Распространение основных групп микроорганизмов в почвах разных типов
Министерство
сельского хозяйства Российской федерации
Российский
государственный аграрный университет - МСХА
Имени К. А.
Тимирязева
(ФГБОУ ВПО
РГАУ - МСХА имени К. А. Тимирязева)
Факультет
почвоведения, агрохимии и экологии
Кафедра
микробиологии и иммунологии
КУРСОВАЯ
РАБОТА
По
дисциплине: Почвенная микробиология
На
тему: “Распространение основных групп микроорганизмов в почвах разных типов”
Выполнил:
Студент 202 группы
Г.А. Ивашенков
Проверил:
доцент А. А. Ванькова
Москва, 2012
Содержание
Аннотация
Введение
Глава 1. Теоретическая часть.
Распространение основных групп микроорганизмов в почвах разных типов
.1 Факторы, влияющие на формирование
микробных ценозов почв различных типов
.1.1 Температурный режим почвы
.1.2 Влажность почвы
.1.3 Воздушный режим почвы
.1.4 Окислительно-восстановительные
условия
.1.5 Кислотность почвы
.2 Численность микроорганизмов в
почвах разных типов
.3 Соотношение основных групп
микроорганизмов
.3.1 Сапротрофная группировка
микроорганизмов
.3.2 Олиготрофные микроорганизмы
.3.3 Хемолитоафтотрофные и
фотолитоавтотрофные микроорганизм
.4 Заключение
Глава 2. Практическая часть
.1. Объекты и методы исследования
.1.1 Объект исследования
.1.2 Метод исследования
.2 Результаты исследований
.2.1 Микроорганизмы, использующие
органическую форму азота (МПА)
.2.2 Микромицеты (СА)
.2.3 Азотфиксирующие бактерии (среда
Эшби)
.2.4 Целлюлозоразрушающие бактерии
(среда Гюйгенса)
.3 Выводы
Аннотация
Данная курсовая работа посвящена микробиоценозам
почв разных типов. Она состоит их двух частей: теоритической и практической. В
первой излагаются данные об основных группах микроорганизмов и их распределении
в почве. Что касается второй, то там делалось описание эксперимента по
выявлению из данной почвенной навески (дерново-подзолистой) конкретных видов
микроорганизмов и их определение.
микроорганизм
почва ценоз
Введение
Почва - одна из наиболее благоприятных сред для
существования микроорганизмов. Особенности почвы как среды обитания микробов
состоит в том, что это трехфазная система с очень развитой твердой
поверхностью, которая соседствует с жидкой и газовой фазами. Твердые частицы и
агрегаты делят почву на многочисленные частично и полностью изолированные
микрозоны, в котором создаются резко отличающиеся, а часто даже противоположные
условия. Клетки микробов имеют микроскопические размеры, и средой их обитания
является микросреда. Сотни и тысячи таких микросред сосредоточены в каждом
грамме почвы. Почва - это комплекс одновременно существующих, но совершенно
различных микросред.
Долгое время микроорганизмы вообще и почвенные в
частности рассматривались как космополиты, более или менее однородно
распределенные по поверхности земного шара. Предполагалось, что почвы
различаются лишь по численности, но не по составу их микронаселения. Однако микроорганизмы
- весьма обобщенное понятие. Оно включает в себя существ, относящихся к разным
таксонам, вплоть до царств и доменов. Поэтому ясно что один тип микробов, в
силу различия своих характеристик и свойств по сравнению с другими группами
микроорганизмов, зачастую будет большей частью своей численности обитать в
одной зоне почвы, в то время как другой тип - в соответственно другой зоне. То
же самое касается и почв: ее типы различаются по физическим, химическим и
прочим свойствам и характеристикам и соответственно часто имеют свою уникальную
биосистему, в том числе и микробиосисему. Поэтому ясно, что удельный вес
различных групп микроорганизмов у различных типов почв отнюдь не одинаков.
Глава 1. Теоретическая часть. Распространение
основных групп микроорганизмов в почвах разных типов.
1.1 Факторы, влияющие на формирование микробных
ценозов почв различных типов
На активность микроорганизмов и формирования их
сообществ в почве влияет ряд природных и антропогенных факторов. Среди них
температура почвы, ее влажность, воздушный режим,
окислительно-восстановительный потенциал, кислотность и механические свойства,
а также биотические факторы.
.1.1 Температурный режим почвы
Температура почвы определяется географическим
фактором и сезоном года. Наблюдается и суточные колебания температуры, которые
сильнее всего сказываются в поверхностном слое почвы. Сезонные колебания
температуры влияют на весь профиль почвы. В криофильных почвах на определенной
глубине залегает постоянный слой, подавляющий активность микроорганизмов.
В одной и той же зоне температурный режим почвы
зависит также от ее способности поглощать тепловые лучи, теплоизлучения, от
характера растительности и т.д.
Что касается микроорганизмов, то по отношению к
температуре их принято делить на следующие группы: психрофилы, размножающиеся
при относительно низких положительных температурах; мезофилы, которые растут
при обычных температурах окружающей среды; термофилы, нуждающиеся в высоких
температурах.
Основная масса почвенных микроорганизмов
принадлежит к мезофилам. Доказано, что оптимум и максимум температуры для
большинства почвенных мезофильных микроорганизмов меняется в зависимости от
климата. Бактерии в южных почвах, как правило, имеют более высокие температрные
оптимум и максимум.
В этом сказывается приспособительная реакция
микроорганизмов к условиям среды. Она отсутствует лишь у видов бактерий,
завершающих процесс минерализации органических остатков (Bacillus
megaterium, B.
