Проблема популяции изменчивости видов рода Hedera L. на Западном Кавказе
МИНИСТЕРСТВО
ОБРАЗОВАНИЯ И НАУКИ РОССИЙСКОЙ ФЕДЕРАЦИИ
Федеральное
государственное бюджетное образовательное учреждение высшего профессионального
образования
«Кубанский
государственный университет»
Географический
факультет
Кафедра
геоэкологии и природопользования
дипломная работа
ПРОБЛЕМА ПОПУЛЯЦИОННОЙ ИЗМЕНЧИВОСТИ ВИДОВ РОДА HEDERA
L. НА ЗАПАДНОМ КАВКАЗЕ
Работу выполнила Нестерова Юлия
Сергеевна.
Нормоконтролер доц., канд.
биол. наук Пикалова Н.А.
Краснодар,
2013
Содержание
Введение
.
История изучения видов рода Hedera
L. по известным
флористическим сводкам
.
Виды рода Hedera
L. на Западном
Кавказе
.1
Изучение экологии и географического распространения вида H.
helix L.
subsp. caucasigena
.2
Изучение экологии и географического распространения вида H.
colchica
C. Koch
.
Общая природно-климатическая характеристика Западного Кавказа
.
Популяционное исследование видов рода Hedera
L. на Западном
Кавказе
.1
Биометрические показатели морфологических признаков листа Hedera
L
.2
Влияние абиотических факторов на популяционную изменчивость листа видов рода Hedera
L
.2.1
Влияние абиотических факторов на морфологическую изменчивость листа H.
helix L.
subsp. caucasigena
.2.2
Сравнительный анализ морфологии листа подвида H.
helix L.
subsp. caucasigena
c средиземноморским
видом H. helix
L
.2.3
Влияние абиотических факторов на морфологическую изменчивость листа H.
colchica
C. Koch
Заключение
Список
используемых источников
Приложение
Введение
Уникальность и высокое биологическое
разнообразие Западного Кавказа представляет большой интерес. В настоящее время
стоит проблема сокращения биологического разнообразия и поэтому актуально
изучение каждого вида в отдельности.
Объектом исследования данной работы являются
виды рода Hedera
L.L. - род семейства
Araliaceae, насчитывающий по оценкам разных специалистов около 15 видов
вечнозеленых кустарников [Бородина, 1966, с. 497].
Согласно современным данным, представленных в
Конспекте флоры Кавказа, выделяют следующие виды Hedera
L.: H.
chrysocarpa
Walsh, H.
helix L.
и подвид H. helix
L. subsp.
caucasigena, H.
colchica (C.Koch)
C.Koch,
H. pastuchowii
Woronow [Конспект флоры
Кавказа, 2009, т. 3 (1), с. 52 - 53].
H. colchica
(C.Koch)
C.Koch,
и подвид H. helix
L. subsp.
сaucasigena - основные виды
Hedera L. на Западном Кавказе. Морфологические различия наблюдаются как между
видами, так и между представителями одного вида разных популяций. Такая
неопределенность связана, прежде всего, с большой изменчивостью листьев, по
признакам которых обычно проводится определение.
Целью исследования стало изучение популяционной
изменчивости листа видов рода Hedera
L. на Западном
Кавказе.
Для достижения поставленной цели решались
следующие задачи:
проанализировать биологическое разнообразие
видов рода Hedera
L. произрастающих в
пределах Западного Кавказа по известным флористическим сводкам;
изучить экологию и географическое
распространение Hedera
L. на Западном
Кавказе;
произвести сбор физико-географических сведений о
местах сбора видов рода Hedera
L.;
произвести анализ биометрических показателей морфологических
признаков листа;
установить влияние абиотических факторов на
популяционную изменчивость видов H.
helix L.
subsp. сaucasigena
и H. colchica
(C.Koch)
C.Koch
(длина черешка, количество лопастей, глубина лопасти) и изучит морфометрические
особенности разных популяций видов Hedera L.
произвести сравнительный анализ подвида
Западного Кавказа (H. helix
L. subsp.
caucasigena) с видом H.
helix L.,
произрастающего в Средиземноморье.
Актуальность работы заключается в том, что
морфологические различия Hedera
L. наблюдаются как
между видами, так и между представителями одного вида разных популяций. Такая
неопределенность связана, прежде всего, с большой изменчивостью листьев, по
признакам которых обычно проводится определение. Морфология листьев H.
helix L.
subsp. caucasigena
имеет значительные отличия от средиземноморских видов H.
helix L.,
что обуславливает возможность выделения подвида H.
helix L.
subsp. caucasigena
в самостоятельный вид, встречающийся на Западном Кавказе.
Новизна исследований заключается в том, в
дипломной работе впервые для региона проводится сравнительный анализ
изменчивости признаков листа различных популяций Hedera
L. в зависимости от
условий местообитания и представлены их морфометрические характеристики. А
также проводится сравнительный анализ изменчивости листьев подвида H.
helix L.
subsp. caucasigena
с средиземноморским видом H.
helix L.
1. История изучения видов рода Hedera
L. по известным
флористическим сводкам
Для изучения данного рода было обработано 4
флористические сводки: А.А. Гроссгейм «Определитель растений Кавказа»
[Гроссгейм, 1949], И.С. Косенко «Определитель высших растений Северо-Западного
Кавказа и предкавказья» [Косенко, 1970], А.И. Галушко «Определитель флоры
Северного Кавказа» [Галушко, т. 3, 1978] А.С. Зернов «Определитель флоры
Северо-Западного Кавказа» [Зернов, 2006] .
Распространение видов рода Hedera
L. на Западном
Кавказе описано в соответствии с данными, представленными А.Л. Тахтаджяном в
«Конспекте флоры Кавказа» том 1 (2003).
По Гроссгейму на Кавказе выделены следующие виды
рода Hedera
L. [Гроссгейм, 1949,
с. 204]:
1. H.
chrysocarpa
Walsh.
Культурное растение. Зрелая костянка желтая.
Цвет листьев желтовато-зеленый. Жизненная форма - кустарник. Родиной этого вида
является Средиземье.
. H.
helix L.
Листья более мелкие, без запаха, на нецветущих
побегах угловато трех-пятилопастные. Жизненная форма - кустарник.
Распространение: Западный Кавказ - Восточный Кавказ - Черкесия - Западное
Закавказье - Центральное Закавказье - Карабах. Произрастает в лесах, по ущельям,
оврагам.
. H.
colchica
C. Koch
Листья широко-треугольные до округлых, в живом
виде с характерным слегка мускатным запахом. Распространение: Западный Кавказ -
Западное Закавказье - Центральное Закавказье (редко) - Юго-Западное Закавказье.
До среднего горного пояса. Произрастает на стеблях деревьев, по откосам и т.д.
. H.
Pastuchowii
G. Wor.
Листья продолговато-ромбические до
продолговато-ланцетных, без запаха. Распространение: Восточный Кавказ -
Центральное Закавказье (на востоке) - Талыш. Простирается до среднего горного
пояса. Произрастает на деревьях.
По Косенко [Косенко, 1970, с. 248] на Западном
Кавказе выделено два вида Hedera
L:
1. H.
colchica
C. Koch
Опушение из бурых чешуевидных, по краям лучистых
волосков. Листья вегетативных побегов обычно яйцевидные, с клиновидным или
яйцевидным основанием, вверху заостренные. Зонтики 12-35-цветковые. Лепестки
сверху зеленовато-оранжевые. Высота до 30 м. Древесная лиана. Произрастает во
влажных лесах, на стволах деревьев, на скалах и лесных откосах. Распространена
на кубанском склоне в верховьях Лабы и Белой, в верхнем горном поясе; на
черноморском склоне от нижних до средних горных поясов. Используется для
вертикального озеленения.
. H.
helix L.
(H. сaucasigena
Pojark.)
Опушение из сероватых звездчатых волосков.
Листья побегов вегетативных 3-5-7-лопастные, с сердцевидным основание. Зонтики
8-12-цветковые. Высота до 20 м. Древесная лиана. Произрастает в лесах, на
стволах деревьев, по скалам и лесным откосам. От нижнего до среднего горных
поясов.
На Северном Кавказе А. И. Галушко выделил
следующие виды рода Hedera
L. [Галушко, 1978,
с. 298]:
1. H.
сaucasigena
Pojark.
2. H.
colchica
C. Koch
3. H.
Pastuchowii
G. Wor.
По А.С. Зернову на Северо - западном Кавказе,
выделяются два вида рода Hedera
L. [Зернов, 2006, с.
405]:
1. H.
colchica
C. Koch
Почки, молодые стебли и черешки молодых листьев покрыты звездчато-чешуевидными
волосками, с короткими лучами. Листья вегетативных побегов цельные или неясно
трехлопастные, с верхней стороны обычно глянцевые. Блестящие. Растение с
приятным запахом. Деревянистая лиана. Произрастает в тенистых лесах и ущельях.
Фанерофит. Распространен в следующих районах Северо-западного Кавказа:
Сочинский, Хадыженский, Майкопский.
2. H.
helix
L.
Почки, молодые стебли и черешки молодых листьев
покрыты звездчатыми волосками с длинными лучами. Листья вегетативных побегов
3-5-лопастные, с верхней стороны обычно матовые. Растение без приятного запаха.
Произрастает в лесах, иногда на щебляке. Деревянистая лиана. Фанерофит.
Распространен в следующих районах Северо-западного Кавказа: Новороссийский,
Сочинский, Абинский. Хадыженский, Майкопский.
По имеющимся данным проведем анализ по
количеству видов, произрастающих на территории Западного Кавказа. Сравнивая
данные флористов по региону (Гроссгейм, 1949; Косенко, 1970; Галушко, 1978;
Зернов, 2006) констатируем соответствия в таксономии видов рода Herdera
L. (таблица 1).
Таблица 1 - Количества видов Hedera
L. на Западном
Кавказе по известным флористическим сводкам
|
Автор
|
Вид
|
|
А.А.
Гроссгейм
|
2
|
|
И.С.
Косенко
|
2
|
|
А.С.
Зернов
|
2
|
|
А.И.
Галушко
|
2
|
Флористы выделяют два основных вида: H.
helix L.
и H. colhica
C. Koch.
В определителе Косенко [Косенко, 1970, с. 248] представлен подвид H.
helix L.
- H. caucasigena
Pojark. А. И. Галушко
выделяет H. caucasigena
Pojark. в самостоятельный
вид, произрастающий на Западном Кавказе [Галушко, 1978, с. 297]. Согласно
данным, представленных во Флоре СССР, H.
helix L.
subsp. caucasigena
является кавказской расой H.
helix L.
и в свою очередь имеет ряд отличий от вида. H.
caucasigena отличается от H.
helix мелкими зонтиками,
сидящих на тонких ножках и мелкими цветками. На стерильных побегах у нее
преобладают стреловидные листья, трехлопастные [Флора СССР, 1950, т. 16, с.17].
Согласно современным данным, представленных в
Конспекте флоры Кавказа, выделяют следующие виды Hedera
L.: H.
chrysocarpa
Walsh, H.
helix L.
и подвид H. helix
L. subsp.
caucasigena, H.
colchica (C.
Koch) C.
Koch, H.
pastuchowii
Woronow. Общий ареал и
географическое распространение указано в соответствии с районированием
предложенным для «Конспекта флоры Кавказа» Ю.Л. Меницким (1991). Помимо этого,
для каждого вида составлена карта иллюстрирующая картину его географического
распространения на Кавказе (рисунок 1) [Конспект флоры Кавказа, 2008, т. 1, с.
52-53].
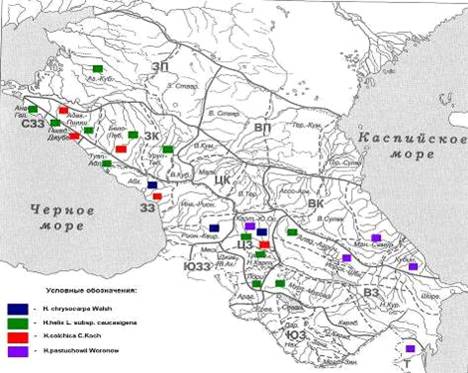
Рисунок 1 - Карта районов флоры Кавказа
(Меницкий, 1991)
Примечание - ЗП - Западное Предкавказье:
Аз.-Куб. - Азово-Кубанский, З.Ставр. - Западноставропольский; ВП - Восточное
Предкавказье: В. ставр. - Восточноставропольский, Тер.-Кум. - Терско-Кумский,
Тер.-Сулак. - Терско-Сулакский; ЗК - Западный Кавказ: Адаг.-Пшиш. -
Адагум-Пшишский, Бело-Лаб. - Бело-Лабинский, Уруп-Теб. - Уруп-Тебердинский,
В.Куб. - Верхнекубанский; ЦК - Центральный Кавказ: В.Кум. - Верхнекумский,
Малк. - Малкинский, В. Тер. - Верхнетерский; ВК - Восточный Кавказ: Ассо-Арг. -
Ассо-Аргунский, В.Сулак. - Верхнесулакский, Ман.-Самур. - Манас-Самурский,
Кубин. - Кубинский;СЗЗ-Северо-Западное Закавказье: Анап.-Гел. -
Анапа-Геленджикский, Пшад.-Джубг. -Пшадско-Джубгский; ЗЗ - Западное Закавказье:
Туан.-Адл. - Туапсе-Адлерский, Абх. - Абхазский, Инг.-Рион. - Ингури-Рионский,
Рион.-Квир. - Рионо-Квирильский, Адж. - Аджарский; ЦЗ - Центральное Закавказье:
Карт.-Ю.Ос. - Карталинско-Югоосетинский, Триал.-Н.Карт. - Триалетско-Нижнекарталинский,
Лори - Лорийский; ВЗ - Восточное Закавказье: Алаз.-Агрич. -
Алазань-Агричайский, Ширв. - Ширванский, Иорск.-Шек. - Иорско-Шекинский,
Мурк.-Муровд. - Мургуз-Муровдагский, Н.Кур. - Нижнекурильский, Караб. -
Карабахский; ЮЗЗ - Юго-Западное Закавказье: Месх. - Месхетский, Джавю-В.Ах. -
Джавахетско-Верхнеахурянский, Араг. - Арагатский; ЮЗ - Южное Закавказье: Ерев.
- Ереванский, Севан.- Севанский, Дар. - Дарелегисский, Нах. - Нахичеванский,
Занг. - Зангезурский, Мегр.-Зан. - Мегри - Зенгеланский, Ю. Караб. -
Южнокарабахский; Т - Талыш [Конспект флоры Кавказа, 2008, с.21].
2. Виды рода Hedera
L. на Западном
Кавказе
На территории Западного Кавказа произрастает два
вида рода Hedera L.: H.
helix L.
subsp. caucasigena
и H. colchica (C.Koch) C.Koch.
.1 Изучение экологии и географического
распространения H.helix
L. subsp.
сaucasigena
Описан из Западного Закавказья. Typus:
«Сухум. Округ, в лесах с. Юрьевского в 25 верстах от Сухума, на деревьях»
[Конспект флоры Кавказа, 2009, т. 3 (1), с. 52].. helix L. subsp. сaucasigena
не требователен к почве, влаголюбив, переносит значительные морозы,
теневынослив, опыляется насекомыми (энтомофил), плоды и семена распространяются
птицами (орнитохор), обитает в среде с более или менее достаточным, но не избыточным
увлажнением почвы (мезофит), растет при сравнительно низких температурах
окружающей среды (рисунок 2).

Рисунок 2 - H. helix L. subsp. сaucasigena (фото
Ю. Нестерова)
Лазящая лиана до 20 м высотой. От H. helix L.
отличается следующими морфологическими особенностями: на стерильных побегах
всегда имеются в большом, а часто и в преобладающем числе Продолговатые листья
стреловидной формы с длинно вытянутой средней лопастью и короткими двумя, реже
четырьмя нижними лопастями. Прочие листья этих побегов широкосердцевидные,
угловато, большей частью трехлопастные, реже пятилопастные, с короткой средней
лопастью; лопасти большей частью тупые, реже острые; листья лазящих стерильных
побегов также большей часть трёхлопастные яйцевидно или округло сердцевидные;
на плодущих побегах листья всегда цельные, более мелкие чем у H. helix L., до 7
(9) см длиной и 4.5-6 (6.5) см шириной, и в общем более узкие, в очертании
яйцевидные с усеченным основанием или ромбически-яйцевидные с клиновидным
основанием, часто с примесью более узких ланцетовидных и ланцето-ромбических;
края гладкие или волнистые. Общее соцветие более удлиненное и узкое, иногда в
виде продолговатой кисти, из (3) 5-12 зонтиков, изредка зонтики одиночные;
зонтики более мелкие, чем у H. helix L.: верхушечные и одиночные 25-27 (32) мм
в диаметре, боковые (15) 17-23 (27) мм в диаметре, из меньшего числа цветков,
полушаровидные, редко и главным образом. Верхушечные шаровидные; ножки зонтиков
2-3.8 мм длиной, более тонкие, 0.6-0.8 мм в поперечнике; цветоножки тонкие, 5-9
(12) мм длиной, при плодах часто заметно удлиняющиеся, 8-15(18) мм длиной;
опушение соцветия не столь плотное, из звездчатых, 5-6 лучевых волосков с малой
примесью 7-8 лучевых; цветы более мелкие, с лепестками 2-2.5 мм, изредка до
трех мм длиной и 1.5 мм шириной; плоды 5-9 мм в диаметре, обычно с 2-4 (5)
семенами. Цветет в сентябре - ноябре; зрелые плоды весной и летом (апрель -
июнь) следующего года.
Общее распространение.: Балкано-Малоазиатский
(Лазистан). Описан из с. Юрьевского близ Сухуми [Флора СССР, 1950, т. 16, с.
13-14].
На Кавказе имеет следующее распространение
(Карта районов флоры Кавказа (Меницкий, 1991)): Западное Предкавказье. На
западе ограничено побережьем Черного и Азовского морей, на востоке водоразделом
рек Кума и Калаус и проходит через населенные пункты Курсавка - Светлоград -
Ипатово - Дивное. Район включает лесостепные территории правокалаусских высот.
Южная граница проходит южнее р. Кубани по линии ст. Варениковская - р. Адагум -
ст. Северская - г. Белореченск - ст. Темиргоевская - г. Невинномысск - ст.
Курсавка. ЗП включает в себя Азово-Кубанский район, восточная граница которого
совпадает с западной административной границей Ставропольского края. Район
включает южную часть Ростовской области и северную равнинную часть
Краснодарского края; Западный Кавказ. Западная граница проходит по западным
отрогам Большого Кавказа (меридиан г. Анапы). H. helix L. subsp. сaucasigena
встречается в Адагум-Пшишском районе, восточная граница которого проходит по
водоразделу рек Пшиши Пшеха, и Бело-Лабинском районе [Конспект флоры Кавказа,
2009, т 3 (1), с. 23].
.2 Изучение экологии и географического
распространения H. colchica
C. Koch
H.
colchica
C. Koch, 1859,
Wochenschr. Gärtn.
Pflanzenk.
