Общая характеристика анализаторов
Министерство образования и науки РФ
ФГБОУ ВПО
Нижнетагильская государственная
социально-педагогическая академия
Институт психолого-педагогического
образования
Кафедра безопасности
жизнедеятельности
Контрольная работа
по дисциплине:
Физиология высшей нервной
деятельности и сенсорных систем
Тема:
Общая характеристика анализаторов
Выполнила: Калганова А.В.,
студентка 422 группы
Проверила: доцент,
к.б.н. Мишина Е.Г.
Нижний Тагил - 2015
Задание:
. Используя рекомендуемую учебную литературу изучить и составить
конспект:
Что такое сенсорная система, каково её строение и функция?
Суть кодирования информации в сенсорных системах
Заполнить таблицу
Таблица
Общий план строения сенсорных систем
|
Название анализатора
|
Природа раздражителя
|
Периферический отдел
|
Проводниковый отдел
|
Центральный отел
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2. Рассмотреть строение и принцип работы сенсорной системы - зрительный
анализатор. Составить опорную схему проводящих путей анализатора
1. Строение и функции сенсорной
системы
Представление о сенсорных системах было сформулировано
И.П. Павловым в учении об анализаторах в 1909 г. при исследовании им высшей
нервной деятельности. Анализатор - совокупность центральных и
периферических образований, воспринимающих и анализирующих изменения внешней и
внутренней сред организма. Понятие сенсорная система, появившееся позже,
заменило понятие анализатор, включив механизмы регуляции различных его отделов
с помощью прямых и обратных связей. Наряду с этим по-прежнему бытует понятие орган
чувств как периферическое образование, воспринимающее и частично
анализирующего факторы окружающей среды. Главной частью органа чувств являются
рецепторы, снабженные вспомогательными структурами, обеспечивающими оптимальное
восприятие. Так, орган зрения состоит из глазного яблока, сетчатой оболочки, в
составе которой имеются зрительные рецепторы, и ряда вспомогательных структур:
век, мышц, слезного аппарата. Орган слуха состоит из наружного, среднего и
внутреннего уха, где кроме спирального (кортиева) органа и его волосковых (рецепторных)
клеток имеется также ряд вспомогательных структур. Органом вкуса можно считать
язык. При непосредственном воздействии различных факторов окружающей среды с
участием анализаторов в организме возникают ощущения, которые
представляют собой отражения свойств предметов объективного мира. Особенностью
ощущений является их модальность, т.е. совокупность ощущений,
обеспечиваемых каким-либо одним анализатором. Внутри каждой модальности в
соответствии с видом (качеством) сенсорного впечатления можно выделить разные
качества, или валентности. Модальностями являются, например, зрение,
слух, вкус. Качественные типы модальности (валентности) для зрения - это
различные цвета, для вкуса - ощущение кислого, сладкого, соленого, горького.
Деятельность анализаторов обычно связывают с
возникновением пяти чувств - зрения, слуха, вкуса, обоняния и осязания, с
помощью которых осуществляется связь организма с внешней средой. Однако в
реальной действительности их значительно больше. Например, чувство осязания в
широком понимании кроме тактильных ощущений, возникающих от прикосновения,
включает чувство давления и вибрации. Температурное чувство включает ощущения
тепла или холода, но существуют также и более сложные ощущения, такие как
ощущения голода, жажды, половой потребности (либидо), обусловленные особым
(мотивационным) состоянием организма. Ощущение положения тела в пространстве
связано с деятельностью вестибулярного, двигательного анализаторов и их
взаимодействием со зрительным анализатором. Особое место в сенсорной функции
занимает ощущение боли. Кроме того, мы можем, хотя и «смутно», воспринимать и
другие изменения, причем не только внешней, но и внутренней сред организма, при
этом формируются эмоционально окрашенные ощущения. Так, коронароспазм в
начальной стадии заболевания, когда еще не возникают болевые ощущения, может
вызвать чувство тоски, уныния. Таким образом, структур, воспринимающих
раздражение из среды обитания и внутренней среды организма, в действительности
значительно больше, чем принято считать.
В основу классификации анализаторов могут быть
положены различные признаки: природа действующего раздражителя, характер
возникающих ощущений, уровень чувствительности рецепторов, скорость адаптации и
многое другое.
Но наиболее существенной является классификация анализаторов,
в основе которой лежит их назначение (роль). В связи с этим выделяют несколько
видов анализаторов.
Внешние анализаторы воспринимают и анализируют изменения
внешней среды. Сюда следует включить зрительный, слуховой, обонятельный,
вкусовой, тактильный и температурный анализаторы, возбуждение которых
воспринимается субъективно в виде ощущений.
Внутренние (висцеральные)
анализаторы, воспринимающие
и анализирующие изменения внутренней среды организма, показателей гомеостазиса.
Колебания показателей внутренней среды в пределах физиологической нормы у
здорового человека обычно не воспринимается субъективно в виде ощущений. Так,
мы не можем субъективно определить величину артериального давления, особенно
если оно нормальное, состояние сфинктеров и пр. Однако информация, идущая из
внутренней среды, играет важную роль в регуляции функций внутренних органов,
обеспечивая приспособление организма к различным условиям его
жизнедеятельности. Значение этих анализаторов изучается в рамках курса
физиологии (приспособительная регуляция деятельности внутренних органов). Но в
то же время изменение некоторых констант внутренней среды организма может
восприниматься субъективно в виде ощущений (жажда, голод, половое влечение),
формирующихся на основе биологических потребностей. Для удовлетворения этих
потребностей включаются поведенческие реакции. Например, при возникновении
чувства жажды вследствие возбуждения осмо- или волюморецепторов формируется
поведение, направленное на поиск и прием воды.
Анализаторы положения тела воспринимают и анализируют изменения
положения тела в пространстве и частей тела друг относительно друга. К ним
следует отнести вестибулярный и двигательный (кинестетический) анализаторы.
Поскольку мы оцениваем положение нашего тела или его частей друг относительно
друга, эта импульсация доходит до нашего сознания. Об этом свидетельствует, в
частности, опыт Д. Маклоски, который он поставил на самом себе. Первичные
афферентные волокна от мышечных рецепторов раздражались пороговыми
электрическими стимулами. Увеличение частоты импульсации этих нервных волокон
вызывало у испытуемого субъективные ощущения изменения положения
соответствующей конечности, хотя ее положение в действительности не изменялось.
Болевой анализатор отдельно следует выделить в связи с
его особым значением для организма - он несет информацию о повреждающих
действиях. Болевые ощущения могут возникать при раздражении как экстеро-, так и
интерорецепторов.
Структурно-функциональная
организация анализаторов
Согласно
представлению И.П. Павлова (1909), любой анализатор имеет три отдела:
периферический, проводниковый и центральный, или корковый. Периферический отдел
анализатора представлен рецепторами. Его назначение - восприятие и
первичный анализ изменений внешней и внутренней сред организма. В рецепторах
происходит трансформация энергии раздражителя в нервный импульс, а также
усиление сигнала за счет внутренней энергии метаболических процессов. Для
рецепторов характерна специфичность (модальность), т.е. способность
воспринимать определенный вид раздражителя, к которому они приспособились в
процессе эволюции (адекватные раздражители), на чем основан первичный анализ.
Так, рецепторы зрительного анализатора приспособлены к восприятию света, а
слуховые рецепторы - звука и т.д. Та часть рцепторной поверхности, от которой
сигнал получает одно афферентное волокно, называется его рецептивным полем.
Рецептивные поля могут иметь различное количество рецепторных образований (от 2
до 30 и более), среди которых есть рецептор-лидер, и перекрывать друг друга. Последнее
обеспечивает большую надежность выполнения функции и играет существенную роль в
механизмах компенсации.
Рецепторы характеризуются большим разнообразием.
В классификации рецепторов центральное место занимает
их деление в зависимости от вида воспринимаемого раздражителя. Существует пять
типов таких рецепторов.
. Механорецепторы возбуждаются при их механической
деформации, расположены в коже, сосудах, внутренних органах,
опорно-двигательном аппарате, слуховой и вестибулярной системах.