Subtilis, B.
Mesentericus и др.). В основном
это спорообразующие виды. Для их существования температурная адаптация не имеет
столь большого значения.
В почвах южной зоны, как правило, обитают
микроорганизмы, предъявляющие повышенные требования к теплу. Например, южные
почвы значительно богаче теплолюбивыми грибами рода Aspergillus,
в то время как в северных преобладают виды рода Penicillium,
способные развиваться при более низких температурах.
Теплолюбивые микроорганизмы при оптимальной для
них температуре более активны, чем психрофильные. Термофилы обладают
исключительно высокой биохимической активностью. В связи с этим в южных почвах
при благоприятных условиях микробиологические процессы протекают более
энергично, чем в северных. Однако термофильных микроорганизмов даже в южных
почвах мало, и существенной роли в почвенных процессах они не играют. Это
объясняется тем, что при сильном нагревании почва быстро пересыхает, создавая
неблагоприятную обстановку для размножения термофилов [2].
Сопоставление температуры почвы разных
климатических зон с потребностью мезофильных форм бактерий в тепле
|
Пункт
|
Средняя
температура почвы за май и август (a)
|
Примерная
оптимальная температура для бактерий (б)
|
Разница
(б-а)
|
|
Архангельск
|
10,5
|
28,5
|
18,0
|
|
Москва
|
12,7
|
30,0
|
|
Курск
|
16,4
|
34,0
|
17,6
|
|
Северный
Кавказ
|
22,4
|
35,5
|
13,1
|
|
Ташкент
|
30,0
|
38,0
|
8,0
|
1.1.2 Воздушный режим почвы
Воздушный режим почвы также имеет большое
значение для микробиологических процессов, чем температура и влажность. Воздух
содержит в почвенных порах, которые составляют в отдельных случаях от 25 до 70%
общего объема почвы. Содержание воздуха в почве подвержено сильным колебаниям в
зависимости от ее увлажнения, уплотнения и т.д.
В связи с потреблением почвенными организмами и
корнями растений содержание кислорода в почвенном воздухе ниже, чем в
атмосфере. В затопленных почвах иногда кислорода почти нет. В воздухе
содержащейся почвы кислорода около 2%, в хорошо взрыхленной - до 20%.
В почвенном воздухе обычно повышено содержание
диоксида углерода, который выделяют микроорганизмы и корни растений. Чаще всего
CO2 составляет
0,3-1,5 % в почвенном воздухе (в атмосферном - 0,03%). В затопленных почвах
количество CO2 повышается
до 10%. В атмосфере щелочных почв диоксид углерода почти отсутствует. В целом
газовый состав почвенного воздуха подвержен суточным и сезонным колебаниям.
Большое количество CO2
ежесуточно выделяется из почвы, а вместо него из атмосферы поступает воздух.
Данный процесс, получивший название “дыхание почвы”, имеет огромное значение
для жизни растительного мира.
Аэробные микроорганизмы хорошо переносят
повышенное содержание в воздухе CO2.
Нередко при этом отмечается даже улучшение их роста при обогащении воздуха
диоксидом углерода. Тем не менее при 1-1,5% и более активность некоторых групп
микроорганизмов подавляется. Вероятно, отчасти по этой причине в нижних
горизонтах пахотного слоя меньше микроорганизмов, чем в верхних.
Вместе с тем аэробные цианобактерии лучше
развиваются при повышенном содержании CO2
в почве. Оптимальная для них концентрация углекислого газа равна 1%,
максимальная для ряда представителей достигает 12% и выше. [2]
.1.3 Влажность почвы
Огромное влияние на жизнедеятельность
микроорганизмов оказывает влажность почвы. Вода, составляющая жидкую фазу
почвы, содержит то или иное количество растворенных веществ. Растения и
микроорганизмы усваивают питательные вещества в основном из почвенного
раствора.
По величине осмотического давления клетки
микроорганизмов почвы существенно различаются. Большинство из них способны
развиваться в почве при наличии по крайней мере гигроскопичной влаги. В южных
почвах микробы адаптированы к более сухим условиям существования, чем в
северных. Некоторые актиномицеты и ряд грибов развиваются даже при крайне
низкой влажности почвы.
Можно считать, что мобилизационные,
агрономически желательные процессы лучше всего протекают при влажности почвы,
приближающейся к 60% полной влагоемкости. При таком увлажнении в почве
достаточно воды и воздуха, находящихся между почвенными агрегатами. При более
сильном увлажнении воздух из почвы вытесняется, что подавляет аэробные
микробиологические процессы. Подобная обстановка, например, создается на
залитых водой рисовых полях, где относительно благоприятные аэробные условия
имеются лишь в самом поверхностном слое почвы.
Установлено, что аммонификация и нитрификация
лучше всего проходят при влажности почвы, равной 60% полной влагоемкости.
Оптимум фиксации азота несколько сдвинут в сторону более высокой влажности.
Это, очевидно, объясняется тем, что молекулярный азот связывает многие
бактерии, склонные к анаэробиозу.
В природной обстановке влажность почвы сильно
колеблется, особенно резко в неорошаемых почвах южной зоны. Здесь в период
дефицита влаги нередко подавляется деятельность бактерий, но активизируются
актиномицеты и грибы. При значительном иссушении активная деятельность
микроорганизмов в почве вообще прекращается. В сухих степях наиболее энергичные
микробиологические процессы вообще протекают не летом, а весной и осенью, т.е.
при более низкой температуре, но достаточной влажности.
