|
Индекс
онтогенетичес- кого
состояния
|
Признак
|
|
p
|
Смешанное
питание (за счет веществ семени и 1 собственной ассимиляции первых листьев);
наличие морфологической связи с семенем (или семядолей); наличие зародышевых
структур: семядолей, первичного (зародышевого) корня и побега.
|
|
j
|
Простота
организации, несформированность признаков и свойств, присущих взрослым
растениям. Наличие листьев иной формы и расположения, чем у взрослых особей;
иной тип нарастания и ветвления (или отсутствие ветвления побегов); возможно
изменение типа корневой системы. Сохранение некоторых зародышевых структур
(корня, побега); потеря связи с семенем; как правило, - отсутствие семядолей.
|
|
im
|
Наличие свойств
и признаков, переходных от ювенильных растений к взрослым: развитие листьев и
корневой системы переходного (полувзрослого) типа, появление отдельных
взрослых черт в структуре побегов (например, смена типов нарастания, начало
ветвления, появление плагиотропных побегов и др.). Сохранение отдельных
элементов первичного побега.
|
|
v
|
Появление
основных черт типичной для вида жизненной формы. Растения имеют характерные
для вида взрослые листья, побеги и корневую систему. Генеративные органы пока
отсутствуют.
|
|
g1
|
Появление
генеративных органов. Преобладание процессов новообразования над отмиранием,
проявляющееся в разных формах. В некоторых случаях (полурозеточные формы и
др.) окончательное формирование взрослых структур.
|
|
g2
|
Уравновешивание
процессов новообразования и отмирания. Максимальный ежегодный прирост
биомассы, максимальная семенная продуктивность.
|
|
g3
|
Преобладание
процессов отмирания над процессами новообразования: резкое снижение
генеративной функции, ослабление процессов корне- и побегообразования. В
некоторых случаях - упрощение жизненной формы (выражающееся в потере
способности к образованию побегов разрастания).
|
|
ss
|
Полное
отсутствие плодоношения (возможно наличие абортивных цветков или соцветий, а
так же скрытогенеративных побегов). Резкое преобладание процессов отмирания
над новобразованием. Возможно упрощение жизненной формы, проявляющееся в
смене способа нарастания, вторичное появление листьев переходного
(имматурного) типа.
|
|
s
|
Накопление
отмерших (прекративших рост) частей растения. Предельное упрощение жизненной
формы, вторичное появление некоторых ювенильных черт организации (форма
листьев, характер побегов и др.). В некоторых случаях - полное отсутствие
почек возобновления и других новобразований.
|
В таблице приводятся наиболее общие качественные признаки
онтогенетических состояний особей разных видов (Ценопопуляции…, 1976).
Выделение имматурных и виргинильных растений представляет
наибольшие трудности. Оба состояния по сути характеризуют переход от детских
структур к взрослым и некоторыми авторами объединяются в одно.
В зависимости от характера жизненной формы качественные
признаки онтогенетических состояний имеют специфическое морфологическое
выражение.


Рис. 1. Ювенильная и имматурные ососби J. cоmmunis
.2 Особенности жизненных форм видов рода Juniperus
L
Анализ литературных материталов выявляет, что исследование
жизненных форм у растений проводили до настоящего времени, главным образом, по
трем основным направлениям: 1) изучение многообразия жизненных форм и решение
вытекающей отсюда проблемы их классификации; 2) изучение отношений жизненных
форм к окружающей среде; 3) изучение эволюции жизненных форм у покрытосеменных
растений. Преимущественно с первым направлением совпадает исследование
закономерностей морфогенеза у разных групп жизненных форм, приобретшее теперь
сравнительно морфологический и онтогенетический характер. В сущности,
наибольшее развитие до настоящего времени получило первое,
классификационное направление. Важнейшую роль в становлении жизненной формы
играет внешняя среда. Но это, конечно, не означает, что жизненная форма любого
растения бесконечно пластична и зависит только от непосредственно
действующих в данный момент условий. Каждый вид растений реагирует на внешние
воздействия в рамках своих наследственно закрепленных возможностей. В трактовке
И.Г. Серебрякова (1964), «жизненная форма - это своеобразная внешняя форма
организмов, обусловленная биологией развития и внутренней структурой их
органов, формируется в определенных почвенно-климатических условиях, как
приспособление жизни к этим условиям», т.e. это форма организмов,
приспособившихся к своей среде обитания под длительным влиянием комплекса
факторов. Его же определение, но более короткое: «Жизненная форма растения это
его габитус (внешняя форма вида), связанный с ритмом развития и приспособленный
к современным и прошлым условиям среды». В начале XX в. появляется серия работ
К. Раункиера (цит. по Серебряков, 1962), определивших на ряд десятилетий общее
развитие учения о жизненных формах. Им была создана система жизненных форм
наземных сосудистых растений, резко отличающаяся от предыдущих своей простотой,
ясностью и законченностью. Жизненные формы К. Раункиер рассматривал как итог и
результат приспособления растений к климатическим условиям страны, возникшие в
процессе исторического развития. В связи с этим жизненные формы, свойственные
той или иной стране могут служить индикатором климата, а само учение о
жизненных формах, по Раункиеру, является основой ботанической географии.
Исследователи выделяют разновидности и формы J. cоmmunis на основе
габитуса, формы кроны, цвета и формы хвои, формы побегов. Деревья…, 1949, 1966;
Гулидова, 1959; Шиманюк, 1964; В. Janusz, 1969; Лучник, 1970; Буторина,
1981; Коропачинский, 1983; Ваганов, 2000; Михеева, 2002).
Чаще всего в литературе J. cоmmunis описывают как кустарник,
либо небольшое деревце с множеством разновидностей и форм. Их разнообразие
связано с обширным ареалом и разнообразием экологических условий произрастания
растения. Описание типичной биоморфы J. cоmmunis. представлено во многих
сводках и определителях (Деревья и кустарники…, 1971; Василевич, 1975; Ареалы
деревьев…, 1977; Определитель…, 1979; Мухамедшин, 1980, 1988, 2000; Гиляров,
1990; Определитель…, 1994; Коропачинекий, Встовская, 2002).


а. б.


в. г.
Рис. 2. Жизненные формы J. cоmmunis. а) дерево, б)
немногоствольное дерево, в) кустарник, г) стланец.
J. cоmmunis встречается в виде
мелкого кустарника (рис. 2.), пирамидального куста и настоящего дерева с
конической вершиной до 12 м высоты и 0,5 м в диаметре. В некоторых областях европейской
части России, в Скандинавии и в северной части Норвегии нередко попадаются
настоящие можжевеловые деревья, достигающие 12-17 м высоту и более 20-24 см в
диаметре возрастом 85-95 лет (Дендрология Узбекистана, 1965; Шиманюк, 1949).
Древовидные формы J. cоmmunis распространены на хорошо
дренированных и относительно богатых почвах (Козубов, Евдокимов, 1965;
Горчаковский, 1968). В популяциях J. cоmmunis под пологом разреженных
древостоев преобладает одноствольные особи (Денисова, 1985; Аши. 1989; Князева,
1999).
У экземпляров, произрастающих в суровых условиях, высоко в
горах, на бедных щебенистых почвах, при сильном затенении главный стебель
наклоняется полегает на землю, со временем сгнивает и на месте одного
экземпляра образуется целая группа особей - куртина. Это не просто подавленные
в росте растения, как считают некоторые исследователи (Мамаев, 1983), а
своеобразные жизненные формы, образовавшаяся в суровых условиях произрастания
(Денисова, Пилипенко, 1978; Грин, 1993). Такие кустистые формы распространены
по берегам морей и на скалистых островах (Козубов, Евдокимов, 1965).Однако
исследователи отмечают, что даже в локальных популяциях могут встречаться самые
разнообразные жизненные формы: стелющиеся, древовидные, кустарниковые.
Морфологическое разнообразие подтверждается высоким уровнем генетической
изменчивости особей внутри популяции (Грейг-Смит, 1967; Камышев, 1986).
Поливариантность жизненных форм выявлена и у особей одного онтогенетического
состояния (Киричок, 1998; Князева, 2000, 2007).
1.3 Систематические и ареалогические особенности
видов рода Juniperus L
Вид Juniperus communis L. относят
к классу Coniferopsida, в котором род Juniperus L. относится к сем.
Кипорисовые (Cupressaceae Rich. ex. Bartl.) и представляет трибу Junipereae (Крюссман,
1986).