. Хеморецепторы воспринимают химические изменения
внешней и внутренней среды организма. К ним относятся вкусовые и обонятельные
рецепторы, а также рецепторы, реагирующие на изменение состава крови, лимфы,
межклеточной и цереброспинальной жидкости (изменение напряжения О2 и
СО2, осмолярности и рН, уровня глюкозы и других веществ). Такие
рецепторы есть в слизистой оболочке языка и носа, каротидном и аортальном
тельцах, гипоталамусе и продолговатом мозге.
. Терморецепторы воспринимают изменения температуры.
Они подразделяются на тепловые и холодовые рецепторы и находятся в коже,
слизистых оболочках, сосудах, внутренних органах, гипоталамусе, среднем,
продолговатом и спинном мозге.
. Фоторецепторы в сетчатке глаза воспринимают световую
(электромагнитную) энергию.
. Ноцицепторы, возбуждение которых сопровождается
болевыми ощущениями (болевые рецепторы). Раздражителями этих рецепторов
являются механические, термические и химические (гистамин, брадикинин, К+,
Н+ и др.) факторы. Болевые стимулы воспринимаются свободными нервными окончаниями,
которые имеются в коже, мышцах, внутренних органах, дентине, сосудах.
С психофизиологической точки зрения рецепторы подразделяют в соответствии
с органами чувств и формируемыми ощущениями на зрительные, слуховые, вкусовые,
обонятельные и тактильные.
По расположению в организме рецепторы делят на экстеро- и
интерорецепторы.
К экстерорецепторам относятся рецепторы кожи, видимых
слизистых оболочек и органов чувств: зрительные, слуховые, вкусовые,
обонятельные, тактильные, болевые и температурные. К ин-терорецепторам
относятся рецепторы внутренних органов (висцерорецепторы), сосудов и ЦНС.
Разновидностью интерорецепторов являются рецепторы опорно-двигательного
аппарата (проприорецепторы) и вестибулярные рецепторы. Если одна и та же
разновидность рецепторов (например, хеморецепторы, чувствительные к СО3)
локализована как в ЦНС (в продолговатом мозге), так и в других местах
(сосудах), то такие рецепторы подразделяют на центральные и периферические.
По скорости адаптации рецепторы делят на три группы: быстро
адаптирующиеся (фазные), медленно адаптирующиеся (тонические) и смешанные
(фазнотонические), адаптирующиеся со средней скоростью. Примером быстро
адаптирующихся рецепторов являются рецепторы вибрации (тельца Пачини) и
прикосновения (тельца Мейснера) к коже. К медленно адаптирующимся рецепторам
относятся проприорецепторы, рецепторы растяжения легких, болевые рецепторы. Со
средней скоростью адаптируются фоторецепторы сетчатки, терморецепторы кожи.
По структурно-функциональной
организации различают
первичные и вторичные рецепторы. Первичные рецепторы представляют собой
чувствительные окончания дендрита афферентного нейрона. Тело нейрона
расположено в спинно-мозговом ганглии или в ганглии черепных нервов. В
первичном рецепторе раздражитель действует непосредственно на окончания
сенсорного нейрона. Первичные рецепторы являются филогенетически более древними
структурами, к ним относятся обонятельные, тактильные, температурные, болевые
рецепторы и проприорецепторы.
Во вторичных рецепторах имеется специальная клетка,
синаптически связанная с окончанием дендрита сенсорного нейрона. Это клетка,
например фоторецептор, эпителиальной природы или нейроэктодермального
происхождения.
Данная классификация позволяет понять, как возникает
возбуждение рецепторов.
Механизм возбуждения рецепторов. При действии стимула на рецепторную
клетку в белково-липидном слое мембраны происходит изменение пространственной
конфигурации белковых рецепторных молекул. Это приводит к изменению
проницаемости мембраны для определенных ионов, чаще всего для ионов натрия, но
в последние годы открыта еще и роль калия в этом процессе. Возникают ионные
токи, изменяется заряд мембраны и происходит генерация рецепторного потенциала
(PП). А далее процесс возбуждения
протекает в разных рецепторах по-разному. В первично чувствующих рецепторах,
которые являются свободными голыми окончаниями чувствительного нейрона
(обонятельных, тактильных, проприоцептивных), РП воздействует на соседние,
наиболее чувствительные участки мембраны, где генерируется потенциал действия (ПД),
который далее в виде импульсов распространяется по нервному волокну.
Преобразование энергии внешнего стимула в ПД в первичных рецепторах может
происходить как непосредственно на мембране, так и при участии некоторых
вспомогательных структур. Так, например, происходит в тельце Пачини. Рецептор
здесь представлен голым окончанием аксона, которое окружено
соединительнотканной капсулой. При сдавливании тельца Пачини регистрируется РП,
который далее преобразуется в импульсный ответ афферентного волокна. Во вторично
чувствующих рецепторах, которые представлены специализированными клетками
(зрительные, слуховые, вкусовые, вестибулярные), РП приводит к образованию и
выделению медиатора из пресинаптического отдела рецепторной клетки в
синаптическую щель рецепторно-афферентного синапса. Этот медиатор воздействует
на постсинаптическую мембрану чувствительного нейрона, вызывает ее
деполяризацию и образование постсинаптического потенциала, который называют
генераторным потенциалом (ГП). ГП, воздействуя на внесинаптические участки
мембраны чувствительного нейрона, обусловливает генерацию ПД. ГП может быть как
де-, так и гиперполяризационным и соответственно вызывать возбуждение или
тормозить импульсный ответ афферентного волокна.
Свойства и особенности рецепторного и
генераторного потенциалов
Рецепторный и генераторный потенциалы - это
биоэлектрические процессы, которые обладают свойствами местного или локального
ответа: распространяются с декрементом, т.е. с затуханием; величина зависит от
силы раздражения, так как подчиняются «закону силы»; величина зависит от
скорости нарастания амплитуды стимула во времени; способны суммироваться при
применении быстро следующих друг за другом раздражений.
Итак, в рецепторах происходит преобразование энергии
стимула в нервный импульс, т.е. первичное кодирование информации,
преобразование информации в сенсорный код.
Большая часть рецепторов обладает так называемой
фоновой активностью, т.е. в них возникает возбуждение в отсутствии каких-либо
раздражителей.
Проводниковый отдел анализатора включает афферентные (периферические)
и промежуточные нейроны стволовых и подкорковых структур центральной нервной
системы (ЦНС), которые составляют как бы цепь нейронов, находящихся в разных
слоях на каждом уровне ЦНС. Проводниковый отдел обеспечивает проведение
возбуждения от рецепторов в кору большого мозга и частичную переработку
информации. Проведение возбуждения по проводниковому отделу осуществляется
двумя афферентными путями:
) специфическим проекционным путем (прямые афферентные
пути) от рецептора по строго обозначенным специфическим путям с переключением
на различных уровнях ЦНС (на уровне спинного и продолговатого мозга, в
зрительных буграх и в соответствующей проекционной зоне коры большого мозга);
) неспецифическим путем, с участием ретикулярной
формации. На уровне ствола мозга от специфического пути отходят коллатерали к
клеткам ретикулярной формации, к которым могут конвергировать различные
афферентные возбуждения, обеспечивая взаимодействие анализаторов. При этом
афферентные возбуждения теряют свои специфические свойства (сенсорную
модальность) и изменяют возбудимость корковых нейронов. Возбуждение проводится
медленно через большое число синапсов. За счет коллатералей в процесс
возбуждения включаются гипоталамус и другие отделы лимбической системы мозга, а
также двигательные центры. Все это обеспечивает вегетативный, двигательный и
эмоциональный компоненты сенсорных реакций.
Центральный, или корковый, отдел анализатора, согласно
И.П. Павлову, состоит из двух частей: центральной части, т.е. «ядра»,
представленной специфическими нейронами, перерабатывающими афферентную
импульсацию от рецепторов, и периферической части, т.е. «рассеянных элементов»
- нейронов, рассредоточенных по коре большого мозга. Корковые концы
анализаторов называют также «сенсорными зонами», которые не являются строго
ограниченными участками, они перекрывают друг друга. В настоящее время в
соответствии с цитоархитектоническими и нейрофизиологическими данными выделяют
проекционные (первичные и вторичные) и ассоциативные третичные зоны коры.