Таким образом, в почвенных процессах огромное
значение имеет сочетание условий температуры и влажности: как высокая, так и
низкая температура подавляют эти процессы в почве.
Следует отметить, что при низкой влажности
почвы, не обеспечивающей сколько-нибудь интенсивного развития организмов, в ней
могут достаточно активно работать разнообразные ферменты. Это показывает, что в
период дефицита влаги почва не представляет собой инертный субстрат. В полевых
условиях благоприятный водный режим может быть создан обработкой почвы,
поливом, мелиорацией и т.д.
Температурно-водный режим оказывает большое
влияние на формирование микробных сообществ почвы. Наблюдения свидетельствуют о
закономерной смене микробных сообществ в ходе распада органического вещества.
Так, в первые фазы разложения растительных остатков на них развиваются грибы и
неспорообразующие бактерии. Позднее увеличивается число бацилл и актиномицетов.
Происходит также смена систематических групп микроорганизмов. В разных
климатических зонах разложение органического вещества идет с неодинаковой
скоростью, что существенно сказывается на составе микрофлоры почвы [2].
.1.4 Окислительно-восстановительные условия
почвы
Обеспеченность почвы кислородом связана с ее
окислительно-восстановительными условиями, характеризующимися величиной
окислительно-восстановительного потенциала (Eh).
В верхнем слое подзолов значение Eh
обычно колеблется в пределах 600-750 мВ, в черноземах - 350-600, в сероземах -
350-400 мВ. Приведенные показатели усреднены, так как почва представляет собой
гетерогенную среду и в отдельных ее зонах наблюдаются существенные колебания
значений Eh.
В нормально увлажненных почвах создаются более
или менее аэробные условия. Относительно анаэробную обстановку можно наблюдать
лишь внутри отдельных комочков почвы, потому что микроорганизмы, находящиеся на
поверхности агрегатов, поглощают кислород, не пропуская его внутрь комка. В
течение вегетационного периода значение окислительно-восстановительного
потенциала в почве также более или менее колеблется.
К аэробным микроорганизмам почвы относят
мицельные грибы, большинство актиномицетов и значительную часть других
бактерий. Аэробные бактерии и актиномицеты могут существовать при относительно
небольших запасах кислорода, что делает понятным из размножение в глубоких
слоях почвы.
Большинство аэробных бактерий при росте на
жидких питательных средах снижают значение Eh
до 100-120 мВ. Многие из них могут жить и в анаэробных условиях (например, за
счет восстановления нитратов и т.д.).
Возникает вопрос о месте обитания в почве
анаэробных микроорганизмов, к которым в основном относятся бактерии.
Максимальная численность анаэробных бактерий наблюдается в верхних слоях почвы.
Это связано с тем, что здесь имеется небольшое количество органических веществ,
а анаэробные микрозоны, в которых эти микроорганизмы размножаются, возникают в
каждом комочке почвы [3].
.1.5 Кислотность почвы
На характер микрофлоры большое влияние оказывает
активная кислотность почвы. По величине pH
почвы могут быть разделены на следующие группы: сильнокислые (pH
3-4), кислые (4-5), слабокислые (5-6), нейтральные (6-7), щелочные (7-8),
сильнощелочные (8-9) и выше.
В подзолах значение pH
- 7,5.
Одна и та же величина реакции среды может иметь
неодинаковое значение в жизнедеятельности микроорганизмов в разных почвах. Так,
в подзолах уже небольшое снижение pH
вызывает освобождение алюминия, токсичного для ряда микроорганизмов; такого не
наблюдается в богатых кальцием черноземах. Поэтому подкисление подзолов
вызывает более сильное подавление микробиологических процессов, чем подкисление
в той же степени черноземов.
К алюминию особо чувствительны актиномицеты,
азотобактер и многие водоросли. Повышенное содержание этого элемента переносят
грибы и ряд бактерий. Отчасти вследствие этого северные почвы бедны
актиномицетами и азотобактером.
Микроорганизмы одной и той же систематической
группы неодинаково относятся к кислотности среды. Так, большинство бактерий
почвы не развивается при значении реакции среды ниже pH
4,5-5, но некоторые из них (например, Thiobacillus
thiooxidans) могут сохраняться
в жизнеспособном состоянии даже при pH
0,9.
Минимальные значения кислотности среды для грибов составляет pH
2-3, но некоторые грибы рода Mortierella
не выносят подкисления. Чувствительны к низким значениям реакции среды
актиномицеты.
Не смотря на разную кислото- и
щелочеустойчивость, все группы микроорганизмов проявляют активную
жизнедеятельность в нейтральной среде. Поэтому нейтрализация кислых и снижение pH
щелочных почв приводят к активизации микробиологических процессов,
благоприятных для возделывания культурных растений [3].
.2. Численность микроорганизмов в почвах разных
типов
Детальное изучение специфики микрофлоры
почвенных типов началось в нашей стране в сороковых годах прошлого столетия.
Сейчас работу по изучению микробных ценозов различных почв проводят во многих
научно-исследовательских учреждениях нашей страны.
Численный состав микроскопических существ почв
отличается большой динамичностью. Даже за относительное короткие промежутки
времени число микроорганизмов в почве может меняться. Это следствие динамики
температуры и влажности почвы, состояния растительного покрова и т.д. Почти во
всех почвах наблюдается большая или меньшая активизация деятельности
микроорганизмов весной. Очевидно, это связано с обогащением почв отмершей за
осенне-зимний период растительностью и достаточным увлажнением.