Рис. 3. Можжевельник обыкновенный: 1-ветвь
с женскими цветками и ягодообразными шишками; 2 - женский цветок; 3-семяпочки;
4-ветвь с мужскими колосками; 5-мужской колосок; 6-чешуйки с тычинками;
7-поперечный разрез шишки; 8-семя; 9 - семя в увеличенном виде; 10 - семя в
разрезе; 11 - разрез хвои; 12 - сеянец
На основе признаков кариотипа можжевеловые
выделяют в отдельное семейство (Джанаева, 1969; Nagano, et al., 1991).
Основанием для выделения можжевельников в самостоятельное подсемейство
послужили особенности мегаспорофилла. После оплодотворения семязачатков чешуи
мегаспорофиллов быстро разрастаются, становятся мягкими, затем срастаются между
собой, образуя сочную шишку - шишкоягоду (Рис. 3.).
Однако ряд авторов считают, что этих
признаков для выделения семейства можжевеловых недостаточно (Исмаилов, 1974;
Козубов, Муратова, 1986; Михеева и др., 2005; Janusz, 1969).
Род Juniperus насчитывает около 70
видов, распространенных в Северном полушарии от Арктики по всему умеренному
поясу; немногие растут в горах (Центральная Америка, Вест-Индия, Восточная
Африка) (Матюхин, 2007).
Род представлен вечнозелеными однодомными
или двудомными небольшими деревьями или кустарниками, иногда стелющимися.
Характерным признаком растений рода является форма листьев: у молодых растений
листья игловидные, у взрослых растений ряда видов эта форма сохраняется, однако
у некоторых видов взрослые растения имеют листья чешуевидной формы. Почки
голые, без чешуй, иногда окруженные прижатыми и укороченными листьями (J.
communis L.) и только у одного вида (J. drupacea L.) с
многочисленными плотными чешуйками. Для наших видов игловидные листья
отстоящие, линейно-ланцетные, с низбегающим основанием, сверху с устьичной
полоской, нераздельной или раздельной срединной продольной жилкой листья
(Матюхин, 2007). Чешуйчатые листья мелкие, прижатые к побегам, попарно
супротивные или, реже, в трехчленных мутовках (Деревья и кустарники…, 1949;
Исмаилов, 1974; Денисова, Пилипенко, 1978).
Существует несколько классификаций рода Juniperus.
По системе А.Л. Тахтаджяна (1978) род разделяют на три подрода. Подроды Caryocedrus
(1 вид) и Juniperus или Oxycedrus (собственно можжевельник, более
14 видов), обладают игловидными отстоящими листьями, мясистыми шишками,
содержащими по три семени. У представителя подрода Caryocedrus семена
срастаются в единую косточку. Семена у видов второго подрода не сросшиеся. У видов
третьего подрода - Sabina (около 40 видов), иногда выделяемого в особый
род, листья все или частично чешуевидные, прижатые к побегу, мясистые, шишки с
1-6 (до 12) семенами (Денисова, Пилипенко, I 1978; Козубов, Муратова, 1986).
М.И. Исмаилов (1974) в обработке рода Juniperus
предложил подобную же систему, но дополнительно разбил подроды на секции:
.первый подрод Juniperus на 3
секции; Oxycedroides (3-5 видов), Regioides (4-6 видов), Recurvoides
(6-8 видов).
.второй подрод Sabina тоже 3
секции; Polycarpoides (20-25 видов), Virginoides (18-20 видов),
Pseudosabinoides (18-20 видов).
. третий подрод Caryocedrus
монотипен.
В подрод Juniperus входят
можжевельники обыкновенный (Juniperus communis), красный (J.
oxycedrus), прибрежный (J. conferta L.), твердый (J. rigida
L.) и др. К подроду Sabina относятся можжевельники высокий (J. excelsa
Bieb.), казацкий (J. sabina L.), и др. Все виды внутри каждого из
подродов довольно близки между собой как по морфологическим признакам, так и по
особенностям биологии и экологии (Денисова, 1978; Имханицкая, 1990).
Современное распространение, форма и конфигурация ареала J. cоmmunis - результат длительного
процесса эволюции. Изучение эволюционного развития ареалов - сложная задача,
требующая синтеза знаний из различных областей науки.
J. communis встречается в
самых разнообразных экологических условиях, что обусловливает его изменчивость
и разнообразие жизненных форм. Ареал J. cоmmunis
циркумполярный в северном полушарии между 30 и 70° с.ш. (Ареалы деревьев и
кустарников, 1977, Деревья и кустарники СССР, 1949, Коропачинский, 2002).
J. cоmmunis - это
единственный вид рода Juniperus, рапространенный в
Восточном и Западном полушариях. В отличие от других родов J. communis имеет весьма
обширный ареал: Северная и Средняя Европа; европейская часть СНГ от
Мурманска и верховий р. Лозьвы на Урале до Киева, Харькова, Саратова, В Сибири
от 66-70° с.ш. на юг до степной зоны; в Северной Америке от Южной Канады, Новой
Шотландии до британской Колумбии и на юг до Нью-Джерси, Пенсильвании,
Мичигана, Небраски и по Скалистым горам до Нью-Мексико (Матюхин, 2007).
На территории России его ареал охватывает Европейскую часть,
Урал и
Сибирь, в восточной части ареала данный вид изучен слабо. По мнению И.Ю.
Коропачинского и Т.Н. Встовской (2002), на большей части территории Сибири
произрастает близкий вид J. sibirica (Burgsd.) Rydb., очень сходный с J. communis пo морфологическим
признакам. Типичный J. communis найден в
Сибири лишь в Томской обл. и некоторых районах Предуралья. На всей
остальной части Сибири произрастает J. sibirica (Burgsd.) Rydb. (Михеева, Муратова, 2005).
В Татарстане широко распространен подвид J. cоmmunis var. на севере республики - в
Предкамье входит в состав лесных сообществ, встречается и в Предволжье, а также
единичными экземплярами в южных районах РТ. Северные районы РТ, где
распространены хвойные и хвойно-широколиственные леса, является южной частью фитоценотического
ареала J. cоmmunis var. communis. По определению А.П.
Шенникова (1942)
фитоценотнческнй ареал - ареал в условиях конкуренции с другими видами
(Любарский, 1976). В рамках своего фитоценотического ареала J. cоmmunis участвует в
сложении смешанных
лесов в составе подлеска, сохраняется после рубок леса и участвует в составе
кустарниковых зарослей как доминантный вид или совместно с другими видами.
По данным гербария Казанского государственного университета
были выявлены районы и географические точки, где когда-либо были собраны
гербарные образцы J. cоmmunis. Первые сборы датируются 1865 г. В 20 веке J. communis, отмечался в
Зеленодольском, Высокогорском, Верхнеуслонском, Балтасинском, Арском, Агрызском,
Актанышском, Мамадышском, Елабужском, Тюлячинском, Атнинском, Кукморском,
Сабинском, Кайбицком и Апастовском районах (Салахов, 2009).
На сегодняшний день J. communis в южных и восточных
районах РТ представлен единичными экземплярами, например: д. Берлибаш Апастовкого
р-на он встречается в сосновом лесу; близ д. Черки-Гришино Буинского р-на, д. Л.
Моркваши Верхне-Услонского р-на и д. Аккузево Актанышского р-на отмечен в подлеске
липняка разнотравного (Салахов, 2009).
По данным Р.П. Адамский (2003) вид J.
соmmunis подразделяется на несколько подвидов, которые распространены по
ареалу вида неравномерно. В западном полушарии произрастают преимущественно
подвиды J. com. var. J. cpressa Pursh, J. com. var.
hemispherica (C. Prest) Nyman., J. com. var. oblonga (M. Bieb)
Loudon., в восточном полушарии, в основном, встречаются J. com. var.
communis L, J. соm. var saxatilis Pall., в Японии - подвид J. com.
var. japponica (Maxim) E.H. Wilson. Наиболее сложная проблема для
идентификации подвидов - это разделение между подвидами, J.com. var.
communis L. и J. com. var. saxatilis L. Результаты анализа ДНК,
выделенного из растений разных географических точек, (Adams. 2003) показывают
высокую степень сходства между этими подвидами. Морфологические параметры также
очень близки. Подвид J. cоmmunis var. communis чаще всего имеет
древовидную форму (Adams. 2003. Матюхин, 2007), а подвид J. cоmmunis var.
saxatilis, стланиковую форму с побегами, имеющими более широкие хвоинки
(Adams, 2003).
Необходимость ревизии подвидовых таксонов
отмечается не только у можжевельника обыкновенного, а также у групп
близкородственных кавказских (J.excelsa Bieb. J. polycarpos С.
Koch. J. isophvllos C. Koch) и среднеазиатических (J. seravschanica
Кот., J. turkestanika Кот.,) видов (Имханицкая, 1990: Антонаж, 1991).
Неопределенное систематическое положение многих видов вызывает разногласия у
систематиков по поводу их названий в соответствии с ботанической номенклатурой.
J. communis - дерево до 8-12 м
высоты с конусовидной кроной или кустарник яйцевидной или конусовидной формы,
более узкой у мужских экземпляров и с более или менее восходящими или
простертыми, иногда «на вершине» свисающими ветвями у женских экземпляров. Побеги
красновато - бурые. Кора серо-бурая, продольно шелушащаяся. Листья 1 - 1,5 см
длины и 0,7-1,5 мм ширины, сверху неглубокожелобчатые, с одной нераздельной или
иногда до середины раздельной беловатой устъечной полоской, снизу
блестяще-зеленые с тупым килем, плотные, на конце вытянутые, в колючее острие.
Шишкоягоды почти сидячие, 5-9 мм в диаметре, шаровидные; зрелые - черно-синие с
голубым налетом или без него, созревающие на 2-3й год, состоят из 3 или 6
чешуй. В шишке 3 (иногда 1-2) семени, удлиненно - яйцевидных или
яйцевидно-конических, бурых. Древесина ядровая. Заболонь узкая, белая с узкими
годичными слоями. Ядро серовато-коричневое с матовым блеском. Годичные слои
узкие, извилистые, хорошо заметные на всех разрезах. Ранняя часть годичного
слоя резко переходит к поздней, которая слабо развита. Сердцевинные лучи
простым глазом не видны. Смоляных ходов нет. Древесина J. communis имеет
очень прочные твердообразные свойства (Арсентьева, 1998; Матюхин, 2007).
Вследствие широкого распространения в
горах можжевеловые леса имеют большое водоохранное, водорегулирующее и
почвозащитное значение. Подсчитано, что на крутых склонах гор, в тех местах,
где можжевельники встречаются лишь одиночно или мелкими группами, с 1 га
ежегодно смывается 5000 м почвы, а в густых можжевеловых зарослях почвенной
эрозии почти не наблюдается.
Можжевеловые леса и редколесья в зоне
широколиственных лесов J. cоmmunis занимает свободные от конкурентов
участки и образует самостоятельные насаждения, где его конкуренты произрастать
не могут. Такими участками оказываются скалистые или крутые склоны с бедными
почвами, малопригодные для широколиственных деревьев (Каппер, 1954). Мощные
разветвленные корни J. cоmmunis внедряются в расщелины скал, удерживая
растение даже на отвесных скалах.
У экземпляров, произрастающих в
неблагоприятных условиях, главный ствол наклоняясь к земле, раскидывает во все
стороны разросшиеся боковые ветви, которые укореняются. К 30-40 годам жизни
ствол совершенно теряется среди лежачих и приподнимающихся мощных боковых ветвей,
длина которых намного превышает длину головного стебля. К 50-60 годам жизни на
месте одного экземпляра образуется целая группа особей - клон или куртина.
Каждая партикула в клоне живет до 90-100 лет, поэтому возраст клона может
измеряться несколькими столетиями (Исмаилов, 1974).
1.4 Изучение биоморфологических и популяционных
особенностей J. соmmunis в разных зональных условиях
Первые исследования J.communis в
Республике Татарстан проводились Н.В. Салаховым (2009). В ходе этих
исследований были выявлены следующие особенности: В экосистемах республики
Татарстан J.communis широко распространен на севере и северо-западе и
постепенно редеет к югу и юго-востоку. Наиболее благоприятные для вида