Возбуждение от соответствующих рецепторов в первичные зоны направляется по
быстропроводяшим специфическим путям, тогда как активация вторичных и третичных
(ассоциативных) зон происходит по полисинаптическим неспецифическим путям.
Кроме того, корковые зоны связаны между собой многочисленными ассоциативными
волокнами. Нейроны по толщине коры распределены неравномерно и обычно образуют
шесть слоев. Основные афферентные пути в кору заканчиваются на нейронах верхних
слоев (III - IV). Эти слои наиболее сильно развиты в центральных отделах
зрительного, слухового и кожного анализаторов. Афферентные импульсы с участием
звездчатых клеток коры (IV
слой) передаются пирамидным нейронам (III слой), отсюда обработанный сигнал уходит из коры к другим структурам мозга.
В коре входные и выходные элементы вместе со
звездчатыми клетками образуют так называемые колонки - функциональные единицы
коры, организованные в вертикальном направлении. Колонка имеет диаметр около
500 мкм и определяется зоной распределения коллатералей восходящего
афферентного таламокортикального волокна. Соседние колонки имеют взаимосвязи,
организующие участие множества колонок для осуществления той или иной реакции.
Возбуждение одной из колонок приводит к торможению соседних.
Корковые проекции сенсорных систем имеют топический
принцип организации. Объем корковой проекции пропорционален плотности
рецепторов. Благодаря этому, например, центральная ямка сетчатки в корковой
проекции представлена большей площадью, чем периферия сетчатки.
Для определения коркового представительства различных
сенсорных систем используют метод регистрации вызванных потенциалов (ВП). ВП
представляет собой один из видов вызываемой электрической активности мозга.
Сенсорные ВП регистрируются при стимуляции рецепторных образований и
используются для характеристики такой важной функции как восприятие.
Из общих принципов организации анализаторов следует
выделить многоуровневостъ и многоканалъностъ.
Многоуровневость обеспечивает возможность
специализации разных уровней и слоев ЦНС по переработке отдельных видов
информации. Это позволяет организму более быстро реагировать на простые
сигналы, анализируемые уже на отдельных промежуточных уровнях.
Существующая многоканальность анализаторных систем
проявляется в наличии параллельных нейронных каналов, т.е. в наличии в каждом
из слоев и уровней множества нервных элементов, связанных со множеством нервных
элементов следующего слоя и уровня, которые в свою очередь передают нервные
импульсы к элементам более высокого уровня, обеспечивая тем самым надежность и
точность анализа воздействующего фактора.
В то же время существующий иерархический принцип построения
сенсорных систем создает условия для тонкого регулирования процессов восприятия
посредством влияний из более высоких уровней на более низкие.
Данные особенности строения центрального отдела
обеспечивают взаимодействие различных анализаторов и процесс компенсации
нарушенных функций. На уровне коркового отдела осуществляется высший анализ и
синтез афферентных возбуждений, обеспечивающие полное представление об
окружающей среде.
Основными свойствами анализаторов являются следующие.
1. Высокая чувствительность к
адекватному раздражителю. Все отделы анализатора, и прежде всего рецепторы, обладают высокой
возбудимостью. Так, фоторецепторы сетчатки могут возбуждаться при действии лишь
нескольких квантов света, обонятельные рецепторы информируют организм о
появлении единичных молекул пахучих веществ. Однако при рассмотрении этого
свойства анализаторов предпочтительнее использовать термин «чувствительность»,
а не «возбудимость», поскольку у человека оно определяется по возникновению
ощущений.
Порог ощущения (абсолютный порог) - минимальная сила
раздражения, вызывающая такое возбуждение анализатора, которое воспринимается
субъективно в виде ощущения.
Порог различения (дифференциальный порог) -
минимальное изменение силы действующего раздражителя, воспринимаемое
субъективно в виде изменения интенсивности ощущения. Эту закономерность установил
Э. Вебер в опыте с определением по ощущению испытуемым силы давления на ладонь.
Оказалось, что при действии груза в 100 г необходимо было для ощущения прироста
давления добавить груз 3 г, при действии груза в 200 г необходимо добавить 6 г,
400 г - 12 г и т.д. При этом отношение прироста силы раздражения (DL) к силе действующего раздражителя (L) есть величина постоянная (С):
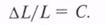
У разных анализаторов эта величина различна, в данном
случае она равна примерно 1/30 силы действующего раздражителя. Подобная
закономерность наблюдается и при уменьшении силы действующего раздражителя.
Интенсивность ощущений при одной и той же силе раздражителя
может быть различной, поскольку это зависит от уровня возбудимости различных
структур анализатора на всех его уровнях. Эту закономерность изучил Г. Фехнер,
показавший, что интенсивность ощущения прямо пропорциональна логарифму силы
раздражения. Это положение выражено формулой:

где Е - интенсивность ощущений,
К - константа,
L - сила действующего раздражителя,
L0 - порог ощущения (абсолютный порог).
Законы Вебера и Фехнера недостаточно точны, особенно
при малой силе раздражения. Психофизические методы исследования, хотя и
страдают некоторой неточностью, широко используются при исследованиях
анализаторов в практической медицине, например при определении остроты зрения,
слуха, обоняния, тактильной чувствительности, вкуса.
2. Инерционность - сравнительно медленное
возникновение и исчезновение ощущений. Латентное время возникновения ощущений
определяется латентным периодом возбуждения рецепторов и временем, необходимым
для перехода возбуждения в синапсах с одного нейрона на другой, временем
возбуждения ретикулярной формации и генерализации возбуждения в коре больших
полушарий. Сохранение на некоторый период ощущений после выключения
раздражителя объясняется явлением последействия в ЦНС - в основном циркуляцией
возбуждения. Так, зрительное ощущение не возникает и не исчезает мгновенно.
Латентный период зрительного ощущения равен 0,1 с, время последействия -0,05 с.
Быстро следующие одно за другим световые раздражения (мелькания) могут давать
ощущение непрерывного света (феномен «слияния мельканий»). Максимальная частота
вспышек света, которые воспринимаются еще раздельно, называется критической
частотой мельканий, которая тем больше, чем сильнее яркость стимула и выше
возбудимость ЦНС, и составляет около 20 мельканий в секунду. Наряду с этим,
если два неподвижных стимула последовательно с интервалом в 20-200 мс
проецировать на разные участки сетчатки, возникает ощущение движения объекта.
Это явление получило название «Фи-феномена». Такой эффект наблюдается даже в
том случае, когда один стимул несколько отличается по форме от другого. Эти два
феномена: «слияние мельканий» и «Фи-феномен» - лежат в основе кинематографии. В
силу инерционности восприятия зрительное ощущение от одного кадра длится до
появления другого, отчего и возникает иллюзия непрерывного движения. Обычно
такой эффект возникает при быстром последовательном предъявлении неподвижных
изображений на экране со скоростью 18-24 кадра в секунду.
3. Способность сенсорной системы к адаптации при
постоянной силе длительно действующего раздражителя заключается в основном в
понижении абсолютной и повышении дифференциальной чувствительности. Это
свойство присуще всем отделам анализатора, но наиболее ярко оно проявляется на
уровне рецепторов и заключается в изменении не только их возбудимости и
импульсации, но и показателей функциональной мобильности, т.е. в изменении
числа функционирующих рецепторных структур (П.Г. Снякин). По скорости адаптации
все рецепторы делят на быстро и медленно адаптирующиеся, иногда выделяют и
среднюю по скорости адаптации группу рецепторов. В проводниковом и корковом
отделах анализаторов адаптация проявляется в уменьшении числа активированных
волокон и нервных клеток.
Важную роль в сенсорной адаптации играет эфферентная
регуляция, которая осуществляется путем нисходящих влияний, изменяющих
деятельность нижерасположенных структур сенсорной системы. Благодаря этому
возникает феномен «настройки» сенсорных систем на оптимальное восприятие
раздражителей в условиях изменившейся среды.