Кроме сезонных изменений, в численности
почвенной микрофлоры отмечаются и кратковременные флуктуации (т.е. неожиданные
изменения). О причине последних существуют разные предположения. Некоторые
исследователи допускают, что число бактерий может резко снижаться вследствие
уничтожения их фагами или простейшими. Предполагают также накопление каких-то
токсичных веществ в почве, временно подавляющие развитие определенных групп
микроорганизмов.
Однако скорее всего флуктуации определяются
неравномерным распределением микроорганизмов в почве. В связи с этим каждая
взятая проба отличается по составу микробов от другой, что создает впечатление
существенной динамики численности. Колебания численности микробов не снимают
вопроса о неодинаковой плотности заселения микроорганизмами почв различных
типов. Несмотря на колебания, легко заметить, что в одних почвах микробов
больше, в других меньше. Если ориентироваться на средние цифры, то можно
составить представление о богатстве тех или иных почв микроорганизмами.
Микробиологические анализы дают условные
показатели, но при пользовании одной и той же методикой для изучения разных
почв получают вполне сопоставимые результаты. Все применяющиеся методы (прямое
микроскопирование и посев на разные питательные среды) еще раз свидетельствуют
о большем богатстве микробами южных почв по сравнению с северными.
Не трудно предположить, что по мере перехода от
более холодного северного климата к южному микронаселение почв возрастает; во
многих южных почвах и микробиологические процессы протекают более энергично.
Вообще все микроорганизмы можно разделить на три
основные группы: собственно бактерии, актиномицеты и грибы. Также выделяются
отдельно спорообразующие бактерии. Помимо этого микроорганизмы делят на другие
группы в зависимости от разных характеристик и факторов (прежде всего по
способу питания).
Таблица 1
Численность и соотношения отдельных групп
микроорганизмов в почвах разных типов (учет методом посева)
|
Зоны
|
Почвы
|
Состояние
почв
|
Общее
число микроорганизмов, тыс. на 1 г почвы
|
Бактерии
|
Споры
(из числа бактерий)
|
Актиномицеты
|
Грибы
|
|
|
|
|
%
|
|
Тундра
и тайга Лесо-луговая Луговая степь и степь Сухая степь Пустынная степь и
пустыня
|
Тундрово-глеевые
и глеево-подзолистые Подзолы и дерново-подзолистые Черноземы Каштановые
Бурые и сероземы
|
Целинные
Окультуренные Целинные Окультуренные Целинные Окультуренные Целинные
Окультуренные Целинные Окультуренные
|
2140
4870 1080 2620 3630 4530 3480 6660 4490 7380
|
95,6
98,0 89,3 70,7 63,8 64,4 64,8 67,6 63,4 66,1
|
0,7 0,6 12,0 14,9 21,4 24,5 19,3
23,0 17,7 19,8
|
1,4 1,6 8,1 28,2 35,4 35,1 34,7
32,0 36,1 33,6
|
3,0 0,4 2,6 1,1 0,8 0,5 0,5 0,4
0,5 0,3
|
Как видно из таблицы, численность
микроорганизмов повышается от северных почв к более южным. Удельный вес
бактерий наиболее высок на севере, ближе к югу он понижается в связи
возрастанием численности других микроорганизмов. При этом процентное содержание
спорообразующих бактерий наоборот, возрастает. Содержание актиномицет гораздо
более высоко в южных почвах, чем в северных. Содержание грибов во всех случаях
низкое, хотя на севере их больше, но при этом меньше видового разнообразия.
Также можно заметить, что окультуренные почвы всех зон обычно богаче
микроорганизмами, чем целинные [1].
.3 Соотношение основных групп микроорганизмов
Эту группу еще называют зимогенной (от греч. ɀyme
- закваска). Это наиболее изученная группировка микроорганизмов в различных
почвах. Сапротрофы - микроорганизмы, разлагающие в основном легко доступные
соединения. К ним относятся прежде всего бактерии, актиномицеты и грибы. Обычно
сапротрофов учитывают методом посева на твердые, а иногда и в жидкие
питательные среды, содержащие те или иные органические вещества. Наиболее часто
используют мясо-пептонный агар и крахмало-аммиачный агар, на которых хорошо
выявляются бактерии и, в частности, актиномицеты. Для учета микроскопических
грибов чаще используют подкисленный сусло-агар, среду Чапека и т.д.
Отдельные почвы существенно различаются по
глубине микробиологического профиля. С углублением в почву количество
микроорганизмов постепенно уменьшается и меняется их состав. Снижение
численности клеток с глубиной до известной степени связано с уменьшением
количества гумуса в нижележащих слоях почвы, но прямая корреляция отсутствует.
Обычно с увеличением глубины численность микроорганизмов снижается более резко,
чем уменьшается содержание гумуса. В гумусных и нейтральных почвах
микробиологический профиль, как правило, все же более глубокий. В более
глубоких слоях относительно больше бацилл и часто актиномицетов. Это особенно
заметно в черноземах и сероземах.
Как видно из таблицы 2, в составе зимогенной
микрофлоры богато представлены бактерии, особенно неспорообразующие формы. Так,
можно считать доказанным, что гнилостные бактерии Pseudomonas
fluorescens, являющиеся
пионерами освоения органических остатков, более богато представлены в почвах
севера, где медленно идет минерализация. В почвах юга они обнаруживаются в
значительном числе лишь в течение краткого времени после внесения растительных
остатков.
Представители рода Arthrobacter,
по ряду признаков родственные актиномицетам, в большем числе встречаются в
почвах южной зоны. Они характерны для более поздних стадий распада
органического вещества и предпочитают нейтральную среду. В почвах севера очень
часто присутствует значительное количество коринебактерий (Corynebacterium).