фитоценотические условия складываются в подтаежных хвойно-широколиственных
лесах на севере и западе РТ. В пределах Республики Татарстан у J.communis
встречаются различные жизненные формы. В южной тайге наиболее важные факторы -
достаточное освещение и подгон видов - эдификаторов, способствуют формированию
биоморфы типичного одноствольного хвойного дерева. Образование
немногоствольного дерева связано с недостатком освещенности. В условиях
можжевеловых зарослей при максимальной освещенности образуется прямостоячий
аэроксильный кустарник. Жизненная форма стелющийся кустарник - стланец,
преобладает на сильно эродированных склонах водоразделов. На основе
морфологических показателей Н.В. Салаховым (2009) охарактеризованы
онтогенетические состояния J.communis: проростки (р), ювенильные (j),
имматурные (im), виргинильные (v), молодые генеративные (gl), средневозрастные
генеративные (g2), старые генеративные (g3) и сенильные (s) особи. В лесных
фитоценозах базовые спектры, по данным автора, полночленные. В большинстве
случаев максимум приходится на прегенеративные особи - левосторонний
онтогенетический спектр, который свойственен растениям с длительным
онтогенезом. Онтогенетический спектр ценопопуляции можжевеловых зарослей
неполночленен. Максимум приходится на генеративные особи - одновершинный
центрированный онтогенетический спектр (Салахов, 2009).
Н.А. Михеева (2005) занималась изучением
кариологических особенностей J. communis в гидроморфных и суходольных
условиях произрастания. Оказалось, что кариотип можжевельника обыкновенного в
условиях различного гидрологического режима является стабильным при высокой
изменчивости биоморфологических признаков вида. Ксероморфные признаки
фотосинтезирующего аппарата (чешуек) J. communis свидетельствуют об
адаптивной реакции вида к условиям повышенного увлажнения почв. Для расчета
площади поверхности и объема хвои J. communis необходимо вводить
поправочные коэффициенты, поскольку форма поперечного сечения хвои не
соответствует правильным геометрическим фигурам.
Е.А. Тишкина (2009) занималась
определением внутривидовой дифференциации форм и экологической приуроченностью J.
communis на Урале. В заключение выяснили, что фитоценотическим и
экологическим оптимумом местообитания можжевельника обыкновенного на Урале
следует считать ельники и сосняки зеленомошные II-III класса бонитета с
сомкнутостью древесного полога 0,6-0,7. Краевые популяции можжевельника
обыкновенного, как и центральные могут быть центрами образования его
декоративных внутривидовых форм. На охраняемых территориях отмечается
повышенная урожайность можжевельника, чем на территориях, подверженных
антропогенному влиянию. Локальные популяции - самая распространенная форма
существования можжевельника обыкновенного, как в горных экосистемах Урала, так
и лесных ценозах и на открытых пространствах.
Н.В. Герлинг (2010) изучавший
продуктивность и физиологические особенности хвои видов рода Juniperus на
Северо-Востоке европейской части России определил, что продолжительность роста
побегов у можжевельника составляет 90, а хвои - 60 дней. Фаза активного
развития листового аппарата отмечается в третьей декаде июня и сопровождается
утолщением покровных тканей и снижением относительного объема мезофилла.
Сезонные изменения в структуре мезофилла развивающейся хвои можжевельника
обыкновенного сопровождаются увеличением парциального объема межклетников, клеточных
оболочек и вакуолей в полтора, а число хлоропластов и митохондрий в цитоплазме
- два раза. В однолетней хвое в течение вегетационного сезона усиливается
вакуолизация мезофилла. Максимальное развитие гранальной структуры в
хлоропластах и пигментной системы в хвое разного возраста отмечается в начале
августа. Однако, у можжевельника сибирского на Северном Урале снижаются размеры
хвои и усиливается развитие эпидермы и смоляных каналов. В мезофилле хвои этого
вида парциальный объем гиалоплазмы выше, а объемная доля межклетников ниже, чем
у можжевельника обыкновенного. В условиях низкой интенсивности физиологически
активной радиации (ФАР) под пологом древостоя елового леса можжевельник
обыкновенный характеризуется высоким содержанием пигментов в хвое и доли
хлорофиллов в светособирающем комплексе хлоропластов. По мере роста хвои
концентрация пигментов в ней увеличивается от 3.8 до 4.6 мг г-1 сухой массы. В
однолетней хвое с марта по август концентрация пигментов возрастает от 3.9 до
4.9 мг г-1 сухой массы. У можжевельника сибирского в горах Урала содержание
суммы пластидных пигментов и доля хлорофилла в составе светособирающего
комплекса снижается в два раза. Видимый фотосинтез можжевельника обыкновенного
под пологом елового фитоценоза продолжается 13-15 ч в сутки, снижение
интенсивности СО2 - газообмена хвои происходит в дни с жаркой и сухой погодой.
Величина максимальной интенсивности видимого фотосинтеза можжевельника
обыкновенного и можжевельника сибирского под пологом елового фитоценоза одинакова.
У можжевельника сибирского в условиях гор формируется фотосинтетический аппарат
«светового» типа, у можжевельника обыкновенного под пологом елового фитоценоза
фотосинтетический аппарат имеет свойства «теневого» типа.
В степном Башкирском Зауралье, проводились
исследования популяционной структуры можжевельника казацкого на горе Йелбаш
(Путенихин, 2009). При снижении жизненности по виталитету, фенотипическое (и
генетическое) разнообразие ценопопуляций выражено достаточно существенно. Это
означает, что имеющийся высокий внутрипопуляционный потенциал не реализуется
достаточно полно в засушливых степных условиях обитания, но потенциальные
адаптационные возможности велики. Это подтверждается хорошим жизненным
состоянием особей, присутствием индивидуумов разных возрастных категорий,
сбалансированной половой структурой, представительством различных биоморф. ЦП
можжевельника казацкого на горе Йелбаш представляют один из уникальных
форпостов вида в степном юго-восточном Башкирском Зауралье.
2. Физико-географическое описание мест
исследования
Физико-географические характеристики Волжско-Вятского
возвышенно - равнинного региона темнохвойно-широколиственных
неморально-травяных лесов с фрагментами южнотаежных елово - пихтовых и сосново
- еловых зеленомошных лесов, где проходили наши исследования описан по сводке
«Сосудистые растения» (2000).
Республика Татарстан (РТ), занимающая площадь 67838 км2
расположена на юго-восточной части Русской равнины. Характерным для территории
республики является наличие границ крупных единиц районирования практически по
всем основным ландшафтообразующим компонентам.
В широтном направлении здесь проходит изолиния
гидротермического коэффициента теплого времени года (май-сентябрь), равного
единице, определяющая деление территории по климатическим условиям на лесную
достаточно увлажненную зону, и лесостепную зону, испытывающую дефицит
увлажнения.
В соответствии с почвенно-географическим районированием по
территории проходит граница между бореальным и суббореальным почвенным поясом.
Сюда заходят Среднерусская провинция дерново - подзолистых среднегумусированных
почв, Вятко - Камская провинция дерново - подзолистая высокогумусированных
почв, Среднерусская провинция серых лесных почв, Прикамская провинция серых
лесных почв, Среднерусская лесостепная провинция оподзоленных, выщелоченных и
типичных среднегумусных и тучных мощных черноземов и серых лесных почв и
Заволжская лесостепная провинция оподзоленных, выщелоченных и типичных тучных
среднемощных черноземов.
Волжско-Вятский возвышенно-равнинный регион темнохвойно-широко
- лиственных несморально травяных лесов с фрагментами южнотаежных елово -
пихтовых и сосново - еловых зеленомошных лесов. Регион расположен к северу от
Камы, занимает водоразделы Волги, Вятки и Камы и входит в состав возвышенности
Западного Предкамья, нередко именуемой Кукморской. Максимальные отметки Вятско
- Волжского водораздела близ правобережья Вятки 200 - 250 м. Восточные склоны
плато круто обрываются к реке Вятке и сильно расчленены притоками реки и
оврагами.
Климат региона умеренно континентальный с холодной зимой и
теплым летом. Среднегодовая температура +2,30С, средняя температура
января -13,90С (минимальная до -52,0 0С), июня +19,0 0С.
Средняя высота снежного покрова 30 - 40 см. Атмосферных осадков в среднем
выпадает 403 - 509 мм (максимум осадков в Западно-Казанском ландшафтном районе
- более 600 мм). Гидротермический коэффициент составляет 1,3-1,4. (Бакин и др.,
2000).
Растительный покров территории весьма не однороден. В
условиях возвышенно-равнинных водоразделов и приводораздельных склонов
коренными являются подтаежные елово - липово - дубовые (Picea fennica, Tilia cordata, Quercus robur) c пихтой (Abies sibirica) кислично (Oxalis acetosella) - неморальные (Aegopodium podagraria, Mercutialis perenis, Carex pilosa) леса с таежными видами
(Linnea borealis, Pyrola rotundifolia, Vaccinium myrtillus), по крутым осыпающимся
склонам овражно - балочных систем и водораздельным склонам на обнажениях
коренных пермских пород - с участием кальцефитных видов (Cephalanthera rubra, Cypripedium calceolus, Asplenium ruta - muraria).
В Илетско-Ашитском районе отмечаются небольшие фрагменты
южно-таежных елово-пихтовых лесов. Среди них: пихтовые с елью и сосной (Pinus
sylvestris) кислично-мшистые (Hylocomium sp., Dicranum
scoparium, Pleurozium schreberi) с участием Carex rhizirina и костяники (Rubus
saxatilis) на коричнево-серой дерново-подзолистой почве элювиальных
мергелей водораздельных и склоново-водораздельных поверхностей; еловые с пихтой
и липой неморально-кислично-мшистые на водораздельных склонах северной
экспозиции; пихтовые с елью и липой лещиновые (Corylus avellana) на перегнойно-карбонатной
почве верхних частей склонов глубоких логов водораздельно-склоновых
поверхностей.
Наряду с хвойно-широколиственными лесами в регионе на
возвышенных участках водоразделов и речных террас, на материковых склонах южной
экспозиции небольшими фрагментами отмечаются широколиственные - дубовые и
дубово-липовые насаждения с елью: дубово-липовые с елью
пролесниково-корневищноосоково-снытевые (Mercurialis perennis, Carex rhizina, Aegopodium podagraria) на суглинках плато;
дубовые с елью и липой лещиново-снытевые с бореальными видами на плоских
выровненных поверхностях водоразделов; дубовые с липой, кленом (Acer platanoides) и вязами (Ulmus laevis, U. glabra) лещиново-снытевые на
серых лесных суглинистых почвах высоких останцовых поверхностей в долине Волги;
дубовые волосистоосоково (Carex pilosa) - снытевые по склонам
южной, юго-западной и юго-восточной экспозиции с участием лесостепных видов (Pulmonaria mollis, Euphorbia semivillosa); дубовые с липой и
сосной снытевые на материковых склонах долин малых рек.
3. Объекты и методы исследования
.1 Объекты исследований
Объектами наших исследований являются ЦП J.communis, приуроченные к разным
ландшафтно-природным районам РТ и различным эколого-фитоценотическим условиям
(табл. 2). Исследования проводились в следующих ландшафтно-природных районах:
Западно-Казанский террасово-долинный район (Зеленодольский р-н РТ), Казанский
