
0,4% of total amount of the liquid and gaseous products released into the atmosphere.
Among them chloride hydrogen, vapors of hydrochloric acid, formaldehyde and
prevails.substances, nonspecific for production of antibiotics, in emissions
are caught by gas-and-dust cleaning installations
for 90%, gaseous emissions of boiler rooms dissipate by means of high pipes.
Specific to production of antibiotics firm substances from air emissions for
92,5%, organic solvents - for 10%, 5,4% of the volume of air emissions at
biosynthesis of antibiotics are neutralized.
In rooms of storage of finished
goods, collecting condensate, preliminary processing of barrels, pump station
of reverse water supply, the foreman, the supervising foreman the all-exchange
supply and exhaust ventilation is provided. Supply of stitched air and removal
of the exhaust is carried out from the top zone, for rooms of packaging of
ointment - from the lower zone. In the stitched P-1 installation external air
is cleared of dust in the filter 3 classes, warmed up in the superficial heat
exchanger and moistened during the cold period of year, during the warm period
- is only cleared of dust.
Thus,
this system of ventilation of air is effective since provides necessary
parameters of air for technological process, favorable microclimatic
conditions, deletes harmful substances from air of a working zone.
4. Ecological
conservation
Ecological factors
influencing the effects of antibiotic production were explored experimentally
and theoretically. A spatially structured model was used to model the dynamics
of antibiotic-producing and nonproducing bacteria in which growth of the
nonproducers was reduced by neighbouring antibiotic producers. Various factors
affecting spatial interactions between the bacteria were examined for their
impact on antibiotic producers. Spatial clustering had a positive impact on the
effect of antibiotic production, as measured by the decline in growth of the
nonproducing strain, while increasing the initial density of the nonproducing
strain had a negative impact. Experiments examined the growth of
antibiotic-producing Streptomyces species and a nonproducing, antibiotic-sensitive
strain of Bacillus subtilis that were coinoculated on surface media. There was
an effect of the Streptomyces on Bacillus growth in some experiments but not in
others. In light of the predictions from the model, unintentional clustering of
cells is a more likely explanation for this finding than different initial
Bacillus densities. The importance of spatial structure seen in this study is
consistent with a terrestrial rather than an aquatic distribution of
antibiotic-producing bacteria, and may have implications in the search for
novel antibiotics.
Over the last 40 years, there has been a steady supply of novel,
useful antibiotics produced by microbes isolated from soil and other natural
environments. The increased efficiency of screening procedures in the last
decade has played a major part in maintaining this supply. However, the
selection and sampling of natural environments are still essentially random
processes. The main reasons for this are an almost total lack of knowledge of
the significance of antibiotics in nature, deficiencies in the taxonomy of
antibiotic-producing microbes and its application, and lack of information
about the distribution and ecology of known or potential antibiotic producers.
The origins of these problems are discussed and some possible solutions are
suggested.
A new perspective on the topic of antibiotic resistance is
beginning to emerge based on a broader evolutionary and ecological
understanding rather than from the traditional boundaries of clinical research
of antibiotic-resistant bacterial pathogens. Phylogenetic insights into the
evolution and diversity of several antibiotic resistance genes suggest that at
least some of these genes have a long evolutionary history of diversification
that began well before the ‘antibiotic era’. Besides, there is no indication
that lateral gene transfer from antibiotic-producing bacteria has played any
significant role in shaping the pool of antibiotic resistance genes in
clinically relevant and commensal bacteria. Most likely, the primary antibiotic
resistance gene pool originated and diversified within the environmental
bacterial communities, from which the genes were mobilized and penetrated into
taxonomically and ecologically distant bacterial populations, including
pathogens. Dissemination and penetration of antibiotic resistance genes from
antibiotic producers were less significant and essentially limited to other
high G+C bacteria.