Таблица 2
Изменение соотношения Pseudomonas
fluorescens и Arthrobacter
в различных почвах, % колоний, учтенных на питательной среде
|
Зона
|
Почва
|
Pseudomonas
|
Arthrobacter
|
|
Тундра
|
Тундровый
подзол
|
До
80
|
0-5
|
|
Лесо-луговая
|
Подзол
и дерново-подзолистая
|
До
20
|
0-5
|
|
Сухая
степь
|
Чернозем
|
До
15
|
До
10
|
|
Пустынная
степь
|
Серозем
|
До
8
|
50-60
|
Из неспорообразующих бактерий виды рода Beijetinkia
распространены только в кислых субтропических почвах (латеритах и желтоземах).
Представители рода Enterobacter
в большом количестве встречаются в лесных почвах средней полосы, а рода Spirillum
- в южной зоне.
Основательнее изучены группмровки
спорообразующих бактерий, развитие которых связано с присутствием в почве более
переработанного органического вещества. Каждому типу почв свойствен характерный
набор преобладающих видов бацилл. Другие виды здесь могут быть, но в очень
малом количестве.
В почве с энергичными мобилизационными
процессами преобладают бациллы, использующие не только азот органический, но и
минеральный азот (Bacillus
megaterium, B.
mesentericus, B.
subtilis) Наоборот, в
почвах со слабо протекающими процессами минерализации органических веществ
доминируют спорообразующие бактерии, для которых необходим органический азот (B.
cereus, B.
mycoides и др.). В этом
проявляется глубокая связь физиологии микроорганизмов со свойствами среды их
обитания.
В.Т. Емцевым детально изучена экология
спорообразуюющих азотфиксирующих бактерий рода Clostridium.
Некоторые из них, например C.
pasteurianum, в больших
количествах встречаются только в северных почвах. В почвах южной зоны
доминируют C. acetobutylicum.
При окультуривании почвы состав почвенной микрофлоры, в том числе бацилл и
клостридий, существенно меняется, появляются спорообразующие виды, свойственные
более южной зоне.
Иногда с органическими удобрениями (навоз и
компост) в почву вносят специфичные для этих удобрений бациллы. Так, в
разогревающемся при созревании навозе содержится много клеток Bacillis
mesentericus, B.
subtilis и термофильных
бактерий.
С вертикальной поясностью почв связана в
основном такая же смена бациллярных и клостридиальных форм, как и при зональном
распределении. Однако почвы вертикальной поясности нельзя считать полным
аналогами горизонтально-зональных, поэтому и в составе их микроорганизмов
имеются некоторые различия [3].
Актиномицеты
Группа актиномицетов бактерий актиномицетной
линии чрезвычайно обширна. В последние годы в изучении экологии и географии
этих микроорганизмов достигнуты большие успехи. Типичные формы, относящиеся к
аэробам и образующие мицелий, широко распространены в почве. Подобно бациллам,
стрептомицеты бедно представлены в северных почвах, но в южных их численность
резко возрастает, что подтверждается всеми методами исследования.
Широкое распространение актиномицетов в природе
определяется их устойчивостью к высушиванию, временному отсутствию питательных
веществ в среде обитания и эффективностью расселения их спор. Многие
актиномицеты в лабораторных культурах способны медленно расти на водном агаре и
других средах с низким содержанием питательных веществ. В почве активно
растущий вегетативный мицелий обнаруживается преимущественно в микрозонах с
повышенным содержанием органического вещества. Корреляция между содержанием актиномицет
в почве и количеством органического вещества в ней еще в 1958 году отмечал Н.А.
Красильников. Обладая небольшой скоростью роста, актиномицеты при попадании в
почву веществ, легкодоступных многим микроорганизмам, обычно не оказываются в
первом эшелоне r-стратегов и
доминируют лишь тогда, когда создают условия для использования сравнительно
труднодоступных субстратов. Большинство актиномицетов - нейтрофилы (оптимальные
для роста значения pH
6-8), однако, существуют сведения о присутствии в почве ацидофильных и
алкалофильных форм.
Представители почти всех известных родов
актиномицетов выделены из почвы или обнаружены в почве. Однако закономерности
распределения мицелиальных прокариот в почвах до недавнего времени были
исследованы только у представителей родов Streptomyces
и Streptoverticillium.
Введение понятия комплекса почвенных актиномицетов, структура которого отражает
пространственно-временное соотношение отдельных таксонов (родов, видов)
мицелиальных прокариотных микроорганизмов, способствовало выявлению общих
закономерностей распределения этих организмов в природных субстратах и, в
первую очередь, в почве. Структура комплекса почвенных актиномицетов
определяется составом и численностью типичных родов и видов и величиной
видового спектра. Выделить представителей всех известных в настоящее время
родов актиномицетов не представляется возможным, поскольку для выявления многих
представителей данного порядка требуются специальные методы , включающие ряд
селективных приемов, связанных со свойствами как спор, так и мицелия этих
организмов, характеризующихся сложными жизненными циклами. Обычно используют
традиционный метод поверхностного посева из разведений на плотные питательные
среды: казеин-глицериновый агар и среду с пропионатом натрия. В отдельных случаях
для выделения некоторых малоизученных родов актиномицетов применяли
предпосевную обработку почвенных образцов (нагреванием при 100 С в течение 1
часа).