2:74
- H.
helix
L.
var. colchica
C.Koch,1842,
Linnaea, 16: 365. - H. rhombifolia Rupr.
ex Regel, 1860.
Описан с
Кавказа
без
указания
местообитания(“in
itinere per Caucasum, Georgiam, Armeniaque…”) [Конспект
флоры Кавказа, 2009, т. 3 (1), с. 53].. colchica плохо переносит сухость
воздуха. Опыляется насекомыми (энтомофил), плоды и семена распространяются
птицами (орнитохор), теневыносливый, мезофит, мезотерм [Литвинская, 1993, с.
4].
Крупнолистная лиана с тонкими стерильными
побегами, несущими ползучие короткие корни-присоски, при помощи которых она
взбирается на деревья и скалы, и с более толстыми, до 1 см диаметром плодущими
побегами, усаженными, особенно в молодости, желтыми чешуевидными многолучевыми
волосками (рисунок 3). Листья грубые, плотнокожие, матовые, темно-зеленые,
сверху голые, снизу с рассеянными чешуевидными волосками, в старости иногда
отсутствующими, до 20 см длиной и 17 см шириной; на стерильных побегах листья
большей частью яйцевидные или округло яйцевидные, реже ромбически-яйцевидные с
клиновидным, сердцевидным или усеченным основанием и обычно коротко
приостренной верхушкой, чаще цельные, реже трёхлопастные, изредка
пятилопастные; на плодущих побегах листья в очертании от яйцевидных и
ромбически-яйцевидных до ланцетовидных и ланцетно-ромбических или узко-ромбических,
коротко или длинно заостренные; черешки варьируют от равных по длине пластинки
или даже несколько превышающих ее до коротких, едва равных 1/3
пластинки. Зонтики шаровидные из 12-35 цветков, одиночные или собранные в виде
кисти в числе 3-8, на ножках 2-3 см длиной при основании с маленькими
треугольными пленчатыми листьями; все части соцветия, включая завязь и наружную
сторону лепестков, густо усажены желто-бурыми, многолучевыми чешуевидными
волосками; цветоножка 4-7 мм длиной при плодах до 17 мм длиной. При основании с
малыми чешуевидными, 1-2 мм длиной перепончатыми опадающими прицветниками.
Цветки обычно обоеполые, но наблюдаются и тычиночные с недоразвитой завязью;
чашелистики развитые, 1-1.5 мм длиной, яйцевидно-треугольные, острые; лепестки
зеленовато-желтые, 4-4.5 мм длиной и 2-2.5 мм шириной. Продолговато-яйцевидные,
заостренные; тычинки немного короче лепестков с овальными пыльниками 1.75 мм
длиной, и 1мм шириной; стебель 1.5 мм длиной; плоды черные, до 10-13 мм в
диаметре, с 2-5 семенами. Цветение сентябрь - октябрь; плоды созревают в
течение весны и лета следующего года [Флора СССР, 1950, т. 16, с.5-6].
Общее распространение: Балк.-Малоаз. ( Лазистан,
Трапезонд), Арм.-Курд. (Артвинский округ). Описан из Колхиды. Тип был в
Берлине.
На Кавказе имеет следующее распространение
(Карта районов флоры Кавказа (Меницкий, 1991)): Западное Предкавказье. На
западе ограничено побережьем Черного и Азовского морей, на востоке водоразделом
рек Кума и Калаус и проходит через населенные пункты Курсавка - Светлоград -
Ипатово - Дивное. Район включает лесостепные территории правокалаусских высот.
Южная граница проходит южнее р. Кубани по линии ст. Варениковская - р. Адагум -
ст. Северская - г. Белореченск - ст. Темиргоевская - г. Невинномысск - ст.
Курсавка.

Рисунок 3 - H.
colchica
C.Koch
(фото Ю. Нестерова)
ЗП включает в себя Азово-Кубанский район,
восточная граница которого совпадает с западной административной границей
Ставропольского края. Район включает южную часть Ростовской области и северную
равнинную часть Краснодарского края; Западный Кавказ. Западная граница проходит
по западным отрогам Большого Кавказа (меридиан г. Анапы). H. colchica C.Koch
встречается в Адагум-Пшишском районе, восточная граница которого проходит по
водоразделу рек Пшиш и Пшеха, и Бело-Лабинском районе, восточную границу
которого на юге образуют водоразделы рек Большая Лаба и Большой Зеленчук, а на
севере (до г. Армавира) - рек Синюха и Уруп и на Северо-Западном Закавказье,
территории с северо-востока ограниченной водоразделом западной оконечности
Главного Кавказского хребта, с юго-запада - побережьем Черного моря;
простирается между г. Анапой и г. Туапсе. Анап -Геленджикский район занимает
сектор территории между г. Анапа и с. Джанхот. Пщадско-Джубгский район занимает
сектор территории от с. Джанхот до г. Туапсе [Конспект флоры Кавказа, 2009, т.
3(1), с. 23].
3. Общая природно-климатическая характеристика
Западного Кавказа и районов сбора видов рода Hedera
L
Западный Кавказ - это площадь Кавказских гор от
Тамани до Эльбруса. Часть Западного Кавказа от Анапы до Фишт характеризуется
низкогорным и среднегорным рельефом (так называемый Северо-Западный Кавказ).
Далее к востоку от Эльбруса горная система приобретает типичный альпийский
облик с многочисленными ледниками и высокогорными формами рельефа
[Гвоздецкий,1954, с. 32].
Климат. Кавказ расположен на границе умеренного
и субтропического поясов. Особенности климата Западного Кавказа определяются
его географическим положением, близостью Черного и Каспийского морей,
сложностью и разнообразием рельефа. Можно выделить климатические пояса, во
многом совпадающие с природными зонами области.
От северных границ до отрогов Пастбищного хребта
распространен умеренно континентальный климат. Его формируют в основном
северные и северо-восточные воздушные массы. Здесь среднегодовая температура
составляет + 9°С, среднегодовое количество осадков - 520 мм. Для этой части
области характерно сухое, жаркое лето со средней температурой + 20°С (редко
максимальная температура достигает 40°С).
Заморозки начинаются здесь в конце сентября, а
заканчиваются в начале апреля. Среднеянварская температура: -4,2°С, минимальная
достигает: -33°С. Зима малоснежная.
Для низкогорной части области (на высоте до
800-1200 м) характерен умеренно теплый климат. В этом поясе среднегодовая
температура воздуха составляет 8-9°С, максимальная доходит до + 38°С,
минимальная до -34°С. Средняя зимняя температура воздуха -3,4°С.
В среднегорной зоне климат умеренно влажный.
Здесь, на невысоких плато, в межгорных котловинах, расположенных на высоте
1200-2000 м над уровнем моря, в формировании микроклимата велика роль
Скалистого и Бокового хребтов. Они служат надежной защитой от холодных ветров и
туманов, проникающих с северо-востока и юго-запада.
На Черноморском побережье Западного Кавказа
выделяются следующие типы климатов: на участке от Тамани до Анапы умеренный
морской, на участке от Анапы до Туапсе - средиземноморский климат, от Туапсе до
Адлера и далее за пределы России - субтропический влажный (близкий влажным
субтропикам Абхазии, Колхиды, Понта, Аджарии и Грузии). Причиной формирования
этих двух различных типов климата является рельеф, точнее - высота гор. До
Туапсе их высота не поднимается выше 1000 м, и они не являются серьезным
орографическим барьером для влагонесущих потоков воздушных масс с юго-запада,
после Туапсе высота гор достигает 3,000 и более метров, на западных наветренных
их склонах весь год выпадает большое количество осадков.
Средняя температура января: -4°С на равнине,
+5°С на Черноморском побережье, июля: 22-24°С [Тонконоженко, 1979, с.151-167].
Геологическое строение и рельеф. На всей
территории Западного Кавказа можно встретить отложения всех геологических
возрастов. Примерно 200-220 млн. лет назад Кавказской горной страны не
существовало, здесь бушевали воды океана Тетис. Современную форму рельефа она
приобрела в эпоху альпийского горообразования. Таким образом. Кавказ - молодая
горная страна. В геологическом отношении на территории области выделяются три
зоны: 1) северная: равнинно-холмистая, где в основном развиты третичные, или
совсем молодые, отложения; 2) средняя: предгорная, где залегают породы мелового
и юрского периодов; 3) южная: районы Бокового и Главного Кавказского хребтов,
где распространены в основном древние докембрийские и палеозойские породы
[Любимова, Бондаренко, 2009, с. 42-43].
Гидрографическая сеть. Горный рельеф Кавказа и
большое количество осадков в его пределах обусловили формирование густой,
причудливо разветвляющейся речной сети. Подавляющая часть рек Западного Кавказа
начинается в горах. Крупнейшие реки Западного Кавказа - Кубань (длина 900 км,
площадь бассейна 61,5 тыс. км2) и Терек (длина 591 км, площадь
бассейна 43,7 тыс. км2). Реки принадлежат к бассейнам Каспийского
(Терек, Кума, Сулак, Самур), Азовского (Кубань) и Черного (Мзымта, Сочи и др.)
морей. Подавляющая часть рек - горные. Они имеют быстрое течение, не замерзают,
имеют резкие колебания стока, подчас даже в течение суток. При выходе на
равнину характер рек сохраняется. Реки несут массу взвешенного материала с гор
и отлагают его на равнине. Терек и Кубань имеют обширные дельты. Собственно
равнинных рек, начинающихся в Предкавказье, очень мало: Калаус, Егорлык, Маныч,
Ея, Бейсуг и некоторые другие. Они имеют медленное течение, небольшое весеннее
половодье, связанное с таянием снега, и зимний ледостав. Летом многие из них
пересыхают [Мельникова, 2009, с.68 - 69].
Почвы. На Западном Кавказе преобладают степные
почвы - черноземы. А именно: черноземы мицелярно-карбонатные (приазовские,
предкавказские), черноземы выщелоченные и оподзоленные, черноземы типичные
малогумусные и сверхмощные, лугово-черноземные почвы (большей частью
солонцеватые). Морфологическими признаками типичных черноземов является большая
мощность гумусовых горизонтов: 160-170 см, темно-серая окраска, светлеющая с
глубиной и приобретающая буровые и каштановые тона, хорошо выраженная структура
- комковато-зернистая сверху и комковато-ореховая во второй половине профиля
[Соляник, 2004, с. 38]. В прибрежной полосе Черного моря, на древних морских
террасах на высоте до 450 м над уровнем моря, от Туапсе до границы с Грузией,
распространены желтоземы и подзолисто-желтоземные почвы. На них растут
широколиственные леса колхидского типа (граб, каштан) и вечнозеленые растения
(лавровишня, рододендрон, лианы, папоротники). Перегнойно-карбонатные почвы
распространены преимущественно в низкогорной части. Коричневые почвы
распространены в границах низких гор, преимущественно полосой, вытянутой вдоль
Черного моря в районе Анапа - Новороссийск. Развиваются под сухими лесами и
кустарниками на элювии известняков и мергелей. Среди них выделяются подтипы:
карбонатные, типичные выщелоченные. Основная площадь желтоземов находится в
районе Сочи - Адлер, на морских и речных террасах Черноморского побережья, где
они формируются преимущественно на делювии глинистых сланцев и песчаников.
Влажный субтропический климат создает благоприятные условия для выщелачивания
почв [Коровин, 1979, с. 151-167].
4. Популяционное исследование видов рода Hedera
L. на Западном
Кавказе
Популяционное изучение рода Hedera
L. проводилось
маршрутно-экспедиционным методом. Первичный материал был собран в ходе
однодневных поездок. Были осуществлены полевые исследования и осуществлен сбор
гербарного материала. В полевом дневнике, помимо флористических и
геоботанических сведений, фиксировались наблюдения фенологического и
экологического характера.
Сбор данных для этой работы проводился на
Западном Кавказе: Туапсинский район (дубовый лес, с. Малое Псеушхо, южный
склон; верховье реки Цыпка, дубово-буковый лес), Геленджикский район (дубовый
лес, 2 км до с. Дтвноморское, южный склон), Новороссийский район (дубовый лес,
1 км до поселка Верхне-Баканский), район МО города-курорта Анапы
(широколиственный лес, 1 км до станицы Гостагаевская, северный склон), Крымский
район (дубовый лес, река Баканка, северный склон), Апшеронский район (пойменный
лес, 3 км до села Черниговское; буковый лес, вход в Гуамское ущелье), Сочинский
район (поселок Совет Кваджи, склон у трассы) в 2010 - 2013 г (рисунок 4).
Также для более полного выявления популяционной
изменчивости вида, образцы видов Западного Кавказа были сравнены с образцами H.
helix, произрастающих в
Испании (провинция Каталония, берег Средиземного моря).
Для последующего проведения работы, в каждом
исследуемом районе были собраны листья Hedera L. в количестве 100 штук.
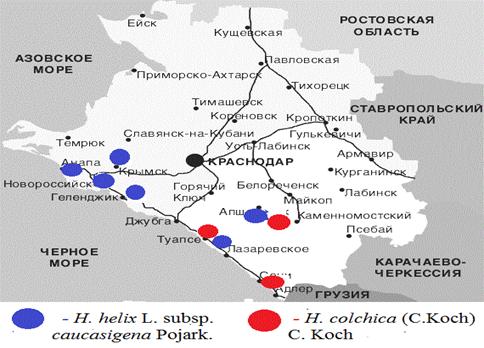
Рисунок 4 - Места сбора Hedera
L. на Западном
Кавказе
В Туапсинском районе произрастает два вида
Hedera L. В дубовом лесу (окрестностях с. Малое Псеушхо) выявлен H. helix L.
subsp. сaucasigena (рисунок 4). Этот вид произрастает на бурых лесных почвах,
при средних температурах июля - +26 и средних январских - +4°С, годовое
количество осадков 800 - 1600 мм [Атлас Краснодарского края, 1996, с. 13].
В дубово-буковом лесу, в верховье реки Цыпка был
отмечен H. colchica на бурых лесных почвах, в тех же климатических условиях,
что и H. helix L. subsp. сaucasigena.
В Геленджикском районе произрастает H. helix L.
subsp. сaucasigena (рисунок 4). Вид был собран на южном склоне (обезлесенные
пологие склоны и шлейфы с редкими группами деревьев и кустарников, в сообществе
с дубом скальным, дубом пушистым и буком), на дерново-карбонатных и бурых
лесных почвах, в 2 км до с. Дивноморское. Район относится к зоне сухих
субтропиков средиземноморского типа. Среднегодовая температура воздуха
составляет 13°С. Среднегодовое количество осадков 700-800 мм в год [Атлас
Краснодарского края, 1996, с. 13]. В районе МО города-курорта Анапы растет H.
helix L. subsp. сaucasigena (рисунок 4). Этот вид был собран в широколиственном
лесу, с преобладанием дуба пушистого (напочвенное покрытие). Произрастает на
западном склоне на дерново-карбонатных почвах. Среднегодовое количество осадков
составляет менее 500 мм в год [Атлас Краснодарского края, 1996, с. 13].
Новороссийский район. В этом районе H. helix L.
subsp. сaucasigena был собран в двух местах (рисунок 4). Первое местообитание в
3 км до г. Новороссийска на южном склоне в дубом лесу с преобладанием дуба
скального и пушистого, растет на коричневых почвах. Также вид был собран на
южном склоне, в дубовом лесу в 1 км от п. Верхне-Баканского в сторону г.
Новороссийска, только уже на дерново-карбонатных почвах. Среднегодовое
количество осадков составляет 500 - 600 мм в год [Атлас Краснодарского края,
1996, с. 13].
На территории Крымского района произрастает H.
helix L. subsp. сaucasigena (рисунок 4). Вид был собран в широколиственном лесу
(преобладанием дуба) вблизи р. Баканка на дерново-карбонатных почвах, северный
склон, при умеренно-континентальном климате. В июле средняя температура воздуха
составляет +23ºС, в январе -4ºС.
Среднегодовое количество осадков от 600 до 700 мм в год [Атлас Краснодарского
края, 1996, с. 22].
Апшеронский район. H.
helix L.
subsp. сaucasigenа
был собран на пойменном лугу, в 3 км до с. Черниговское (рисунок 4).
Местообитание второго экземпляра (H.
colchica) - буковый лес,
вход в Гуамское ущелье, является напочвенным покрытием образов. Растет на
темно-серых и серых лесных почвах. Среднемесячная температура января 0--4ºС,
июля +23ºС.
Среднегодовое количество осадков - 800-1600 мм в год [Атлас Краснодарского
края, 1996, с. 13].
Испания. Провинция Каталония. На большей части
территории Каталонии преобладает средиземноморский климат с жарким и сухим
летом и мягкой и относительно дождливой зимой. Среднегодовая температура около
15 градусов. Общегодовое количество осадков 400-600 мм. Сбор H.
helix проводился в трех
местах: гора Монсеррат (Средиземноморье, 700 м над уровнем моря, на щебляке),
гора Тосса-де-Мар (берег моря), гора Льорет-де-Мар (берег Средиземного моря).
.1 Биометрические показатели морфологических
признаков листа Hedera
L
Материал для исследования получен в результате
измерения листа Hedera
L. Отдельная
листовая пластинка была измерена по трем показателям: длина черешка (L);
количество лопастей (n)
и длина каждой лопасти (l).
Для листовых пластинок, имеющих только одну лопасть, измерялись максимальная
ширина (a) и длина (b).
Измерения проводились с помощью линейки (рисунок 6).


Рисунок 5 - Схема измерения листовой пластинки Hedera
L.
По схеме, представленной на рисунке 5, было
измерено 1200 листовых пластинок Hedera L., морфологические признаки которых
приведены в таблицах 2-11 (Приложение Листовые пластинки Hedera L., над
которыми проводились исследования и результаты статистической обработки).
Все необходимые вычисления выполнены на ПК с
использованием компьютерной программы Excel.
Таблица 2 - Биометрические показатели
морфологических признаков листа H.
helix L.
subsp. caucasigena
(Туапсинский район, дубовый лес, с. Малая Псеушхо южный склон)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
36,21
± 1,9
|
8
|
80
|
26
|
17,49
|
48,5
|
|
Количество
лопастей, шт
|
5
± 0,2
|
3
|
7
|
3
|
1,58
|
31,6
|
|
Глубина
1-ой лопасти, мм
|
8,48
± 0,62
|
2
|
21
|
3
|
5,74
|
56,2
|
|
Глубина
2-ой лопасти, мм
|
21,69
± 1,13
|
2
|
52
|
14
|
10,4
|
48,1
|
|
Глубина
3-ей лопасти, мм
|
17,59
± 1,25
|
2
|
51
|
5
|
11,54
|
65,6
|
|
Глубина
4-ой лопасти, мм
|
18,25
± 1,0
|
5
|
40
|
15
|
6,9
|
37,8
|
|
Глубина
5-ой лопасти, мм
|
10,0
± 1,0
|
1
|
25
|
5
|
6,9
|
60
|
|
Глубина
6-ой лопасти, мм
|
14,7
± 1,5
|
2
|
27
|
16
|
6,7
|
45,5
|
|
Глубина
7-ой лопасти, мм
|
5,3
± 0,8
|
2
|
14
|
2
|
3,5
|
66
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Как видно из таблицы, наиболее изменчивым
признаком для этой популяции является глубина залегания 7-ой лопасти (V=
66%),а наиболее постоянными признаками H.
helix L.
subsp. caucasigena
является количество лопастей (V=31,6%).
