Влажность как экологический фактор
КУРСОВАЯ
РАБОТА
Влажность
как экологический фактор
вода экологический гумидный
ВВЕДЕНИЕ
вода экологический
Часто встречается перевод термина ≪экология≫
как учения о доме, жилище. Это не точно. Древние греки понимали этот термин
значительно шире. Экосом они называли любое место пребывания человека: и
хороший пляж, где люди собирались для купания, и горное пастбище, где пастухи
пасли овец.
В середине XX в. экологию стали понимать как
науку об экосистемах и биосфере. Начало такому пониманию положили работы В.И.
Вернадского, В.В. Докучаева, Ю.П. Одума, А.Дж. Тенсли, Н.В.
Тимофеева-Ресовского и других известных ученых. В результате стало ясно, что
современная биосфера, среда обитания всех живых организмов, является продуктом
их жизнедеятельности: неустанного воспроизведения, метаболизма (обмена веществ)
и посмертного разложения мириадов живых существ. Почвенная, водная, наземная,
воздушная среда жизни есть результат постоянного взаимодействия и
взаимопроникновения живого и неживого веществ. Ни один вид живых организмов не
может существовать среди себе подобных. Жизнь возможна только в сообществах
(биоценозах) и в строго определенной совокупности условий, характеризующей
место их обитания (биотоп). Единство биотопа и биоценоза - основная концепция современной
экологии, концепция экосистемы [Николайкин, Николайкина, Мелехова, 2004, с. 6].
Целью данной курсовой работы является изучение
влажности как экологического фактора. Для достижения поставленной цели
предполагается решить комплекс взаимосвязанных задач: рассмотреть свойства воды
и ее биологическую роль, дать характеристику воды как экологического фактора,
определить аридные и гумидные условия, исследовать водный баланс организмов,
источники получения воды, рассмотреть водный дефицит, а также изучить потери
воды и солевой баланс, определить эколого-физиологические показатели,
характеризующие водный режим организмов, рассмотреть воду как среду обитания
организмов, изучить экологию водных организмов, а также хроматическую
адаптацию, дать характеристику редким водным животным Красной книги
Краснодарского края, а также редким видам гигрофитов Западного Кавказа.
1. Свойства воды и ее биологическая роль
Вода − уникальное вещество и все её
аномальные свойства: высокая температура кипения, значительная растворяющая и
диссоциирующая способность, малая теплопроводность, высокая теплота испарения и
другие обусловлены строением её молекулы и пространственной структурой. У
отдельно взятой молекулы воды есть качество, которое проявляется только в
присутствии других молекул: способность образовывать водородные мостики между
атомами кислорода двух оказавшихся рядом молекул, так, что атом водорода
располагается на отрезке, соединяющем атомы кислорода. Свойство образовывать
такие мостики обусловлено наличием особого межмолекулярного взаимодействия, в
котором существенную роль играет атом водорода. Это взаимодействие называется
водородной связью. Каждая из присоединённых к данной молекул воды сама способна
к присоединению дальнейших молекул. Этот процесс можно называть "полимеризацией".
Если только одна из двух возможных связей участвует в присоединении следующей
молекулы, а другая остаётся вакантной, то "полимеризация" приведёт к
образованию либо зигзагообразной цепи, либо замкнутого кольца.
Вода −превосходный растворитель для
полярных веществ. К ним относятся ионные соединения, такие как соли, у которых
заряженные частицы (ионы) диссоциируют в воде, когда вещество растворяется, а
также некоторые неионные соединения, например сахара и простые спирты, в
молекуле которых присутствуют заряженные (полярные) группы (-OH).Присущие воде
свойства растворителя означают также, что вода служит средой для транспорта
различных веществ. Эту роль она выполняет в крови, в лимфатической и
экскреторных системах, в пищеварительном тракте и во флоэме и ксилеме растений.
Удельной теплоёмкостью воды называют количество
теплоты в джоулях, которое необходимо, чтобы поднять температуру 1 кг воды на
1° C. Вода обладает большой теплоёмкостью (4,184 Дж/г). Это значит, что
существенное увеличение тепловой энергии вызывает лишь сравнительно небольшое
повышение её температуры. Объясняется такое явление тем, что значительная часть
этой энергии расходуется на разрыв водородных связей, ограничивающих
подвижность молекул воды. Большая теплоёмкость воды сводит к минимуму
происходящие в ней температурные изменения. Благодаря этому биохимические
процессы протекают в меньшем интервале температур, с более постоянной скоростью
и опасность нарушения этих процессов от резких отклонений температуры грозит им
не столь сильно. Вода служит для многих клеток и организмов средой обитания,
для которой характерно довольно значительное постоянство условий [Коробкин,
2000, с.52−60].
Скрытая теплота испарения есть мера количества
тепловой энергии, которую необходимо сообщить жидкости для её перехода в пар,
то есть для преодоления сил молекулярного сцепления в жидкости. Испарение воды
требует довольно значительных количеств энергии (2494 Дж/г). Это объясняется
существованием водородных связей между молекулами воды. Именно в силу этого
температура кипения воды - вещества со столь малыми молекулами - необычно
высока.
Скрытая теплота плавления есть мера тепловой
энергии, необходимой для расплавления твёрдого вещества (льда). Воде для
плавления (таяния) необходимо сравнительно большое количество энергии.
Справедливо и обратное: при замерзании вода должна отдать большое количество
тепловой энергии. Это уменьшает вероятность замерзания содержимого клеток и
окружающей их жидкости. Кристаллы льда особенно губительны для живого, когда
они образуются внутри клеток.
Плотность воды (максимальна при +4° С) от +4 до
0° С понижается, поэтому лёд легче воды и в воде не тонет. Вода - единственное
вещество, обладающее в жидком состоянии большей плотностью, чем в твёрдом, так
как структура льда более рыхлая, чем структура жидкой воды.
Когезия - это сцепление молекул физического тела
друг с другом под действием сил притяжения. На поверхности жидкости существует
поверхностное натяжение - результат действующих между молекулами сил когезии,
направленных внутрь. Благодаря поверхностному натяжению жидкость стремится
принять такую форму, чтобы площадь её поверхности была минимальной (в идеале -
форму шара). Значительная когезия, характерная для молекул воды, играет важную
роль в живых клетках, а также при движении воды по сосудам ксилемы в растениях.
Многие мелкие организмы извлекают для себя пользу из поверхностного натяжения:
оно позволяет им удерживаться на воде или скользить по её поверхности[Дедю,
1990].