Структура комплексов почвенных актиномицетов
специфична для каждого биогеоценоза. Лесной биогеоценоз характеризуется
наличием одного доминирующего рода Streptomyces
с типичными доминирующими видами только одной секции и серии. Виды почвенных
актиномицетов лесного биогеоценоза функционируют в условиях, где основными
средообразующими факторами является подстилка, невысокое содержание гумуса,
низкое значение pH
среды. Эти условия определяют заполнение основной части пространства
экологических возможностей (65%) в лесной почве только стрептомицетами секции и
серии Cinereus
Achromogenes. Показано, что
экологические ниши изученных актиномицетных популяций дерново-подзолистой почвы
не разобщены полностью, а в той или иной мере перекрываются. Виды секций и
серий Cinereus
Achromogenes И Albus
Albus имеют сходные
экологические позиции, степень перекрывания их экологических ниш более 70%. То
же можно сказать о видах секций и серий Roseus
Lavendulae-roseus
и Cinereus
Chromogenes. Если степень
перекрывания ниш рассматривать как коэффициент конкуренции видов, то
конкурентные взаимоотношения в почве наиболее вероятны между представителями
секций и серий Cinereus
Achromogenes и Albus
Albus; Roseus
Lavendulae-roseus
и Cinereus
Chromogenes.
Слабый рост актиномицетов в почвах северной зоны
может быть объяснен как замедленным темпом разложения здесь органического
вещества, так и слабой толерантностью организмов к почвенной кислотности.
Имеющиеся сведения нередко противоречивы, но несомненно, что почвы южной зоны
не только богаче актиномицетами, но и имеют более разнообразный их видовой
состав.
Очевидно, некоторые актиномицеты распространены
чрезвычайно широко (группы albus,
griseus, globisporus,
violaceus и аспоргенные
формы albus). Однако
группы violaceus
и аспорогенные albus
богаче представлены в южных почвах. Некоторые группы актиномицетов (fradis,
flavus, cromogenes,
rubroaurantiacus) в заметных
количествах обнаружены в серых лесных почвах. К югу их численность также
возрастает. Создается впечатление, что группа verticillatus
и аспорогенные формы flavus
и chromogenes также тяготеет к
наиболее южным почвам.
Смена состава актиномицетов в разных почвах
хорошо выявляется на примере пигментированных культур. Они гораздо больше
распространены в почвах, формирующихся в условиях теплого климата. Установлено,
что и виды рода Acinomadura
широко распространены повсеместно, но их видовое разнообразие значительно
богаче в южных почвах [2].
Грибы и дрожжи
Северные почвы, имеющие кислую реакцию, наиболее
богаты грибами. Вообще в разлагающейся растительной массе и в верхних слоях
почвы их биомасса значительно больше бактериальной. Учет массы грибного мицелия
в разных почвах, проведенной Т.Г. Мирчинк, показал, что в тундре на 1 г почвы
приходится 4 мг мицелия грибов, в лиственных лесах - до 1 мг, а в почвах южной
зоны - 0,4-0,7 мг.
В почвах южной зоны родовой и видовой состав
микроскопических грибов более разнообразен, чем в северных. В первых доминирует
представители рода Aspergillus,
а во вторых - Penicillium.
По имеющимся данным, род Penicillium
в северных почвах представлен 35 - 40 видами, а в южных - лишь 10-15. Обратная
картина наблюдается для грибов рода Aspergillus:
в северных почвах в небольшом числе встречаются три-пять видов рода, в южных -
15 - 20.
Северные почвы беднее, чем южные, грибами рода Fusarium,
которые особенно обильно размножаются в каштановых почвах и сероземах.
Некоторые виды, например Fusarium
sambicinum, свойственны
только щелочным почвам. Мукоровыми грибами богаты почвы северных районов,
однако некоторые роды (Choanephora,
Cunninghamella, Rhizopus)
приурочены к южным почвам.
В почвах обычно встречаются грибы с
темнопигментированным мицеллием (Dematium,
Cladosporium, Macrosporium,
Alternaria и т.д.). Их
экология плохо изучена, но отмечается, что представители рода Dematium
более распространены в почвах с малоактивным мобилизационными процессами, т.е.
в основном в северной зоне, а виды рода Alternaria
чаще встречаются в окультуренных почвах.
Выявлено, что одни виды рода Mortierella
(M. vanaceae,
M. usabellina)
распространены в кислых почвах, другие (M.
alpine, M.
dichotoma) - в нейтральных.
Установлены индикаторные микроскопические грибы для определения типов почв.
По данным И.П. Бабьевой, в тундре при большой
пестроте почвенного покрова основная часть дрожжей сосредоточена на мхах и
торфе. Доминантные виды дрожжей в тундровых почвах не относятся к типичным
педобионтам и более характерны для живых и отмирающих частей растений. Они
представлены базидиомицетами (роды Rhodotorula,
Rhdosporidium, Cryptococcus).
В лесных биогеоценозах много дрожжей имеется в
подстилке. Они составляют группу базидальных грибов (виды родов Candida,
Trichosporon и др.). В
минеральных горизонтах почвы дрожжей значительно меньше. Здесь доминируют
типичные педобионты - из аскоспоровых грибов Lipomyces
starkeyi, из базидомицетов
- виды Candida
и Cryptococcus.
В степном биогеоценозе травяной опад
также богат дрожжами. Здесь встречается до 14 видов родов Cryptococcus,
Aureobasidium, Rhodosporidium
и др. Для почвы характерно доминирование Lipomyces
tetrasporium.