Изменчивость большей части признаков растений имеет средний и высокий уровень
варьирования.Длина черешка изменятся от 8 мм до 80 мм. Наиболее часто
встречаемая длина - 26 мм. Встречаются трех-пяти-семилопастные листья,
Преобладают трехлопастные листья.
Таблица 3 - Биометрические показатели
морфологических признаков листа H.
helix L.
subsp. caucasigena
(Геленджикский район, южный склон, 2 км до с. Дивноморское)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
откл.
|
V,%
|
|
Длина
черешка, мм
|
43,94
± 1,6
|
10
|
84
|
45
|
16,03
|
36,4
|
|
Количество
лопастей, шт
|
4,04
± 0,1
|
3
|
7
|
3
|
1,34
|
33,1
|
|
Глубина
1-ой лопасти, мм
|
7,39
± 0,4
|
1
|
25
|
5
|
4,94
|
66,8
|
|
Глубина
2-ой лопасти, мм
|
19,8
± 1,0
|
2
|
45
|
5
|
10,63
|
53,6
|
|
Глубина
3-ей лопасти, мм
|
16,28
± 1,1
|
3
|
60
|
10
|
11,2
|
68,7
|
|
Глубина
4-ой лопасти, мм
|
13,9
± 1,0
|
3
|
26
|
20
|
6,3
|
45,3
|
|
Глубина
5-ой лопасти, мм
|
4,4
± 0,5
|
1
|
15
|
3
|
3,5
|
79,5
|
|
Глубина
6-ой лопасти, мм
|
7,4
± 1,1
|
2
|
14
|
7
|
3,6
|
48,6
|
|
Глубина
7-ой лопасти, мм
|
2,7
± 0,5
|
1
|
7
|
1
|
2,05
|
75,9
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Изменчивость большей части признаков растений из
этой популяции средняя и высокая. Наиболее изменчивым признаком для этой
популяции является глубина залегания 5-ой лопасти (V=
79,5%), 7-ой лопасти (V=75,9
%),а наиболее постоянными признаками H.
helix L.
subsp. caucasigena
является количество лопастей(V=33,1%)
и длина черешка (V=36,4%). Длина черешка изменятся от 10 до 84 мм, наиболее часто
встречаемая длина - 45 мм. Количество лопастей варьирует от 3 до 7 штук,
преобладают трехлопастные листья.
Таблица 4 - Биометрические показатели
морфологических признаков листа H.
helix L.
subsp. caucasigena
(Новороссийский район, дубовый лес, 1 км до поселка Верхне-Баканский, северный
склон)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
откл.
|
V,%
|
|
Длина
черешка, мм
|
63,8
± 2,0
|
15
|
125
|
70
|
27,6
|
43,2
|
|
Количество
лопастей, шт
|
4
± 0,1
|
3
|
5
|
5
|
1,0
|
25
|
|
Глубина
1-ой лопасти, мм
|
10,4
± 0,6
|
1
|
35
|
9
|
6,6
|
63,4
|
|
Глубина
2-ой лопасти, мм
|
27,2
± 1,1
|
6
|
60
|
21
|
11,1
|
40,8
|
|
Глубина
3-ей лопасти, мм
|
29,5
± 1,8
|
5
|
75
|
10
|
18,2
|
61,6
|
|
Глубина
4-ой лопасти, мм
|
18,9
± 1,0
|
5
|
36
|
21
|
7,5
|
39,6
|
|
Глубина
5-ой лопасти, мм
|
5,8
± 0,5
|
1
|
20
|
3
|
3,9
|
67,2
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Наиболее изменчивым признаком для этой популяции
является глубина залегания 1-ой (V=
63,4 %) и 5-ой лопасти (V=
67,2 %). Наиболее постоянным признаком - количество лопастей (V=
25 %). Длина черешка варьирует от 15 мм до 125. Часто встречаемая длина - 70
мм. Количество лопастей изменяется от 3 до 5 штук. Большинство листьев
пятилопастные.
Таблица 5 - Биометрические показатели
морфологических признаков листа H.
helix L.
subsp. caucasigena
(Анапский район, широколиственный лес, 1 км до ст. Гостагаевская)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
86,5
± 2,2
|
25
|
147
|
27,4
|
31,6
|
|
Количество
лопастей, шт
|
4,16
± 0,1
|
3
|
5
|
3
|
1,0
|
24
|
|
Глубина
1-ой лопасти, мм
|
13,54
± 1,0
|
1
|
43
|
5
|
10,0
|
71,8
|
|
Продолжение
таблицы 5
|
|
Глубина
2-ой лопасти, мм
|
35,71
± 1,8
|
2
|
90
|
20
|
19,5
|
54,6
|
|
Глубина
3-ей лопасти, мм
|
6,46
± 0,6
|
1
|
24
|
5
|
4,9
|
75,1
|
|
Глубина
4-ой лопасти, мм
|
24,1
± 1,2
|
8
|
57
|
31
|
9,8
|
40,6
|
|
Глубина
5-ой лопасти, мм
|
6,4
± 0,6
|
1
|
24
|
5
|
4,8
|
75
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Как видно из таблицы, наиболее изменчивым
признаком для этой популяции является глубина залегания 7-ой лопасти (V=
75%),а наиболее постоянными признаками H.
helix L.
subsp. caucasigena
является количество лопастей (V=24%)
и длина черешка(V=31,6%).
Изменчивость большей части признаков растений имеет средний и высокий уровень
варьирования. Длина черешка изменяется от 25 мм до 147 мм. Часто встречаемая
длина - 86 мм. Количество лопастей варьирует от 3 до 5 штук. Преобладают
трехлопастные листья.
Таблица 6 - Биометрические показатели
морфологических признаков листа H.
helix L.
subsp. caucasigena
(Крымский район, дубовый лес, река Баканка, северный склон)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
82,7
± 2,1
|
10
|
142
|
80
|
23,1
|
28
|
|
Количество
лопастей, шт
|
4
± 0,1
|
3
|
7
|
5
|
1,04
|
26
|
|
Глубина
1-ой лопасти, мм
|
8,6
± 0,7
|
1
|
28
|
3
|
7,2
|
83
|
|
Глубина
2-ой лопасти, мм
|
26,9
± 1,7
|
3
|
90
|
10
|
17,4
|
64
|
|
Глубина
3-ей лопасти, мм
|
25,2
± 1,6
|
2
|
67
|
9
|
16,8
|
66
|
|
Глубина
4-ой лопасти, мм
|
13,9
± 0,9
|
2
|
32
|
14
|
6,7
|
48
|
|
Глубина
5-ой лопасти, мм
|
3,3
± 0,4
|
1
|
19
|
2
|
3,04
|
92
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Как видно из таблицы, наиболее изменчивым
признаком для этой популяции является глубина залегания 5-ой лопасти (V=
92%) и 1-ой лопасти (V=
83%) ,а наиболее постоянными признаками H.
helix L.
subsp. caucasigena
является количество лопастей (V=26%)
и длина черешка (V=28%).
Изменчивость большей части признаков растений имеет средний и высокий уровень
варьирования. Длина черешка изменятся от 10 мм до 142 мм. Часто встречаемая
длина - 80 мм. Встречаются трех-пяти-семилопастные листья. Преобладают
пятилопастные.
Таблица 7 - Биометрические показатели
морфологических признаков листа H.
helix L.
subsp. caucasigena
(Апшеронский район, пойменный лес, 3 км до с. Черниговское)
|
Признаки
|
χ¯±
Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
36,73
± 1,5
|
13
|
100
|
33
|
15,5
|
42,1
|
|
Количество
лопастей, шт
|
4,2
± 0,1
|
3
|
5
|
3
|
1,0
|
23
|
|
Глубина
1-ой лопасти, мм
|
6,63
± 0,5
|
2
|
22
|
3
|
4,2
|
63
|
|
Глубина
2-ой лопасти, мм
|
18,02
± 1,1
|
5
|
59
|
11
|
11,4
|
63
|
|
Глубина
3-ей лопасти, мм
|
23,59
± 1,3
|
3
|
57
|
10
|
13,07
|
55
|
|
Глубина
4-ой лопасти, мм
|
10,6
± 0,7
|
4
|
41
|
10
|
6,2
|
58
|
|
Глубина
5-ой лопасти, мм
|
4,8
± 0,3
|
2
|
18
|
5
|
2,7
|
56
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Изменчивость большей части признаков растений из
этой популяции средняя и высокая. Наиболее изменчивым признаком для этой
популяции является глубина залегания 1-ой лопасти и 2-ой лопасти (V=
63%), а наиболее постоянными признаками H.
helix L.
subsp. caucasigena
является количество лопастей (V=23%).
Длина черешка изменятся от 13 мм до 100 мм. Часто встречаемая длина - 33 мм.
Преобладают трехлопастные листья.
Таблица 8 - Биометрические показатели
морфологических признаков листа H.
helix (Испания,
провинция Каталония, берег Средиземного моря)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
120,7
± 0,2
|
46
|
190
|
140
|
34,3
|
28
|
|
Количество
лопастей, шт
|
3,9
± 0,1
|
3
|
5
|
5
|
1,0
|
25
|
|
Глубина
1-ой лопасти, мм
|
14,57
± 0,8
|
2
|
39
|
15
|
8,6
|
59
|
|
Глубина
2-ой лопасти, мм
|
52,87
± 1,2
|
32
|
100
|
50
|
12,9
|
24
|
|
Глубина
3-ей лопасти, мм
|
14,81
± 0,9
|
2
|
45
|
10
|
9
|
60
|
|
Глубина
4-ой лопасти, мм
|
28,08
± 1,2
|
11
|
47
|
20
|
9,0
|
32
|
|
Глубина
5-ой лопасти, мм
|
14,81
± 1,0
|
2
|
45
|
10
|
9,0
|
60
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Наиболее изменчивым признаком для этой популяции
является глубина залегания 5-ой лопасти (V=
60%), а наиболее постоянными признаками H.
Helix L.
является количество лопастей (V=25%)
и длина черешка (V=28%). Длина
черешка изменятся от 46 мм до 190 мм. Часто встречаемая длина - 140 мм.
Количество лопастей - от 3 до 5 штук. Преобладают пятилопастные листья.
Таблица 9 - Биометрические показатели
морфологических признаков листа H.
colchica (Апшеронский
район, буковый лес, вход в Гуамское ущелье)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
61
± 1,0
|
20
|
101
|
70
|
17,2
|
28,1
|
|
Количество
лопастей, шт
|
1
± 0
|
1
|
1
|
1
|
0
|
0
|
|
Ширина
(а), мм
|
69
± 1,0
|
26
|
134
|
67
|
13,1
|
18,9
|
|
Продолжение
таблицы 9
|
|
Длина
(b), мм
|
91,3
± 1,1
|
56
|
160
|
110
|
18,5
|
20,2
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Наиболее изменчивым признаком для этой популяции
является длина черешка (V=
28,1%), а постоянными признаками H.
colchica является
количество лопастей (V=0%).
Длина черешка изменятся от 20 мм до 101 мм. Часто встречаемая длина черешка
составляет 70 мм. Количество лопастей не изменяется.
Таблица 10 - Биометрические показатели
морфологических признаков листа H.
colchica (Туапсинсий район,
верховье р. Цыпка, дубово-буковый лес)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
67
± 1,3
|
20
|
130
|
45
|
25,03
|
37,3
|
|
Количество
лопастей, шт
|
1
± 0
|
1
|
1
|
1
|
0
|
0
|
|
Ширина
(а), мм
|
69,5
± 1,9
|
37
|
132
|
71
|
19,5
|
28
|
|
Длина
(b), мм
|
95
± 1,7
|
58
|
168
|
110
|
17,5
|
18
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
Наиболее изменчивым признаком для этой популяции
является длина черешка (V=
37,3%), а постоянным признаком H.
colchica является
количество лопастей (V=0%).
Длина черешка меняется от 20 мм до 130 мм. Преобладают листья, у которых длина
черешка составляет 45 мм. Количество лопастей не изменяется.
Таблица 11 - Биометрические показатели
морфологических признаков листа H.
colchica (Сочинский район,
п. Совет Кваджи, склон у трассы)
|
Признаки
|
χ¯ ± Sχ¯
|
min
|
max
|
Mo
|
Стандартное
отклонение
|
V,%
|
|
Длина
черешка, мм
|
62
± 1,3
|
34
|
105
|
67
|
21,8
|
35
|
|
Количество
лопастей, шт
|
1
± 0
|
1
|
1
|
1
|
0
|
0
|
|
Ширина
(а), мм
|
67
± 1,3
|
21
|
94
|
61
|
17,4
|
26
|
|
Длина
(b), мм
|
90,7
± 1,0
|
22
|
153
|
101
|
17,5
|
19
|
Примечание: χ¯ - средняя
арифметическая; Sχ¯
-
ошибка репрезентативности выборочной средней; min-max
- минимальное и максимальное значения параметра; Mo
- мода; V - коэффициент
вариации.
В популяции данного вида наиболее изменчивым
является длина черешка (V=
35 %), а постоянным признаком - количество лопастей. Длина черешка изменятся от
34 мм до 105 мм. Часто встречаемая длина - 67 мм. Количество лопастей не
изменяется.
Таким образом, у H.
helix L.
subsp. caucasigena
наибольшей биометрической изменчивости подвергается глубина залегания лопастей,
а наиболее постоянными признаки являются длина черешка и количество лопастей.
Для H.
colchica характерно наличие
одной лопасти, а значит этот признак является постоянным. Размеры ширины и
длины в значительной степени варьируют.
.2 Влияние абиотических факторов на
популяционную изменчивость листьев видов рода Hedera
L
Одна из задач работы заключается в изучении
влияния внешних факторов на популяционную изменчивость листа видов рода Hedera
L. В работе мы
рассматриваем прежде всего морфологический и экологический критерии вида.
Морфологический критерий вида основан на существовании морфологических
признаков, характерных для одного вида, но отсутствующих у других видов.
Экологический критерий основан на том, что два вида не могут занимать одну
экологическую нишу. Следовательно, каждый вид характеризуется своими
собственными отношениями со средой обитания. Морфологические признаки листа
зависят как от вида, так и от влияния абиотических факторов, например:
температура, влажность, экспозиция склона и т.д. Изменение какого-либо внешнего
фактора может приводить к колебаниям биометрических показателей. К факторам
относятся вид и местообитание растения, а признаки - длина черешка, количество и
глубина лопасти. Для того чтобы изучит влияние факторов на изменчивость
признака, необходимо произвести расчет наименьшей существенной разности. НСР0,5
рассчитывалась по формулам:
 (1)
(1)

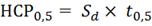 (2)
(2)
где: Sd - ошибка
разности средней ошибки; Sх- средняя ошибка среднего; t0,5
- стандартный t- критерий.
флористический биометрический лист кавказ
4.2.1 Влияние абиотических факторов на
морфологическую изменчивость листа H. helix L. subsp. сaucasigenа
В результате проведенного анализа было
установлено, что местообитание H. helix L. subsp. сaucasigenа по первому
признаку (длина черешка) доказывает как достоверные различия, так и недостоверные
(таблица 12).
Таблица 12 - Оценка разности средних значений
длины черешка
Длина
черешка, мм
|
|
|
S
|
|
Туапсинский
район
|
36
|
1,9
|
|
Геленджикский
район
|
44
|
1,6
|
|
МО
город-курорт Анапа
|
87
|
2,7
|
|
Крымский
район
|
83
|
2,3
|
|
Апшеронский
район
|
37
|
1,6
|
|
Примечание:
НСР05 = 6,2
|
Согласно полученным данным (таблица 12),
наименьшая существенная разность по длине черешка относительно местообитания
вида, следующая:
Туапсинский - МО город-курорт Анапа - разница
достоверна (Δ > НСР05);
Туапсинский - Новороссийский - разница достоверна (Δ
>
НСР05);
Туапсинский - Крымский - разница достоверна (Δ
>
НСР05);
Туапсинский - Апшеронский - разница недостоверна
(Δ
<
НСР05);
Геленджикский - МО город-курорт Анапа - разница
достоверна (Δ > НСР05);
Геленджикский - Новороссийский - разница
достоверна (Δ > НСР05);
Геленджикский - Крымский - разница достоверна (Δ
>
НСР05);
Геленджикский - Апшеронский - разница достоверна
(Δ
>
НСР05);
МО город-курорт Анапа - Новороссийский - разница
достоверна (Δ > НСР05);
МО город-курорт Анапа - Крымский - разница
недостоверна (Δ < НСР05);
МО город-курорт Анапа - Апшеронский - разница
достоверна (Δ > НСР05);
Новороссийский - Крымский - разница достоверна (Δ
>
НСР05);
Новороссийский - Апшеронский - разница
достоверна (Δ > НСР05);
Крымский - Апшеронский - разница достоверна (Δ
>
НСР05).
Следовательно, если Δ
<
НСР05, то отличие недостоверно и местообитание не влияет на длину
черешка. Например: МО город-курорт Анапа - Крымский - виды произрастают в
схожих условиях среды (северная экспозиция склонов, одинаковый температурный
режим и среднегодовое количество осадков). В этих районах наблюдаются
наибольшие длины черешков.
Если Δ >
НСР05,то отличие достоверно и местообитание влияет на длину черешка. Например:
Туапсинский - МО город-курорт Анапа - виды произрастают в условиях с различным
температурным режимом, среднегодовым количеством осадков. Значительную роль
играет нахождение на разных склонах (Туапсинский - южный склон, МО город-курорт
Анапа - западный). В МО город-курорт Анапа районе длина черешка значительно превышает
длину черешка в Туапсинском районе (рисунок 6).

Рисунок 6 - Средняя длина черешка по районам
(мм)
Таким образом, наибольшие длины черешка
наблюдаются в Крымском, Новороссийском районах и МО города-курорта Анапа.
Наименьшие средние значения длин в Туапсинском, Апшеронском и Геленджикском
районах.
Затем был произведен анализ влияние факторов на
изменчивость количество лопастей (таблица 13).