Биологическое значение воды определяется и тем,
что она представляет собой один из необходимых метаболитов, то есть участвует в
метаболических реакциях. Вода используется, например, в качестве источника
водорода в процессе фотосинтеза, а также участвует в реакциях гидролиза.
2 Характеристика воды как экологического
фактора. Аридные и гумидные условия
.1 Характеристика воды как экологического
фактора
Тело растения на 50-90% состоит из воды.
Особенно богата водой цитоплазма (85-90%), много ее и в органеллах клетки.
Весьма богаты водой сочные плоды, мягкая листва, корни, однако семена, особенно
маслянистые, могут содержать незначительные количества воды. Вода имеет
первостепенное значение в жизни растений. Все биохимические процессы растения
протекают в водной среде. Растворяя почти все питательные вещества вода служит
и транспортным средством для них.
Вода играет непосредственную роль в обеспечении
роста растений, являющегося функцией растяжения клеток. Тургорное давление,
производимое изнутри на молодые оболочки клеток водой, вызывает их растяжение,
пока они еще не эластичны. Это способствует увеличению размера клетки. Поэтому
сухость замедляет рост, так как клетка полностью не растягивается. С
уменьшением пластичности оболочки (при отложении целлюлозы) полного увеличения
размера клетки не происходит даже при достаточном обеспечении растения водой.
Днем в период недостатка воды стебли растений имеют меньший диаметр, чем ночью,
что обусловлено разницей в тургоре[Березина, 2009,с.36−39].
Но для растения важно не только количество воды,
поступающее в его распоряжение, но также и ее состояние в атмосфере и в почве.
Очень велика в жизни растений и для закономерностей формирования растительного
покрова роль относительной влажности, воздуха (процентное отношение реального
давления водяного пара к давлению насыщенного пара при той же температуре). Она
показывает долю (процент) от того максимального количества влаги, которое могло
бы содержаться при полном насыщении данного объема воздуха при данной
температуре. Более теплый воздух, естественно, может содержать влаги больше,
чем холодный. При охлаждении воздуха относительная влажность его увеличивается,
и когда достигает 100% (так называемая точка росы), начинается конденсация
паров в виде капель (рис. 1).
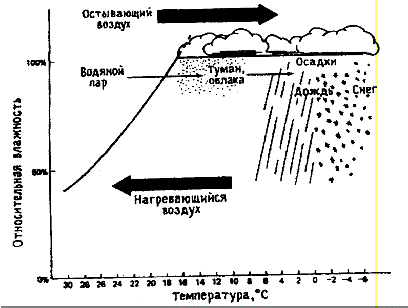
Рисунок 1 - Влияние температуры на относительную
влажность воздуха[по ГорышинойТ., 1979]
Относительной влажность обычно измеряют
психрометром, по разности температур, показываемых сухим и смоченным
термометрами прибора. Применяют также гигрометры и гигрографы, в которых
использован волос, имеющий свойство сокращаться или удлиняться при изменении
влажности. В отличие от дождя, количество которого можно измерить дождемером,
или снега, количество которого определяют растаиванием определенного его
объема, росу и туман измерить дождемером невозможно. Но роса и туман имеют
также важное значение для растений и растительности. Подсчеты показали, что
выпадение осадков в виде росы может достигать иногда 10−20% от годовой
суммы осадков в данном месте. Однако количество конденсирующейся при выпадении
росы влаги в течение суток невелико. Имеются данные, что количество росы,
выпадающее за одну ночь, как правило, не превышает 0,2 мм. Поэтому, например, в
пустынях, где малое количество осадков, юго-западной части Африки, где, кстати,
очень часты туманы, они почти не имеют значения в жизни растений, поскольку
могут смочить слой песка толщиной не более 1−2 см, а такой слой полностью
пересыхает днем. Но в засушливых местах и в пустынях роса может поддерживать
водный режим листьев, и, кроме того, имеет довольно большое значение для
существования низших растений (лишайников и водорослей), покрывающих иногда
значительным слоем поверхность песка, почвы или скал. Определенную роль играет
туман как источник увлажнения листьев и почвы в поясе так называемых туманных
лесов во многих горных системах мира, особенно в Новой Зеландии, в Северной и
Южной Америке и др. Очень сильное образование росы наблюдается во влажных
тропиках. Но в этих районах так часты осадки, что добавление влажности за счет
росы почти не имеет значения, разве что для многочисленных эпифитов[Горышина,
1979, с. 104−107].
.2 Аридные и гумидные условия
Характерными аридными условиями являются сухой
климат с высокими температурами воздуха, испытывающими большие суточные
колебания, и малым количеством атмосферных осадков (100−150 мм/год) или
полным их отсутствием. Аридами называют фазы иссушения климата в зонах пустынь
и полупустынь, внеледниковых областей, примерно соответствующие межледниковьям
областям, подвергавшихся оледенению во время антропогенового периода.
Вся получаемая влага быстро испаряется. Реки,
протекающие через пустыню из соседних более влажных областей, здесь мелеют и
часто заканчиваются бессточными котловинами с солёными озёрами. Обнажённая
земная поверхность испытывает резкие колебания температуры в течение суток,
из-за чего даже плотные горные породы разрушаются и превращаются в песок. Ветер
беспрепятственно переносит массы сухого песка, создавая волнистый рельеф
песчаных барханов и дюн.
Аридный климат в своих наиболее ярких формах характерен
для тропических и субтропических широт (Сахара, пустыни Аравийского
полуострова, Австралии). В более высоких широтах аридный климат связан или с
защитным действием горных хребтов, препятствующих приносу влаги с океана
(пустыни Северной и Южной Америки), или с удалённостью от океанов (пустыни
Центральной и Средней Азии).
К гумидным условиям относятся области с
избыточным увлажнением, при котором количество атмосферных осадков больше, чем
может испариться и просочиться в почвогрунты. Это формирует обильный
поверхностный сток ручьёв и рек, что способствует развитию эрозионных форм
рельефа, густой гидрографической сети и процветанию влаголюбивых форм
растительности. Различают два типа гумидного климата:
.Полярный тип гумидного климата (наблюдается в субарктическом
и умеренном географическом поясах) характерен для зон с многолетнемерзлыми
грунтами, которые тормозят грунтовое питание рек и усиливают сток атмосферных
осадков;
.Фреатический, или тропический тип гумидного
климата (наблюдается в экваториальном поясе) с частичным просачиванием осадков
в почву и наличием грунтового питания[Культиасов, 1982, с. 48−50].