4. Взаимодействие анализаторов. С помощью анализаторов организм
познает свойства предметов и явлений окружающей среды, полезные и негативные
стороны их воздействия на организм. Поэтому нарушения функции внешних
анализаторов, особенно зрительного и слухового, чрезвычайно сильно затрудняют
познание внешнего мира (очень беден окружающий мир для слепого или глухого).
Однако только аналитические процессы в ЦНС не могут создать реального
представления об окружающей среде. Способность анализаторов взаимодействовать
между собой обеспечивает образное и целостное представление о предметах
внешнего мира. Например, качество дольки лимона мы оцениваем с помощью
зрительного, обонятельного, тактильного и вкусового анализаторов. При этом
формируется представление как об отдельных качествах - цвете, консистенции,
запахе, вкусе, так и о свойствах объекта в целом, т.е. создается определенный
целостный образ воспринимаемого объекта. Взаимодействие анализаторов при оценке
явлений и предметов лежит также в основе компенсации нарушенных функций при
утрате одного из анализаторов. Так, у слепых повышается чувствительность
слухового анализатора. Такие люди могут определить местоположение крупных
предметов и обойти их, если нет посторонних шумов. Это осуществляется за счет
отражения звуковых волн от находящегося впереди предмета. Американские
исследователи наблюдали за слепым человеком, который достаточно точно определял
местоположение большой картонной пластинки. Когда испытуемому залепили уши
воском, он не смог больше определять местоположение картона.
Взаимодействия сенсорных систем могут проявляться в
виде влияния возбуждения одной системы на состояние возбудимости другой по
доминантному принципу. Так, прослушивание музыки может вызвать обезболивание
при стоматологических процедурах (аудиоаналгезия). Шум ухудшает зрительное
восприятие, яркий свет повышает восприятие громкости звука. Процесс
взаимодействия сенсорных систем может проявляться на различных уровнях.
Особенно большую роль в этом играет ретикулярная формация ствола мозга, кора
большого мозга. Многие нейроны коры обладают способностью отвечать на сложные
комбинации сигналов разной модальности (мультисенсорная конвергенция), что
очень важно для познания окружающей среды и оценки новых раздражителей.
Кодирование
информации в анализаторах
Понятия. Кодирование - процесс преобразования информации в
условную форму (код), удобную для передачи по каналу связи. Любое
преобразование информации в отделах анализатора является кодированием. В
слуховом анализаторе механическое колебание перепонки и других звукопроводящих
элементов на первом этапе преобразуется в рецепторный потенциал, последний
обеспечивает выделение медиатора в синаптическую щель и возникновение
генераторного потенциала, в результате действия которого в афферентном волокне
возникает нервный импульс. Потенциал действия достигает следующего нейрона, в
синапсе которого электрический сигнал снова превращается в химический, т. е.
многократно меняется код. Следует отметить, что на всех уровнях анализаторов не
происходит восстановления стимула в его первоначальной форме. Этим
физиологическое кодирование отличается от большинства технических систем связи,
где сообщение, как правило, восстанавливается в первоначальном виде.
Коды нервной системы. В вычислительной технике используется
двоичный код, когда для образования комбинаций всегда используются два символа
- 0 и 1, которые представляют собой два состояния. Кодирование информации в
организме осуществляется на основе недвоичных кодов, что позволяет при той же
длине кода получить большее число комбинаций. Универсальным кодом нервной
системы являются нервные импульсы, которые распространяются по нервным
волокнам. При этом содержание информации определяется не амплитудой импульсов
(они подчиняются закону «Все или ничего»), а частотой импульсов (интервалами
времени между отдельными импульсами), объединением их в пачки, числом импульсов
в пачке, интервалами между пачками. Передача сигнала от одной клетки к другой
во всех отделах анализатора осуществляется с помощью химического кода, т.е.
различных медиаторов. Для хранения информации в ЦНС кодирование осуществляется
с помощью структурных изменений в нейронах (механизмы памяти).
Кодируемые характеристики
раздражителя. В
анализаторах кодируются качественная характеристика раздражителя (например,
свет, звук), сила раздражителя, время его действия, а также пространство, т.е.
место действия раздражителя и локализация его в окружающей среде. В кодировании
всех характеристик раздражителя принимают участие все отделы анализатора.
В периферическом отделе анализатора кодирование качества раздражителя
(вид) осуществляется за счет специфичности рецепторов, т.е. способности
воспринимать раздражитель определенного вида, к которому он приспособлен в
процессе эволюции, т.е. к адекватному раздражителю. Так, световой луч
возбуждает только рецепторы сетчатки, другие рецепторы (обоняния, вкуса,
тактильные и т.д.) на него обычно не реагируют.
Сила раздражителя может кодироваться изменением
частоты импульсов в генерируемых рецепторами при изменении силы раздражителя,
что определяется общим количеством импульсов в единицу времени. Это так
называемое частотное кодирование. При этом с увеличением силы стимула обычно
возрастает число импульсов, возникающих в рецепторах, и наоборот. При изменении
силы раздражителя может изменяться и число возбужденных рецепторов, кроме того,
кодирование силы раздражителя может осуществляться различной величиной
латентного периода и временем реакции. Сильный раздражитель уменьшает латентный
период, увеличивает число импульсов и удлиняет время реакции. Пространство
кодируется величиной площади, на которой возбуждаются рецепторы, это
пространственное кодирование (например, мы легко определяем, острым или тупым
концом карандаш касается поверхности кожи). Некоторые рецепторы легче возбуждаются
при действии на них раздражителя под определенным углом (тельца Пачини,
рецепторы сетчатки), что является оценкой направления действия раздражителя на
рецептор. Локализация действия раздражителя кодируется тем, что рецепторы
различных участков тела посылают импульсы в определенные зоны коры большого
мозга.
Время действия раздражителя на рецептор кодируется
тем, что он начинает возбуждаться с началом действия раздражителя и прекращает
возбуждаться сразу после выключения раздражителя (временное кодирование).
Следует заметить, что время действия раздражителя во многих рецепторах
кодируется недостаточно точно вследствие быстрой их адаптации и прекращения
возбуждения при постоянно действующей силе раздражителя. Эта неточность
частично компенсируется за счет наличия on-, off- и
on-off-рецепторов, возбуждающихся соответственно при
включении, выключении, а также при включении и выключении раздражителя. При
длительно действующем раздражителе, когда происходит адаптация рецепторов,
теряется некоторое количество информации о стимуле (его силе и
продолжительности), но при этом повышается чувствительность, т. е. развивается
сенситизация рецептора к изменению этого стимула. Усиление стимула действует на
адаптированный рецептор как новый раздражитель, что также отражается в
изменении частоты импульсов, идущих от рецептора.
В проводниковом отделе анализатора кодирование
осуществляется только на «станциях переключения», т. е. при передаче сигнала от
одного нейрона к другому, где происходит смена кода. В нервных волокнах информация
не кодируется, они исполняют роль проводов, по которым передается информация,
закодированная в рецепторах и переработанная в центрах нервной системы.
Между импульсами в отдельном нервном волокне могут
быть различные интервалы, импульсы формируются в пачки с различным числом,
между отдельными пачками могут быть также различные интервалы. Все это отражает
характер закодированной в рецепторах информации. В нервном стволе при этом
может изменяться также число возбужденных нервных волокон, что определяется
изменением числа возбужденных рецепторов или нейронов на предыдущем переходе
сигнала с одного нейрона на другой. На станциях переключения, например в
зрительном бугре, информация кодируется, во-первых, за счет изменения объема
импульсации на входе и на выходе, а во-вторых, за счет пространственного
кодирования, т.е. за счет связи определенных нейронов с определенными
рецепторами. В обоих случаях чем сильнее раздражитель, тем большее число
нейронов возбуждается.