Таблица 13 - Оценка разности средних значений
количества лопастей
|
Местообитание
вида
|
Количество
лопастей, шт
|
|
|
S
|
|
Туапсинский
район
|
5
|
0,2
|
|
Геленджикский
район
|
4
|
1,3
|
|
МО
город-курорт Анапа
|
4
|
0,1
|
|
Новороссийский
район
|
4
|
0,1
|
|
Крымский
район
|
4
|
0,1
|
|
Апшеронский
район
|
4
|
0,1
|
|
Примечание:
НСР05 = 2,8
|
Согласно полученным данным (таблица 13),
разность количества лопастей по отношению к местообитанию, следующая:
а) Туапсинский-Геленджикский - разница
недостоверна (Δ < НСР05);
б) Туапсинский - МО город-курорт Анапа - разница
недостоверна (Δ < НСР05);
в) Туапсинский - Новороссийский - разница
недостоверна (Δ < НСР05);
г) Туапсинский - Крымский - разница недостоверна
(Δ
<
НСР05);
д) Туапсинский - Апшеронский - разница
недостоверна (Δ < НСР05);
е) Геленджикский - МО город-курорт Анапа -
разница недостоверна (Δ <
НСР05);
ж) Геленджикский - Новороссийский - разница
недостоверна (Δ < НСР05);
з) Геленджикский - Крымский - разница
недостоверна (Δ < НСР05);
и) Геленджикский - Апшеронский - разница
недостоверна (Δ < НСР05);
к) МО город-курорт Анапа - Новороссийский -
разница недостоверна (Δ <
НСР05);
л) МО город-курорт Анапа - Крымский - разница
недостоверна (Δ < НСР05);
м) МО город-курорт Анапа- Апшеронский - разница
недостоверна (Δ < НСР05);
н) Новороссийский - Крымский - разница
недостоверна (Δ < НСР05);
о) Новороссийский - Апшеронский - разница
недостоверна (Δ < НСР05);
п) Крымский - Апшеронский - разница недостоверна
(Δ
<
НСР05).
Во всех перечисленных выше районах Δ
<
НСР05, следовательно, различие недостоверно и факторы (экспозиция
склона, свет, влажность и т.д.) не влияют на количество лопастей листа.
Наиболее часто встречаются трехлопастные листья (МО город-курорт Анапа,
Новороссийский, Крымский и Апшеронский районы), реже - семилопастные (рисунок
7).

Рисунок 7 - Среднее количество
лопастей по районам (шт)
Далее идет анализ влияния факторов
на глубину каждой лопасти. Каждая лопасть рассматривается как отдельный
признак.
Согласно полученным данным (таблица
14), наименьшая существенная разность по глубине первой лопасти относительно
местообитания вида, следующая:
а) Туапсинский - Геленджикский -
разница недостоверна (Δ < НСР05);
б) Туапсинский - Анапский - разница
достоверна (Δ
>
НСР05);
в) Туапсинский - Новороссийский -
разница недостоверна (Δ < НСР05);
г) Туапсинский - Крымский - разница
недостоверна (Δ
<
НСР05);
д) Туапсинский - Апшеронский -
разница недостоверна (Δ
<
НСР05);
е) Геленджикский - Анапский -
разница достоверна (Δ
>
НСР05);
ж) Геленджикский - Новороссийский -
разница недостоверна (Δ
<
НСР05);
з) Геленджикский - Крымский -разница
недостоверна (Δ
<
НСР05);
и) Геленджикский - Апшеронский -
разница недостоверна (Δ
<
НСР05);
к) Анапский - Новороссийский -
разница недостоверна (Δ
<
НСР05);
л) Анапский - Крымский - разница
недостоверна (Δ
<
НСР05);
м) Анапский - Апшеронский - разница
достоверна (Δ
>
НСР05);
н) Новороссийский - Крымский -
разница недостоверна (Δ
<
НСР05);
о) Новороссийский - Апшеронский -
разница недостоверна (Δ
<
НСР05);
п) Крымский - Апшеронский - разница
недостоверна (Δ
<
НСР05).
Таблица 14 - Оценка разности средних
значений 1-ой лопасти (3- лопастной)
|
Местообитание
вида
|
Глубина
1-ой лопасти
|
|
|
S
|
|
Туапсинский
район
|
10,2
|
0,8
|
|
Геленджикский
район
|
10,7
|
0,8
|
|
Анапский
район
|
23,3
|
1,2
|
|
Новороссийский
район
|
15,0
|
1,2
|
|
Крымский
район
|
14,2
|
1,2
|
|
Апшеронский
район
|
10,1
|
0,7
|
|
Примечание:
НСР05 = 2,8
|
Рассматривая первую лопасть как отдельный
признак, можно заметить следующее: климатические экофакторы не влияют на
глубину лопасти. Исключения составляют районы Геленджикский - Анапский и
Анапский - Апшеронский (рисунок 8).
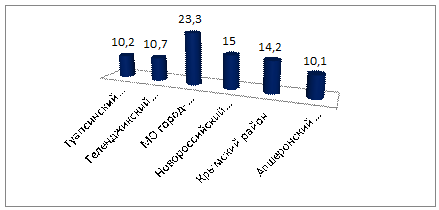
Рисунок 8 - Средняя глубина первой
лопасти по районам (мм)
Согласно полученным данным по второй
лопасти (таблица 15), наименьшая существенная разность по районам следующая:
а) Туапсинский-Геленджикский -
разница недостоверна (Δ
<
НСР05);
б) Туапсинский - Анапский - разница
достоверна (Δ
>
НСР05);
в) Туапсинский - Новороссийский -
разница достоверна (Δ > НСР05);
г) Туапсинский - Крымский - разница
достоверна (Δ
>
НСР05);
д) Туапсинский - Апшеронский - разница
недостоверна (Δ
<
НСР05);
е) Геленджикский - Анапский -
разница достоверна (Δ
>
НСР05);
ж) Геленджикский - Новороссийский -
разница достоверна (Δ
>
НСР05);
з) Геленджикский - Крымский -
разница достоверна (Δ
>
НСР05);
и) Геленджикский - Апшеронский -
разница недостоверна (Δ
<
НСР05);
к) Анапский - Новороссийский -
разница достоверна (Δ
>
НСР05);
л) Анапский - Крымский - разница
недостоверна (Δ
<
НСР05);
м) Анапский - Апшеронский - разница
достоверна (Δ > НСР05);
н) Новороссийский - Крымский -
разница недостоверна (Δ
<
НСР05);
о) Новороссийский - Апшеронский -
разница недостоверна (Δ > НСР05);
п) Крымский - Апшеронский - разница
достоверна (Δ > НСР05).
Таблица 15 - Оценка разности средних
значений 2-ой лопасти (3-лопастной)
|
Местообитание
вида
|
Глубина
2-ой лопасти
|
|
|
S
|
|
Туапсинский
район
|
30,0
|
1,5
|
|
Геленджикский
район
|
27,6
|
1,4
|
|
Анапский
район
|
54,3
|
2,5
|
|
Новороссийский
район
|
35,6
|
1,5
|
|
Крымский
район
|
41,1
|
2,5
|
|
Апшеронский
район
|
30,0
|
1,4
|
|
Примечание:
НСР05 = 5
|
Внешние факторы только в некоторых районах
влияют на глубину лопасти. Наибольшая глубина лопасти в МО город-курорт Анапа
районе (54,3 мм) и Крымском (41,4 мм). Следовательно, достоверно влияние
факторов в районах Анапский - Апшеронский, МО город-курорт Анапа -
Новороссийский, Геленджикский - Крымский, Геленджикский - МО город-курорт
Анапа, Туапсинский - Крымский, Туапсинский - МО город-курорт Анапа (рисунок 9).

Рисунок 9 - Средняя глубина второй
лопасти по районам (мм)
Согласно полученным данным по
третьей лопасти (таблица 16), наименьшая существенная разность по районам
следующая:
а) Туапсинский-Геленджикский - разница
недостоверна (Δ < НСР05);
б) Туапсинский - Анапский - разница достоверна (Δ
>
НСР05);
в) Туапсинский - Новороссийский - разница недостоверна
(Δ
< НСР05);
г) Туапсинский - Крымский - разница недостоверна
(Δ
<
НСР05);
д) Туапсинский - Апшеронский - разница
недостоверна (Δ < НСР05);
е) Геленджикский - Анапский - разница достоверна
(Δ
>
НСР05);
ж) Геленджикский - Новороссийский - разница
недостоверна (Δ < НСР05);
з) Геленджикский - Крымский - разница
недостоверна (Δ < НСР05);
и) Геленджикский - Апшеронский - разница
недостоверна (Δ < НСР05);
к) Анапский - Новороссийский - разница
достоверна (Δ > НСР05);
л) Анапский - Крымский - разница достоверна (Δ
>
НСР05);
м) Анапский - Апшеронский - разница достоверна (Δ
> НСР05);
н) Новороссийский - Крымский - разница
недостоверна (Δ < НСР05);
о) Новороссийский - Апшеронский - разница
достоверна (Δ > НСР05);
п) Крымский - Апшеронский - разница недостоверна
(Δ
< НСР05).
Таблица 16 - Оценка разности средних значений
3-ей лопасти (3-лопастной)
|
Местообитание
вида
|
Глубина
3-ей лопасти
|
|
|
S
|
|
Туапсинский
район
|
11
|
1,0
|
|
Геленджикский
район
|
11
|
1,0
|
|
Анапский
район
|
21,9
|
1,2
|
|
Новороссийский
район
|
14,7
|
1,1
|
|
Крымский
район
|
12,6
|
1,6
|
|
Апшеронский
район
|
9,6
|
0,7
|
|
Примечание:
НСР05 = 3,2
|
Если Δ < НСР05,
то отличие недостоверно и местообитание не влияет на глубину третьей
лопсати (Туапсинский-Геленджикский, Туапсинский - Новороссийский, Туапсинский -
Крымский, Туапсинский - Апшеронский, Геленджикский - Новороссийский,
Геленджикский - Крымский, Новороссийский - Крымский, Крымский - Апшеронский).
А в таких районах как Туапсинский - МО
город-курорт Анапа, Геленджикский - МО город-курорт Анапа, МО город-курорт
Анапа - Новороссийский, МО город-курорт Анапа - Крымский, МО город-курорт Анапа
- Апшеронский, Новороссийский - Апшеронский Δ
> НСР05, следовательно, экологические факторы будут влиять на
глубину лопасти (рисунок 10).

Рисунок 10 - Средняя глубина третьей лопасти по
районам (мм)
Таким образом, мы видим, что один и тот же вид Hedera
L.в зависимости от
условий местообитания популяции (абиотических факторов) изменчив по
морфологическим признакам.
.2.2 Сравнительный анализ морфологии листа
подвида H. helix
L. subsp.
caucasigena
c средиземноморским
видом H. helix
L
Для более полного изучения популяционной
изменчивости вида Hedera
helix (также на примере
листа), проводилось сравнение популяций подвидов Западного Кавказа (H.helix L.
subsp. сaucasigenа) и популяций видов средиземноморских местообитаний (Испания,
провинция Каталония) (таблица 17).
Получив значение НСР, можно сделать вывод, что
различия существенны Δ > НСР0,5,
следовательно разница достоверна и местообитание влияет на длину черешка.
На рисунке 11 показаны средние значения длин
черешков растения, произрастающего на Западном Кавказе (H. helix L. subsp.
сaucasigenа) и Средиземноморье (Испания) (H. helix L.).
Из рисунка видно, что длина черешка видов,
произрастающих в Средиземноморье на много отличается от Западно-Кавказских
видов и составляет 120 мм, тогда как виды западного Кавказа имеют среднюю длину
58 мм.
Таблица 17 - Средние значения длины черешка
популяций на Западном Кавказе (H. helix L. subsp. сaucasigenа) и
Средиземноморье (Испания) (H. helix L.)
|
Местообитание
вида
|
Длина
черешка (мм)
|
|
χ¯±
Sχ¯
|
|
Западный
Кавказ
|
58
± 2,1
|
|
Средиземноморье
(Испания)
|
120
± 3,4
|
|
Примечание:
НСР0,5= 7,6
|
На Западном Кавказе преобладают трехлопастные
листья (65%) (рисунок 12), у вида, произрастающего в Испании - пятилопастные
листья (68%) (рисунок 13). Сравнительный анализ глубины лопасти проводился на
трехлопастных листьях. Каждая лопасть рассматривается как отдельный признак
(таблица 18, 19, 20). Таким образом, очевидно, что вид H.
helix, произрастающий в
Средиземноморье имеет более крупную листовую пластинку по сравнению с
Западно-кавказским подвидом H.
helix L.
subsp. сaucasigena
(рисунок 14).
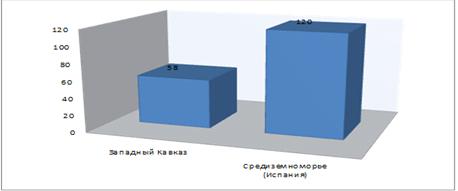
Рисунок 11 - Средняя длина черешка (мм)
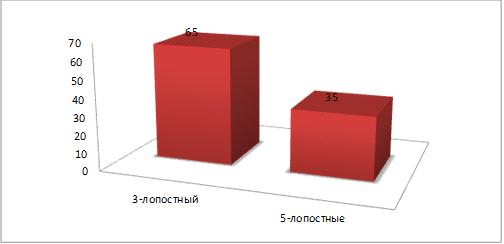
Рисунок 12 - Количество лопастей вида Западного
Кавказа (%)

Рисунок 13 - Количество лопастей вида
Средиземноморья (%)
Таблица 18 - Оценка разности средних значений
1-ой лопасти (3-лопастной)
|
Местообитание
вида
|
Глубина
1-ой лопасти
|
|
χ¯±
Sχ¯
|
|
Западный
Кавказ
|
13,9
± 1,0
|
|
Средиземноморье
(Испания)
|
14,5
± 1,0
|
|
Примечание:
НСР0,5= 2,8
|
Таблица 19 - Оценка разности средних значений
2-ой лопасти (3-лопастной)
|
Местообитание
вида
|
Глубина
2-ой лопасти
|
|
χ¯±
Sχ¯
|
|
Западный
Кавказ
|
36,4
± 1,8
|
|
Средиземноморье
(Испания)
|
52,87
± 1,3
|
|
Примечание:
НСР0,5= 4,5
|
Таблица 20 - Оценка разности средних значений
3-ей лопасти (3- лопастной)
|
Местообитание
вида
|
Глубина
3-ей лопасти
|
|
χ¯±
Sχ¯
|
|
Западный
Кавказ
|
13,4
± 1,0
|
|
Средиземноморье
(Испания)
|
14,8
± 0,9
|
|
Примечание:
НСР0,5= 2,6
|
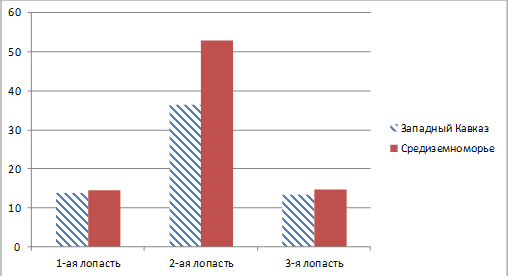
Рисунок 14 - Средние значения глубины лопасти
(мм)
.2.2 Влияние факторов на морфологическую
изменчивость листа H. colchica
(C.Koch)
C.Koch
В результате проведенного анализа было
установлено, что местообитание H.
colchica по первому
признаку (длина черешка) доказывает как достоверные различия, так и
недостоверные (таблица 21).
Таблица 21 - Оценка разности средних значений
длины черешка
|
Местообитание
вида
|
Длина
черешка
|
|
|
S
|
|
Апшеронский
район
|
61
|
1,9
|
|
Туапсинский
район
|
67
|
2,8
|
|
Сочинский
район
|
62
|
3,7
|
|
Примечание:
НСР0,5= 6,8
|
В результате проведенного анализа оценки разности
средних значений по длине черешка, можно сделать следующий вывод: Δ
< НСР05. Следовательно, существенное различие между средними
значениями недостоверно, а значит, местообитание как фактор не влияет на длину
черешка. Как видно из таблицы 19, длина черешка Туапсинского района немного
превышает длину черешка Апшеронского и Сочинского районов (рисунок 15).

Рисунок 15 - Средние значения длины черешка H.
colchica, мм
H. colchica
имеет только одну лопасть, следовательно для анализа измерялась ширина и длина
(таблица 22).
Таблица 22 - Оценка средних значений ширины и
длины листовой пластинки
|
Местообитание
|
Листовая
пластинка Ширина (мм) Длина (мм)
|
|
|
SS
|
|
|
|
Апшеронский
район
|
69
|
1,9
|
91,3
|
1,9
|
|
Туапсинский
район
|
69,5
|
1,9
|
95
|
1,7
|
|
Сочинский
район
|
67
|
1,3
|
90,7
|
1,0
|
|
Примечание:
|
НСР0,5=
4,8
|
НСР0,5=
4,3
|
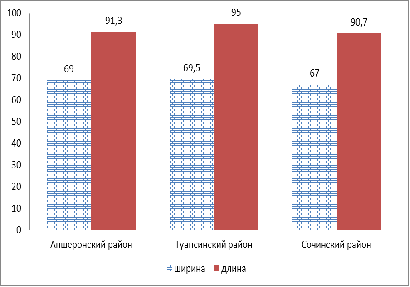
Рисунок 16- Средние значения длины и ширины
листовой пластинки H. colchica
(мм)
На рисунке 16 видно, что листовые пластинки H.
colchica имеют практически
одинаковые размеры, а это значит, что местообитание не влияет на популяционную
изменчивость этого вида.
Заключение
Согласно данным флористических сводок на
территории Западного Кавказа произрастают два вида Hedera
L.: H.
helix L.
(H. helix
L. subsp.
caucasigena) и H.
colchica (C.Koch)
C.Koch.
В определителе Косенко [Косенко, 1970, с. 248] представлен подвид H.
helix L.
- H. caucasigena
Pojark. А.И. Галушко
выделяет H. caucasigena
Pojark. в самостоятельный
вид, произрастающий на Западном Кавказе [Галушко, 1978, с. 297].
Сбор данных для этой работы проводился в
следующих районах: Туапсинский район (дубовый лес, с. Малое Псеушхо, южный
склон; верховье реки Цыпка, дубово-буковый лес), Геленджикский район (дубовый
лес, 2 км до с. Дтвноморское, южный склон), Новороссийский район (дубовый лес,
1 км до поселка Верхне-Баканский), район МО города-курорта Анапы
(широколиственный лес, 1 км до станицы Гостагаевская, северный склон), Крымский
район (дубовый лес, река Баканка, северный склон), Апшеронский район (пойменный
лес, 3 км до села Черниговское; буковый лес, вход в Гуамское ущелье), Сочинский
район (поселок Совет Кваджи, склон у трассы) в 2010 - 2013 г
В работе использовались методы: метод измерения
и статистический метод. Отдельная листовая пластинка была измерена по трем
показателям: длина черешка (L);
количество лопастей (n)
и длина каждой лопасти (l).
Все необходимые вычисления выполнены на ПК с использованием компьютерной
программы Excel.
Произведя биометрическое измерение
морфологических признаков листа, мы видим, что у популяций H.
helix L.
subsp. caucasigena
наибольшей биометрической изменчивости подвергается глубина залегания лопастей,
а наиболее постоянными признаки являются длина черешка и количество лопастей.
Для H. colchica
характерно наличие одной лопасти, а значит, этот признак является постоянным.
Размеры ширины и длины в значительной степени варьируют.
Одна из задач работы заключается в изучении
влияния абиотических факторов на популяционную изменчивость видов H. helix L.
subsp. сaucasigena и H.
colchica (C.Koch)
C.Koch
(длина черешка, количество лопастей, глубина лопасти) и изучит морфометрические
особенности разных популяций видов Hedera L.