3. Водный баланс организмов. Источник получения
воды. Водный дефицит. Потери воды и солевой баланс
.1 Водный баланс организмов
Все животные могут выдерживать определенные
отклонения от нормального содержания воды в теле (обычно около 10%). Чтобы не
допустить опасные для жизни отклонения содержания воды, многим из них
приходится вырабатывать соответствующие адаптации. Приведем несколько примеров:
. у жаб на брюхе находятся особые
трубчатые железы, служащие не для выделения веществ, а для впитывания воды из
влажной почвы;
. те вещества, в составе которых
выводятся продукты расщепления азотсодержащих соединений, отличаются у разных
групп животных, те из них, которым важнее всего экономить воду, выводят азот в
форме мочевой кислоты;
. по мере развития яиц змей и ящериц их
масса увеличивается, так как они получают из окружающей среды дополнительную
воду, необходимую для их развития;
. некоторые насекомые способны впитывать
воду непосредственно из влажного воздуха через свои трахеи;
. если потерпевшие кораблекрушение люди
пьют морскую воду, они только усиливают обезвоживание своего организма, так как
концентрация солей в ней выше концентрации солей в моче; морские черепахи, в
отличие от людей, могут пить морскую воду благодаря тому, что имеют находящиеся
в углах глаз солевые железы, выделяющие концентрированный раствор соли; у
альбатросов и других трубконосых птиц аналогичную функцию выполняют трубчатые железы
у основания клюва;
. высокая концентрация молока у
китообразных и ластоногих, которое способно плавать по поверхности воды, как
суфле, уменьшает потерю воды организмом кормящей матери.
.2 Источник получения воды
Растения потребляют воду в основном в жидком
состоянии. В умеренной и холодных зонах огромное косвенное влияние на растения
оказывает вода в твердой фазе.
Снег помимо большого его значения для теплового
режима местообитания играет немалую роль в весенний период, когда осадков
обычно мало и тающий снег пополняет водные запасы почвы, как раз тогда, когда
растущим растениям требуется повышенное количество влаги.
Лед оказывает на растение большей частью вредное
влияние, а при возникновении в межклетниках причиняет механические повреждения.
Зимой в случае образования слоя льда на поверхности почвы могут погибнуть
культивируемые и дикорастущие растения. Ожеледь, т. е. накопление льда на
органах растений на поверхности почвы при внезапных потеплениях после дождя,
часто вызывает механические повреждения растений.
Изморозь или рыхлый лед на ветвях деревьев и
листьях трав во время туманов, в некоторой степени способствует накоплению
воды, при таянии часть ее может попасть в почву, часть испаряется в атмосферу.
Подобное же значение имеет иней. Град обычно является нежелательным фактором,
поскольку часто приводит к механическим повреждениям. В вегетационный период
влага из атмосферы поступает в основном в жидкой фазе. Дожди, их количество
сроки, частота выпадения, интенсивность, а также связь их с другими факторами -
температурой, ветром и т. д. − имеют огромное значение. Количество
осадков принято выражать толщиной слоя (в мм), который образуется на
поверхности при их выпадении. Один миллиметр осадков соответствует выпадению 1
л воды на 1 м2[Культиасов, 1982, с. 91−93].
.3 Водный дефицит
Растения испытывают водный дефицит, когда
скорость транспирации превосходит скорость поглощения воды корневой системой.
Ниже показана интенсивность транспирации растений в дневные часы (по данным
разных авторов, дан диапазон наиболее встречающихся величин) на рисунке 2.Такая
ситуация возникает не только при засухе, но и в условиях почвенного засоления,
а также при низких температурах. В двух последних случаях дегидратация является
компонентом стрессов «неводной» природы и развивается в растениях, когда
количество воды в почве чаще всего не ограничено, однако эта вода недоступна
для растения. Например, при почвенном засолении вода не поглощается корневой
системой из-за высокого осмотического давления (низкого водного потенциала)
почвенного раствора.

Рисунок 2 - Диапазон наиболее часто
встречающихся величин интенсивности транспирации растений в дневные часы [по
Горышиной Т., 1979]
Снижение содержания воды в клетках при водном
дефиците и сопутствующее обезвоживанию увеличение концентрации ионов в
цитоплазме вызывают различного рода нарушения в структуре и функциях
биополимеров, в частности происходит денатурация белков и подавляется их
ферментативная активность, изменяется структура липидного бислоя мембран и
нарушается их целостность. Деструктивные изменения в мембранах в свою очередь
приводят к нарушению внутриклеточнойкомпартментации веществ и подавлению
электрогенеза. На клеточном уровне водный дефицит выражается в потере тургора.
На уровне целого растения водный дефицит часто проявляется в обращении
градиента водного потенциала в системе почва -корень - побег и ингибировании
роста. Способность растения адекватно отвечать на водный дефицит и выживать в
условиях водного дефицита зависит от эффективности защитных механизмов
растения. Некоторые ответные реакции растения на водный дефицит могут
наблюдаться уже через несколько секунд после начала действия стрессора
[Горышина, 2009, с. 114−119]
.4 Потери воды
В условиях полевой культуры нижние листья
окружены преимущественно влажным воздухом в результате транспирации других
листьев. Только верхние листья соприкасаются непосредственно с более сухой
атмосферой. Кроме того, листья, расположенные ниже верхнего слоя, настолько
ограничивают движение воздуха, что между растениями он становится насыщенным
влагой. Поэтому нижние листья обладают значительно более низкой интенсивностью
транспирации. Молекулы водяных паров диффундируют из насыщенного воздушного
слоя в посеве в сухой воздух над посевом, и при относительно ровной поверхности
насаждения (как, например, в посевах злаков) эффективной транспирирующей
поверхностью становится, по существу, поверхность посева, а не всех отдельных
листьев. Тогда безразлично, сколько растений в посеве или сколько листьев на
растении; если листовой полог покрывает всю почву, потеря воды будет
определяться площадью посева. Такой посев теряет относительно много воды - в
дневное время примерно столько же, сколько испаряет открытая водная
поверхность, такая, как озеро. Поскольку ночью большинство растений закрывает
свои устьица, ограничивая таким образом транспирацию, суточная потеря воды
посевом несколько ниже испарения с поверхности открытого водоема. Так как
отдача воды каждой культурой составляет обычно определенную долю испарения с
открытой водной поверхности, ее можно рассчитать по этому испарению или по
количеству солнечного тепла, которое может расходоваться на испарение. Такие
данные сейчас обычно можно получить от метеостанций.