В вышележащих отделах ЦНС наблюдаются уменьшение
частоты разрядов нейронов и превращение длительной импульсации в короткие пачки
импульсов. Имеются нейроны, возбуждающиеся не только при появлении стимула, но
и при его выключении, что также связано с активностью рецепторов и
взаимодействием самих нейронов. Нейроны, получившие название «детекторов»,
избирательно реагируют на тот или иной параметр стимула, например на стимул,
движущийся в пространстве, или на светлую либо темную полоску, расположенную в
определенной части поля зрения. Количество таких нейронов, которые лишь
частично отражают свойства стимула, возрастает на каждом последующем уровне
анализатора. Но в то же время на каждом последующем уровне анализатора имеются
нейроны, дублирующие свойства нейронов предыдущего отдела, что создает основу
надежности функции анализаторов. В сенсорных ядрах происходят тормозные
процессы, которые осуществляют фильтрацию и дифференциацию сенсорной
информации. Эти процессы обеспечивают контроль сенсорной информации. При этом
снижается шум и изменяется соотношение спонтанной и вызванной активности
нейронов. Такой механизм реализуется за счет разновидностей торможения
(латерального, возвратного) в процессе восходящих и нисходящих влияний.
В корковом конце анализатора происходит частотно-пространственное
кодирование, нейрофизиологической основой которого является пространственное
распределение ансамблей специализированных нейронов и их связей с определенными
видами рецепторов. Импульсы поступают от рецепторов в определенные зоны коры с
различными временными интервалами. Поступающая в виде нервных импульсов
информация перекодируется в структурные и биохимические изменения в нейронах
(механизмы памяти). В коре мозга осуществляется высший анализ и синтез
поступившей информации.
Анализ заключается в том, что с помощью возникающих
ощущений мы различаем действующие раздражители (качественно - свет, звук и
т.д.) и определяем силу, время и место, т.е. пространство, на которое действует
раздражитель, а также его локализацию (источник звука, света, запаха).
Синтез реализуется в узнавании известного предмета,
явления или в формировании образа, впервые встречаемого предмета, явления.
Известны случаи, когда у слепых от рождения зрение
появлялось только в подростковом возрасте. Так, девушка, которая обрела зрение
лишь в 16 лет, не могла с помощью зрения узнать предметы, которыми она
многократно пользовалась ранее. Но стоило ей взять предмет в руки, как она с
радостью называла его. Ей пришлось, таким образом, практически заново изучать
окружающий ее мир с участием зрительного анализатора, подкреплением информацией
от других анализаторов, в частности от тактильного. При этом тактильные
ощущения оказались решающими. Об этом свидетельствует, например, и давний опыт
Стратона. Известно, что изображение на сетчатке глаза является уменьшенным и
перевернутым. Новорожденный видит мир именно таким. Однако в раннем онтогенезе
ребенок все трогает руками, сопоставляет и сличает зрительные ощущения с
тактильными. Постепенно взаимодействие тактильных и зрительных ощущений ведет к
восприятию расположения предметов, каким оно является в реальной
действительности, хотя на сетчатке изображение остается перевернутым. Стратон
надел очки с линзами, которые перевернули изображение на сетчатке в положение,
соответствующее реальной действительности. Наблюдаемый окружающий мир
перевернулся «вверх ногами». Однако в течение 8 дней он с помощью сравнения
тактильных и зрительных ощущений снова стал воспринимать все вещи и предметы
как обычно. Когда экспериментатор снял очки-линзы, мир снова «перевернулся»,
нормальное восприятие вернулось через 4 дня.
Если информация о предмете или явлении поступает в
корковый отдел анализатора впервые, то формируется образ нового предмета,
явления благодаря взаимодействию нескольких анализаторов. Но и при этом идет
сличение поступающей информации со следами памяти о других подобных предметах
или явлениях. Поступившая в виде нервных импульсов информация кодируется с
помощью механизмов долговременной памяти.
Итак, процесс передачи сенсорного сообщения
сопровождается многократным перекодированием и завершается высшим анализом и
синтезом, который происходит в корковом отделе анализаторов. После этого уже
происходит выбор или разработка программы ответной реакции организма.
сенсорный рецепторный зрительный
анализатор
Таблица
Общий план строения сенсорных систем
|
Название анализатора
|
Природа раздражителя
|
Периферический отдел
|
Проводниковый отдел
|
Центральный отел
|
|
зрительный
|
Электромагнитные колебания,
отраженные или излученные объектами внешнего мира и воспринимаемые органами
зрения.
|
Палочковые и колбочковые
нейросенсорные клетки, наружные сегменты которых имеют соответственно
палочковидную («палочки») и колбочковидную («колбочки») формы. Палочки
являются рецепторами, воспринимающими световые лучи в условиях слабой
освещенности, т.е. бесцветное, или ахроматическое, зрение. Колбочки же
функционируют в условиях яркой освещенности и характеризуются разной
чувствительностью к спектральным свойствам света (цветное или хроматическое
зрение)
|
Первый нейрон
проводникового отдела зрительного анализатора представлен биполярными
клетками сетчатки. Аксоны биполярных клеток в свою очередь конвергируют на
ганглиозные клетки (второй нейрон). Биполярные и ганглиозные клетки
взаимодействуют между собой за счет многочисленных латеральных связей,
образованных коллатералями дендритов и аксонов самих клеток, а также с
помощью амакриновых клеток
|
Расположен в затылочной
доле. Имеются сложные и сверхсложные рецептивные поля детекторного типа. Эта
особенность позволяет выделять из цельного изображения лишь отдельные части линий
с различным расположением и ориентацией, при этом проявляется способность
избирательно реагировать на эти фрагменты.
|
|
слуховой
|
Звуки, т. е. колебательные
движения частиц упругих тел, распространяющихся в виде волн в самых различных
средах, включая воздушную среду, и воспринимающиеся ухом
|
Превращающий энергию
звуковых волн в энергию нервного возбуждения, представлен рецепторными
волосковыми клетками кортиева органа (орган Корти), находящимися в улитке.
Внутреннее ухо (звуковоспринимающий аппарат), а также среднее ухо
(звукопередающий аппарат) и наружное ухо (звукоулавливающий аппарат)
объединяются в понятие орган слуха
|
Представлен периферическим
биполярным нейроном, расположенным в спиральном ганглии улитки (первый
нейрон). Волокна слухового (или кохлеарного) нерва, образованные аксонами
нейронов спирального ганглия, заканчиваются на клетках ядер кохлеарного
комплекса продолговатого мозга (второй нейрон). Затем после частичного
перекреста волокна идут в медиальное коленчатое тело метаталамуса, где опять
происходит переключение (третий нейрон), отсюда возбуждение поступает в кору
(четвертый нейрон). В медиальных (внутренних) коленчатых телах, а также в
нижних буграх четверохолмия располагаются центры рефлекторных двигательных
реакций, возникающих при действии звука.
|
Находится в верхней части
височной доли большого мозга. Важное значение для функции слухового
анализатора имеют поперечные височные извилины (извилины Гешля).
|
|
Вестибулярный
|
Обеспечивает так называемое
акселерационное чувство, т.е. ощущение, возникающее при прямолинейном и
вращательном ускорении движения тела, а также при изменениях положения
головы. Вестибулярному анализатору принадлежит ведущая роль в
пространственной ориентации человека, сохранении его позы.
|
Представлен волосковыми
клетками вестибулярного органа, расположенного, как и улитка, в лабиринте
пирамиды височной кости. Вестибулярный орган (орган равновесия, орган
гравитации) состоит из трех полукружных каналов и преддверия. Преддверие
состоит из двух мешочков: круглого (саккулюс), расположенного ближе к улитке,
и овального (утрикулюс), расположенного ближе к полукружным каналам. Для
волосковых клеток преддверия адекватными раздражителями являются ускорение
или замедление прямолинейного движения тела, а также наклоны головы. Для
волосковых клеток полукружных каналов адекватным раздражителем является
ускорение или замедление вращательного движения в какой-либо плоскости
|
К рецепторам подходят
периферические волокна биполярных нейронов вестибулярного ганглия,
расположенного во внутреннем слуховом проходе (первый нейрон). Аксоны этих
нейронов в составе вестибулярного нерва направляются к вестибулярным ядрам
продолговатого мозга (второй нейрон). Вестибулярные ядра продолговатого мозга
(верхнее - ядро Бехтерева, медиальное - ядро Швальбе, латеральное - ядро
Дейтерса и нижнее - ядро Роллера) получают дополнительную информацию по
афферентным нейронам от проприорецепторов мышц или от суставных сочленений
шейного отдела позвоночника. Эти ядра вестибулярного анализатора тесно
связаны с различными отделами центральной нервной системы. Благодаря этому
обеспечиваются контроль и управление эффекторными реакциями соматического,
вегетативного и сенсорного характера. Третий нейрон расположен в ядрах
зрительного бугра, откуда возбуждение направляется в кору полушарий.