эрозионно-расчленённый район (Арский, Балтасинский р-ны РТ),
Правобережно-Вятский эрозионно-равнинный район (Кукморский р-н РТ) (Бакин и
др., 2000).
Таблица. 2. Эколого-географическая характеристика ценопопуляций J.communis
|
Местоположение
|
Фитоценоз
|
Освещенность
|
Тип
природопользования
|
|
Сосняки с елью
(ЦП 1; 2; 3)
|
РТ.
Зеленодольский район. Раифа кв. 132 около болота «гнилое»
|
Сосняк с
березой (единично ель) бруснично - ландышево-мшистый
|
70%
|
Заповедник
|
|
|
|
|
|
|
|
|
РТ.
Зеленодольский район. Раифа кв. 132/135 вдоль просеки
|
Сосняк с елью и
березой чернично - мшистый
|
50%
|
Заповедник
|
|
|
Арский район.
Сев. часть дер. Сурнары.
|
Сосняк с елью
ландышево-мшистый
|
50%
|
Не охраняемая,
рядом с деревней
|
|
Сосняки (ЦП
4; 5; 6; 7)
|
г. Казань.
Кировский р-н. лесопарк Лебяжье рядом с оз Б. Голубое
|
Сосняк
орляково-разнотравный
|
80%
|
Рекреационная
зона г. Казани
|
|
|
г. Казань.
Кировский р-н. лесопарк Лебяжье около оз Лебьяже в сторону п. Юдино.
|
Сосняк
вейниковый
|
|
Рекреационная
зона г. Казани
|
|
|
РТ. Арский
район. Сурнарское уч. лесничество. Кв. 126.
|
Сосняк
вейниково зеленомошный
|
|
Лесничество
|
|
|
РТ.
Балтасинский райаон. Лесной массив» Шубан», ООПТ «Карадуван»
|
Сосняк
|
|
ООПТ,
рекреационная зона
|
|
Можжевеловые
заросли (ЦП8; 9; 10)
|
РТ.
Балтасинский район. Северная часть р.ц. Балтаси.
|
Можжевеловые
заросли
|
100%
|
Пригородная
зона р.ц. Балтаси.
|
|
|
РТ. Балтасинский
район. Сев.-западная часть деревни Куюк.
|
Можжевеловые
заросли
|
|
Придорожная
зона. склон.
|
|
|
РТ. Кукморский
район. около деревни Яныль.
|
Можжевеловые
заросли
|
|
Придорожная
зона. склон
|
3.2 Методы исследования
Геоботанический, на основе общепринятых
методов (Воронов, 1976); 2. Популяционный метод (Ценопопуляции растений, 1976,
1977); 3. биоморфологический; 4. для пространственного анализа применялась
среда R.
Для оценки сходства видового состава двух сообществ
использовали коэффициент общности Серенсена-Чекановского (КО):
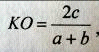
где а - число видов в первом сообществе, b - число видов во
втором сообществе, с - число общих видов для первого и второго сообществ
(Рогова Т.В. и др., 2000).
На основе морфометрических показателей разных особей J. communis (Салахов, 2009)
определялся возрастной, половой и биоморфологический состав, а также изучалось
пространственное распределение разных групп особей (онтогенетических, половых,
жизненных форм). Далее составлялась электронная база J. communis, с учетом перечисленных
групп и разных видов деревьев, произрастающих на пробной площадке.
Метод построения К-функции был предложен Рипли для
характеристики точечных паттернов (point pattern) индивидов растений и их
групп, у которых определены местоположения (Ripley, 1976; Ripley, 1977). Его преимущество
в том, что этот метод позволяет оценить распределение особей (или др. групп) на
разных масштабах.
Далее для анализа внутрипопуляционного распределения в
пространстве особей (или других исследуемых групп) нами использовалась К(r)
функция Рипли, реализованная в языке статистического программирования R. К(r) показывает
среднее число особей, находящихся на расстоянии r от любой случайно
выбранной особи. Для вычисления значения функции вокруг каждой особи условно
очерчивают круг радиуса r, центр которого задается координатами
местоположения особи, затем подсчитывают число особей, находящихся внутри
окружностей, и делят полученную сумму на произведение площади картированной
площадки на плотность особей, находящихся в центрах условных окружностей и
плотность особей, попавших в окружности.
В случае полной пространственной случайности (complete spatial randomness, CSR), когда особи
распределены в пространстве случайным образом и не наблюдается их значительных
скоплений или стремления к регулярному расположению в пространстве, значение
функции Рипли равно площади круга с радиусом, равным r: КCSR(r)=pr2. По отклонению значений К(r),
вычисленных на исследуемой точечной мозаике, от значений КCSR(r), ожидаемых при
пространственной случайности, судят о пространственной кластеризации или
регулярности наблюдаемого расположения особей для данного радиуса. Превышение
функции К(r) над КCSR(r) свидетельствует о том,
что на данном расстоянии r или на чуть меньшем расстоянии встречается больше
особей, чем могло бы быть при их случайном, независимом пространственном
распределении, что свидетельствует о наличии скоплений радиусом r. Если кривая функции К(r)
проходит ниже значений КCSR(r), то это означает, что
приблизительно на данном расстоянии r было встречено меньше
особей, чем могло бы быть при их случайном распределении, что свидетельствует о
регулярности расположения точек, или их разрежении. Расстояния r, на которых наблюдаются
эти отклонения, называются размером (масштабом) наблюдаемого вида
неоднородности.
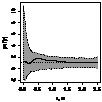
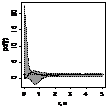
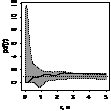
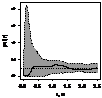
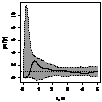
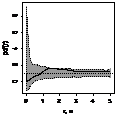
Графически функцию Рипли представляют в виде функции L(r) - r, т.к. в случае полной пространственной
случайности LCSR(r) - r=0
при любых расстояниях r (рис. 1), при этом L(r) вычисляется как: 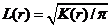 .
.
Для определения значимости отклонений поведения L(r) функции Рипли от LCSR(r) функции
Рипли, соответствующей случайному пространственному распределению особей,
использован метод симуляций Монте-Карло. Данный метод уже описан нами.
При расчёте расстояния между особями разных классов (виды,
онтогенетические группы), распределение характеризуется либо «агрегированием» -
формированием скопления, либо «отталкиванием» - под зоной отталкивания мы
понимали область наименьшей плотности особей, обусловленной разграничением
территории распространения разных возрастных групп или разных видов и
формированием равномерной структуры. Если наблюдаемая функция Рипли выходит из
коридоров доверия функции Рипли, который соответствует случайному распределению
особей, значит, наблюдается значительное / значимое отклонение от случайного
распределения на этих расстояниях (отложенных по оси абсцисс).
По вертикали откладывается L(r) - r, по горизонтали - расстояние, для которого рассчитывалось значение
функции Рипли, (рис. 4).
Все картированные особи можно разделить на несколько классов -
например, по онтогенетическому состоянию, индексу жизненности или на основании
любой другой информации. Для анализа пространственных закономерностей между
расположением двух классов особей используется кросс-функция Рипли. Анализ
точечных мозаик был проведён с помощью пакета SPATSTAT.
Таким образом, мы можем проанализировать «притягивание» или
«отталкивание» между деревьями разных возрастных групп и разных пород. Нами
были построены графики функции J-cross. Выбираются две группы. Функция
Пунктирные линии - коридоры доверия. Если наблюдаемая функция (сплошная черная
линия) выходит из коридоров доверия, значит, наблюдается значительное /
значимое отклонение от случайности распределения на малых расстояниях. Если
значение функции <1 то наблюдается кластеризация (агрегирование), если >1
то наблюдается регулярное распределение, или прореживание на данном расстоянии.
Парно-корреляционная функция чётче показывает размер скоплений,
расстояний между скоплениями и вновь повторяющихся формирований скоплений.
Функция Рипли в этом плане чаще всего показывает увеличение плотности и
формирование очень крупных скоплений (Wiegand, 2007).
Для выявления количественной оценки использования каждого
экологического фактора тем или иным видом предложено понятие «экологической
валентности», а для комплекса факторов - «толерантности», или «бионтности»
видов. Экологическую валентность и толерантность характеризуют с применением
экологических шкал, которые отражают балловые оценки оптимума существования
видов растений.
Потенциальную экологическую валентность (PEV) вида
рассматривают как меру приспособленности популяций конкретного вида к изменению
только одного экологического фактора. Потенциальная экологическая валентность
рассчитывается как отношение числа ступеней конкретной шкалы, занятой данным
видом, к общей протяженности шкалы в ступенях. Величина PEV равна доле
диапазона ступеней конкретного вида от всей шкалы:
 ,
,
где Аmax и Аmin - максимальные и минимальные значения
ступеней шкалы, занятых отдельным видом; n - общее число ступеней в шкале; 1 - добавляется как 1-е деление
шкалы, с которого по данному фактору начинается диапазон вида.
При проведении исследований конкретных ценопопуляций (ЦП) или
сообществ реализованную экологическую валентность (REV) также можно представить
в виде следующей формулы:
 ,
,
где Аmax и Аmin - максимальные и минимальные значения
ступеней шкалы, занятые конкретными ценопопуляциями на шкале; n - общее число ступеней в шкале; 0,01 - добавляется как 1-е
деление шкалы, с которого встречаются изученные ценопопуляций.
Эффективность освоения экологического пространства вида
конкретными ценопопуляциями по каждому фактору оценивается при помощи
коэффициента экологической эффективности (K.ec.eff.), который представлен
следующей формулой:
 ,
,
где PEV - потенциальная экологическая валентность, REV -
реализованная экологическая валентность.
Соотношение REV/PEV определяет степень использования
экологических потенций вида изученными ценопопуляциями и дают возможность
произвести анализ экологического предпочтения видов в сообществах разных
растительных зон, а также вскрыть экологические механизмы устойчивости
популяций растений (Жукова и др., 2010).
4. Результаты и обсуждение исследований
.1 Особенности эколого-фитоценотических условий
местообитаний ЦП J. communis
На основе геоботанических описаний мест произрастания ЦП J.
communis была определена оценка сходства видового состава растительных
сообществ (табл. 3).
Таблица 3. Коэффициент общности сообществ по Сёрсену-Чекановскому
|
Сосняки с елью
|
Сосняки
|
Можжевеловые
заросли
|
|
|
|
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
8
|
9
|
10
|
|
1
|
|
0.44
|
0.34
|
0.54
|
0.32
|
0.44
|
0.15
|
0.43
|
0.1
|
0.14
|
|
2
|
0.44
|
|
0.93
|
0.52
|
0.2
|
0.32
|
0.16
|
0.55
|
0.1
|
0.17
|
|
3
|
0.34
|
0.93
|
|
0.55
|
0.2
|
0.3
|
0.01
|
0.53
|
0.07
|
0.28
|
|
4
|
0.54
|
0.52
|
0.55
|
|
0.29
|
0.39
|
0.04
|
0.61
|
0.16
|
0.31
|
|
5
|
0.32
|
0.2
|
0.2
|
0.29
|
|
0.19
|
0.33
|
0.21
|
0.06
|
0.18
|
|
6
|
0.44
|
0.32
|
0.3
|
0.39
|
0.19
|
|
0.05
|
0.32
|
0.07
|
0.27
|
|
7
|
0.15
|
0.16
|
0.01
|
0.04
|
0.33
|
0.05
|
|
0.06
|
0.01
|
0.06
|
|
8
|
0.43
|
0.55
|
0.53
|
0.61
|
0.21
|
0.32
|
0.06
|
|
0.42
|
0.88
|
|
9
|
0.1
|
0.1
|
0.07
|
0.16
|
0.06
|
0.07
|
0.01
|
0.42
|
|
0.56
|
|
10
|
0.14
|
0.17
|
0.28
|
0.31
|
0.18
|
0.27
|
0.06
|
0.88
|
0.56
|
|
На основе коэффициента общности, как сосняки с елью, так и
сосняки свежие либо сухие более или менее сходны между собой по видовому
составу (50 до 90%). И только можжевеловые заросли на остепненных склонах имеют
низкий коэффициента общности (КО) с лесными фитоценозами.
Экологический анализ флористического состава исследуемых