В зависимости от биологии данной культуры потери
влаги могут варьировать от очень малой величины до количества, превышающего
испарение с водной поверхности. Голая поверхность почвы быстро высыхает,
ограничивая тем самым потерю воды из нижележащих слоев. Вот почему для
сохранения воды в районах с ограниченным количеством осадков и без орошения
почву оставляют под паром. Таким образом в почве можно сохранять большую часть
годовых осадков. Когда растения начинают расти, корни могут доставать воду из
нижележащих слоев почвы. Потери воды затем возрастают до тех пор, пока посев не
закроет полностью почву и не сформирует транспирирующую поверхность.
Транспирация у рядковой культуры или плодовых деревьев может превышать потерю
воды ровным посевом, так как они имеют большую транспирируюшую поверхность, чем
площадь земли, на которой растут. Кроме того, между рядками может дуть ветер,
перемешивая сухой воздух с влажным. Поэтому норма орошения для рядковых посевов
может быть больше, чем для сплошных, и она зависит в основном от общей
поверхности посевах [Культиасов, 1982, с. 57−59].
3.5 Солевой баланс
Высокие концентрации NaCl и других солей в
оросительной воде повреждают растения, так как осмотические эффекты тормозят
поглощение воды, а натрий ингибирует ферменты в цитоплазме. Поэтому при
накоплении соли растения становятся чахлыми и в конце концов перестают расти.
Поскольку в условиях сухого климата фактически вся поливная вода испаряется,
засоленность почвы увеличивается несмотря даже на то, что оросительная вода
обычно содержит лишь незначительные концентрации солей. Соли остаются в почве,
и их количество медленно возрастает до тех пор, пока концентрация не начнет
повреждать растения. В этот момент нужно либо прекратить сельскохозяйственное
использование почвы, либо промыть ее избыточным количеством воды для удаления
соли. Однако большие количества воды, необходимые для промывания, часто
недоступны в зонах, требующих орошения[Бигон, 1989, с. 99−100].
4. Эколого-физиологические показатели,
характеризующие водный режим организмов
Количество влаги, поступающее в растение через
корни, редко удается определить. Обычно пользуются показателем, дающим
представление о водоотнимающей силе растения, - величиной осмотического
давления клеточного сока, выражаемой в Паскалях. Как видно на рисунке 3, в
зависимости от условий оно варьируется от 500-700 до 7000-9000 кПа( в отдельных
случаях достигает 10000 кПа и более).
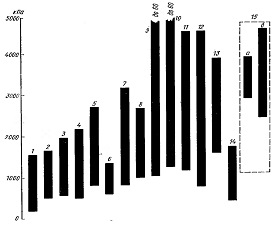
Растения: 1 - водные, 2 - весенние эфемероиды, 3
- луговые, 4 - культурные, 5 - сорняки, 6 - травы влажных лесов, 7 - травы
сухих лесов, 8 - лиственные деревья и кустарники, 9 - хвойные деревья, 10 -
жестколистные средиземноморские древесные породы, 11 - кустарники, 12 -
растения степные, 13 - пустынные, 14 - суккуленты, 15 - галофиты (а - растения
мангров, б - растения засоленных лугов).
Рисунок 3 − Амплитуды осмотического
давления клеточного сока в листьях растений различных экологических типов [по WalterH.,
1960]
В экологических работах определяется
интенсивность транспирации - количество испаренной воды за 1 ч в расчете на
единицу массы растения в граммах сырой или сухой массы или на единицу площади
листа. Экологическую значимость имеют не только величины транспирации, но и
характер во времени: дневной(суточный) ход и сезонный ход. Содержание воды у
растений разных экологических групп сильно варьируется. Здесь нет прямой связи
с водообеспеченностью местообитания, поскольку оводненность растения очень
сильно зависит от его анатомо-морфологических особенностей ( прежде всего от
количества механических, проводящих, покровных тканей). Содержание воды в
растениях не постоянно. Наиболее резко оно колеблется у пойкилогидридных
растений. В сезонном ходе содержания воды обычно отмечается падение его к концу
вегетации, которое может быть связанно как с ухудшением условий водоснабжения,
так и с возрастными изменениями листьев(старением). Вода в растениях содержится
в двух формах: свободной и коллоидно - связанной. Обычно доля связанной воды
велика у растений, испытывающих по тем или иным причинам затруднение в
пополнении расходов воды, что может рассматриваться как своего рода защитная
реакция растения против излишней транспирации [Горышина, 1979, с. 110−118;
Чернова и др., 1988].
5. Вода как среда обитания организмов. Экология
водных организмов. Хроматическая адаптация
.1 Вода как среда обитания организмов
Экологические группы растений по отношению к
воде
Гидатофиты - это водные растения, целиком или
почти целиком погруженные в воду. Среди них - цветковые, которые вторично
перешли к водному образу жизни (элодея, рдесты, водяные лютики, валлиснерия,
уруть и др.). Вынутые из воды, эти растения быстро высыхают и погибают. У них
редуцированы устьица и нет кутикулы. Транспирация у таких растений отсутствует,
а вода выделяется через особые клетки -гидатоды.
Гидрофиты - это растения наземно-водные,
частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на
болотах. Встречаются в районах с самыми разными климатическими условиями. К ним
можно отнести тростник обыкновенный, частуху подорожниковую, вахту трехлистную,
калужницу болотную и другие виды. У них лучше, чем у гидатофитов, развиты
проводящие и механические ткани. Хорошо выражена аэренхима. В аридных районах
при сильной инсоляции их листья имеют световую структуру. У гидрофитов есть
эпидерма с устьицами, интенсивность транспирации очень высока, и они могут
расти только при постоянном интенсивном поглощении воды.Так, для зимовки
многолетних гидрофитов в реках и озерах большое значение имеет вертикальное
распределение температур подо льдом. Наиболее плотная и наименее холодная вода
с температурой 4°С располагается в придонном слое, куда опускаются зимующие
почки (турионы) роголистника, пузырчатки, водокраса и др. (рис. 4), а также
целые облиственные растения, такие, как ряска, элодея.