Для того, чтобы изучить влияние абиотических
факторов (местообитания) на признаки, был проведен анализ вариационного ряда и
затем, в результате полученных данных, произведен расчет наименьшей
существенной разницы (НСР05), с помощью которой возможно доказать
существенность различий между средними значениями.
В результате проведенного анализа было
установлено, что изменчивость H. helix L. subsp. сaucasigenа зависит от условия
местообитания популяции. Длина черешка и глубина залегания лопасти в
значительной степени варьируют. В местах с повышенной влажность, тенистостью
(расположение на северных склонах) лист имеет более крупные размеры (Крымский,
Новороссийский, Анапский районы). Также было уставлено, что количество лопастей
не зависит от абиотических факторов. Наиболее встречаемы трехлопастные листья.
Затем, для более полного изучения популяционной
изменчивости вида Hedera
helix (также на примере
листа), проводилось сравнение популяций видов Западного Кавказа и популяций
видов Средиземноморских местообитаний (Испания, провинция Каталония). Длина
черешка видов, произрастающих в Средиземноморье на много отличается от
Западно-Кавказских видов и составляет 120 мм, тогда как виды западного Кавказа
имеют среднюю длину 58 мм. На Западном Кавказе преобладают трехлопастные листья
(65%), у вида, произрастающего в Испании - пятилопастные листья (68%).
Сравнительный анализ глубины лопасти проводился на трехлопастных листьях.
Каждая лопасть рассматривается как отдельный признак. Очевидно, что вид H.
helix, произрастающий в
Средиземноморье имеет более крупную листовую пластинку по сравнению с
Западно-кавказским подвидом H.helix
L. subsp.
сaucasigena. Морфология
листьев H. helix
L. subsp.
caucasigena имеет значительные
отличия от средиземноморских видов H.
helix L.,
что обуславливает выделения подвида H.
helix L.
subsp. caucasigena
в самостоятельный вид, встречающийся на Западном Кавказе.
Таким образом, мы видим, что один и тот же вид Hedera
L.в зависимости от
условий местообитания (внешних факторов) изменчив по морфологическим признакам.
Проведя анализ влияния абиотических факторов на
морфологическую изменчивость листа H. colchica
(C.Koch)
C.Koch
мы убедились, что местообитание как фактор не влияет на биометрию признаков.
Это связано с тем, что взятые образцы данного вида произрастают в схожих
условиях.
Список используемых источников
Атлас
Краснодарского края и республики Адыгея // Минск, 1996. 48 c.
Бородина
Н.А., Некрасов В.И., Некрасова Н.С., Петрова И.П., Плотникова Л.С., Смирнова
Н.Г. Деревья и кустарники СССР. М., 1966. 637 с.
Галушко
А.И. Флора Северного Кавказа. Ростов н/Д., 1978. Т. 3. 320 с.
Гроссгейм
А.А. Определитель растений Кавказа. М., 1949. 747 с.
Гвоздецкий
Н.А. Физическая география Кавказа. М., 1954. 234 с.
Зернов
А.С. Флора Северо-Западного Кавказа. М., 2006. 664 с.
Конспект
флоры Кавказа / отв. ред. А.Л. Тахтаджян. Т. 1 / под. ред. Ю.Л. Меницкого, Т.Н.
Поповой. СПб., 2006. 204 с.
Конспект
флоры Кавказа / отв. ред. А.Л. Тахтаджян. Т. 3 (1) / под. ред. Ю.Л. Меницкого,
Т.Н. Поповой. СПб., 2009. 469 с.
Косенко
И.С. Определитель высших растений северо-западного Кавказа и Предкавказья. М.,
1970. 613 с.
Литвинская
С.А. Деревья и кустарники Кубани. Ростов н/Д., 1993. 230 с.
Любимова
Т.В., Бондаренко Н.А., Куропаткина Т.Н. Инженерно-геологические условия
Черноморского побережья Северо-Западного Кавказа. Краснодар, 2009. 211 с.
Мильков
Ф.Н., Гвоздецкий Н.А. Физическая география СССР. М., 1976. 307 с.
Покровская
В.С. Природа Краснодарского края, 1979. 275 с.
Соляник
Г.М. Почвы Краснодарского края. Краснодар, 2004. 70 с.
Тонконоженко
Е.В. Природа Краснодарского края, Почвы. Краснодар, 1979. 275 с.
Флора
СССР / отв. ред. Б. К. Шишкин. Т. 16 / под. ред. Е.Г. Боброва, Е.П. Коровина,
А.М. Крищтофовича, И.А. Линчевской, Б.А. Федченко. М., 1950. 339 с.
Приложение. Листовые пластинки Hedera L., над
которыми проводились исследования и результаты статистической обработки
Таблица А. 1 - Морфометрическая характеристика
вида H. helix
L. subsp.
caucasigena (Туапсинский
район, дубовый лес, с. Малая Псеушхо)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Глубина