участков, проведенный по шкалам Д.Н. Цыганова, позволил выявить экологические
условия местообитаний, определить степень их благоприятности для произрастания
J. communis и рассчитать индексы экологической валентности и толерантности (табл. 4.), а также коэффициента
экологической эффективности, которые характеризуют приспособленность конкретной
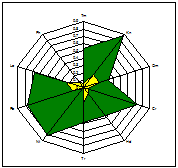
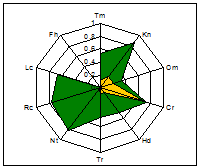
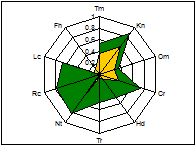
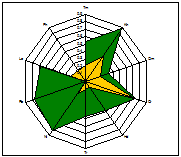
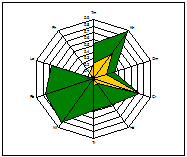
ценопопуляции к условиям данного местообитания. На основании расчетов индексов PEV, REV и коэффициента
экологической эффективности построены полиграммы, отражающие диапазон
потенциальной и реализованной экологической валентности J.communis в разных
эколого-фитоценотических условиях. Полиграммы демонстрируют различие между
потенциальной экологической валентностью (внешняя граница полиграммы), и
границей экологического пространства (реализованной экологической
валентностью).
Таблица 4. Экологическая валентность J.communis и экологические условия
ценопопуляции
|
Сосняки с елью
|
|
 ЦП 1 ЦП 1
|
 ЦП 2 ЦП 2
|
 ЦП 3 ЦП 3
|
|
Малонарушенные
сосняки
|
|
 ЦП 6 ЦП 6
|
 ЦП 7 ЦП 7
|
|
Рекреационно
нарушенные сосняки
|
|
 ЦП 4 ЦП 4
|
 ЦП 5 ЦП 5
|
|
Можжевеловые
заросли
|
|
 ЦП 8 ЦП 8
|
 ЦП 9 ЦП 9
|
 ЦП 10 ЦП 10
|
|
|
|
|
Условные обозначения шкал: Tm - термоклиматическая, Kn - континентальности
климата, Om - аридности-гумидности, Cr - криоклиматическая, Hd - увлажнения почв, Tr - солевого режима, Rc - кислотности почв, Nt - богатства почв азотом,
Lc -
освещенности-затемнения. зеленая - pev; желтая - rev.
Самые неблагоприятные условия для произрастания можжевельника
складываются в сосняке орляково-ландышево-разнотавном на Лебяжьем, находящимся
вблизи основной зоны отдыха горожан на Оз. Глубокое. В целом местообитания
благоприятны для произрастания J.communis, по всем основным шкалам
4.2 Популяционная организация J. communis, на основе численности,
плотности, возрастной, половой и биоморфологической структуры
Для анализа популяционной организации J. communis проводились исследования
численности, плотности, возрастной структуры и их динамики в различных
фитоценозах, что позволило выявить особенности популяционной структуры в разных
условиях произрастания и природопользования.
J. communis - двудомное растение, у
которого в генеративной стадии по репродуктивным органам хорошо определяются
половые группы. Ряд авторов считают, что у растений большее количество мужских
особей развиваются в неблагоприятных условиях, в нормальных условиях половая
структура должна быть уравновешена. В сосново-еловых сообществах отмечается
более иди менее уравновешенная половая структура (рис. 7.). Напротив, большее
количество женских особей встречаются на склоновых участках, что обусловлено,
как нам кажется, низкой конкуренцией с другими видами растений (биотический
фактор) и для закрепления на склоновых участках, по-видимому, увеличивается
активность семенного размножения. С точки зрения стратегии можжевельник
патиент, поэтому освобождение пространства с ресурсной базой от конкуренции
является важным экологическим условием для благоприятного развития его
популяций. Кроме того, густые можжевеловые заросли снижают влияние почвенной
эрозии на склоновых эродированных либо осыпных участках.
В условиях можжевеловых зарослей вид представлен кустарниками
и стланцами, их доля зависит от прогреваемости, экспозиции и угла наклона
склоновой поверхности. Оказалось, что на более крутых склонах, юго-восточной,
восточной и северных экспозиций преобладают стелющиеся формы - стланцы, что
обусловлено сильной ветровой эрозией (почвы и снега). Часто в неблагоприятных
условиях - на бедных щебенистых почвах или при сильном затенении главный
стебель полегает на землю и со временем сгнивает, на его месте образуется целая
группа вегетативных клонов и формируется куртина. Этот процесс, как отмечают
ряд авторов, не подавленные роста побега первого порядка, а своеобразные
жизненные формы - кустарниковые либо стелющиеся (стланцы).
В благоприятных лесных сообществах древесные и кустарниковые
жизненные формы уравновешены. И только в условиях сухих, рекреационно
нарушенных сосняков, у можжевельника преобладает кустарниковая жизненная форма,
что является механизмом приспособления и устойчивости вида, обусловленное
интенсивностью вегетативного размножения в результате партикуляции (разделения)
старых кустов. Поливариантность жизненных форм в неблагоприятных
эколого-фитоценотических условиях способствует выживанию вида, его ЦП-ий и
является проявлением толерантности и приспособленности.
Во всех четырех типах местообитаний в ЦП можжевельника
преобладают генеративные особи, доля прегенеративных незначительная (около
10-20%), что связано с низким семенным возобновлением, особенно за последние 3
года засухи (2009-2011 гг.). Полночленная возрастная структура ЦП J. cоmmunis отмечалась лишь на
территории заповедника и лесных массивов, удаленных от городов и мало
подверженных антропогенной нагрузке в сосняках и сосняках с елью зеленомошных (рис.
9.).
Отмечено, что для древесных видов характерно чаще семенное
возобновление, поэтому в их популяциях доля прегенеративных особей должна быть
высокой. Однако, на границе лесной зоны, и особенно в нарушенных местообитаниях
- рекреационная зона г. Казани либо эродированные и осыпные склоны,
преобладающей жизненной формой является кустарник, преимущественно
размножающийся вегетативным способом (укореняющиеся ветви, распад крупных
особей на партикулы). также преобладание генеративных особей в возрастной
структуре обусловлено типом онтогенеза - генеративные особи живут значительно
дольше. Поэтому их доля варьирует от 60 до 70%, а доля постгенеративных
(сенильных и субсенильных) составляет 10-18%. Хотя доля проростков и ювенильных
особей незначительна, наличие практически всех онтогенетических групп говорит о
поддержание онтогенеза, в результате активизации вегетативного размножения.
Выводы: 1. В пограничных условиях ареала (южная граница
тайги), где сказываются неблагоприятные климатические условия (малоснежные
зимы, летние засухи) и фитоценотические условия - неморализация
хвойно-широколиственных лесов и псевдосемиаридизации (ложное или антропогенное
остепнение) ландшафтов на севере РТ, обуславливают плохое семенное размножение
можжевельника.
. Поэтому для стабильности популяционной организации на юге
лесной зоны в ЦП J. cоmmunis встречаются различные жизненные формы
более подходящие для эколого-фитоценотических условий местообитаний и
определенный тип возрастной структуры - центрированный, с преобладанием
генеративных особей, как более устойчивых и преимущественно вегетативное
размножение.
. В оптимальных условиях зеленомошных сосняков с елью и
свежих сосняков доля женских особей составляет 55%, а мужских 45% - половой
спектр уравновешен; в пессимальных условиях антропогенно деградированных
сосняков начинают преобладать мужские особи, напротив, на сухих остепненных
склонах - женские.
4.3 Особенности пространственной структуры ЦП J.communis
в разных условиях произрастания
Для анализа пространственно-онтогенетической структуры была
создана электронная база с учетом разных групп и произрастающих совместно видов
деревьев. Первоначально были построены карты схемы и дана динамика абсолютной
численности по онтогенетическим группам особей.
В пределах ареала популяции вида, как и особи в пределах
ценопопуляций, распределены, как правило, неравномерно, что обусловлено рядом
причин. С помощью пространственной организации реализуются основные
популяционные адаптации; с пространственной структурой связана ее устойчивость
и продуктивность, и, наконец, пространственное распределение способствует
достижению оптимальной плотности населения на определенной площади (Наумов,
1965). С одной стороны, определенный тип пространственного размещения вида
позволяет наиболее эффективно использовать природные ресурсы местообитания,
конкурировать с другими видам, произрастающими в фитоценозе, с другой - снижать
внутривидовую конкуренцию, обеспечивая нормальное взаимодействие особей внутри
популяции для поддержания устойчивости в постоянно меняющихся экологических
условиях.
Принято считать, что случайное и регулярное распределение
растений встречается редко. Даже при искусственных регулярных посадках,
например в лесных культурах, регулярное размещение растений со временем
меняется из-за неравномерной элиминации растений, в силу их разной
жизнеспособности, конкуренции друг с другом и влиянии внешних причин (Морозов, 1928;
Ипатов, Кирикова, 1997).
Пространственный анализ первоначально был сделан для разных
половых групп J. communis во всех ЦП на исследуемых
участках (табл. 5.). Как правило, женские и мужские особи относительно друг
друга распределяются случайным образом. Это дает возможность крупным
генеративным особям эффективно использовать природный ресурс, снижая
конкуренцию друг с другом, укрепляя тем, самым свои позиции по отношению к
другим видам в сообществе. В оптимальных условиях (сосняков с елью и свежих сосняков)
женские особи стремятся разграничить пространство между собой - в основном
преобладает случайное их распределение. Напротив, мужские особи чаще формируют
локусы повышенной плотности - скопления, радиусом 0,8 - 1,5 м, что отмечалось,
как на охраняемой, так и рекреационно-нарушенной территории.
Изучая пространственное распределение разных жизненных форм
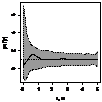
можжевельника было отмечено следующее: 1. Кустарниковые формы во всех
ценопопуляциях образуют скопления, радиусом 0,4-1,2 м либо 1,5- 5 м, что
обусловлено укоренением нижних ветвей, образование кустарниковых форм при
ухудшении эколого-фитоценотических условий и формированием куртины. 2. Исконные
формы можжевельника древесные, напротив, распределяются случайным образом, это
способствует снижению внутривидовой конкуренции; 3. При взаимном распределение
кустарников, деревьев и немногоствольных деревьев только в сосняках с елью
образуются небольшие скопления, а в свежих сосняках распределение носит
случайный характер (табл. 6.).
При анализе пространственного распределения возрастных групп
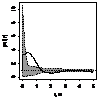
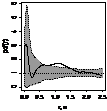
было выявлено, что генеративные особи, как правило, стремятся к агрегированию,
радиус скоплений варьирует от 1 до 2,5 м (табл. 7.). Плотность прегенеративных
особей небольшая, поэтому распределение носит случайный характер, что
обусловлено недостатком семенного размножения и преобладанием вегетативного.
При взаимном распределении прегенеративных и генеративных особей первоначально
образуется «зона отталкивания» - радиусом 0,75-1,5 м, что снижает внутривидовую
конкуренцию и способствует росту и развитию молодых особей можжевельника в
«светлых окнах», затем начинают образовываться скопления радиусом 1,5 - 3 м.
Таблица.5. Особенности пространственного
распределения половых групп
|
|
Сосняки с
елью
|
Сухие
сосняки
|
Свежие
сосняки
|
Можжевеловые
заросли
|
|
Карта-
схема
|