Рисунок 4 - Водокрас (Hydrochariasmorsusranae)
осенью. Видны зимующие почки, опускающиеся на дно [по Кернеру А., 1896]
Гигрофиты - наземные растения, живущие в
условиях повышенной влажности воздуха и часто на влажных почвах. Среди них
различают теневые и световые. Теневые гигрофиты - это растения нижних ярусов
сырых лесов в разных климатических зонах (недотрога, цирцея альпийская, бодяк
огородный, многие тропические травы и т. п.). Из-за высокой влажности воздуха у
них может быть затруднена транспирация, поэтому для улучшения водного обмена на
листьях развиваются водяные устьица, выделяющие капельно-жидкую воду. Листья
часто тонкие, с теневой структурой, со слабо развитой кутикулой, содержат много
свободной и малосвязанной воды. Обводненность тканей достигает 80 % и более.
При наступлении даже непродолжительной и несильной засухи в тканях создается
отрицательный водный баланс, растения завядают и могут погибнуть.
Мезофитымогут переносить непродолжительную и не
очень сильную засуху. Это растения, произрастающие при среднем увлажнении,
умеренно теплом режиме и достаточно хорошей обеспеченности минеральным
питанием. К мезофитам можно отнести вечнозеленые деревья верхних ярусов
тропических лесов, листопадные деревья саванн, древесные породы влажных
вечнозеленых субтропических лесов, летнезеленые лиственные породы лесов
умеренного пояса, кустарники подлеска, травянистые растения дубравного
широкотравья, растения заливных и не слишком сухих суходольных лугов, пустынные
эфемеры и эфемероиды, многие сорные и большинство культурных растений. Из
приведенного перечня видно, что группа мезофитов очень обширна и неоднородна.
По способности регулировать свой водный обмен одни приближаются к гигрофитам
(мезогигрофиты), другие к засухоустойчивым формам (мезоксерофиты).
Ксерофитырастут в местах с недостаточным
увлажнением и имеют приспособления, позволяющие добывать воду при ее
недостатке, ограничивать испарение воды или запасать ее на время засухи.
Ксерофиты лучше, чем все другие растения, способны регулировать водный обмен,
поэтому и во время продолжительной засухи остаются в активном состоянии. Это
растения пустынь, степей, жестколистных вечнозеленых лесов и кустарниковых
зарослей, песчаных дюн. Примером приспособления к недостаточной влажности,
может служить длинный корень у растений, как на рисунке 5.

Рисунок 5 - Корневая система пустынного
кустарника Pituranthostortuosus[по
Вальтеру Г., 1968]
Ксерофиты подразделяются на два основных типа:
суккуленты и склерофиты.
Суккуленты − сочные растения с сильно
развитой водозапасающей паренхимой в разных органах. Стеблевые суккуленты -
кактусы, стапелии, кактусовидные молочаи. Листовые суккуленты - алоэ, агавы,
мезембриантемумы, молодило, очитки. Корневые суккуленты - аспарагус. В пустынях
Центральной Америки и Южной Африки суккуленты могут определять облик ландшафта
(рис. 6).

Рисунок 6 -Суккулентное растение - древовидный
кактус из пустыни Аризона
Листья, а в случае их редукции стебли
суккулентов имеют толстую кутикулу, часто мощный восковой налет или
густоеопушение. Устьица погруженные, открываются в щель, где задерживаются
водяные пары. Водозапасающие ткани у них могут быть развиты в стеблях или
листьях, поэтому они подразделяются на стеблевые суккуленты (кактусы, молочаи)
и листовые суккуленты (алоэ, агава, молодило). Тело суккулентов, как правило,
покрыто толстым кутинизированным эпидермисом и восковым налетом. Устьиц на
поверхности тела почти нет. А если есть, то они мелкие, располагаются в ямках и
большую часть времени закрыты. Открываются только на ночь. Все это предельно
сокращает транспирацию. Характерной особенностью суккулентов является высокая
поглощающая способность. В период дождей отдельные виды поглощают в себя
большое количество воды. Накопленную влагу суккуленты в дальнейшем медленно
расходуют. Суккуленты произрастают в районах с жарким сухим климатом. Там, где
хотя бы изредка проходят дожди кратковременные, но обильные, ливневые.
Склерофиты (от греч. «склерос» - твердый,
жесткий) не накапливают в себе влагу, а испаряют ее в большом количестве,
постоянно доставая из глубоких слоев почвы. Тело этих растений жестковатое,
сухое, иногда одревесневшее, с большим количеством механической ткани. При
продолжительном прекращении подачи воды может наблюдаться сбрасывание листьев
или части побегов, что приводит к сокращению испарения. Многие из ксерофитов
переносят засушливое время года в состоянии вынужденного покоя. Примерами
являются пустынная полынь, ковыль, саксаул.
Стипаксерофиты является группой узколистных
дерновинных злаков (ковыли, тонконоги, типчак и др.).Отличаются повышенными
скоростями физиологического реагирования на изменение внешних условий. Они
быстро и наиболее полно используют влагу кратковременных ливневых осадков, а
также сравнительно хорошо переносят перегрев. Весьма чувствительны к
обезвоживанию, могут переносить лишь кратковременный недостаток влаги в почве.
Характеризуются низкой транспирацией в засушливый период и могут переносить
особенно сильное обезвоживание тканей[Чернова, 2004, с. 79-83].
.2 Экология водных организмов
Водные растения называют гидрофитами. По образу
жизни и строению среди них можно выделить погруженные растения и растения с
плавающими листьями. Погруженные растения подразделяют на укореняющиеся в
донном грунте и взвешенные в толще воды. Из высших растений к первым
принадлежат полушник озерный, телорез, шильник водяной, в эту же группу водят
водоросли прикрепленные к грунту. Важной особенностью адаптации растений к
обитанию в водной среде является и то, что листья, погруженные в воду, как
правило, очень тонкие. Часто хлорофилл в них располагается в клетках
эпидермиса, что способствует усилению интенсивности фотосинтеза при слабом
освещении. Такие анатомо-морфологические особенности наиболее четко выражены у
водяных мхов (Riccia, Fontinalis),
валиснерии (Vallisneriaspiralis),
рдестов (Potamageton).
От вымывания у водных растений из клеток
минеральных солей или выщелачивания защитой является выделение специальными
клетками слизи и образование эндодермы из более толстостенных клеток в виде
кольца.