лопасти (l), мм
|
|
|
|
1
|
2
|
3
|
4
|
5
|
7
|
|
1
|
51
|
7
|
3
|
17
|
5
|
23
|
5
|
14
|
2
|
|
2
|
53
|
5
|
5
|
20
|
29
|
21
|
5
|
-
|
-
|
|
3
|
52
|
5
|
2
|
17
|
35
|
11
|
2
|
-
|
-
|
|
4
|
26
|
3
|
7
|
28
|
10
|
-
|
-
|
-
|
-
|
|
5
|
13
|
3
|
7
|
29
|
8
|
-
|
-
|
-
|
-
|
|
6
|
49
|
7
|
6
|
26
|
21
|
25
|
16
|
24
|
5
|
|
7
|
35
|
3
|
9
|
26
|
9
|
-
|
-
|
-
|
-
|
|
8
|
8
|
3
|
12
|
31
|
9
|
-
|
-
|
-
|
-
|
|
9
|
42
|
5
|
4
|
14
|
29
|
14
|
4
|
-
|
-
|
|
10
|
39
|
5
|
20
|
14
|
22
|
15
|
23
|
-
|
-
|
|
11
|
60
|
5
|
21
|
15
|
30
|
16
|
20
|
-
|
-
|
|
12
|
44
|
7
|
2
|
16
|
16
|
17
|
10
|
16
|
2
|
|
13
|
65
|
7
|
21
|
16
|
7
|
26
|
16
|
27
|
6
|
|
14
|
27
|
5
|
6
|
23
|
41
|
19
|
5
|
-
|
-
|
|
15
|
33
|
5
|
14
|
12
|
21
|
11
|
15
|
-
|
-
|
|
16
|
31
|
5
|
16
|
7
|
20
|
6
|
17
|
-
|
-
|
|
17
|
59
|
3
|
20
|
40
|
22
|
-
|
-
|
-
|
-
|
|
18
|
21
|
3
|
7
|
33
|
13
|
-
|
-
|
-
|
-
|
|
19
|
33
|
7
|
2
|
13
|
7
|
21
|
7
|
14
|
2
|
|
20
|
20
|
3
|
11
|
39
|
15
|
-
|
-
|
-
|
-
|
|
21
|
40
|
5
|
3
|
10
|
24
|
16
|
3
|
-
|
-
|
|
22
|
51
|
5
|
16
|
14
|
23
|
15
|
18
|
-
|
-
|
|
23
|
12
|
3
|
3
|
23
|
3
|
-
|
-
|
-
|
-
|
|
24
|
73
|
5
|
3
|
26
|
51
|
32
|
7
|
-
|
-
|
|
25
|
53
|
5
|
16
|
11
|
32
|
15
|
20
|
-
|
-
|
|
26
|
55
|
7
|
5
|
7
|
22
|
40
|
24
|
9
|
11
|
|
27
|
15
|
3
|
8
|
23
|
10
|
-
|
-
|
-
|
-
|
|
29
|
53
|
5
|
20
|
13
|
26
|
15
|
24
|
-
|
-
|
|
30
|
80
|
3
|
18
|
52
|
18
|
-
|
-
|
-
|
-
|
|
31
|
63
|
3
|
16
|
39
|
18
|
-
|
-
|
-
|
-
|
|
32
|
51
|
5
|
6
|
15
|
31
|
16
|
9
|
-
|
-
|
|
33
|
55
|
7
|
10
|
11
|
2
|
16
|
2
|
6
|
9
|
|
34
|
26
|
3
|
20
|
40
|
22
|
-
|
-
|
-
|
-
|
|
35
|
55
|
7
|
5
|
19
|
13
|
31
|
14
|
16
|
5
|
|
36
|
45
|
7
|
6
|
33
|
20
|
26
|
16
|
27
|
|
37
|
62
|
7
|
2
|
20
|
14
|
28
|
15
|
21
|
7
|
|
38
|
45
|
5
|
2
|
6
|
25
|
9
|
21
|
-
|
-
|
|
39
|
28
|
5
|
16
|
11
|
16
|
10
|
11
|
-
|
-
|
|
40
|
35
|
5
|
2
|
8
|
27
|
10
|
2
|
-
|
-
|
|
41
|
43
|
5
|
3
|
30
|
41
|
28
|
5
|
-
|
-
|
|
42
|
38
|
5
|
5
|
26
|
41
|
21
|
5
|
-
|
-
|
|
43
|
71
|
5
|
5
|
23
|
44
|
15
|
25
|
-
|
-
|
|
44
|
38
|
3
|
16
|
45
|
23
|
-
|
-
|
-
|
-
|
|
45
|
39
|
3
|
18
|
46
|
19
|
-
|
-
|
-
|
-
|
|
46
|
60
|
3
|
20
|
35
|
25
|
-
|
-
|
-
|
-
|
|
47
|
42
|
7
|
19
|
14
|
5
|
20
|
6
|
9
|
10
|
|
48
|
56
|
5
|
2
|
19
|
35
|
18
|
2
|
-
|
-
|
|
49
|
28
|
3
|
11
|
41
|
12
|
-
|
-
|
-
|
-
|
|
50
|
30
|
5
|
14
|
9
|
22
|
6
|
11
|
-
|
-
|
|
51
|
15
|
3
|
10
|
20
|
10
|
-
|
-
|
-
|
-
|
|
52
|
45
|
7
|
3
|
15
|
11
|
22
|
10
|
16
|
4
|
|
53
|
41
|
5
|
2
|
14
|
26
|
14
|
5
|
-
|
-
|
|
54
|
26
|
3
|
13
|
41
|
12
|
-
|
-
|
-
|
-
|
|
55
|
52
|
5
|
2
|
19
|
38
|
20
|
2
|
-
|
-
|
|
56
|
61
|
5
|
9
|
20
|
35
|
16
|
7
|
-
|
-
|
|
57
|
50
|
7
|
3
|
19
|
18
|
23
|
12
|
20
|
2
|
|
58
|
32
|
7
|
5
|
12
|
12
|
23
|
12
|
11
|
3
|
|
59
|
22
|
3
|
11
|
33
|
11
|
-
|
-
|
-
|
-
|
|
60
|
22
|
3
|
10
|
27
|
5
|
-
|
-
|
-
|
-
|
|
61
|
18
|
3
|
6
|
30
|
10
|
-
|
-
|
-
|
-
|
|
62
|
12
|
3
|
7
|
23
|
10
|
-
|
-
|
-
|
-
|
|
63
|
50
|
5
|
2
|
18
|
40
|
18
|
2
|
-
|
-
|
|
64
|
32
|
7
|
2
|
11
|
6
|
20
|
6
|
10
|
3
|
|
66
|
26
|
7
|
8
|
2
|
2
|
11
|
2
|
2
|
14
|
|
67
|
28
|
3
|
9
|
25
|
7
|
-
|
-
|
-
|
-
|
|
68
|
18
|
3
|
7
|
23
|
12
|
-
|
-
|
-
|
-
|
|
69
|
18
|
3
|
7
|
28
|
8
|
-
|
-
|
-
|
-
|
|
70
|
11
|
3
|
7
|
21
|
5
|
-
|
-
|
-
|
-
|
|
71
|
10
|
3
|
10
|
25
|
10
|
-
|
-
|
-
|
-
|
|
72
|
12
|
3
|
11
|
28
|
7
|
-
|
-
|
-
|
-
|
35
|
7
|
6
|
14
|
5
|
17
|
7
|
13
|
3
|
|
74
|
48
|
7
|
3
|
16
|
7
|
21
|
10
|
15
|
7
|
|
75
|
15
|
3
|
7
|
25
|
7
|
-
|
-
|
-
|
-
|
|
76
|
18
|
3
|
11
|
26
|
8
|
-
|
-
|
-
|
-
|
|
77
|
26
|
7
|
2
|
11
|
10
|
20
|
7
|
10
|
2
|
|
78
|
21
|
3
|
9
|
26
|
9
|
-
|
-
|
-
|
-
|
|
79
|
10
|
3
|
5
|
13
|
9
|
-
|
-
|
-
|
-
|
|
80
|
20
|
5
|
10
|
6
|
16
|
5
|
10
|
-
|
-
|
|
81
|
11
|
3
|
5
|
30
|
5
|
-
|
-
|
-
|
-
|
|
82
|
23
|
3
|
5
|
16
|
6
|
-
|
-
|
-
|
-
|
|
83
|
19
|
3
|
7
|
31
|
6
|
-
|
-
|
-
|
-
|
|
84
|
40
|
5
|
5
|
14
|
25
|
15
|
5
|
-
|
-
|
|
85
|
33
|
3
|
8
|
33
|
5
|
-
|
-
|
-
|
-
|
|
86
|
47
|
3
|
11
|
5
|
9
|
-
|
-
|
-
|
-
|
|
87
|
28
|
3
|
4
|
14
|
6
|
-
|
-
|
-
|
-
|
|
88
|
21
|
3
|
5
|
16
|
3
|
-
|
-
|
-
|
-
|
|
89
|
38
|
3
|
12
|
28
|
3
|
-
|
-
|
-
|
-
|
|
90
|
11
|
5
|
2
|
6
|
17
|
5
|
11
|
-
|
-
|
|
91
|
45
|
3
|
10
|
23
|
11
|
-
|
-
|
-
|
-
|
|
92
|
35
|
3
|
8
|
23
|
5
|
-
|
-
|
-
|
-
|
|
93
|
17
|
5
|
3
|
5
|
16
|
5
|
4
|
-
|
-
|
|
94
|
25
|
5
|
6
|
8
|
12
|
8
|
4
|
-
|
-
|
|
95
|
41
|
5
|
7
|
9
|
19
|
6
|
2
|
-
|
-
|
|
96
|
18
|
3
|
4
|
12
|
5
|
-
|
-
|
-
|
-
|
|
97
|
50
|
5
|
7
|
14
|
19
|
12
|
6
|
-
|
-
|
|
98
|
23
|
3
|
6
|
18
|
8
|
-
|
-
|
-
|
-
|
|
99
|
43
|
3
|
12
|
25
|
11
|
-
|
-
|
-
|
|
|
100
|
29
|
3
|
4
|
16
|
7
|
-
|
-
|
-
|
-
|
Таблица А. 2 - Морфометрическая характеристика
вида H. helix
L. subsp.
caucasigena (Крымский район,
дубовый лес, река Баканка)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Глубина
лопасти (l), мм
|
|
|
|
1
|
2
|
3
|
4
|
5
|
6
|
7
|
|
1
|
110
|
3
|
24
|
90
|
15
|
-
|
-
|
-
|
-
|
|
2
|
95
|
5
|
3
|
35
|
46
|
21
|
2
|
-
|
-
|
|
3
|
133
|
3
|
20
|
69
|
13
|
-
|
-
|
-
|
-
|
|
4
|
81
|
5
|
3
|
10
|
32
|
14
|
2
|
-
|
-
|
|
5
|
115
|
3
|
22
|
44
|
31
|
-
|
-
|
-
|
-
|
|
6
|
80
|
5
|
2
|
21
|
49
|
20
|
3
|
-
|
-
|
73
|
5
|
3
|
18
|
50
|
17
|
3
|
-
|
-
|
|
8
|
55
|
3
|
25
|
55
|
21
|
-
|
-
|
-
|
-
|
|
9
|
33
|
3
|
15
|
33
|
14
|
-
|
-
|
-
|
-
|
|
10
|
78
|
3
|
16
|
35
|
17
|
-
|
-
|
-
|
-
|
|
11
|
77
|
3
|
7
|
41
|
3
|
-
|
-
|
-
|
-
|
|
12
|
67
|
3
|
7
|
24
|
5
|
-
|
-
|
-
|
-
|
|
13
|
75
|
3
|
7
|
42
|
9
|
-
|
-
|
-
|
-
|
|
14
|
81
|
3
|
17
|
51
|
20
|
-
|
-
|
-
|
-
|
|
15
|
85
|
7
|
5
|
12
|
2
|
14
|
3
|
13
|
5
|
|
16
|
93
|
5
|
6
|
16
|
35
|
14
|
2
|
-
|
-
|
|
17
|
100
|
5
|
2
|
13
|
34
|
13
|
1
|
-
|
-
|
|
18
|
75
|
3
|
14
|
26
|
12
|
-
|
-
|
-
|
-
|
|
19
|
55
|
3
|
12
|
30
|
7
|
-
|
-
|
-
|
-
|
|
21
|
80
|
5
|
5
|
8
|
19
|
9
|
2
|
-
|
-
|
|
22
|
74
|
5
|
3
|
10
|
40
|
12
|
2
|
-
|
-
|
|
23
|
100
|
3
|
17
|
39
|
9
|
-
|
-
|
-
|
-
|
|
24
|
71
|
5
|
6
|
17
|
38
|
25
|
7
|
-
|
-
|
|
25
|
87
|
3
|
5
|
60
|
6
|
-
|
-
|
-
|
-
|
|
26
|
87
|
3
|
20
|
35
|
18
|
-
|
-
|
-
|
-
|
|
27
|
90
|
3
|
7
|
34
|
9
|
-
|
-
|
-
|
-
|
|
28
|
60
|
3
|
11
|
49
|
16
|
-
|
-
|
-
|
-
|
|
29
|
61
|
3
|
9
|
27
|
9
|
|
-
|
-
|
-
|
|
30
|
142
|
5
|
5
|
12
|
30
|
11
|
4
|
-
|
-
|
|
31
|
89
|
3
|
7
|
32
|
7
|
-
|
-
|
-
|
-
|
|
32
|
92
|
5
|
16
|
4
|
20
|
16
|
3
|
-
|
-
|
|
33
|
70
|
5
|
1
|
10
|
30
|
6
|
2
|
-
|
-
|
|
34
|
107
|
3
|
5
|
62
|
4
|
-
|
-
|
-
|
-
|
|
35
|
60
|
3
|
21
|
43
|
20
|
-
|
-
|
-
|
-
|
|
36
|
81
|
3
|
7
|
26
|
5
|
-
|
-
|
-
|
-
|
|
37
|
84
|
3
|
5
|
20
|
3
|
-
|
-
|
-
|
-
|
|
38
|
85
|
5
|
1
|
10
|
30
|
8
|
1
|
-
|
-
|
|
39
|
107
|
5
|
1
|
12
|
60
|
15
|
3
|
-
|
-
|
|
40
|
90
|
3
|
25
|
35
|
17
|
-
|
-
|
-
|
-
|
|
41
|
68
|
3
|
4
|
53
|
7
|
-
|
-
|
-
|
-
|
|
42
|
60
|
3
|
18
|
45
|
17
|
-
|
-
|
-
|
-
|
|
43
|
118
|
12
|
50
|
14
|
-
|
-
|
-
|
-
|
|
44
|
110
|
5
|
9
|
33
|
44
|
32
|
9
|
-
|
-
|
|
45
|
83
|
3
|
22
|
47
|
29
|
-
|
-
|
-
|
-
|
|
46
|
115
|
3
|
28
|
56
|
19
|
-
|
-
|
-
|
-
|
|
47
|
75
|
5
|
4
|
20
|
39
|
12
|
2
|
-
|
-
|
|
48
|
76
|
3
|
27
|
47
|
20
|
-
|
-
|
-
|
-
|
|
49
|
10
|
3
|
9
|
35
|
8
|
-
|
-
|
-
|
-
|
|
50
|
104
|
3
|
17
|
41
|
16
|
-
|
-
|
-
|
-
|
|
51
|
130
|
5
|
5
|
20
|
60
|
21
|
3
|
-
|
-
|
|
52
|
70
|
5
|
3
|
15
|
43
|
14
|
3
|
-
|
-
|
|
53
|
20
|
3
|
14
|
34
|
9
|
-
|
-
|
-
|
|
|
54
|
60
|
3
|
20
|
35
|
11
|
-
|
-
|
-
|
-
|
|
55
|
100
|
5
|
1
|
6
|
28
|
14
|
4
|
-
|
-
|
|
56
|
72
|
3
|
10
|
25
|
8
|
-
|
-
|
-
|
-
|
|
57
|
53
|
3
|
6
|
12
|
6
|
-
|
-
|
-
|
-
|
|
58
|
121
|
5
|
3
|
5
|
34
|
6
|
3
|
-
|
-
|
|
59
|
73
|
5
|
4
|
17
|
45
|
14
|
4
|
-
|
-
|
|
60
|
85
|
5
|
4
|
20
|
55
|
26
|
10
|
-
|
-
|
|
61
|
88
|
5
|
14
|
25
|
57
|
19
|
5
|
-
|
-
|
|
62
|
92
|
3
|
4
|
46
|
6
|
-
|
-
|
-
|
-
|
|
63
|
82
|
5
|
4
|
16
|
55
|
22
|
5
|
-
|
-
|
|
64
|
78
|
5
|
3
|
20
|
37
|
20
|
1
|
-
|
-
|
|
65
|
79
|
5
|
18
|
3
|
23
|
3
|
19
|
-
|
-
|
|
66
|
140
|
5
|
1
|
9
|
27
|
7
|
1
|
-
|
-
|
|
67
|
90
|
5
|
2
|
27
|
50
|
26
|
3
|
-
|
-
|
|
68
|
52
|
3
|
13
|
46
|
4
|
-
|
-
|
-
|
-
|
|
69
|
95
|
5
|
2
|
33
|
43
|
25
|
7
|
-
|
-
|
|
70
|
55
|
5
|
2
|
7
|
29
|
8
|
1
|
-
|
-
|
|
71
|
85
|
5
|
3
|
13
|
40
|
10
|
1
|
-
|
-
|
|
72
|
93
|
5
|
1
|
5
|
35
|
10
|
1
|
-
|
-
|
|
73
|
92
|
5
|
3
|
7
|
30
|
9
|
2
|
-
|
-
|
|
74
|
95
|
5
|
2
|
7
|
15
|
9
|
3
|
-
|
-
|
|
75
|
70
|
3
|
10
|
25
|
10
|
-
|
-
|
-
|
-
|
|
76
|
95
|
3
|
21
|
50
|
22
|
-
|
-
|
-
|
-
|
|
77
|
80
|
5
|
5
|
10
|
46
|
15
|
1
|
-
|
-
|
|
78
|
94
|
5
|
2
|
21
|
8
|
2
|
-
|
-
|
|
79
|
110
|
5
|
4
|
10
|
42
|
2
|
7
|
-
|
-
|
|
80
|
62
|
5
|
2
|
10
|
25
|
12
|
1
|
-
|
-
|
|
81
|
65
|
5
|
4
|
20
|
52
|
16
|
1
|
-
|
-
|
|
82
|
100
|
3
|
14
|
41
|
9
|
0
|
-
|
-
|
-
|
|
83
|
70
|
5
|
1
|
40
|
67
|
26
|
3
|
-
|
-
|
|
84
|
69
|
3
|
20
|
43
|
15
|
-
|
-
|
-
|
-
|
|
85
|
53
|
5
|
1
|
6
|
20
|
5
|
2
|
-
|
-
|
|
86
|
92
|
5
|
10
|
9
|
42
|
11
|
2
|
-
|
-
|
|
87
|
75
|
5
|
2
|
14
|
22
|
13
|
-
|
-