Conclusion
Antibiotics are biotechnological products that inhibit bacterial
growth or kill bacteria. They are naturally produced by microorganisms, such as
fungi, to attain an advantage over bacterial populations
<https://www.boundless.com/definition/population/>. Antibiotics are
produced on a large scale by cultivating and manipulating fungal cells. Many
antibacterial compounds are classified on the basis of their chemical or
biosynthetic origin into natural, semisynthetic, and synthetic. Another
classification system is based on biological activity. In this classification,
antibiotics are divided into two broad groups according to their biological
effect on microorganisms: bactericidal
<https://www.boundless.com/definition/bactericidal/> agents kill
bacteria, and bacteriostatic
<https://www.boundless.com/definition/bacteriostatic/> agents slow down
or stall bacterial growth. of some locally
isolated actinomycetes for the production of antibiotic(s). A survey of four
locally isolated Streptomyces strains for antibiotic production was carried out
in static and shaken cultures. It was generally observed that the growth and
antibacterial activity obtained in static cultutres were higher than shaken
cultures. Streptomyces astreogriseus showed the longest incubation time (12
days) needed to obtain maximum antibacterial activity, while Streptomyces
violatus showed a relatively short time (7-days) and produced the highest
activity among the tested strains. Streptomyces violatus [30] was also characterised by its broader antibacterial activity,
because it affected the growth of all the tested bacteria, showing a stronger
activity on S. aureus and B. subtilis. Accordingly, S. violatus was selected
for further investigation.Antibiotics are produced
industrially by a process of fermentation, where the source microorganism is
grown in large containers (100,000-150,000 liters or more) containing a liquid
growth medium. Oxygen concentration, temperature, pH
<http://www.chemeurope.com/en/encyclopedia/PH.html>and nutrient <http://www.chemeurope.com/en/encyclopedia/Nutrient.html>
levels must be optimal, and are closely monitored and adjusted if necessary.
The
effectiveness of individual antibiotics varies with the location of the
infection, the ability of the antibiotic to reach the site of infection, and
the ability of the microbe to inactivate or excrete the antibiotic. Some
anti-bacterial antibiotics destroy bacteria (bactericidal), whereas others
prevent bacteria from multiplying (bacteriostatic).Oral antibiotics are simply
ingested, while intravenous
<http://www.bionity.com/en/encyclopedia/Intravenous.html> antibiotics are
used in more serious cases, such as deep-seated
systemic infections
<http://www.bionity.com/en/encyclopedia/Systemic_infection.html>.
Antibiotics may also sometimes be administered topically, as with eye drops or
ointments.the last few years, three new classes of antibiotics have been
brought into clinical use. This follows a 40-year hiatus in discovering new
classes of antibiotic compounds. These new antibiotics are of the following
three classes: cyclic lipopeptides (daptomycin), glycylcyclines (tigecycline),
and oxazolidinones (linezolid). Tigecycline is a broad-spectrum antibiotic,
while the two others are used for gram-positive infections. These developments
show promise as a means to counteract the growing bacterial resistance to
existing antibiotics.
References
1 Běhal
V. Bioactive products from Streptomyces. Adv Appl Microbiol 2000;47:113-156.
2Omura S, Sasaki Y, Iwari Y and
Takeshima H. Staurosporine, a potentially important gift from a microorganism.
J Antibiot 1995;48:535-548.
Běhal
V. Nontraditional microbial bioactive metabolites.
Folia Microbiol 2001;46: (6) 000-000, (in press).
4
Umezawa
K, Aoyagi T, Suda T, Hamada M and Takeuchi T. Bestatin, an inhibitor of
aminopeptidase B, produced by actinomycetes. J Antibiot 1976;30:170-173.
Bennett
JW and Bentley R. What is a name?-Microbial secondary metabolites. Adv Appl
Microbiol 1989;35:1-28.
6
Lipman
F. Attempts to map a prcoess evolution of peptide biosynthesis. Science
1971;173:875-884.
7
Laland
SG and Zimmer T-L. Bioactive peptides produced by microorganisms. Essay Biochem
1973;9:31-57.