|
Центральный отдел
вестибулярного анализатора локализуется в височной области коры большого
мозга, несколько кпереди от слуховой проекционной зоны (21 - 22 поля по
Бродману, четвертый нейрон).
|
|
Двигательный
|
Обеспечивает формирование
так называемого мышечного чувства при изменении напряжения мышц, их оболочек,
суставных сумок, связок, сухожилий. В мышечном чувстве можно выделить три
составляющих: чувство положения, когда человек может определить положение
своих конечностей и их частей относительно друг друга; чувство движения,
когда, изменяя угол сгибания в суставе, человек осознает скорость и
направление движения; чувство силы, когда человек может оценить мышечную
силу, нужную для движения или удерживания суставов в определенном положении
при подъеме или перемещении груза. Наряду с кожным, зрительным, вестибулярным
двигательный анализатор оценивает положение тела в пространстве, позу,
участвует в координации мышечной деятельности
|
Представлен
проприорецепторами, расположенными в мышцах, связках, сухожилиях, суставных
сумках, фасциях. К ним относятся мышечные веретена, тельца Гольджи, тельца
Пачини, свободные нервные окончания. Мышечное веретено представляет собой
скопление тонких коротких поперечно-полосатых мышечных волокон, которые
окружены соединительнотканной капсулой. Мышечное веретено с интрафузальными
волокнами расположено параллельно экстрафузальным, поэтому возбуждаются при
расслаблении (удлинении) скелетной мышцы. Тельца Гольджи находятся в
сухожилиях. Это гроздевидные чувствительные окончания. Тельца Гольджи,
располагаясь в сухожилиях, включены относительно скелетной мышцы
последовательно, поэтому они возбуждаются при ее сокращении вследствие
натяжения сухожилия мышцы. Рецепторы Гольджи контролируют силу мышечнюго
сокращения, т.е. напряжения. Тельца Панины представляют собой
инкапсулированные нервные окончания, локализуются в глубоких слоях кожи, в
сухожилиях и связках, реагируют на изменения давления, которое возникает при
сокращении мышц и натяжении сухожилий, связок и кожи.
|
Представлен нейронами,
которые располагаются в спинальных ганглиях (первый нейрон). Отростки этих
клеток в составе пучков Голля и Бурдаха (задние столбы спинного мозга)
достигают нежного и клиновидного ядер продолговатого мозга, где располагаются
вторые нейроны. От этих нейронов волокна мышечно-суставной чувствительности,
совершив перекрест, в составе медиальной петли доходят до зрительного бугра,
где в вентральных заднелатеральном и заднемедиальном ядрах располагаются
третьи нейроны.
|
Центральным отделом двигательного
анализатора являются нейроны передней центральной извилины.
|
|
Внутренние (висцеральные)
|
Осуществляют анализ и
синтез информации о состоянии внутренней среды организма и участвуют в
регуляции работы внутренних органов. Можно выделить: 1) внутренний анализатор
давления в кровеносных сосудах и давления (наполнений) во внутренних полых
органах (периферическим отделом этого анализатора являются механорецепторы);
2) анализатор температуры; 3) анализатор химизма внутренней среды организма;
4) анализатор осмотического давления внутренней среды.
|
К механорецепторам
относятся все рецепторы, для которых адекватными стимулами являются давление,
а также растяжение, деформация стенок органов (сосуды, сердце, легкие,
желудочно-кишечный тракт и другие внутренние полые органы). К хеморецедторам
относят всю массу рецепторов, реагирующих на различные химические вещества:
это рецепторы аортального и каротидного клубочков, рецепторы слизистых
оболочек пищеварительного тракта и органов дыхания, рецепторы серозных
оболочек, а также хеморецепторы головного мозга. Осморецепторы локализованы в
аортальном и каротидном синусах, в других сосудах артериального русла, в
интерстициальной ткани вблизи капилляров, в печени и других органах. Часть
осморецепторов является механорецепторами, часть - хеморецепторами.
Терморецепторы локализованы в слизистых оболочках пищеварительного тракта,
органов дыхания, мочевого пузыря, серозных оболочках, в стенках артерий и
вен, в каротидном синусе, а также в ядрах гипоталамуса.
|
От интерорецепторов
возбуждение в основном проходит в одних стволах с волокнами вегетативной
нервной системы. Первые нейроны находятся в соответствующих чувствительных
ганглиях, вторые нейроны - в спинном или продолговатом мозге. Восходящие пути
от них достигают заднемедиальное ядро таламуса (третий нейрон) и затем
поднимаются в кору больших полушарий (четвертый нейрон).
|
Корковый отдел локализуется
в зонах С1 и С2 соматосенсорной области коры и в
орбитальной области коры большого мозга. Восприятие некоторых интероцептивных
стимулов может сопровождаться возникновением четких, локализованных ощущений,
например при растяжении стенок мочевого пузыря или прямой кишки. Но
висцеральная импульсация (от интерорецепторов сердца, сосудов, печени, почек
и др.) может и не вызывать ясно осознаваемых ощущений. Обусловлено это тем,
что такие ощущения возникают в результате раздражения различных рецепторов,
входящих в ту или иную систему органов. В любом случае изменения внутренних
органов оказывают значительное влияние на эмоциональное состояние и характер
поведения человека
|
|
Температурный
|
Обеспечивает информацию о
температуре внешней среды и формирование температурных ощущений
|
Представлен двумя видами
рецепторов: одни реагируют на холодовые стимулы, другие - на тепловые.
Тепловые рецепторы - это тельца Руффини, а холодовые - колбы Краузе.
Рецепторы холода расположены в эпидермисе и непосредственно под ним, а
рецепторы тепла - преимущественно в нижнем и верхнем слоях собственно кожи и
слизистой оболочки.
|
От рецепторов холода
отходят миелинизированные волокна типа А, а от рецепторов тепла -
немиелинизированные волокна типа С, поэтому информация от холодовых
рецепторов распространяется с большей скоростью, чем от тепловых. Первый
нейрон локализуется в спинальных ганглиях. Клетки задних рогов спинного мозга
представляют второй нейрон. Нервные волокна, отходящие от вторых нейронов
температурного анализатора, переходят через переднюю комиссуру на
противоположную сторону в боковые столбы и в составе латерального
спинно-таламического тракта доходят до зрительного бугра, где находится
третий нейрон. Отсюда возбуждение поступает в кору полушарий большого мозга.
|
Центральный отдел
температурного анализатора локализуется в области задней центральной извилины
коры большого мозга.
|
|
Тактильный
|
Представлен различными
рецепторными образованиями, раздражение которых приводит к формированию
специфических ощущений. На поверхности кожи, лишенной волос, а также на
слизистых оболочках на прикосновение реагируют специальные рецепторные клетки
(тельца Мейснера), расположенные в сосочковом слое кожи. На коже, покрытой
волосами, на прикосновение реагируют рецепторы волосяного фолликула,
обладающие умеренной адаптацией.
|
От большинства
механорецепторов в спинной мозг информация поступает в центральную нервную
систему по А-волокнам и лишь от рецепторов щекотки - по С-волокнам. Первый
нейрон находится в спинальных ганглиях. В заднем роге спинного мозга
происходит первое переключение на интернейроны (второй нейрон), от них
восходящий путь в составе заднего столба достигает ядер заднего столба в
продолговатом мозге (третий нейрон), где происходит второе переключение,
далее через медиальную петлю путь следует к вентро-базальным ядрам
зрительного бугра (четвертый нейрон), центральные отростки нейронов
зрительного бугра идут в кору больших полушарий.