|
-
|
|
88
|
103
|
3
|
14
|
28
|
10
|
-
|
-
|
-
|
-
|
|
89
|
107
|
3
|
10
|
31
|
8
|
-
|
-
|
-
|
-
|
|
90
|
81
|
3
|
9
|
33
|
3
|
-
|
-
|
-
|
-
|
|
91
|
80
|
5
|
5
|
21
|
32
|
19
|
3
|
-
|
-
|
|
93
|
44
|
5
|
3
|
10
|
35
|
8
|
1
|
-
|
-
|
|
94
|
97
|
5
|
5
|
12
|
50
|
11
|
6
|
-
|
-
|
|
95
|
53
|
5
|
2
|
6
|
67
|
5
|
1
|
-
|
-
|
|
96
|
103
|
5
|
5
|
25
|
36
|
21
|
3
|
-
|
-
|
|
97
|
62
|
3
|
10
|
65
|
12
|
-
|
-
|
-
|
-
|
|
98
|
103
|
5
|
3
|
24
|
45
|
14
|
6
|
-
|
-
|
|
99
|
80
|
5
|
1
|
20
|
11
|
16
|
4
|
-
|
-
|
|
100
|
40
|
3
|
18
|
22
|
9
|
-
|
-
|
-
|
-
|
Таблица А. 3 - Морфометрическая характеристика
вида H. helix L. subsp. caucasigena (МО город-курорт Анапа, широколиственный
лес, 1 км до ст. Гостагаевская)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Глубина
лопасти (l), мм
|
|
|
|
1
|
2
|
3
|
4
|
5
|
|
1
|
79
|
5
|
5
|
23
|
62
|
31
|
9
|
|
2
|
86
|
5
|
1
|
31
|
56
|
31
|
5
|
|
3
|
88
|
3
|
21
|
56
|
22
|
-
|
-
|
|
4
|
75
|
5
|
12
|
31
|
52
|
29
|
10
|
|
5
|
86
|
5
|
4
|
25
|
52
|
28
|
5
|
|
6
|
113
|
3
|
17
|
53
|
16
|
-
|
-
|
|
7
|
95
|
3
|
37
|
74
|
42
|
-
|
-
|
|
8
|
111
|
5
|
5
|
37
|
57
|
29
|
1
|
|
9
|
79
|
3
|
27
|
67
|
25
|
-
|
-
|
|
10
|
78
|
5
|
2
|
25
|
58
|
26
|
1
|
|
11
|
129
|
3
|
26
|
65
|
24
|
-
|
-
|
|
12
|
131
|
3
|
26
|
64
|
27
|
-
|
-
|
|
13
|
140
|
5
|
1
|
33
|
74
|
31
|
5
|
|
15
|
88
|
3
|
17
|
53
|
24
|
-
|
-
|
|
16
|
121
|
3
|
29
|
65
|
19
|
-
|
-
|
|
17
|
135
|
5
|
15
|
41
|
45
|
3
|
|
18
|
108
|
3
|
29
|
90
|
36
|
-
|
-
|
|
19
|
100
|
5
|
9
|
15
|
70
|
15
|
7
|
|
20
|
118
|
5
|
5
|
29
|
54
|
29
|
6
|
|
21
|
98
|
5
|
7
|
36
|
60
|
46
|
8
|
|
22
|
109
|
5
|
11
|
34
|
65
|
31
|
8
|
|
23
|
130
|
5
|
1
|
36
|
86
|
29
|
6
|
|
24
|
86
|
5
|
19
|
29
|
43
|
28
|
17
|
|
25
|
95
|
5
|
2
|
39
|
71
|
43
|
6
|
|
26
|
112
|
3
|
30
|
59
|
26
|
-
|
-
|
|
27
|
65
|
5
|
11
|
33
|
63
|
24
|
7
|
|
28
|
95
|
5
|
8
|
30
|
60
|
31
|
8
|
|
29
|
95
|
3
|
17
|
43
|
16
|
-
|
-
|
|
30
|
110
|
5
|
7
|
20
|
46
|
21
|
8
|
|
31
|
55
|
5
|
31
|
4
|
21
|
57
|
7
|
|
32
|
108
|
3
|
23
|
54
|
19
|
-
|
-
|
|
33
|
102
|
5
|
3
|
20
|
45
|
23
|
5
|
|
34
|
113
|
5
|
5
|
25
|
52
|
23
|
4
|
|
35
|
53
|
5
|
18
|
25
|
46
|
23
|
21
|
|
36
|
80
|
3
|
15
|
53
|
23
|
-
|
-
|
|
37
|
119
|
5
|
14
|
37
|
55
|
40
|
11
|
|
38
|
81
|
5
|
9
|
38
|
65
|
33
|
5
|
|
39
|
98
|
3
|
25
|
57
|
18
|
-
|
-
|
|
40
|
82
|
3
|
31
|
57
|
17
|
-
|
-
|
|
41
|
54
|
3
|
35
|
60
|
20
|
-
|
-
|
|
42
|
115
|
3
|
26
|
49
|
25
|
-
|
-
|
|
43
|
85
|
5
|
5
|
18
|
40
|
16
|
5
|
|
44
|
55
|
3
|
18
|
48
|
23
|
-
|
-
|
|
45
|
96
|
3
|
30
|
59
|
30
|
-
|
-
|
|
46
|
53
|
3
|
21
|
53
|
14
|
-
|
-
|
|
47
|
130
|
3
|
32
|
70
|
23
|
-
|
-
|
|
48
|
30
|
3
|
15
|
52
|
18
|
-
|
-
|
|
49
|
65
|
5
|
5
|
27
|
65
|
25
|
1
|
|
50
|
106
|
3
|
21
|
45
|
15
|
-
|
-
|
|
51
|
130
|
5
|
11
|
27
|
71
|
35
|
10
|
|
52
|
51
|
5
|
3
|
21
|
35
|
16
|
3
|
|
53
|
51
|
5
|
3
|
20
|
52
|
19
|
8
|
|
54
|
96
|
5
|
5
|
2
|
55
|
22
|
3
|
|
55
|
81
|
3
|
21
|
63
|
26
|
-
|
-
|
|
56
|
106
|
3
|
19
|
35
|
17
|
-
|
-
|
|
57
|
90
|
5
|
2
|
21
|
52
|
20
|
1
|
|
58
|
37
|
5
|
8
|
6
|
54
|
8
|
7
|
|
59
|
55
|
5
|
4
|
23
|
58
|
19
|
3
|
|
60
|
80
|
3
|
24
|
62
|
30
|
-
|
-
|
|
61
|
75
|
5
|
7
|
42
|
22
|
4
|
|
62
|
145
|
5
|
5
|
24
|
59
|
19
|
4
|
|
63
|
45
|
3
|
17
|
3
|
21
|
-
|
-
|
|
64
|
52
|
5
|
3
|
13
|
46
|
14
|
3
|
|
65
|
95
|
3
|
43
|
79
|
33
|
-
|
-
|
|
66
|
75
|
5
|
7
|
19
|
60
|
24
|
5
|
|
67
|
96
|
5
|
6
|
20
|
44
|
22
|
8
|
|
68
|
52
|
3
|
35
|
50
|
34
|
-
|
-
|
|
70
|
83
|
5
|
5
|
21
|
53
|
18
|
5
|
|
71
|
147
|
5
|
7
|
17
|
33
|
12
|
8
|
|
72
|
123
|
3
|
15
|
48
|
11
|
-
|
-
|
|
73
|
61
|
5
|
7
|
25
|
40
|
23
|
7
|
|
74
|
55
|
3
|
16
|
47
|
16
|
-
|
-
|
|
75
|
111
|
3
|
15
|
31
|
20
|
-
|
-
|
|
76
|
77
|
5
|
14
|
24
|
58
|
32
|
24
|
|
77
|
98
|
5
|
6
|
14
|
61
|
22
|
7
|
|
78
|
69
|
5
|
5
|
12
|
39
|
9
|
10
|
|
79
|
63
|
3
|
14
|
62
|
12
|
-
|
-
|
|
80
|
86
|
5
|
3
|
20
|
38
|
14
|
2
|
|
81
|
32
|
3
|
19
|
42
|
19
|
-
|
-
|
|
82
|
78
|
3
|
17
|
41
|
12
|
-
|
-
|
|
83
|
60
|
3
|
24
|
48
|
16
|
-
|
-
|
|
84
|
109
|
5
|
2
|
20
|
43
|
20
|
2
|
|
85
|
76
|
5
|
4
|
20
|
40
|
17
|
5
|
|
86
|
90
|
5
|
2
|
12
|
30
|
11
|
2
|
|
87
|
120
|
3
|
18
|
74
|
24
|
-
|
-
|
|
88
|
98
|
3
|
12
|
46
|
11
|
-
|
-
|
|
89
|
110
|
3
|
32
|
65
|
31
|
-
|
-
|
|
90
|
98
|
5
|
7
|
15
|
46
|
7
|
6
|
|
91
|
60
|
5
|
2
|
12
|
32
|
14
|
2
|
|
92
|
58
|
5
|
11
|
23
|
31
|
25
|
10
|
|
93
|
65
|
5
|
2
|
17
|
62
|
20
|
2
|
|
94
|
40
|
5
|
5
|
7
|
39
|
10
|
2
|
|
95
|
67
|
5
|
4
|
12
|
47
|
15
|
4
|
|
96
|
25
|
3
|
20
|
37
|
15
|
-
|
-
|
|
97
|
86
|
5
|
10
|
9
|
71
|
21
|
7
|
|
98
|
96
|
5
|
2
|
10
|
41
|
10
|
2
|
|
99
|
48
|
3
|
15
|
37
|
15
|
-
|
-
|
|
100
|
75
|
5
|
11
|
40
|
59
|
38
|
23
|
Таблица А. 5 - Морфометрическая характеристика
вида H. helix L. subsp.
caucasigena (Новороссийский
район, дубовый лес, 1 км до п. Верхне - Баканский)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Глубина
лопасти (l), мм
|
|
|
|
1
|
2
|
3
|
4
|
5
|
|
1
|
110
|
5
|
11
|
32
|
50
|
35
|
9
|
|
2
|
85
|
3
|
14
|
45
|
16
|
-
|
-
|
|
4
|
74
|
5
|
2
|
14
|
35
|
21
|
3
|
|
5
|
77
|
3
|
29
|
53
|
24
|
-
|
-
|
|
6
|
5
|
9
|
22
|
64
|
25
|
10
|
|
7
|
70
|
5
|
5
|
26
|
51
|
18
|
8
|
|
8
|
53
|
3
|
9
|
45
|
15
|
-
|
-
|
|
9
|
78
|
5
|
13
|
21
|
53
|
16
|
7
|
|
10
|
82
|
5
|
4
|
9
|
46
|
15
|
5
|
|
11
|
44
|
3
|
14
|
27
|
14
|
-
|
-
|
|
12
|
65
|
5
|
2
|
17
|
37
|
20
|
2
|
|
13
|
97
|
5
|
4
|
21
|
47
|
31
|
3
|
|
14
|
99
|
5
|
15
|
27
|
56
|
7
|
11
|
|
15
|
74
|
5
|
9
|
20
|
35
|
18
|
6
|
|
16
|
49
|
3
|
10
|
31
|
15
|
-
|
-
|
|
17
|
47
|
3
|
11
|
35
|
10
|
-
|
-
|
|
18
|
46
|
5
|
2
|
9
|
33
|
9
|
4
|
|
19
|
21
|
3
|
10
|
34
|
10
|
-
|
-
|
|
20
|
87
|
5
|
9
|
13
|
34
|
21
|
3
|
|
21
|
84
|
5
|
6
|
17
|
47
|
21
|
7
|
|
22
|
93
|
5
|
1
|
24
|
43
|
19
|
5
|
|
23
|
75
|
5
|
5
|
21
|
45
|
25
|
3
|
|
24
|
67
|
5
|
7
|
25
|
38
|
24
|
9
|
|
25
|
77
|
3
|
23
|
45
|
23
|
-
|
-
|
|
26
|
35
|
3
|
5
|
30
|
9
|
-
|
-
|
|
27
|
81
|
3
|
21
|
47
|
22
|
-
|
-
|
|
28
|
95
|
5
|
14
|
32
|
36
|
19
|
5
|
|
29
|
73
|
5
|
4
|
22
|
34
|
25
|
9
|
|
30
|
79
|
5
|
4
|
21
|
60
|
17
|
5
|
|
31
|
87
|
3
|
20
|
45
|
21
|
-
|
-
|
|
32
|
82
|
3
|
22
|
30
|
15
|
-
|
-
|
|
33
|
68
|
5
|
9
|
22
|
55
|
21
|
7
|
|
34
|
55
|
5
|
2
|
10
|
34
|
9
|
3
|
|
35
|
53
|
3
|
10
|
33
|
9
|
-
|
-
|
|
36
|
64
|
5
|
3
|
7
|
43
|
8
|
4
|
|
37
|
70
|
3
|
17
|
34
|
18
|
-
|
-
|
|
38
|
64
|
5
|
9
|
20
|
35
|
19
|
9
|
|
39
|
96
|
3
|
21
|
43
|
21
|
-
|
-
|
|
40
|
66
|
5
|
4
|
9
|
45
|
9
|
3
|
|
41
|
70
|
3
|
20
|
37
|
19
|
-
|
-
|
|
42
|
84
|
3
|
13
|
30
|
6
|
-
|
-
|
|
43
|
73
|
5
|
4
|
20
|
44
|
21
|
5
|
|
44
|
71
|
5
|
10
|
23
|
49
|
18
|
11
|
|
45
|
50
|
5
|
3
|
18
|
29
|
14
|
5
|
|
46
|
73
|
3
|
13
|
27
|
15
|
-
|
-
|
|
47
|
113
|
5
|
16
|
32
|
56
|
21
|
10
|
|
48
|
93
|
3
|
35
|
60
|
33
|
-
|
-
|
|
49
|
122
|
3
|
17
|
51
|
20
|
-
|
-
|
62
|
5
|
15
|
21
|
47
|
20
|
16
|
|
51
|
90
|
5
|
7
|
23
|
46
|
18
|
8
|
|
52
|
120
|
5
|
17
|
37
|
64
|
36
|
20
|
|
53
|
70
|
5
|
7
|
21
|
35
|
19
|
2
|
|
54
|
30
|
3
|
15
|
34
|
14
|
-
|
-
|
|
55
|
100
|
5
|
4
|
31
|
64
|
35
|
6
|
|
56
|
80
|
5
|
7
|
33
|
51
|
32
|
1
|
|
57
|
22
|
3
|
5
|
32
|
5
|
-
|
-
|
|
58
|
90
|
5
|
8
|
20
|
49
|
20
|
11
|
|
59
|
42
|
3
|
9
|
33
|
7
|
-
|
-
|
|
60
|
51
|
5
|
2
|
15
|
34
|
9
|
1
|
|
61
|
35
|
3
|
17
|
35
|
17
|
-
|
-
|
|
62
|
62
|
5
|
3
|
10
|
20
|
10
|
2
|
|
63
|
33
|
3
|
10
|
35
|
9
|
-
|
-
|
|
64
|
25
|
3
|
14
|
28
|
7
|
-
|
-
|
|
65
|
21
|
3
|
6
|
26
|
5
|
-
|
-
|
|
66
|
103
|
3
|
19
|
42
|
13
|
-
|
-
|
|
67
|
86
|
5
|
6
|
31
|
52
|
19
|
4
|
|
68
|
88
|
3
|
17
|
31
|
24
|
-
|
-
|
|
69
|
96
|
3
|
29
|
40
|
25
|
-
|
-
|
|
70
|
20
|
3
|
7
|
27
|
11
|
-
|
-
|
|
71
|
25
|
3
|
10
|
21
|
10
|
-
|
-
|
|
72
|
15
|
3
|
9
|
19
|
10
|
-
|
-
|
|
73
|
81
|
3
|
20
|
41
|
19
|
-
|
-
|
|
74
|
125
|
3
|
8
|
35
|
8
|
-
|
-
|
|
75
|
20
|
3
|
10
|
26
|
9
|
-
|
-
|
|
76
|
40
|
5
|
4
|
23
|
48
|
19
|
1
|
|
77
|
20
|
3
|
9
|
20
|
10
|
-
|
-
|
|
78
|
33
|
3
|
7
|
35
|
7
|
-
|
-
|
|
79
|
31
|
5
|
3
|
14
|
27
|
12
|
2
|
|
80
|
83
|
5
|
7
|
17
|
64
|
21
|
9
|
|
81
|
41
|
3
|
10
|
23
|
13
|
-
|
-
|
|
82
|
34
|
3
|
10
|
25
|
10
|
-
|
-
|
|
83
|
35
|
3
|
10
|
20
|
9
|
-
|
-
|
|
84
|
24
|
3
|
15
|
43
|
14
|
-
|
-
|
|
85
|
30
|
3
|
15
|
30
|
11
|
-
|
-
|
|
86
|
75
|
5
|
3
|
7
|
54
|
5
|
3
|
|
87
|
25
|
3
|
20
|
25
|
16
|
-
|
-
|
|
88
|
55
|
3
|
10
|
36
|
13
|
-
|
-
|
|
89
|
17
|
3
|
10
|
35
|
11
|
-
|
-
|
|
90
|
34
|
3
|
14
|
35
|
10
|
-
|
-
|
|
91
|
92
|
5
|
9
|
15
|
52
|
21
|
5
|
|
92
|
61
|
3
|
22
|
52
|
15
|
-
|
-
|
|
93
|
44
|
5
|
1
|
10
|
30
|
22
|
|
94
|
54
|
5
|
6
|
25
|
30
|
19
|
1
|
|
95
|
18
|
3
|
9
|
21
|
11
|
-
|
-
|
|
96
|
54
|
5
|
4
|
14
|
45
|
5
|
6
|
|
97
|
45
|
5
|
7
|
19
|
50
|
21
|
3
|
|
98
|
30
|
3
|
16
|
35
|
10
|
-
|
-
|
|
99
|
65
|
5
|
2
|
6
|
50
|
7
|
2
|
|
100
|
95
|
5
|
7
|
25
|
75
|
21
|
10
|
Таблица А. 6 - Морфометрическая характеристика
вида H. helix L. subsp.