В биогеоценозах полупустынь и пустынь на
растительности доминируют дрожжи родов Sporodiobolus,
Tilletiopsis и Sporobolomyces,
образующие баллистоспоры, рассеивающие токами воздуха и имеющие в жизненном
цикле стадии, устойчивые к засухе, - хламидоспоры. Численность их видов
невилика. Дрожжи в указанных регионах приурочены не к поверхностному слою
почвы, а обитают на некоторой глубине - ниже 20 см. Род Lipomyces
в почве пустынь отсутствует. Доминирую криптококки [3].
Целлюлозоразрушающие микроорганизмы
Процесс распада целлюлозы, вызываемый как
бактериями, так и грибами, представляет существенный интерес для познания
почвообразования. Большая часть растительных остатков состоит из целлюлозы.
Изучение микроорганизмов, разрушающих ее, показало, что их состав существенно
меняется в разных почвах. В северных почвах (тундра) этот процесс связан с
деятельностью некоторых медленно растущих грибов, относящихся главным образом к
родам Dematium
и Penicillium. В зоне тайги в
составе данной группы появляются микобактерии и виды рода Cellvibrio.
В южных почвах грибы в значительной степени вытесняются как уже указанными видами
бактерий, так и представителями рода Cytophaga
и др. В заметных количествах здесь появляются грибы рода Chaetomium.
Биоценоз описаного состава в отличие от северного разрушает целлюлозу быстро.
Изучение особенностей физиологии разных групп
целлюлозо-разлагающихся микроорганизмов позволило объяснить их своеобразную
экологию. Особенности выражаются главным образом в требовательности к
источникам азотного питания. Микроорганизмы северных почв (в основном грибы),
хотя и медленно, растут и на бедных азотным основанием средах. Микроорганизмы
южных почв, разрушающие целлюлозу, нуждаются в высоком уровне азотного питания.
На юге процесс минерализации азота протекает значительно энергичнее, что
благоприятствует развитию микроорганизмов, более требовательных к условиям
среды [3].
.3.2 Олиготрофные микроорганизмы
Олиготрофы - микроорганизмы
<#"791664.files/image001.gif"> 100=90%=0,9 от одного грамма почвы.
100=90%=0,9 от одного грамма почвы.
После взбалтывания мы взяли колбу c суспензией,
где 1 мл соответствует разведению  , и сделали последующие разведения -
, и сделали последующие разведения -
 ;
; 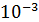 ;
; 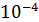 ;
;  . Для этого из каждого предыдущего
разведения отдельной стерильной пипеткой брали 1 мл суспензии и переносят в
отдельную пробирку с 9 мл воды. Всякий раз пипетку ополаскивали и отстаивали, а
последующие пробирки встряхивали по 1 минуте.
. Для этого из каждого предыдущего
разведения отдельной стерильной пипеткой брали 1 мл суспензии и переносят в
отдельную пробирку с 9 мл воды. Всякий раз пипетку ополаскивали и отстаивали, а
последующие пробирки встряхивали по 1 минуте.
Из полученных разведений делают
посев на плотные и жидкие среды. Набор сред зависит от задач
бактериологического анализа почвы. В данном случае я использовал глубинный
метод посева. Для этого взяли 8 стерильных чашек Петри, вносим в каждую из них
по 1 мл суспензии. Для МПА сделали четыре образца с разведением 
 по 2 на каждое разведение. Для СА
сделали то же самое, только разведение было и .
по 2 на каждое разведение. Для СА
сделали то же самое, только разведение было и .
Когда численность отдельных групп
микроорганизмов в почве небольшая, их выявляют методом обрастания комочков
почвы по Виноградскому. Для этого мы использовали две среды - Эшби и Гетчинсона.
По поверхности сред раскладывают по трафарету 50 комочков почвы диаметром 1-2
мм. Чашки помещают во влажную камеру и ставят в термостат. Если в комочках
почвы находятся соответствующие микроорганизмы, то они начинают развиваться и
формируют вокруг комочков колонии. Затем вычисляют количество обросших комочков
почвы (в % от исходного числа). После определенного срока инкубации при 28-30 °C культуры
анализируют [4].
.2 Результаты исследований
.2.1 Микроорганизмы, использующие
органическую форму азота (МПА)
Мясо-пептонный агар - плотная
универсальная питательная среда, состоящая из мясо-пептонного бульона с
добавлением 0,5 - 2% агара и применяемая для выявления микроорганизмов,
использующие органические формы азота. После инкубации мы вынули чашки с засеянными
средами из термостата и подсчитали число колоний, которые отражают число живых
клеток микроорганизмов в почве. После подсчета количества всех колоний на чашке
мы их сгруппировали по культуральным признакам: характеру роста на агаре,
цвету, краю, форме, размеру (приблизительный) и консистенции колонии - для
качественного анализа сапротрофной микрофлоры на МПА. Мы выбрали четвертое
разведение, потому что колонии на среде были выделены более четко и не
перекрывались друг другом.
Доминирующие колонии были
бледно-кремового цвета, округлой формы, с матовым блеском, ровными краями,
однородной структурой и плотной консистенцией.
Таблица 1
Численная характеристика
микроорганизмов на МПА
|
Разведение
|
Общее
число КОЕ
|
Число
доминирующих КОЕ
|
Количество
микроорганизмов в 1 г
|
Доминирующие
формы (численность в 1 г)
|
Содержание
доминирующей группы в %
|
56
|
45
|
12444444
|
10000000
|
80
|
Расчеты
количество микроорганизмов в 1 г абсолютно сухой
почвы:
56×20× / 0,9 = 12444444.
/ 0,9 = 12444444.
Количество микроорганизмов
доминирующей формы в 1 г абсолютно сухой почвы: 45×20×/ 0,9 =10000000.
Доля доминирующих форм в общей
численности микроорганизмов: 10000000/12444444×100% = 80 %.