|
Локализуется в 1 и II
зонах соматосенсорной области коры большого мозга (задняя центральная
извилина).
|
|
Вкусовой
|
Возникающее чувство вкуса
связано с раздражением не только химических, но и механических, температурных
и даже болевых рецепторов слизистой оболочки полости рта, а также
обонятельных рецепторов. Вкусовой анализатор определяет формирование вкусовых
ощущений, является рефлексогенной зоной.
|
Рецепторы вкуса (вкусовые
клетки с микроворсинками) - это вторичные рецепторы, они являются элементом
вкусовых почек, в состав которых входят также опорные и базальные клетки. Во
вкусовых почках обнаружены клетки, содержащие серотонин, и клетки, образующие
гистамин. Эти и другие вещества играют определенную роль в формировании
чувства вкуса. Отдельные вкусовые почки являются полимодальными
образованиями, так как могут воспринимать различные виды вкусовых
раздражителей. Вкусовые почки в виде отдельных включений находятся на задней
стенке глотки, мягком нёбе, миндалинах, гортани, надгортаннике и входят также
в состав вкусовых сосочков языка как органа вкуса.
|
Внутрь вкусовой почки
входят нервные волокна, которые образуют рецепторно-афферентные синапсы.
Вкусовые почки различных областей полости рта получают нервные волокна от
разных нервов: вкусовые почки передних двух третей языка - от барабанной
струны, входящей в состав лицевого нерва; почки задней трети языка, а также
мягкого и твердого нёба, миндалин - от языкоглоточного нерва; вкусовые почки,
расположенные в области глотки, надгортанника и гортани, - от
верхне-гортанного нерва, являющегося частью блуждающего нерва
|
Локализуется в нижней части
соматосенсорной зоны коры в области представительства языка. Большая часть
нейронов этой области мультимодальна, т.е. реагирует не только на вкусовые,
но и на температурные, механические и ноцицептивные раздражители. Для
вкусовой сенсорной системы характерно то, что каждая вкусовая почка имеет не
только афферентные, но и эфферентные нервные волокна, которые подходят к
вкусовым клеткам из ЦНС, благодаря чему обеспечивается включение вкусового
анализатора в целостную деятельность организма.
|
|
Обонятельный
|
Запахи
|
Первично-чувствующие
рецепторы, которые являются окончаниями дендрита так называемой
нейросекреторной клетки. Верхняя часть дендрита каждой клетки несет 6-12
ресничек, а от основания клетки отходит аксон. Реснички, или обонятельные
волоски, погружены в жидкую среду - слой слизи, вырабатываемой боуменовыми
железами. Наличие обонятельных волосков значительно увеличивает площадь
контакта рецептора с молекулами пахучих веществ. Движение волосков
обеспечивает активный процесс захвата молекул пахучего вещества и контакта с
ним, что лежит в основе целенаправленного восприятия запахов. Рецепторные
клетки обонятельного анализатора погружены в обонятельный эпителий,
выстилающий полость носа, в котором кроме них имеются опорные клетки,
выполняющие механическую функцию и активно участвующие в метаболизме
обонятельного эпителия. Часть опорных клеток, располагающихся вблизи базальной
мембраны, носит название базальных
|
Первым нейроном
обонятельного анализатора следует считать нейросенсорную или нейрорецепторную
клетку. Аксон этой клетки образует синапсы, называемые гломерулами, с главным
дендритом митральных клеток обонятельной луковицы, которые представляют
второй нейрон. Аксоны митральных клеток обонятельных луковиц образуют
обонятельный тракт, который имеет треугольное расширение (обонятельный
треугольник) и состоит из нескольких пучков. Волокна обонятельного тракта
отдельными пучками идут в передние ядра зрительного бугра. Некоторые
исследователи считают, что отростки второго нейрона идут прямо в кору
большого мозга, минуя зрительные бугры.
|
Локализуется в передней
части грушевидной доли коры в области извилины морского коня.
|
|
Болевой
|
Боль - это «сенсорная
модальность» подобно слуху, вкусу, зрению и пр., она выполняет сигнальную
функцию, которая заключается в информации о нарушении таких жизненно важных
констант организма, как целостность покровных оболочек и определенный уровень
окислительных процессов в тканях, обеспечивающих их нормальную
жизнедеятельность. В то же время боль можно рассматривать как
психофизиологическое состояние, сопровождаемое изменениями деятельности
различных органов и систем, а также возникновением эмоций и мотиваций.
|
Представлен рецепторами
боли, которые по предложению Ч. Шеррингтона называют ноцицепторами. Это
высокопороговые рецепторы, реагирующие на разрушающие воздействия. По
механизму возбуждения ноцицепторы делят на механоноцицепторы и
хемоноцицепторы. Механоноцицепторы расположены преимущественно в коже,
фасциях, сухожилиях, суставных сумках и слизистых оболочках пищеварительного
тракта. Хемоноцицепторы расположены также на коже и в слизистых оболочках, но
превалируют во внутренних органах, где локализуются в стенках мелких артерий.
|
Проведение болевого
возбуждения от рецепторов осуществляется по дендритам первого нейрона,
расположенного в чувствительных ганглиях соответствующих нервов,
иннервирующих определенные участки организма. Аксоны этих нейронов поступают
в спинной мозг к вставочным нейронам заднего рога (второй нейрон). Далее
проведение возбуждения в центральной нервной системе осуществляется двумя
путями: специфическим (лемнисковым) и неспецифическим (экстралемнисковым).
Специфический путь начинается от вставочных нейронов спинного мозга, аксоны
которых в составе спиноталамического тракта поступают к специфическим ядрам
таламуса (в частности, в вентробазальное ядро), которые представляют третьи
нейроны. Отростки этих нейронов достигают коры. Неспецифический путь
начинается также от вставочного нейрона спинного мозга и по коллатералям идет
к различным структурам мозга. В зависимости от места окончания выделяют три
основных тракта - неоспиноталамический, спиноретикулярный,
спиномезенцефалический. Последние два тракта объединяются в
спиноталамический. Возбуждение по этим трактам поступает в неспецифические
ядра таламуса и оттуда во все отделы коры больших полушарий.
|
Специфический путь
заканчивается в соматосенсорной области коры большого мозга. Согласно
современным представлениям выделяют две соматосенсорные зоны. Первичная
проекционная зона находится в области заднецентральной извилины. Здесь
происходит анализ ноцицептивных воздействий, формирование ощущения острой,
точно локализованной боли. Кроме того, за счет тесных связей с моторной зоной
коры осуществляются моторные акты при воздействии повреждающих стимулов.
Вторичная проекционная зона, которая находится в глубине сильвиевой борозды,
участвует в процессах осознания и выработке программы поведения при болевом
воздействии. Неспецифический путь распространяется на все области коры.
Значительную роль в формировании болевой чувствительности играет
орбитофронтальная область коры, которая участвует в организации
эмоционального и вегетативного компонентов боли.
|
2. Зрительный анализатор
Зрительный анализатор представляет собой совокупность
структур, воспринимающих световую энергию в виде электромагнитного излучения с
длиной волны 400-700 нм и дискретных частиц фотонов, или квантов, и формирующих
зрительные ощущения. С помощью глаза воспринимается 80 - 90% всей информации об
окружающем мире.
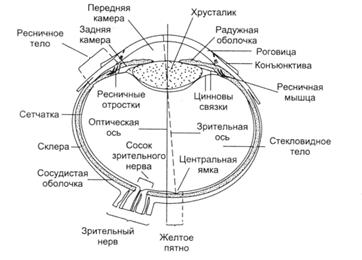
Рис. 2.1 Орган зрения
Благодаря деятельности зрительного анализатора
различают освещенность предметов, их цвет, форму, величину, направление
передвижения, расстояние, на которое они удалены от глаза и друг от друга. Все
это позволяет оценивать пространство, ориентироваться в окружающем мире,
выполнять различные виды целенаправленной деятельности.