Billich
A and Zocher R. Enzymatic synthesie of cyclosporine A. J Biol Chem
1987;262:17258-17259.
10
Běhal
V and Hunter IS. In: Vining LC and Stuttard C (eds)
Genetics and Biochemistry of Antibiotics Production. Butterworth-Heinemann,
Boston, 1995;359-384.
11
Kleinkauf
H von Doehren H In: Kleinkauf H., von Doehren H, Dornauer H and Nasemann G
(eds) Regulation of Secondary Metabolite Formation, VCH Verlagsgesselshaft,
Weinheim, 1986:173-207.
12
Roland
I, Froyshov O and Laland G. A rapid method for the preparation of three enzymes
of bacitracin synthetase essentialy free from other proteins. FEBS Lett
1977;84:22-24.
13
Ishihara
HM, Hara N and Iwabuchi T. Molecular cloning and expression in Escherichia coli
of Bacillus licheniformis bacitracin synthetase gene 2 gene. J Bacteriol
1989;171: 1705-1711.
14
von
Doehren H. In: Vining LC and Stuttard C (eds) Genetics and Biochemistry of
Antibiotic Production, Butterworth-Heinemann, Boston, 1995;129-171.
15
Martin
JF and Liras P. Beta-lactams. Adv Biochem Eng 1989;39:153-187.
Jensen
SE and Demain AL. In: Vining LC and Stuttard C (eds) Genetics and Biochemistry
of Antibiotics Production, Butterworth-Heinemann, Boston, 1995;239-268.
17
Newton
GGF and Abraham EP. Isolation of cephalosporin C, penicillin-like antibiotic
containing D-α-aminoadipic acid. Biochem J
1956;62:651-658.
18
Martin
JF. Α-aminoadipyl-cysteinyl-valine santhetases in β-lactam producing
organisms. J Antibiot 2000;53:1008-1021.
19
Abraham
EP. In: Kleinkauf H, von Doehren H, Dornauer H and Nesemann G (eds) Regulation
of secondary metabolite formation. VCH Verlagegesselshaft, Weinheim,
1986;115-132.
20
Parenti
F and Cavalleri B. Proposal to name the vancomycin-ristoceti like glycopeptides
as dalbaheptides. J Antibiot 1989;42:1882-1883.
21
Williams
DH, Stone MJ, Mortishire-Smith RJ and Hauck PR. Molecular recognition by
secondary metabolites. Biochem Pharmacol 1990;40:27-34.
22
Bentley
R and Bennett JW. Constructinc polyketides: From Collie to combinatorial
biosynthesis. Ann Rev Microbiol 1999;53:411-446.
23
Hopwood
DA and Sherman DH. Molecular genetic of polyketides and its comparison to fatty
acid biosynthesis. Ann Rev Genet 1990;14:37-66.
24
Dimroth
P, Walter H and Lynen F. Biosynthesis von 6-methylsalicylsaure. Eur J Biochem
1970;13:98-110.
25
Dimroth
P, Ringelmann E and Lynen F. 6-methylsalicylic acid from Penicillium patulum.
Eur J Biochem 1976;68: 591-596.
26 Aasen IM, Folkvord K &
Levine DW (1992) Development of a process for large- scale chromatographic
purification of an alginate lyase from Klebsiella pneumoniae. Journal of
Applied Microbiology
and Biotechnology 37: 55-60.
27 Bibb M (1996) The regulation
of antibiotic Production in Streptomyces coelicolor A3(2). Microbiology 142:
1335-1344.
28 Bystrykh LV,
Fernander-Moreno M A, Herrema J K, Malportida F, Hopwood D A & Dijkhuizen L
(1996) Production of actinorhodin-related blue pigments by Streptomyces
coelicolor A3(2). Journal of Bacteriology 178: 2238-2244.
30 Said WY (2001) Production of
antibiotics by immobilized Streptomyces strains. Ph.D. Thesis, Faculty of
Science, Alexandria University, Egypt.
1995;48:535-548.