Относительно низкая температура водной среды
обусловливает отмирание вегетирующих частей у погруженных в воду растений после
образования зимних почек и замену летних тонких нежных листьев более жесткими и
короткими зимними. Низкая температура воды отрицательно сказывается на
генеративных органах водных растений, а высокая ее плотность затрудняет перенос
пыльцы. В связи с этим водные растения интенсивно размножаются вегетативным
путем. Большинство плавающих на поверхности и погруженных растений выносят
цветоносные стебли в воздушную среду и размножаются половым путем. Пыльца
разносится ветром и поверхностными течениями. Плоды и семена, которые
образуются, также распространяются поверхностными течениями. Это явление носит
название гидрохории. К гидрохорным относятся не только водные, а также
многие прибрежные растения. Их плоды имеют высокую плавучесть, длительное время
находятся в воде и не теряют при этом всхожесть. Например, водой переносятся
плоды и семена стрелолиста (Sagittariasagittofolia),
сусака (Butomusumbellatus),
частухи (Alismaplantago-aguatica).
Из растений, взвешенных в толще воды, можно
назвать роголистник погруженный, пузырчатку обыкновенную, а также
многочисленные виды планктонных водорослей. К настоящим водным растениям очень
близко примыкает и обычно вместе с ним рассматривается группа гелофитов. Это
виды береговых и прибрежных местообитаний с избыточным или переменным
увлажнением. Примерами являются стрелолист, частуха, сусак, поручейник,
ежеголовка[Степановских, 1979, с. 121−122].
5.3 Хроматическая адаптация
Так, растения, живущие на поверхности воды, не
испытывают недостатка света, а погруженные и особенно глубоководные относят к
«теневой флоре». Им приходится адаптироваться не только к недостатку света, но
и к изменению его состава выработкой дополнительных пигментов. Это
прослеживается на известной закономерности окраски у водорослей, обитающих на
разных глубинах. В мелководных зонах, где растениям еще доступны красные лучи,
которые в наибольшей степени поглощаются хлорофиллом, как правило, преобладают
зеленые водоросли. В более глубоких зонах встречаются бурые водоросли, имеющие
кроме хлорофилла бурые пигменты фикофеин, фукоксантин и др. Еще глубже обитают
красные водоросли, содержащие пигмент фикоэритрин. Здесь четко прослеживается
способность к улавливанию солнечных лучей с разной длиной волны. Данное явление
получило название хроматической адаптации.
Характерно для сине-зелёных (Cyanophyta)
и красных (Rhodophyta) водорослей [Степановских, 2001, с.114−115].
6. Характеристика редких водных животных,
занесенных в Красную книгу Краснодарского края. Характеристика редких видов
гигрофитов Западного Кавказа
.1 Характеристика редких водных животных,
занесенных в Красную книгу Краснодарского края
. Вид бывает редок, если потенциально пригодные
для его жизни участки редко встречаются или малы по площади. Существование
специфической флоры и фауны может поддерживаться необычным сочетанием
физико-химических параметров среды.
. Вид бывает редок из-за конкурентного
исключения со стороны других видов, сильного воздействия хищников или
паразитов.
. Вид бывает редок из-за недостатка внутри таких
зон жизненно важных ресурсов - пищи, убежищ и т.д.
. Вид бывает редок из-за того, что недостаточная
наследственная изменчивость его представителей сужает диапазон пригодных для
него местообитаний.
. Вид бывает редок из-за того, что недостаточная
фенотипическая пластичность его представителей сужает диапазон пригодных для
него местообитаний.
. Вид бывает редок, поскольку конкуренты,
хищники, паразиты, сборщики или коллекционеры удерживают плотность его
популяций ниже уровня, обеспечиваемого доступными ресурсами. Интересно
отметить, что собираемые людьми виды становятся более «ценными» и интенсивнее
разыскиваемыми именно после того, как начинают встречаться реже[Бигон, 1989, с.
109−112].
Рассмотрим следующих редких водных животных,
занесенных в Красную книгу Краснодарского края:
Горбыль светлый - Umbrinacirrosa
(Linnaeus, 1758). Относится к семействугорбылёвые - Sciaenidae.Тело
прогонистое, горбатое, длиной до 1,5 м. Чаще встречаются экземпляры 23-33 см.
Вес до 32 кг.Распространен в Атлантическом океане у берегов Европы и Африки, в
Средиземном, Мраморном морях. Также обитает вприбрежных водах Черного моря, а
также участках устья Кубани и Протоки в Азовском море, отмечен в
Кизилташскихлиманах. Придонная рыба, предпочитает скалистые берега, а также
песчаные отмели с галькой, ракушечником, реже встречается на участках с илистым
дном. У берегов держится с апреля по декабрь, зимой уходит на глубину. Размножается
в Черном море с апреля по август. Питается червями и рыбой: хамсой, сардиной,
кефалью, барабулей, также в питании присутствуют крабы, креветки и моллюски.
Редкий вид, имеет тенденцию к снижению численности.
Черноморская афалина - TursiopstruncatusponticusBarabasch-nikiforov,
1940. Относится к семейству дельфиновые - Delphinidae. Средняя длина
тела половозрелых афалин 220-240 см (самый крупный экземпляр, пойманный в
Черном море, - 310 см). Глобальный ареал включает всю акваторию Черного моря, а
не только прибрежные районы, как считалось раньше. В РФ населяет внутренние
воды России в Черном море. Регулярно наблюдается в Керченском проливе, но не
заходит в Азовское море. Вместе с косяками рыбы совершает нерегулярные кочевки
по Черному морю. Осенью, ко времени выхода хамсы из Азовского моря, афалины
скапливаются в северо-восточных частях моря вблизи Керченского пролива и у
берегов Северного Кавказа.В море обычно держатся поодиночке, парами или
небольшими группами в несколько особей (до 15-20).Охотно сопровождают суда,
часто выпрыгивают из воды целиком на высоту до 3-4 м. Короткое время могут
плыть со скоростью до 40 км/ч, нырять на глубину до 500 м и оставаться под
водой до 15 минут. Основу питания составляют рыбы: хамса, атерина, барабуля,
камбала, кефаль, лобан, пелингас, пеламида, судак, лещ.В море у афалин нередки
случаи грибковых и бактериальных поражений кожи. Они страдают от пневмоний,
заболеваний желудочно-кишечного тракта и сердечно-сосудистой системы. Афалина
лучше многих других видов переносит неволю, легко поддается дрессировке, долго
может жить в дельфинариях и при хороших условиях размножается.