|
|
|
|
|
pcf_все
|
|

|

|
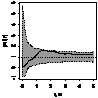
|
|
pcf_ женские
|
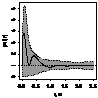
|
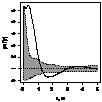
|
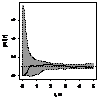
|
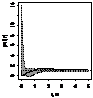
|
|
pcf_жен и муж
|

|
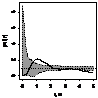
|
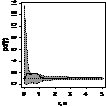
|
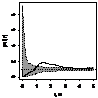
|
|
pcf_ мужские
|
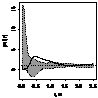
|

|
|
|
Примечание:

 - женские особи,
- женские особи,

 - мужские особи.
- мужские особи.
Таблица.6. Особенности пространственного распределения
жизненных форм.
|
|
Сосняки с
елью
|
Сухие
сосняки
|
Свежие
сосняки
|
Можжев.
заросли
|
|
Карта-
схема
|
|
|
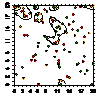
|
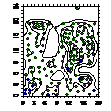
|
|
pcf_все
|

|
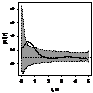
|
|
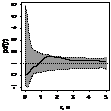
|
|
pcf_ кустарники
|
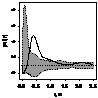
|

|
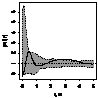
|
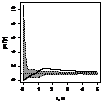
|
|
pcf_деревья
|
|

|
|
|
|
pcf_ немногоствольные деревья
|

|
|

|
|
|
pcf_сланцы
|
|
|
|

|
|
|
|
|
|
|
Примечание:
- кустарники,
- дерево,

 - немногоствольное дерево.
- немногоствольное дерево.
Таблица.7. Особенности пространственного распределения
возрастных групп.
|
|
Сосняки с
елью
|
Сухие
сосняки
|
Свежие
сосняки
|
Можжев.
заросли
|
|
Карта-
схема
|

|
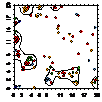
|

|
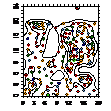
|
|
pcf_все
|
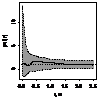
|
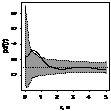
|

|
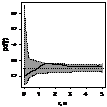
|
|
pcf_ генеративные
|
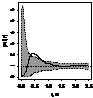
|
|

|
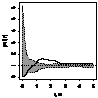
|
|
pcf_генеративные и прегенеративные
|
|
|
|
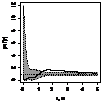
|
|
pcf_ прегенеративные
|
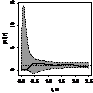
|
|
|
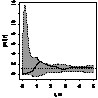
|
При рассмотрении пространственного распределения J. cоmmunis с другими деревьями выявлено,
что растение не образует скоплений с елью. Напротив, со сосной и березой, как
близких по световым требованиям видов, в сухих сосняках образуются небольшие
скопления, что обусловлено одинаковым отношение к свету, так как сосна и береза
встречались в таком же ярусе что и можжевельник, в сосняках с елью и в
можжевеловых зарослях они распределены случайным образом (табл. 8.).
Таблица.8. Особенности пространственного распределения с другими
деревьями.
|
|
Сосняки с
елью
|
Сухие
сосняки
|
Свежие
сосняки
|
Можжев. заросли
|
|
Карта-
схема
|
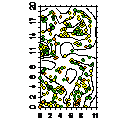
|
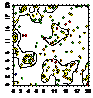
|
|