Далее мы сделали препарат и под
микроскопом определили, что доминирующие бактерии относятся к роду Bacillus.

Рис.1 x 1600
Описание доминирующей формы под
микроскопом
Форма клетки: палочковидная;
Характер расположения клеток: поодиночке или группами; Способность к
спорообразованию: спорообразующие.
.2.2 Микромицеты (СА)
Сусло-агар применяют для первичного
посева материала с целью выделить грибы. Учет проводят также, как и в
предыдущем случае. Разведение . Доминирующие колонии были с
округленной формой, размером 1,2 см, зеленого цвета, мучнистого блеска, с
шероховатою поверхностью, зубчатым краем и однородной структурой.
Таблица 2
Численная характеристика микроорганизмов на СА
|
Разведение
|
Общее
число КОЕ
|
Число
доминирующих КОЕ
|
Количество
микроорганизмов в 1 г
|
Доминирующие
формы (численность в 1 г)
|
Содержание
доминирующей группы в %
|
|
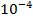
|
13
|
9
|
2888888
|
2000000
|
69
|
Расчеты
количество микроорганизмов в 1 г абсолютно сухой
почвы:
13×20×/ 0,9 =2888888.
Количество микроорганизмов
доминирующей формы в 1 г абсолютно сухой почвы: 9×20×/ 0,9 =2000000.
Доля доминирующих форм в общей
численности микроорганизмов: 2000000/2888888×100% = 69 %.
Далее под микроскопом удалось
определить, что доминирующие грибы - Penicillium glaucum.

Рис. 2 x 64
Описание грибов под микроскопом
Это сумчатые грибы; способность к
спорообразованию: спорообразующие; мицелий: хорошо развитый, многоклеточный;
образуют конидии, напоминающие кисть руки.
.2.3 Азотфиксирующие бактерии (среда
Эшби)
Среда Эшби не содержит ни
минеральных, ни органических форм азота, поэтому ее используют для учета
азотфиксирующих микроорганизмов путем раскладывания комочков почвы. При учете
численности микроорганизмов мы посчитали обросшие колониями комочки почвы и,
зная общее количество комочков, посчитали процентное содержание обросших
комочков.
Таблица 3
Учет обросших комочков после посева
|
Общее
число комочков
|
Число
обросших комочков
|
Процентное
содержание
|
|
50
|
48
|
96
|

Рис. 3
Далее мы сделали препарат и под микроскопом
определили, что это azotobacter
chroococoom
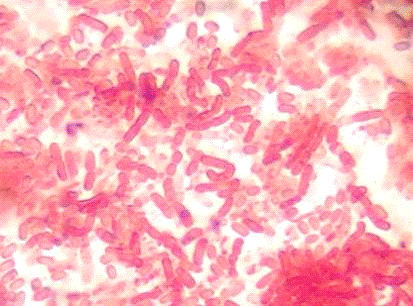
Рис. 4 x
1600
Описание бактерий под микроскопом
Форма клетки: палочки или кокковидные;
способность к спорообразованию: неспорообразующие; характер расположения:
поодиночке
.2.4 Целлюлозоразрушающие бактерии (среда
Гитченсона)
На среде Гетчинсона выявляют
целлюлозоразрушающие микроорганимзы. Эта среда не содержит никаких источников
углерода, кроме целлюлозы в виде фильтровальной бумаги. Другие микроорганизмы,
не разлагающие целлюлозу, не могут развиваться, так как не имеют альтернативных
источников углерода.
Таблица 4
|
Общее
число комочков
|
Число
обросших комочков
|
Процентное
содержание
|
|
50
|
43
|
86
|
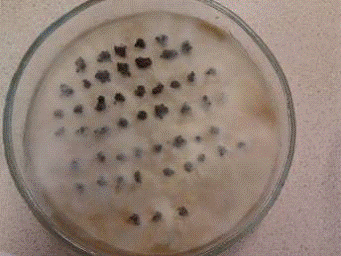
Рис. 5
Далее мы сделали препарат и под микроскопом
определили, что это polyangium
Рис. 6 x
1600
Плодовые тела Polyangium
.3 Выводы
В полученном образце дерново-подзолистой почвы в
ходе микробиологического анализа были обнаружены микроорганизмы рода Bacillus,
как доминирующий вид, играющий одну из главных ролей в процессе минерализации
органических соединений азота. Так же были обнаружены различные формы
микроскопических грибов (Penicillium), что свидетельствует о слегка повышенной
кислотности почвенного образца. Также в исследуемом образце был найден
Azotobacter, а значит процесс азотфиксации протекает достаточно интенсивно.
Наконец, из целлюлозоразрушающих бактерий был обнаружен Polyangium.
Таким образом, с точки зрения
микробиологического анализа, данный образец почвы подходит для выращивания
сельскохозяйственных культур, так как в почве протекают все необходимые для
развития растений микробиологические процессы, а именно - минерализация
органического вещества, азотфиксация.
Список использованной литературы
. Добровольская
Т.Г. Структура бактериальных сообществ почв. М.:
“Академкнига”,
2002.
282 с.
2. Звягинцев
Д.Г., Бабьева И.П., Зенова Г.М. Биология почв. М.:
МГУ,
2005.
445 c.
3. Мишустин
Е.Н., Емцев В.Т. Микробиология. М.: Агропромиздат, 1987. 368 с.
. Теппер
Е. З., Шильникова В. К., Переверзева Г. И. Практикум по микробиологии. М.:
Дрофа,
2004 - 256 c.: ил.
5. www.wikipedia.ru