Пиявкамедицинская - Hirudomedicinalis
Linnaeus,
1758. Относится к семейству гирудиниды - hirudinidae.Вид известен только
из типового местонахождения: Западный Кавказ, Красноалександровская пещера на
р. Аше севернее пос. Лазаревского. Обитает на каменистом грунте чистого
проточного пещерного водоема в условиях малого содержания орга-нического
вещества и большого количества растворенного кислорода. Локально распространенный,
стенобионтный, малочисленный вид. Эндемик Западного Кавказа. Вид редок в
коллекциях, его численность в настоящий момент не поддается точной оценке.
Горошинка пещерная - PisidiumcavaticumShadin,
1952. Относится к семейству горошины - Pisidiidae.Эндемик Западного
Кавказа. Пресноводный троглобионтный вид. Особенности биологии и экологии не
изучены. Вид известен только из двух точек: Западный Кавказ, Краснодарский
край, верховья р. Хосты,ручей в глубине пещеры (типовое местонахождение),
пещера Цебельда (Абхазия). В настоящий момент численность не поддается точной
оценке. Известно менее двух десятков экземпляров данного вида, собранных Я.А.
Бирштейном, С.И. Левушкиным в период с 1950 по 1959 г.
Проанализировав данные Красной книги
Краснодарского края по редким водным животным, были выявлены следующие
тенденции лимитирующих факторов, которые представлены в виде диаграммы на
рисунке 8.
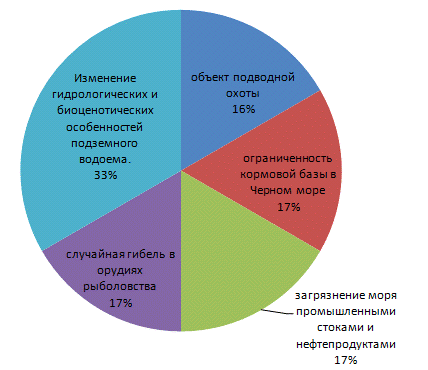
Рисунок 7 - Диаграмма лимитирующих факторов для
редких водных животных Краснодарского края
Изучив данные на диаграмме, можно сделать вывод,
что главным лимитирующим фактором будет являться изменение гидрологических и
биоценотических особенностей подземного водоема. Остальные четыре фактора
пагубно влияют на первый, тем сам усугубляя его.
.2 Характеристика редких видов гигрофитов
Западного Кавказа
Срединския большая - Sredinskyagrandis
(Trautv.) Fed. 1951. Относится к семейству Первоцветные - Primulaceae. В
России распространен на Северном Кавказе, Западном Кавказе, Западном Закавказье.
Гигрофит, гелиофит, микротерм, криптофит. Обитает на влажных альпийских
лужайках, по берегам горных ручьев, у окраин тающего снега.Цветет в июле,
плодоносит в сентябре.Обитает на влажных альпийских лужайках, по берегам горных
ручьев, у окраин тающего снега. Встречается небольшими группами.Охраняется на
территории КГПБЗ и СНП. Вне региона охраняется в Кабардино-Балкарском
заповеднике. Необходим поиск новых местонахождений и контроль за состоянием
популяций.
Сабельник болотный - Comarumpalustre L.
1753. Относится к семейству Розовые - Rosaceae. Распространен в
Скандинавии, Восточной Молдовы и Украины, Средней и Центральной Азии, Северная
Америка, Европейская часть России, Западная и Восточная Сибирь, Дальний Восток,
Северный Кавказ, Западный Кавказ. В высокогорной зоне Западного Кавказа цветет
во второй половине сентября, плоды не вызревают. Размножается вегетативным
путем корневищами. Вид обладает низкой жизненностью. Произрастает на
высокогорных осоково-моховых болотах и заболоченных лугах.Высока вероятность
гибели малочисленных популяций в результате действия естественных факторов или
изменения климата. Популяции вида могут быть уничтожены в случае отторжения
участков территории от КГПБЗ с целью хозяйственного или рекреационного
использования. Усиление антропогенного эвтрофирования водоемов приводит к
снижению жизненности популяций вида
Жирянка обыкновенная - Pinguiculavulgaris
L. 1753. Относится к семейству Пузырчатковые - Lentibulariaceae. Обитает
в Средней и Северной Европе, Восточном Средиземноморье, Юго-Восточной Азии,
Северной Америке, а также в России, а именно: Арктика, Европейская часть,
Дальний Восток, Северный Кавказ, Западный Кавказ. Цветет в июле - августе.
Растение плотоядное. Произрастает на заболоченных лугах, болотистых местах,
местах выхода родниковых вод и схода снега в субальпийском и альпийском
поясах.Основные местообитания расположены в приграничных районах КГПБЗ в
пределах зоны интересов горнолыжной индустрии, отгонного животноводства.
Популяции вида могут быть уничтожены в случае изменения климата, а также
отторжения участков территории от заповедника с целью развития горнолыжных
курортов, выпаса скота, стро-ительства новых автодорог.
Осока топяная - Carexlimosa L. 1753.
Относится к семейству Осоковые - Cyperaceae. Распространен в Северной
Америке, Европе: Атлантической, Северной, Центральной, Южной, Юго-Восточной;
Азия: Северная, Юго-Западная и Восточная. В России вид распространен в
Европейской части, Грузии и Южной Осетии, Северном и Западном Кавказе и Дальнем
Востоке. Цветет в июле - августе. Произрастает преимущественно на верховых
болотах. На Кавказе встречается на заболоченных лугах, низинных осоково-моховых
болотах, надводных сплавинах в субальпийском поясе. Все водоемы характеризуются
преимущественно дождевым питанием и, соответственно, слабокислыми
низкоминерализованными водами.Основные места обитания расположены на границе
КГПБЗ в пределах зоны интересов горнолыжной индустрии. Популяции вида могут
быть уничтожены в случае отторжения участков территории от запо-ведника, развития
горнолыжного туризма, строительства новых автодорог.
Проанализировав данные Красной книги
Краснодарского края по редким видам гигрофитов Западного Кавказа, были выявлены
следующие тенденции лимитирующих факторов, представленных в виде диаграммы на
рисунке 8.
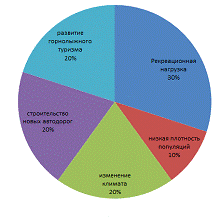
Рисунок 8 - Диаграмма лимитирующих факторов для
редких видов гигрофитов Западного Кавказа
Изучив процентное соотношение всех факторов, на
рисунке видно, что самым большим лимитирующим фактором является рекреационная
нагрузка на территорию, следующими факторами будут развитие горнолыжного
туризма, строительство новых автодорог, изменение климата. Последним фактором
является низкая плотность популяций.