|
|
pcf_все
|

|

|

|
|
|
pcf ель финская
|

|
|
|
|
|
pcf сосна обыкновенная
|

|
|
|
|
|
pcf_ береза повислая
|
|

|
|
|
.4 Динамика пространственной структуры J.
communis на разных участках мезорельефа
Синузия (греч. synusia - совместное
пребывание, сообщество) - часть фитоценоза (биоценоза), совокупность
(объединение, группа) особей одного вида (синузия первого порядка) или сходных
видов (синузии второго и третьего порядков). Или пространственно и экологически
обособленная часть растительного сообщества, состоящая из видов растений одной
или нескольких экологически близких жизненных форм. По Гамсу (Gams, 1918)
выделяют синузии нескольких порядков: Синузия 1 порядка образована растениями
одного вида и, следовательно, одной жизненной формы, например синузия кислицы в
ельнике. Синузия 2 порядка образована растениями разных видов, но одной
жизненной формы, например синузия брусники и черники в сосняке зеленомошном.
Синузия 3 порядка образована растениями разных видов и разных, но близких
жизненных форм (синузия марьянника лугового и черники в ельнике). Синузия 4
порядка образована растениями разных видов и разных жизненных форм,
объединяющая разные ярусы. Синузию 4 порядка также называют микрогруппировкой
или микроценозом, в основе ее лежит 1 или несколько ценоячеек. Примером такой
синузии является группировка таких видов как зеленые мхи, кислица, черника под
пологом молодой ели.
Динамика популяции - это разнонаправленные
или однонаправленные изменения в структуре (например, численности и плотности -
компенсаторных признаков) либо (полового и возрастного состава - консервативные
признаки) популяции в результате действия абиотических (не зависящих от
численности и плотности самой популяции) и биотических (зависящих от
численности и плотности популяции) факторов (Динамика …., 1985).
Пространственная структура популяции - это
особенность размещения особей популяции в пространстве. Она зависит как от
свойств мест обитания, так и от биологических особенностей вида. Она может
изменяться во времени, зависит от сезона года, от численности популяции и т.д.
Пространственное размещение особей может быть: равномерным, случайным и
групповым.
Половая структура популяции - это
количественное соотношение особей по полу. Соотношение полов популяции
устанавливается по генетическим законам, а затем на него влияет среда. У большинства
видов пол будущей особи определяется в момент оплодотворения в результате
перекомбинации половых хромосом. Соотношение особей по полу, особенно
размножающихся самок в популяции, имеет большое значение для дальнейшего роста
ее численности. На соотношении полов в популяции влияют условия среды. У
некоторых видов пол изначально определяется не генетическими, а экологическими
факторами.
Возрастная структура популяции -
определяет все возрастные группы особей, в том числе всех стадий и фаз развития
организма. В популяции животных выделяют три экологических возраста:
предрепродуктивный (довоспроизводящий), репродуктивный (воспроизводящий), и
пострепродуктивный (послевоспроизводящий). Длительность каждого возраста по
отношению к общей продолжительности жизни не одинакова у разных видов. В
жизненном цикле растений выделяют около десяти возрастных состояний.
Биоморфологическая структура популяции -
это своеобразная внешняя форма организмов, обусловленная биологией развития и
внутренней структурой их органов, формируется в определенных
почвенно-климатических условиях, как приспособление жизни к этим условиям, т.е.
это форма организмов, приспособившихся к своей среде обитания под длительным
влиянием комплекса факторов. Изучение биоморфологической структуры, их динамики
исключительно важно для познания закономерностей приспособления популяций и
организмов к тем или иным условиям среды. Сообщества растений и животных
устойчивее и продуктивнее, если они состоят из видов разных жизненных форм.
Такое сообщество полнее использует ресурсы среды и имеет более разнообразные
внутренние связи. Без изучения биоморфологической структуры невозможно изучить
ценотическую структуру - один из основных аспектов структурно-функциональной
организации биогеоценоза (Серебряков.1964).
Изучение особенностей динамики J. communis в пространстве проводили
в трех разных местообитаниях (Лесной массив «Шубан», Лесопарк «Лебяжье», ВКГПБЗ
Раифский участок), у которых выделяются ценопопуляционные локусы, приуроченные
к разным, синузиям, участкам различного освещения, влажности или рельефа.
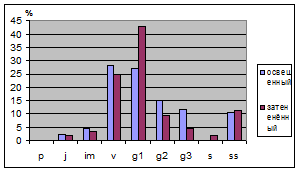
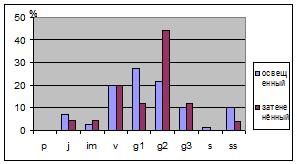
При изучении распределения возрастных
групп во всех трех площадках заметно, что в ненарушенных условиях сильного
различия нет, но в нарушенных преобладают генеративные особи (табл. 9). На
освещенных участках увеличивается доля прегенеративных, так как для их развития
требуется свет. Также видно, что группа имматурных особей отмечаються в
условиях затенения, потому что это удаленный участок от дороги.
Таблица.9. Динамика онтогенетических групп
в разных ценопопуляционных локусах
|
РТ, г. Казань.
Кировский район, лесопарк «Лебяжье», рядом с оз. Б. Голубым.
|

|
|
РТ,
Зеленодольский район, Раифский участок ВКГЗ, кв. 132. около болота «гнилое».
|
|
|
РТ. Балтасинский
район. лесной массив Шубан. ООПТ «Карадуван».
|
|
В распределении жизненных форм можно
сказать, что кустарниковые формы больше всего встречаются в затененных условиях
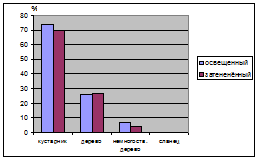
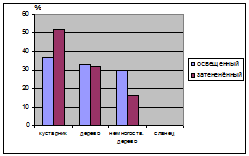
и в сильно нарушенных условиях (Лебяжье) (табл. 10.). Древесные формы
равномерно распределяются как в освещенных так и в затененных условиях, а вот
немногоствольные деревья предпочитают более освещенные участки. Судя по таким
распределениям можно сказать о том, что в освещенных и затененных условиях наиболее
приспособленными являются кустарниковые и древесные формы J. communis.
Таблица.10. Динамика биоморфологических
групп в разных ценопопуляционных локусах
|
РТ, г. Казань.
Кировский район, лесопарк «Лебяжье», рядом с оз. Б. Голубым.
|
|
|
РТ,
Зеленодольский район, Раифский участок ВКГЗ, кв. 132. около болота «гнилое».
|
|
|
РТ.
Балтасинский район. лесной массив Шубан. ООПТ «Карадуван».
|

|
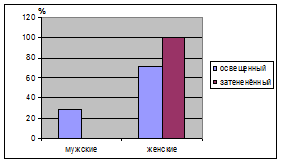
При распределении половых групп с
различными условиями освещенности видно, что и в освещенных и в затененных
условиях преимущественно встречаются женские особи (табл. 11.). Такое
распределением мужских и женских особей в популяции необходима для поддержания
потомства J. communis в этих условиях.
Таблица.11. Динамика половых групп в
разных ценопопуляционных локусах
|
РТ, г. Казань.
Кировский район, лесопарк «Лебяжье», рядом с оз. Б. Голубым.
|

|
|
РТ,
Зеленодольский район, Раифский участок ВКГЗ, кв. 132. около болота «гнилое».
|