caucasigena (Апшеронский
район, пойменный лес, 3 км до с. Черниговское)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей(т), шт
|
Глубина
лопасти (l), мм
|
|
|
|
1
|
2
|
3
|
4
|
5
|
|
1
|
40
|
5
|
7
|
11
|
55
|
10
|
6
|
|
2
|
36
|
5
|
3
|
5
|
52
|
11
|
4
|
|
3
|
60
|
3
|
15
|
44
|
10
|
-
|
-
|
|
4
|
71
|
3
|
17
|
40
|
14
|
-
|
-
|
|
5
|
55
|
5
|
7
|
16
|
31
|
13
|
5
|
|
6
|
60
|
5
|
4
|
15
|
43
|
16
|
4
|
|
7
|
65
|
5
|
10
|
30
|
50
|
32
|
18
|
|
8
|
36
|
5
|
7
|
13
|
30
|
14
|
6
|
|
9
|
100
|
5
|
22
|
43
|
57
|
41
|
13
|
|
10
|
64
|
5
|
5
|
22
|
42
|
23
|
6
|
|
11
|
23
|
5
|
3
|
7
|
17
|
8
|
3
|
|
12
|
29
|
5
|
3
|
8
|
30
|
9
|
4
|
|
13
|
45
|
3
|
10
|
30
|
11
|
-
|
-
|
|
14
|
26
|
5
|
3
|
5
|
40
|
10
|
5
|
|
15
|
36
|
5
|
4
|
11
|
38
|
9
|
4
|
|
16
|
59
|
3
|
17
|
37
|
14
|
-
|
-
|
|
17
|
60
|
5
|
3
|
10
|
28
|
11
|
3
|
|
18
|
33
|
5
|
5
|
10
|
34
|
10
|
5
|
|
19
|
20
|
3
|
10
|
21
|
10
|
-
|
-
|
|
20
|
51
|
3
|
22
|
59
|
25
|
-
|
-
|
|
21
|
31
|
3
|
11
|
30
|
13
|
-
|
-
|
|
22
|
21
|
5
|
2
|
5
|
35
|
7
|
4
|
|
23
|
46
|
3
|
17
|
42
|
15
|
-
|
-
|
|
24
|
50
|
3
|
10
|
36
|
7
|
-
|
-
|
|
25
|
58
|
3
|
10
|
35
|
10
|
-
|
-
|
|
26
|
47
|
3
|
7
|
36
|
5
|
-
|
-
|
|
27
|
23
|
3
|
13
|
30
|
14
|
-
|
-
|
|
28
|
33
|
5
|
5
|
12
|
32
|
9
|
5
|
|
29
|
23
|
5
|
5
|
9
|
43
|
6
|
5
|
|
30
|
52
|
3
|
12
|
25
|
10
|
-
|
-
|
|
31
|
38
|
5
|
3
|
15
|
41
|
12
|
3
|
|
32
|
46
|
5
|
10
|
15
|
34
|
13
|
7
|
|
33
|
42
|
5
|
3
|
16
|
25
|
14
|
4
|
|
34
|
35
|
5
|
6
|
11
|
32
|
7
|
5
|
|
35
|
36
|
5
|
4
|
7
|
28
|
6
|
4
|
|
36
|
71
|
5
|
5
|
51
|
25
|
10
|
|
37
|
27
|
5
|
3
|
11
|
27
|
10
|
3
|
|
38
|
18
|
3
|
6
|
29
|
5
|
-
|
-
|
|
39
|
60
|
5
|
4
|
12
|
34
|
10
|
3
|
|
40
|
37
|
5
|
2
|
7
|
32
|
6
|
2
|
|
41
|
33
|
3
|
5
|
35
|
6
|
-
|
-
|
|
42
|
13
|
3
|
10
|
21
|
10
|
-
|
-
|
|
43
|
33
|
3
|
11
|
15
|
10
|
-
|
-
|
|
44
|
36
|
5
|
3
|
9
|
24
|
8
|
3
|
|
45
|
35
|
3
|
11
|
30
|
9
|
-
|
-
|
|
46
|
45
|
3
|
15
|
33
|
10
|
-
|
-
|
|
47
|
56
|
3
|
10
|
34
|
9
|
-
|
-
|
|
48
|
47
|
5
|
5
|
9
|
29
|
14
|
2
|
|
49
|
33
|
5
|
3
|
10
|
27
|
7
|
2
|
|
50
|
23
|
5
|
3
|
8
|
28
|
8
|
3
|
|
51
|
47
|
5
|
5
|
12
|
31
|
10
|
7
|
|
52
|
35
|
5
|
5
|
7
|
20
|
6
|
7
|
|
53
|
33
|
5
|
7
|
9
|
31
|
11
|
6
|
|
54
|
39
|
5
|
6
|
12
|
30
|
19
|
10
|
|
55
|
44
|
5
|
3
|
14
|
21
|
13
|
2
|
|
56
|
48
|
5
|
3
|
14
|
26
|
10
|
3
|
|
57
|
34
|
3
|
10
|
32
|
5
|
-
|
-
|
|
58
|
31
|
5
|
6
|
13
|
25
|
12
|
5
|
|
59
|
72
|
5
|
5
|
19
|
33
|
10
|
6
|
|
60
|
46
|
5
|
8
|
9
|
35
|
13
|
10
|
|
61
|
29
|
3
|
5
|
23
|
8
|
-
|
-
|
|
62
|
15
|
5
|
2
|
6
|
23
|
9
|
3
|
|
63
|
28
|
5
|
5
|
10
|
36
|
10
|
5
|
|
64
|
16
|
5
|
3
|
7
|
38
|
6
|
2
|
|
65
|
62
|
5
|
3
|
14
|
37
|
15
|
2
|
|
66
|
27
|
5
|
3
|
10
|
30
|
9
|
3
|
|
67
|
46
|
3
|
10
|
25
|
10
|
-
|
-
|
|
68
|
30
|
3
|
15
|
35
|
16
|
-
|
-
|
|
69
|
26
|
3
|
9
|
25
|
8
|
-
|
-
|
|
70
|
15
|
3
|
6
|
21
|
7
|
-
|
-
|
|
71
|
32
|
5
|
5
|
11
|
31
|
10
|
5
|
|
72
|
30
|
5
|
3
|
10
|
30
|
7
|
3
|
|
73
|
26
|
5
|
3
|
11
|
30
|
10
|
3
|
|
74
|
38
|
3
|
8
|
22
|
15
|
-
|
-
|
|
75
|
39
|
5
|
5
|
9
|
36
|
8
|
5
|
|
76
|
25
|
3
|
7
|
22
|
7
|
-
|
-
|
|
77
|
20
|
5
|
6
|
10
|
28
|
10
|
5
|
|
78
|
45
|
5
|
7
|
7
|
35
|
10
|
7
|
|
79
|
30
|
5
|
2
|
11
|
35
|
12
|
5
|
|
80
|
17
|
5
|
7
|
21
|
6
|
5
|
|
81
|
17
|
3
|
5
|
29
|
5
|
-
|
-
|
|
82
|
35
|
5
|
5
|
11
|
25
|
10
|
5
|
|
83
|
22
|
5
|
3
|
6
|
27
|
7
|
3
|
|
84
|
30
|
3
|
6
|
28
|
7
|
-
|
-
|
|
85
|
28
|
3
|
5
|
24
|
3
|
-
|
-
|
|
86
|
15
|
5
|
4
|
7
|
22
|
7
|
5
|
|
87
|
30
|
5
|
5
|
10
|
30
|
5
|
5
|
|
88
|
25
|
3
|
7
|
27
|
7
|
-
|
-
|
|
89
|
20
|
3
|
5
|
20
|
9
|
-
|
-
|
|
90
|
32
|
3
|
8
|
25
|
4
|
-
|
-
|
|
91
|
22
|
5
|
3
|
6
|
20
|
5
|
4
|
|
92
|
41
|
5
|
4
|
6
|
21
|
7
|
5
|
|
93
|
20
|
3
|
7
|
22
|
5
|
-
|
-
|
|
94
|
32
|
3
|
11
|
37
|
14
|
-
|
-
|
|
95
|
38
|
5
|
3
|
8
|
29
|
7
|
3
|
|
96
|
15
|
3
|
9
|
27
|
5
|
-
|
-
|
|
97
|
34
|
5
|
3
|
7
|
25
|
5
|
3
|
|
98
|
27
|
5
|
3
|
11
|
28
|
4
|
2
|
|
99
|
25
|
5
|
3
|
9
|
21
|
6
|
3
|
|
100
|
23
|
5
|
5
|
8
|
28
|
4
|
7
|
Таблица А. 7 - Морфометрическая характеристика
вида H. helix L. (Испания,
провинция Каталония, берег Средиземного моря)
|
№
|
Количество
лопастей (n), шт
|
Глубина
лопасти (l), мм
|
|
|
1
|
2
|
3
|
4
|
5
|
|
1
|
5
|
15
|
30
|
60
|
20
|
10
|
|
2
|
5
|
10
|
18
|
50
|
20
|
5
|
|
3
|
5
|
15
|
18
|
50
|
20
|
15
|
|
4
|
3
|
18
|
-
|
73
|
-
|
15
|
|
5
|
3
|
19
|
-
|
40
|
-
|
12
|
|
6
|
3
|
11
|
-
|
48
|
-
|
20
|
|
7
|
3
|
14
|
-
|
37
|
-
|
15
|
|
8
|
3
|
20
|
-
|
70
|
-
|
20
|
|
9
|
3
|
25
|
-
|
40
|
-
|
20
|
|
10
|
5
|
10
|
30
|
45
|
15
|
10
|
|
11
|
3
|
10
|
-
|
43
|
-
|
10
|
|
12
|
3
|
15
|
-
|
65
|
--
|
22
|
|
13
|
5
|
5
|
7
|
48
|
20
|
7
|
|
14
|
3
|
18
|
-
|
51
|
-
|
21
|
|
15
|
3
|
18
|
-
|
47
|
-
|
14
|
|
16
|
3
|
18
|
-
|
38
|
-
|
19
|
|
17
|
3
|
6
|
-
|
45
|
-
|
19
|
|
18
|
5
|
2
|
30
|
54
|
28
|
7
|
|
19
|
5
|
8
|
33
|
60
|
30
|
14
|
|
20
|
5
|
2
|
12
|
45
|
14
|
2
|
|
21
|
3
|
30
|
-
|
100
|
-
|
33
|
|
22
|
5
|
15
|
48
|
80
|
47
|
15
|
|
23
|
3
|
20
|
-
|
42
|
-
|
14
|
|
24
|
3
|
33
|
-
|
62
|
-
|
32
|
|
25
|
5
|
11
|
20
|
50
|
28
|
5
|
|
26
|
3
|
-
|
48
|
-
|
16
|
|
27
|
5
|
4
|
32
|
42
|
27
|
6
|
|
28
|
3
|
12
|
-
|
51
|
-
|
19
|
|
29
|
3
|
18
|
-
|
45
|
-
|
25
|
|
30
|
3
|
16
|
-
|
41
|
-
|
15
|
|
31
|
5
|
5
|
21
|
65
|
29
|
2
|
|
32
|
5
|
2
|
9
|
35
|
11
|
2
|
|
33
|
3
|
25
|
-
|
56
|
-
|
29
|
|
34
|
3
|
18
|
-
|
40
|
-
|
14
|
|
35
|
3
|
7
|
-
|
46
|
-
|
6
|
|
36
|
5
|
2
|
25
|
52
|
18
|
2
|
|
37
|
5
|
2
|
16
|
39
|
19
|
3
|
|
38
|
3
|
21
|
-
|
61
|
-
|
23
|
|
39
|
5
|
5
|
19
|
38
|
19
|
8
|
|
40
|
3
|
26
|
-
|
58
|
-
|
23
|
|
41
|
3
|
12
|
-
|
41
|
-
|
16
|
|
42
|
5
|
19
|
25
|
60
|
29
|
16
|
|
43
|
5
|
3
|
22
|
51
|
25
|
10
|
|
44
|
5
|
19
|
38
|
59
|
35
|
15
|
|
45
|
3
|
15
|
-
|
65
|
-
|
14
|
|
46
|
3
|
18
|
-
|
50
|
-
|
15
|
|
47
|
3
|
27
|
-
|
51
|
-
|
21
|
|
48
|
5
|
5
|
35
|
56
|
31
|
5
|
|
49
|
5
|
11
|
34
|
70
|
36
|
11
|
|
50
|
5
|
10
|
27
|
35
|
27
|
13
|
|
51
|
5
|
10
|
30
|
65
|
29
|
11
|
|
52
|
5
|
4
|
25
|
32
|
29
|
7
|
|
53
|
3
|
9
|
-
|
45
|
-
|
9
|
|
54
|
3
|
39
|
-
|
75
|
-
|
45
|
|
55
|
3
|
31
|
-
|
52
|
-
|
26
|
|
56
|
3
|
21
|
-
|
45
|
-
|
21
|
|
57
|
3
|
25
|
-
|
69
|
-
|
26
|
|
58
|
5
|
7
|
41
|
50
|
34
|
14
|
|
59
|
5
|
9
|
20
|
36
|
30
|
10
|
|
60
|
5
|
7
|
35
|
47
|
30
|
10
|
|
61
|
3
|
30
|
-
|
70
|
-
|
30
|
|
62
|
5
|
8
|
38
|
65
|
45
|
7
|
|
63
|
5
|
21
|
44
|
61
|
40
|
10
|
|
64
|
5
|
5
|
25
|
56
|
23
|
4
|
|
65
|
3
|
18
|
-
|
44
|
-
|
13
|
|
66
|
3
|
11
|
-
|
45
|
-
|
18
|
|
67
|
5
|
10
|
25
|
34
|
14
|
4
|
|
68
|
3
|
20
|
-
|
50
|
-
|
20
|
|
69
|
3
|
18
|
-
|
51
|
-
|
21
|
|
70
|
3
|
25
|
-
|
61
|
-
|
17
|
|
71
|
5
|
5
|
35
|
85
|
40
|
5
|
|
72
|
5
|
9
|
40
|
71
|
38
|
10
|
|
73
|
3
|
13
|
-
|
52
|
-
|
13
|
|
74
|
3
|
16
|
-
|
55
|
-
|
30
|
|
75
|
5
|
7
|
28
|
68
|
38
|
5
|
|
76
|
3
|
15
|
35
|
-
|
14
|
|
77
|
3
|
26
|
-
|
48
|
-
|
35
|
|
78
|
5
|
12
|
33
|
61
|
40
|
14
|
|
79
|
5
|
5
|
21
|
40
|
19
|
5
|
|
80
|
5
|
4
|
17
|
48
|
19
|
7
|
|
81
|
5
|
5
|
23
|
39
|
21
|
7
|
|
82
|
5
|
14
|
25
|
56
|
35
|
14
|
|
83
|
3
|
15
|
-
|
36
|
-
|
14
|
|
84
|
5
|
5
|
21
|
35
|
25
|
4
|
|
85
|
3
|
15
|
-
|
36
|
-
|
12
|
|
86
|
5
|
14
|
39
|
59
|
40
|
5
|
|
87
|
3
|
28
|
-
|
82
|
-
|
30
|
|
88
|
3
|
39
|
-
|
75
|
-
|
45
|
|
89
|
3
|
31
|
-
|
52
|
-
|
26
|
|
90
|
3
|
21
|
-
|
45
|
-
|
21
|
|
91
|
3
|
25
|
-
|
69
|
-
|
26
|
|
92
|
5
|
7
|
41
|
50
|
34
|
14
|
|
93
|
5
|
9
|
20
|
36
|
30
|
10
|
|
94
|
5
|
7
|
35
|
47
|
30
|
10
|
|
95
|
3
|
30
|
-
|
70
|
-
|
30
|
|
96
|
5
|
8
|
38
|
65
|
45
|
7
|
|
97
|
5
|
21
|
44
|
61
|
40
|
10
|
|
98
|
5
|
15
|
30
|
60
|
20
|
10
|
|
99
|
5
|
10
|
18
|
50
|
20
|
5
|
|
100
|
5
|
11
|
21
|
53
|
22
|
7
|
|
|
|
|
|
|
|
|
Таблица А. 8 - Морфометрическая характеристика
вида H. colhica (Туапсинсий район, верховье р. Цыпка, дубово-буковый лес)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Лопасть
|
|
|
|
|
|
|
|
ширина
(а),мм
|
длина
(b),мм
|
|
1
|
112
|
1
|
71
|
93
|
|
2
|
115
|
1
|
132
|
125
|
|
3
|
95
|
1
|
59
|
86
|
|
4
|
78
|
1
|
55
|
73
|
|
5
|
50
|
1
|
54
|
82
|
|
6
|
52
|
1
|
55
|
81
|
|
7
|
72
|
1
|
71
|
110
|
|
8
|
110
|
1
|
67
|
107
|
|
9
|
90
|
1
|
95
|
115
|
|
10
|
105
|
1
|
92
|
100
|
|
11
|
130
|
1
|
110
|
100
|
|
12
|
120
|
1
|
92
|
110
|
|
13
|
88
|
1
|
110
|
160
|
|
14
|
115
|
1
|
125
|
105
|
|
15
|
40
|
1
|
44
|
70
|
|
16
|
35
|
1
|
37
|
65
|
|
17
|
61
|
1
|
80
|
90
|
|
18
|
100
|
1
|
100
|
120
|
|
19
|
82
|
1
|
98
|
105
|
|
20
|
105
|
1
|
76
|
100
|
|
21
|
25
|
1
|
44
|
60
|
|
22
|
90
|
1
|
100
|
110
|
|
23
|
45
|
1
|
60
|
80
|
|
24
|
95
|
1
|
66
|
105
|
|
25
|
75
|
1
|
67
|
85
|
|
26
|
80
|
1
|
75
|
96
|
|
27
|
62
|
1
|
49
|
72
|
|
28
|
45
|
1
|
60
|
92
|
|
29
|
50
|
1
|
60
|
65
|
|
30
|
80
|
1
|
70
|
100
|
|
31
|
71
|
1
|
65
|
98
|
|
32
|
45
|
1
|
67
|
100
|
|
33
|
55
|
1
|
62
|
73
|
|
34
|
50
|
1
|
55
|
60
|
|
35
|
80
|
64
|
110
|
|
36
|
65
|
1
|
50
|
85
|
|
37
|
45
|
1
|
66
|
70
|
|
38
|
85
|
1
|
70
|
90
|
|
39
|
43
|
1
|
52
|
101
|
|
40
|
89
|
1
|
63
|
85
|
|
41
|
65
|
1
|
70
|
110
|
|
42
|
66
|
1
|
54
|
81
|
|
43
|
79
|
1
|
64
|
91
|
|
44
|
41
|
1
|
44
|
80
|
|
45
|
23
|
1
|
40
|
74
|
|
46
|
61
|
1
|
66
|
86
|
|
47
|
62
|
1
|
65
|
70
|
|
48
|
76
|
1
|
69
|
105
|
|
49
|
46
|
1
|
70
|
105
|
|
50
|
46
|
1
|
60
|
110
|
|
51
|
104
|
1
|
66
|
114
|
|
52
|
86
|
1
|
71
|
85
|
|
53
|
111
|
1
|
81
|
91
|
|
54
|
110
|
1
|
96
|
100
|
|
55
|
95
|
1
|
105
|
120
|
|
56
|
112
|
1
|
116
|
113
|
|
57
|
37
|
1
|
57
|
91
|
|
58
|
70
|
1
|
59
|
115
|
|
59
|
60
|
1
|
62
|
100
|
|
60
|
75
|
1
|
71
|
97
|
|
61
|
70
|
1
|
76
|
90
|
|
62
|
54
|
1
|
62
|
67
|
|
63
|
55
|
1
|
64
|
102
|
|
64
|
102
|
1
|
104
|
117
|
|
65
|
92
|
1
|
92
|
110
|
|
66
|
87
|
1
|
62
|
90
|
|
67
|
76
|
1
|
70
|
111
|
|
68
|
71
|
1
|
49
|
56
|
|
69
|
52
|
1
|
59
|
83
|
|
70
|
100
|
1
|
114
|
95
|
|
71
|
114
|
1
|
66
|
77
|
|
72
|
73
|
1
|
72
|
90
|
|
73
|
55
|
1
|
58
|
105
|
|
74
|
82
|
1
|
64
|
80
|
|
75
|
115
|
1
|
95
|
122
|
|
76
|
68
|
1
|
50
|
85
|
|
77
|
66
|
1
|
74
|
80
|
|
78
|
96
|
1
|
71
|
86
|
|
79
|
77
|
1
|
71
|
80
|
|
80
|
43
|
1
|
44
|
59
|
|
81
|
55
|
1
|
76
|
77
|
|
82
|
50
|
1
|
63
|
86
|
|
83
|
67
|
1
|
73
|
102
|
|
84
|
75
|
1
|
79
|
115
|
|
85
|
97
|
1
|
82
|
101
|
|
86
|
50
|
1
|
58
|
79
|
|
87
|
39
|
1
|
40
|
67
|
|
88
|
113
|
1
|
80
|
133
|
|
89
|
115
|
1
|
70
|
77
|
|
90
|
97
|
1
|
60
|
85
|
|
91
|
51
|
1
|
39
|
63
|
|
92
|
67
|
1
|
67
|
75
|
|
93
|
70
|
1
|
47
|
80
|
|
94
|
45
|
1
|
44
|
84
|
|
95
|
59
|
1
|
52
|
82
|
|
96
|
112
|
1
|
60
|
85
|
|
97
|
31
|
1
|
48
|
79
|
|
98
|
75
|
1
|
67
|
81
|
|
99
|
55
|
1
|
53
|
66
|
|
100
|
63
|
1
|
54
|
56
|
Таблица А. 9 - Морфометрическая характеристика
вида H. colhica (Апшеронский район, буковый лес, вход в Гуамское ущелье)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Глубина
лопасти (l), мм
|
|
|
|
1
|
2
|
3
|
4
|
5
|
|
1
|
100
|
3
|
10
|
40
|
6
|
-
|
-
|
|
2
|
67
|
3
|
9
|
47
|
-
|
-
|
|
3
|
57
|
3
|
3
|
52
|
1
|
-
|
-
|
|
4
|
70
|
3
|
1
|
45
|
2
|
-
|
-
|
|
5
|
65
|
3
|
7
|
46
|
10
|
-
|
-
|
|
6
|
52
|
3
|
3
|
25
|
4
|
-
|
-
|
|
7
|
50
|
3
|
4
|
30
|
3
|
-
|
-
|
|
8
|
95
|
3
|
8
|
25
|
7
|
-
|
-
|
|
9
|
60
|
3
|
14
|
28
|
8
|
-
|
-
|
|
10
|
60
|
3
|
8
|
43
|
10
|
-
|
-
|
|
11
|
64
|
3
|
10
|
52
|
8
|
-
|
-
|
|
12
|
60
|
3
|
10
|
33
|
5
|
-
|
-
|
|
13
|
46
|
3
|
3
|
37
|
2
|
-
|
-
|
|
14
|
57
|
3
|
2
|
43
|
1
|
-
|
-
|
|
15
|
42
|
3
|
3
|
30
|
1
|
-
|
-
|
|
16
|
63
|
3
|
1
|
40
|
1
|
-
|
-
|
|
17
|
48
|
3
|
5
|
38
|
3
|
-
|
-
|
|
18
|
67
|
3
|
14
|
45
|
15
|
-
|
-
|
|
19
|
55
|
3
|
2
|
45
|
1
|
-
|
-
|
|
20
|
30
|
3
|
4
|
53
|
7
|
-
|
-
|
|
21
|
48
|
3
|
9
|
36
|
9
|
-
|
-
|
|
22
|
92
|
3
|
9
|
40
|
3
|
-
|
-
|
|
23
|
61
|
3
|
2
|
52
|
3
|
-
|
-
|
|
24
|
47
|
3
|
3
|
35
|
5
|
-
|
-
|
|
25
|
34
|
3
|
4
|
25
|
1
|
-
|
-
|
|
26
|
35
|
3
|
2
|
23
|
3
|
-
|
-
|
|
27
|
50
|
3
|
5
|
36
|
16
|
-
|
-
|
|
28
|
77
|
3
|
5
|
42
|
3
|
-
|
-
|
|
29
|
51
|
3
|
9
|
39
|
5
|
-
|
-
|
|
30
|
73
|
3
|
6
|
36
|
7
|
-
|
-
|
|
31
|
70
|
3
|
7
|
31
|
7
|
-
|
-
|
|
32
|
50
|
3
|
5
|
40
|
6
|
-
|
-
|
|
33
|
40
|
3
|
3
|
31
|
4
|
-
|
-
|
|
34
|
67
|
3
|
3
|
34
|
2
|
-
|
-
|
|
35
|
62
|
3
|
4
|
42
|
10
|
-
|
-
|
|
36
|
70
|
3
|
10
|
45
|
8
|
-
|
-
|
|
37
|
42
|
3
|
5
|
23
|
2
|
-
|
-
|
|
38
|
63
|
5
|
7
|
3
|
30
|
2
|
8
|
|
39
|
61
|
3
|
4
|
55
|
5
|
-
|
-
|
|
40
|
70
|
3
|
7
|
54
|
7
|
-
|
-
|
|
41
|
70
|
3
|
11
|
43
|
8
|
-
|
-
|
|
42
|
58
|
3
|
5
|
47
|
3
|
-
|
-
|
|
43
|
65
|
3
|
4
|
41
|
6
|
-
|
-
|
|
44
|
65
|
3
|
7
|
27
|
5
|
-
|
-
|
|
45
|
85
|
3
|
6
|
28
|
5
|
-
|
-
|
|
46
|
50
|
3
|
7
|
8
|
-
|
-
|
|
47
|
90
|
3
|
13
|
64
|
9
|
-
|
-
|
|
48
|
69
|
3
|
3
|
44
|
3
|
-
|
-
|
|
49
|
68
|
3
|
8
|
40
|
4
|
-
|
-
|
|
50
|
61
|
3
|
4
|
51
|
3
|
-
|
-
|
|
51
|
48
|
3
|
5
|
25
|
3
|
-
|
-
|
|
52
|
75
|
3
|
9
|
50
|
7
|
-
|
-
|
|
53
|
58
|
3
|
6
|
29
|
4
|
-
|
-
|
|
54
|
53
|
3
|
4
|
39
|
8
|
-
|
-
|
|
55
|
91
|
3
|
13
|
52
|
8
|
-
|
-
|
|
56
|
79
|
3
|
9
|
24
|
4
|
-
|
-
|
|
57
|
38
|
3
|
2
|
33
|
5
|
-
|
-
|
|
58
|
97
|
3
|
11
|
41
|
7
|
-
|
-
|
|
59
|
45
|
3
|
5
|
32
|
5
|
-
|
-
|
|
60
|
64
|
3
|
4
|
36
|
3
|
-
|
-
|
|
61
|
58
|
3
|
5
|
30
|
6
|
-
|
-
|
|
62
|
79
|
3
|
7
|
43
|
6
|
-
|
-
|
|
63
|
58
|
3
|
3
|
24
|
3
|
-
|
-
|
|
64
|
47
|
3
|
9
|
26
|
8
|
-
|
-
|
|
65
|
71
|
3
|
2
|
40
|
6
|
-
|
-
|
|
66
|
27
|
3
|
5
|
28
|
9
|
-
|
-
|
|
67
|
100
|
3
|
11
|
60
|
10
|
-
|
-
|
|
69
|
26
|
3
|
4
|
24
|
3
|
-
|
-
|
|
70
|
20
|
3
|
4
|
26
|
5
|
-
|
-
|
|
71
|
67
|
3
|
5
|
42
|
5
|
-
|
-
|
|
72
|
60
|
3
|
4
|
51
|
9
|
-
|
-
|
|
73
|
69
|
3
|
8
|
39
|
5
|
-
|
-
|
|
74
|
60
|
3
|
4
|
35
|
3
|
-
|
-
|
|
75
|
63
|
3
|
8
|
50
|
6
|
-
|
-
|
|
76
|
35
|
3
|
6
|
28
|
5
|
-
|
-
|
|
77
|
62
|
3
|
4
|
41
|
3
|
-
|
-
|
|
78
|
52
|
3
|
5
|
27
|
7
|
-
|
-
|
|
79
|
101
|
3
|
6
|
45
|
5
|
-
|
-
|
|
80
|
58
|
3
|
7
|
54
|
5
|
-
|
-
|
Таблица А. 10 - Морфометрическая характеристика
вида H. colchica
(Сочинский район, п. Совет Кваджи, склон у дороги)
|
№
п/п
|
Длина
черешка (L), мм
|
Количество
лопастей (n), шт
|
Лопасть
|
|
|
|
ширина
(а), мм
|
длина
(b), мм
|
|
|
|
|
|
|
1
|
1
|
1
|
34
|
53
|
|
2
|
13
|
1
|
52
|
60
|
|
3
|
15
|
1
|
37
|
39
|
|
4
|
7
|
1
|
30
|
34
|
|
5
|
95
|
1
|
102
|
111
|
|
6
|
47
|
1
|
64
|
82
|
|
7
|
15
|
1
|
55
|
65
|
|
8
|
47
|
1
|
60
|
74
|
|
9
|
10
|
1
|
40
|
67
|
|
10
|
13
|
1
|
43
|
50
|
|
11
|
105
|
1
|
123
|
130
|
|
12
|
12
|
1
|
42
|
45
|
|
13
|
17
|
1
|
50
|
70
|
|
14
|
40
|
1
|
50
|
70
|
|
15
|
14
|
1
|
46
|
60
|
|
16
|
1
|
36
|
40
|
|
17
|
10
|
1
|
40
|
57
|
|
18
|
11
|
1
|
29
|
48
|
|
19
|
18
|
1
|
39
|
50
|
|
20
|
13
|
1
|
36
|
56
|
|
21
|
14
|
1
|
49
|
46
|
|
22
|
11
|
1
|
31
|
54
|
|
23
|
81
|
1
|
89
|
43
|
|
24
|
56
|
1
|
118
|
109
|
|
25
|
75
|
1
|
109
|
144
|
|
26
|
65
|
1
|
100
|
110
|
|
27
|
93
|
1
|
86
|
124
|
|
28
|
91
|
1
|
101
|
95
|
|
29
|
34
|
1
|
53
|
119
|
|
30
|
18
|
1
|
52
|
72
|
|
31
|
34
|
1
|
79
|
59
|
|
32
|
58
|
1
|
55
|
96
|
|
33
|
12
|
1
|
45
|
68
|
|
34
|
51
|
1
|
52
|
53
|
|
35
|
50
|
1
|
70
|
69
|
|
36
|
53
|
1
|
73
|
60
|
|
37
|
96
|
1
|
90
|
95
|
|
38
|
72
|
1
|
96
|
99
|
|
39
|
34
|
1
|
61
|
89
|
|
40
|
63
|
1
|
70
|
80
|
|
41
|
40
|
1
|
56
|
95
|
|
42
|
38
|
1
|
62
|
61
|
|
43
|
34
|
1
|
49
|
76
|
|
44
|
99
|
1
|
109
|
57
|
|
45
|
44
|
1
|
87
|
119
|
|
46
|
72
|
1
|
92
|
111
|
|
47
|
72
|
1
|
127
|
102
|
|
48
|
80
|
1
|
85
|
127
|
|
49
|
69
|
1
|
78
|
95
|
|
50
|
95
|
1
|
99
|
88
|
|
51
|
79
|
1
|
105
|
87
|
|
52
|
93
|
1
|
80
|
108
|
|
53
|
85
|
1
|
139
|
108
|
|
54
|
65
|
1
|
84
|
144
|
|
55
|
84
|
1
|
122
|
114
|
|
56
|
72
|
1
|
98
|
107
|
|
57
|
44
|
1
|
77
|
100
|
|
58
|
34
|
1
|
69
|
88
|
|
59
|
45
|
1
|
84
|
78
|
|
60
|
37
|
1
|
68
|
106
|
|
61
|
40
|
1
|
66
|
92
|
|
62
|
38
|
1
|
76
|
90
|
|
63
|
80
|
1
|
80
|
96
|
|
64
|
30
|
1
|
74
|
97
|
|
65
|
48
|
1
|
87
|
89
|
|
66
|
65
|
1
|
100
|
110
|
|
67
|
93
|
1
|
86
|
124
|
|
68
|
91
|
1
|
101
|
95
|
|
69
|
34
|
1
|
53
|
119
|
|
70
|
18
|
1
|
52
|
72
|
|
71
|
34
|
1
|
79
|
59
|
|
72
|
58
|
1
|
55
|
96
|
|
73
|
12
|
1
|
45
|
68
|
|
74
|
51
|
1
|
52
|
53
|
|
75
|
50
|
1
|
70
|
69
|
|
76
|
53
|
1
|
73
|
60
|
|
77
|
96
|
1
|
90
|
95
|
|
78
|
72
|
1
|
96
|
99
|
|
79
|
34
|
1
|
61
|
89
|
|
80
|
63
|
1
|
70
|
80
|
|
81
|
40
|
1
|
56
|
95
|
|
83
|
34
|
1
|
49
|
76
|
|
84
|
99
|
1
|
109
|
57
|
|
85
|
44
|
1
|
87
|
119
|
|
86
|
72
|
1
|
92
|
111
|
|
87
|
72
|
127
|
102
|
|
88
|
80
|
1
|
85
|
127
|
|
89
|
69
|
1
|
78
|
95
|
|
90
|
95
|
1
|
99
|
88
|
|
91
|
79
|
1
|
105
|
87
|
|
92
|
93
|
1
|
80
|
108
|
|
93
|
85
|
1
|
139
|
108
|
|
94
|
65
|
1
|
84
|
144
|
|
95
|
84
|
1
|
122
|
114
|
|
96
|
72
|
1
|
98
|
107
|
|
97
|
44
|
1
|
77
|
100
|
|
98
|
34
|
1
|
69
|
88
|
|
99
|
45
|
1
|
84
|
78
|
|
100
|
37
|
1
|
68
|
106
|