Заключение
Уступая по своему значению температуре, влажность
относится к основным экологическим факторам. На протяжении большей части
истории живой природы органический мир был представлен исключительно водными
нормами организмов. Влажность почвы чрезвычайно важная характеристика, особенно
для произрастающих на ней растений. Она зависит от многочисленных факторов:
режима дождей, глубины залегания слоя, а также физических и химических свойств
почвы, частицы которой в зависимости от их размера, содержания органического
вещества и т.п. Флора сухих и влажных почв неодинакова и на этих почвах нельзя
разводить одни и те же культуры. Фауна почвы также весьма чувствительная к ее
влажности и, как правило не переносит слишком большой сухости. Общеизвестным
примером служат дождевые черви и термиты. Последние иногда вынуждены снабжать
водой свои колонии, проделывая подземные галереи на большой глубине. Однако
слишком высокое содержание воды в почве убивает личинки насекомых в больших
количествах.
Тело растения на 50−90% состоит из воды.
Вода играет непосредственную роль в обеспечении роста растений, являющегося
функцией растяжения клеток. Тургорное давление, производимое изнутри на молодые
оболочки клеток водой, вызывает их растяжение, пока они еще не эластичны.
Характерными аридными условиями являются сухой климат с высокими температурами
воздуха, испытывающими большие суточные колебания, и малым количеством
атмосферных осадков (100−150 мм/год) или полным их отсутствием. К
гумидным условиям относятся области с избыточным увлажнением, при котором
количество атмосферных осадков больше, чем может испариться и просочиться в
почвогрунты. Все животные могут выдерживать определенные отклонения от
нормального содержания воды в теле (обычно около 10%). Чтобы не допустить
опасные для жизни отклонения содержания воды, многим из них приходится
вырабатывать соответствующие адаптации. Растения испытывают водный дефицит,
когда скорость транспирации превосходит скорость поглощения воды корневой
системой. Такая ситуация возникает не только при засухе, но и в условиях
почвенного засоления, а также при низких температурах. У одиночного растения
все листья будут транспирировать, так как они окружены сравнительно сухим
воздухом. Однако в условиях полевой культуры нижние листья окружены
преимущественно влажным воздухом в результате транспирации других листьев.
Только верхние листья соприкасаются непосредственно с более сухой атмосферой.
Кроме того, листья, расположенные ниже верхнего слоя, настолько ограничивают
движение воздуха, что между растениями он становится насыщенным влагой. Поэтому
нижние листья обладают значительно более низкой интенсивностью транспирации.
Высокие концентрации NaCl и других солей в оросительной воде повреждают
растения, так как осмотические эффекты тормозят поглощение воды, а натрий
ингибирует ферменты в цитоплазме. Поэтому при накоплении соли растения
становятся чахлыми и в конце концов перестают расти. Поскольку в условиях
сухого климата фактически вся поливная вода испаряется, засоленность почвы
увеличивается несмотря даже на то, что оросительная вода обычно содержит лишь
незначительные концентрации солей. Содержание воды у растений разных
экологических групп сильно варьируется. Здесь нет прямой связи с
водообеспеченностью местообитания, поскольку оводненность растения очень сильно
зависит от его анатомо-морфологических особенностей (прежде всего от количества
механических, проводящих, покровных тканей). Содержание воды в растениях не
постоянно. Наиболее резко оно колеблется у пойкилогидридных растений. В
сезонном ходе содержания воды обычно отмечается падение его к концу вегетации,
которое может быть связанно как с ухудшением условий водоснабжения, так и с
возрастными изменениями листьев(старением). Также выделяют следующие
экологические группы растений по отношению к воде:
Гидатофиты − это водные растения, целиком
или почти целиком погруженные в воду.
Гидрофиты − это растения наземно-водные,
частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на
болотах.
Гигрофиты наземные растения, живущие в условиях
повышенной влажности воздуха и часто на влажных почвах.
Мезофитымогут переносить непродолжительную и не
очень сильную засуху.
Ксерофитырастут в местах с недостаточным
увлажнением и имеют приспособления, позволяющие добывать воду при ее
недостатке, ограничивать испарение воды или запасать ее на время засухи.
По образу жизни и строению среди водных растений
можно выделить погруженные растения и растения с плавающими листьями.
Погруженные растения подразделяют на укореняющиеся в донном грунте и взвешенные
в толще воды. Из высших растений к первым принадлежат полушник озерный,
телорез, шильник водяной, в эту же группу водят водоросли прикрепленные к
грунту.
Говоря о редких водных животных, занесенных в
Красную книгу Краснодарского края, их насчитывается четыре вида, и редких видов
гигрофитов Западного Кавказа тоже четыре.
И в заключении хотелось бы отметить, что
собираемые людьми виды становятся более «ценными» и интенсивнее разыскиваемыми
именно после того, когда начинают встречаться реже.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Березина Н.А. Экология растений:
учеб.пособие для студ. высш. учеб. заведений - М.: Издательский центр
"Академия", 2009. 400 с.
. Бигон М. Экология особи, популяции
и сообщества: В 2 т. / М. Бигон, Дж. Харпер, К. Таусенд. - М.: Мир, 1989. -
Т.1. - 660 с.; Т.2. 473 с.
. Горышина Т.К. Экология растений.
М.:Высш. Школа, 1979. 368 с.
. Дедю И.И. Экологический
энциклопедический словарь. - Кишинев: Изд-во МСЭ, 1990. 406 с.
. Коробкин, В.И. Экология: Учеб.для
студ. вузов / В.И. Коробкин, Л.В. Передельский. - Ростов н/Д: Феникс, 2000. 576
с.
. Красная книга Краснодарского края.
Т. 1. Растения. Т. 2. Животные. 2007.
. Культиасов ИМ. Экология растений.
М.: Изд-во МГУ. 1982. 381 с.
. Николайкин Н.И., Николайкина Н.Е.,
Мелехова Экология. О.П. 3-е изд., стереотип. - М.: Дрофа, 2004. 624 с
. Новиков Г.А. Основы общей экологии
и охраны природы. - Л.: Изд-во Ленингр. ун-та, 1979. 352 с.
. Степановских А.С. Экология. -
Курган: ГИПП «Зауралье», 1997. 616 с.
. Чернова Н.М., Былова А.М.
Экология. М.:Просвещение,1988. 272 с.