|
|
РТ.
Балтасинский район. лесной массив Шубан. ООПТ «Карадуван».
|
|
Выводы
1) В сосново-еловых лесах наблюдается равновесие между
женскими и мужскими особями. Напротив большее количество женских особей
встречаются на склоновых участках, так как здесь низкая конкуренция с другими
видами (биотический фактор) и для закрепления на склоновых участках,
по-видимому, увеличивается активность семенного размножения.
) В условия можжевеловых зарослей вид представлен
кустарниками, в сосняках с елью древесных и кустарниковых жизненных форм
уравновешено. Уже в условиях сухих сосняков у можжевельника, при ухудшении
абиотических условий и рекреационной нагрузке отмечается преобладание
кустарниковой жизненной формы, что является механизмом устойчивости вида и
способствует интенсивности вегетативного размножения в результате партикуляции
(разделения) старых кустов. У условиях крутых склонов начинают развиваться
новые жизненные формы - сланцы.
) Во всех трех типах местообитаний можжевельник представлен
больше всего генеративными особями, прегенеративных - встречается мало, что
связано с низким семенным возобновлением (за последние 3 года засухи). Наиболее
близкое к полночленному распределению встречается в заповедных или в лесных
массивах мало подверженных к антропогенной нагрузке. В целом во всех
экологических условиях доля прегенеративных особей очень незначительна и
составляет около 10 -12%, а доля генеративных особей наоборот выше в 2-4 раза
) Кустарниковые формы во всех ценопопуляциях формируют
скопления, радиусом 0,4-1,2 м и 1,5- 5 м, что обусловлено биологическим
процессом, так как при ухудшении условий происходит процесс укоренения нижних
ветвей и образование кустарниковых формы, которые образуют куртины. Древесные,
напротив, распределяются случайным образом, это способствует снижению
внутривидовой конкуренции.
) Женские и мужские особи относительно друг друга
распределяются случайным образом. В оптимальных условиях (сосняков с елью и
свежих сосняков) женские особи стремятся разграничить пространство. Напротив,
для взаиморасположения мужских особей чаще характерно формирование скоплений
радиусом 0,8 - 1,5 м, исключение составляют только не нарушенные сообщества
свежих сосняков.
) При взаимном распределении прегенеративных и генеративных
особей первоначально образуется «зона отталкивания» - радиусом 0,75-1,5 м, что
снижает внутривидовую конкуренцию и способствует росту и развитию молодых
особей можжевельника в «светлых окнах», затем начинают образовываться скопления
радиусом 1,5 - 3 м.
) J. cоmmunis с другими деревьями
выявлено, что растение не образует скоплений с елью. Напротив, со сосной и
березой, как близких по в сухих сосняках образуют небольшие скопления, по
видимому это связано с тем, что у них одинаковое отношение к свету, так как
сосна и береза встречались в таком же ярусе что и можжевельник, в остальных же
фитоценозах они распределены случайным образом.
Библиографический список литературы
1.
Алешина Е.Н. Влияние способа культивирования на рост каллусной ткани Juniperus sibirica В. // Тез. докл.
междунар. конф. Звенигород, 8-12 сент. 2008 г. Москва: ИД ФБК - ПРЕСС, 2008. С.
6-7.
.
Ареалы деревьев и кустарников СССР. - Л.: Наука. 1977. - Т.1. 164 с.
.
Арсентьева Т.В. Сравнительно - анатомическое изучение древесины J.communis L. (Cupressaceae) в связи с изменением
жизненных форм // Проблемы ботаники на рубеже 20-21 вв.: Тез. докл. - СПб. -
1998. - Т.1. - С. 6.
.
Аши М. Можжевельник в лесах Верхневолжья: Флора и растительность южной тайги, -
Калинин, 1989 - С. 59 - 63.
.
Бакин О.В., Рогова Т.В., Ситников А.П. Сосудистые растения Татарстана, -
Казань: Изд-во Казан. ун-та. 2000. С. 12 - 18.
.
Буторина А.К., Мурая Л.С., Исаков Ю.Н. Влияние условий произрастания;
экологическая генетика растений и животных, - Кишинев: 1981 г. - Ч. 1. - С.
11-12.
.
Ваганов Е.А., Шашхин А.В. Рост и структура годичных колец хвойных. -
Новосибирск: Наука, 2000. - С. 112.
.
Василевич И.В. Статистические методы в геоботанике, - Л.: Наука. 1969. -232 с.
.
Василевич И.В. Количественные методы изучения структуры растительности // Итоги
науки и техники. - М: ВИНИТИ. 1972. -140 с.
.
Василевич И.В. Выявление границ в растительном покрове // Бюл. МОИП. Отд. биол.
- 1975. - Т.80. - Вып. 3. - С. 94 - 104.
.
Воронцова Л.И., Гатцук Л.Е., Егорова В.Н. Ценопопуляция растений, - М.: Наука,
1976. - С. 217.
.
Восточноевропейские леса / Под ред. О.В. Смирновой. М.: Наука, 2004. 364 с.
.
Герлинг.Н.В., Морфолого - анотомические особенности фотосинтети-ческого
аппарата видов рода Juniperus на севере: дис… кандидата биоло-гических наук.:
защищена 17.03.2010. Институт биологии Коми научного центра Уральского
отделения РАН.
.
Гиляров А.М. Популяционная экология, - М.: изд. МГУ. 1990. С 21-23.
.
Горчаковский П.Л. Растения европейских широколиственных на восточном пределе их
ареала, - Свердловск: Труды института экологии растений и животных УФ АН СССР,
1968. Вып.59. - С. 207.
.
Грейг - Смит. П. Количественная экология растений, - М.: Мир, 1967. - С. 358.
.
Гулидова И.Е. Определение поверхности хвои у ели. - М.: Сообщение института
леса АН СССР, 1959. Вып.11. - С. 49 - 50.
.
Дендрология Узбекистана./ под ред. Ф.Н. Русанова. - Ташкент Таш, 1965. Т.1. С.
112.
.
Денисова Г.А., Пилипенко Ф.С. Семейство кипарисовые (Cupressaceae) // Жизнь растений, -
М.: Просвещение, 1978. Т.4. - С. 393 - 403.
.
Денисова Г.И., Якушина И.И., Денисова Г.М. Физиология роста и развития
растений, - М.: 1985. С. 116.
.
Деревья и кустарники СССР. Дикорастущие, культивируемые и перспективные для
интродукций / Ред. С.Я. Соколова, Б.К. Шишкин: Изд Акад. наук СССР в Лгр.,
1949. Т.1. - С. 57 - 62.
.
Деревья и кустарники СССР. // Н.А Бородина, В.И Некрасов, Н.С Некрасова, И.П
Петрова, Л.С Плотникова, Н.Г Смирнова, - М.: Мысль, 1966. - С. 241.
.
Деревья и кустарники. Голосеменные. Справочник / под ред. Н.Н. Гордиенко. -
Киев: Наука думка, 1971. - С. 161.
.
Джанаева В.М. Определитель семейства можжевельниковых, - Фрунзе: Илим, 1969.
-93 с.
.
Жизнеспособность популяций. Природоохранные аспекты. М.: Мир, 1989. 224 с.
.
Имханицкая И.Н. Критическая заметка о кавказских видах секций Juniperus рода Juniperus L. // Новости систематики
высших растений, - Л.: Наука, 1990. №27. - С. 5 -16.
.
Исмаилов М.И. О системе рода Juniperus L. // Вопросы экологий и
географии растений, - Душанбе.:1974. - С. 138 - 168.
.
Камышев Н.В. Введение в фитоценологии, - Воронеж.: Изд-во Воронежского
Университета, 1986. - С. 202.
.
Капер О.Г. Хвойные породы, - М, - Л.: Гослесбумиздат, 1954. - С. 187.
.
Киричок Е.И. Онтогенез, возрастная и пространственная структура можжевельник
высокого (Juniperus excelsa, Cupressaceae) в редколесьях юго - восточного Крыма // Проблема ботан. на
рубеже 20 - 21 вв. Тез. докл. предствал. 2 (10) Съезду Рус. ботан. о-ва, Санкт
- Петербург 26 - 29 мая, 1998 г. Т.1. - Спб., 1998. - С. 262.
.
Князева С.Г. Биологические особенности прорастания семян можжевельника
сибирского (Burgsd // Плодоводство, семеноводство, интродукция древесных растений:
Материалы 2-й Всероссийской научно - практической конференции с международным
участием, Красноярск, 8- 9 дек., 1999 г. - Красноярск, 1999.С. 43-44
.
Князева С.Г. Изменчивость и морфоструктура природной популяции можжевельника
сибирского: автореф. дис. …канд. биол. наук.: защищена 12.04.2000: утв
24.09.2000, Изд - во Красноярского гос. ин-та, 2000. 21 с.
.
Князева С.Г. половая изменчивость можжевельника обыкновенного // Лесоведение. -
2002. - №6. - С. 73 - 75.
.
Князева С.Г. Опыт применения многомерных методов для определения
таксономического положения Juniperus sibirica (Cupressaceae) // Ботан. журнал. 2004
г. - №2.-С. 236 - 244.
.
Князева С.Г. Изменчивость морфологических признаков сибирских видов
можжевельника // Лесоведение - 2007. - №1. - С. 65 - 69.
.
Козубов Г.М., Муратова Е.Н. Современные голосеменные, - Л.: Наука, 1986. С.
192.
.
Коропачинекий И.Ю., Встовская Т.Н. Древесные растения Азиатской России, -
Новосибирск: СО РАН, филиал Гео, 2002 г. С. 707.
.
Коропачинский И.Ю. Древесные растения Сибири, - Новосибирск: Наука., Сиб.
Отделение АН СССР, 1983. С. 189.
.
Корчагин А.А. Внутривидовой (популяционный) состав растительных сообществ //
Полевая геоботаника. - М, Ленинград: Наука, 1964 г. - С. 63 -130.
.
Красная Книга республики Татарстан, Казань.: Изд-во. «Идель - пресс». 2006 г.
732 с.
.
Крылов Н.П. Флора Западной Сибири. Руководство по определению западносибирских
растении, - Томск: ТГУ, 1955. С. 138.
.
Крюссман Г. Хвойные породы. М., Лесная промышленность, 1986. С. 46 - 53.
.
Мамаев С.А. Виды хвойных на Урале и их использование в озеленении, -
Свердловск: Урал. НЦ АН СССР. 1983. 109 с.
.
Матюхин Д.Л., Громадин А.В. Дендрология. 2-е издание, - М.:Академия, 2007.С. 45
- 54. С. 98 -102.
.
Михеева Н.А., Муратова Е.Н. Кариологическое изучение двух популяции
можжевельника обыкновенного Juniperus communis L. в Западной Сибири.
Цитология, 2005; Т.47. N.8.
.
Мухамедшин К.Д. Арча.-М.: Лесная промышленность. 1980, С. 92.
.
Мухамедшин К.Д., Саргбаев С.К. Чемпионы долголетия. - Алма - Ата: Кайнар, 1988
г. С. 168.
.
Определитель растений Татарской АССР, - Казань: Изд - во. Казанского
Университета, 1979. С. 163.
.
Определитель сосудистых растений среднего Урала // под ред. Горчаковский П.Л,
М.:Наука, 1994. 525 с.
.
Путенихин В.П., Фарукшина Г.Г. - Ценопопуляция можжевельника Казацкого
(Juniperus Sabina L.). // Степи Северной Евразии: Материалы V Международного
симпозиума, 2009 г.
.
Салахов Н.В., Эколого-фитоценотическая приуроченность, жизненные формы и
популяционная биология Juniperus communis L. в Республике Татарстан: дис…
кандидата биологических наук.: защищена 21.12. 2009 г. Моск. пед. гос. ун-т.
52.
Смирнова О.В., Заугольнова Л.Б., Торопова Н.Ф., Фаликов Л.Д. Критерии выделения
возрастных состоянии и особенностей хода онтогенеза у растении разных биоморф
ценопопуляции растении. Основные понятия и структура. М.: Наука, 1976. - 215 с.
.
Смирнова О.В., Зауголънова Л.Б., Попадюк Р.В. Популяционная концепция в
биоценологии // Журн. общ. биологии. 1993. Т. 54, №4. С. 438-48.
.
Смирнова О.В., Калякин В.Н., Турубанова С.А. и др. Современная зональность
Восточной Европы как результат преобразования позднеплейстоценового комплекса
ключевых видов // Мамонт и его окружение: 200 лет изучения. М.: Геос, 2001. С.
200-208.
.
Солбриг О., Солбриг Д. Популяционная биология и эволюция. М., Мир., 1982. С.
488.
.
Тимофеев - Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции.
- М.: Наука, 1973. - 276 с.
.
Тишкина Е.А., Закономерности распространения, формовое разнообразие и
экологическая приуроченность Juniperus communis L. на Урале: диссертация
кандидата сельскохозяйственных наук.: защищена 03.03. 2009 г. Ур. гос.
лесотехн. ун-т.
.
Уранов А.А., Смирнова О.В. Классификация и основные черты развития популяции
многолетних растений. // Бюллетень МОИП, 1969. Т.74. №1.
.
Яблоков А.В. Популяционная биология: Учеб. пособие для биол. спец.
вузов.
- М.: Высш. шк., 1987. - 303 с.
60. Adams R.P., Pandey R.N. Analysis of Juniperus communis and is
varieties based on DNA fingerprinting //Biochem. Syst. Ecol. 2003. V. 31. P.
1271-1278.
. Nagano K., Nakamura M., Toda Y. Chromosomes studieson
Cupressaceae. VIII. Karyotype of Juniperus // Proc. Fac. Agr. Kyushu Tokai
Univ. 1991. V 10. P 75-87.
. Janusz A. Brzozowski, Rina S. Cohen: On Decompositions of
Regular Events. 1969. J. ACM 16 (1): P. 132 -144.
. Ripley, B.D. 1976. The second-order analysis of stationary point
processes. Journal of Applied Probability 13: pp.255-266.
. The mosaic cycle concept of ecosystem. H. Remmert-ed. Ecologecal
studies. // Analysis and syntesis.1991. Vol. 85. 168 p.
. Watt A.S. Pattern and Process in plant communities // J. Ecol.
1947. Vol. 35 P. 1-22.
. Whittaker R.H. A consideration of climax theory: The climax as
population patterns // Ecol. Monogr. 1953. Vol. 23. P. 41-78.
. Whittaker R.H., Lewin A.S. The role of mosaic phenomena in
natural communities 11 Theor. Pop. Biol. 1977. Vol. 12, №2. P. 117-139.
. Wiegand, T., and K.A. Moloney. 2004. Rings, circles, and null -
models for point pattern analysis in ecology. Oikos 104: pp.209 - 229.
. Wiegand T., Gunatilleke S., Gunatilleke N. Species Associations
in a Heterogeneous Sri Lankan Dipterocarp Forest // The American Naturalist.,
2007, №4, P. 77 - 95.