Наряду с понятием зрительного анализатора существует
понятие органа зрения (рис. 2.1)
Орган зрения - это глаз, включающий три различных
в функциональном отношении элемента:
) глазное яблоко, в котором расположены
световоспринимающий, светопреломляющий и светорегулирующий аппараты;
) защитные приспособления, т.е. наружные оболочки
глаза (склера и роговица), слезный аппарат, веки, ресницы, брови; 3)
двигательный аппарат, представленный тремя парами глазных мышц (наружная и
внутренняя прямые, верхняя и нижняя прямые, верхняя и нижняя косые), которые
иннервируются III (глазодвигательный нерв), IV (блоковый нерв) и VI (отводящий нерв) парами черепных
нервов.
Структурно-функциональная
характеристика
Рецепторный (периферический) отдел зрительного анализатора
(фоторецепторы) подразделяется на палочковые и колбочковые нейросенсорные
клетки, наружные сегменты которых имеют соответственно палочковидную
(«палочки») и колбочковидную («колбочки») формы. У человека насчитывается 6 - 7
млн колбочек и 110-125 млн палочек.
Место выхода зрительного нерва из сетчатки не содержит
фоторецепторов и называется слепым пятном. Латерально от слепого пятна в
области центральной ямки лежит участок наилучшего видения - желтое пятно,
содержащее преимущественно колбочки. К периферии сетчатки число колбочек
уменьшается, а число палочек возрастает, и периферия сетчатки содержит одни
лишь палочки.
Различия функций колбочек и палочек лежит в основе
феномена двойственности зрения. Палочки являются рецепторами, воспринимающими
световые лучи в условиях слабой освещенности, т.е. бесцветное, или
ахроматическое, зрение. Колбочки же функционируют в условиях яркой освещенности
и характеризуются разной чувствительностью к спектральным свойствам света
(цветное или хроматическое зрение). Фоторецепторы обладают очень высокой
чувствительностью, что обусловлено особенностью строения рецепторов и
физико-химических процессов, лежащих в основе восприятия энергии светового
стимула. Полагают, что фоторецепторы возбуждаются при действии на них 1-2
квантов света.
Палочки и колбочки состоят из двух сегментов -
наружного и внутреннего, которые соединяются между собой посредством узкой
реснички. Палочки и колбочки ориентированы в сетчатке радиально, а молекулы
светочувствительных белков расположены в наружных сегментах таким образом, что
около 90% их светочувствительных групп лежат в плоскости дисков, входящих в
состав наружных сегментов. Свет оказывает наибольшее возбуждающее действие в
том случае, если направление луча совпадает с длинной осью палочки или
колбочки, при этом он направлен перпендикулярно дискам их наружных сегментов.
Фотохимические процессы в сетчатке
глаза. В рецепторных
клетках сетчатки находятся светочувствительные пигменты (сложные белковые
вещества) - хромопротеиды, которые обесцвечиваются на свету. В палочках на
мембране наружных сегментов содержится родопсин, в колбочках - йодопсин и
другие пигменты.
Родопсин и йодопсин состоят из ретиналя (альдегида
витамина А1) и гликопротеида (опсина). Имея сходство в
фотохимических процессах, они различаются тем, что максимум поглощения
находится в различных областях спектра. Палочки, содержащие родопсин, имеют
максимум поглощения в области 500 нм. Среди колбочек различают три типа,
которые отличаются максимумами в спектрах поглощения: одни имеют максимум в
синей части спектра (430-470 нм), другие в зеленой (500-530), третьи - в
красной (620-760 нм) части, что обусловлено наличием трех типов зрительных
пигментов. Красный колбочковый пигмент получил название «йодопсин». Ретиналь
может находиться в различных пространственных конфигурациях (изомерных формах),
но только одна из них - 11-ЦИС-изомер ретиналя выступает в качестве хромофорной
группы всех известных зрительных пигментов. Источником ретиналя в организме
служат каротиноиды.
Фотохимические процессы в сетчатке протекают весьма
экономно. Даже при действии яркого света расщепляется только небольшая часть
имеющегося в палочках родопсина (около 0,006%).
В темноте происходит ресинтез пигментов, протекающий с
поглощением энергии. Восстановление йодопсина протекает в 530 раз быстрее, чем
родопсина. Если в организме снижается содержание витамина А, то процессы
ресинтеза родопсина ослабевают, что приводит к нарушению сумеречного зрения,
так называемой куриной слепоте. При постоянном и равномерном освещении
устанавливается равновесие между скоростью распада и ресинтеза пигментов. Когда
количество света, падающего на сетчатку, уменьшается, это динамическое
равновесие нарушается и сдвигается в сторону более высоких концентраций
пигмента. Этот фотохимический феномен лежит в основе темновой адаптации.
Особое значение в фотохимических процессах имеет
пигментный слой сетчатки, который образован эпителием, содержащим фусцин. Этот
пигмент поглощает свет, препятствуя отражению и рассеиванию его, что
обусловливает четкость зрительного восприятия. Отростки пигментных клеток окружают
светочувствительные членики палочек и колбочек, принимая участие в обмене
веществ фоторецепторов и в синтезе зрительных пигментов.
Вследствие фотохимических процессов в фоторецепторах
глаза при действии света возникает рецепторный потенциал, который представляет
собой гиперполяризацию мембраны рецептора. Это отличительная черта зрительных
рецепторов, активация других рецепторов выражается в виде деполяризации их
мембраны. Амплитуда зрительного рецепторного потенциала увеличивается при
увеличении интенсивности светового стимула. Так, при действии красного цвета,
длина волны которого составляет 620-760 нм, рецепторный потенциал более выражен
в фоторецепторах центральной части сетчатки, а синего (430-470 нм) - в
периферической.
Синаптические окончания фоторецепторов конвергируют на
биполярные нейроны сетчатки. При этом фоторецепторы центральной ямки связаны
только с одним биполяром.
Проводниковый отдел. Первый нейрон проводникового отдела
зрительного анализатора представлен биполярными клетками сетчатки (рис. 2.2).

Рис. 2.2 Схема строения сетчатки (по данным
электронной микроскопии)
Считают, что в биполярных клетках возникают потенциалы
действия подобно рецепторным и горизонтальным НС. В одних биполярах на включение
и выключение света возникает медленная длительная деполяризация, а в других -
на включение - гиперполяризация, на выключение - деполяризация.
Аксоны биполярных клеток в свою очередь конвергируют
на ганглиозные клетки (второй нейрон). В результате на каждую ганглиозную
клетку могут конвергировать около 140 палочек и 6 колбочек, при этом, чем ближе
к желтому пятну, тем меньше фоторецепторов конвергирует на одну клетку. В
области желтого пятна конвергенция почти не осуществляется и количество
колбочек почти равно количеству биполярных и ганглиозных клеток. Именно это
объясняет высокую остроту зрения в центральных отделах сетчатки.
Периферия сетчатки отличается большой
чувствительностью к слабому свету. Это обусловлено, по-видимому, тем, что до
600 палочек конвергируют здесь через биполярные клетки на одну и ту же
ганглиозную клетку. В результате сигналы от множества палочек суммируются и
вызывают более интенсивную стимуляцию этих клеток.
В ганглиозных клетках даже при полном затемнении
спонтанно генерируются серии импульсов с частотой 5 в секунду. Эта импульсация
обнаруживается при микроэлектродном исследовании одиночных зрительных волокон
или одиночных ганглиозных клеток, а в темноте воспринимается как «собственный
свет глаз».
В одних ганглиозных клетках учащение фоновых разрядов
происходит на включение света (on-ответ),
в других - на выключение света (off-ответ).
Реакция ганглиозной клетки может быть обусловлена и спектральным составом
света.
В сетчатке кроме вертикальных существуют также
латеральные связи. Латеральное взаимодействие рецепторов осуществляется
горизонтальными клетками. Биполярные и ганглиозньте клетки взаимодействуют
между собой за счет многочисленных латеральных связей, образованных
коллатералями дендритов и аксонов самих клеток, а также с помощью амакриновых
клеток.
Горизонтальные клетки сетчатки обеспечивают регуляцию
передачи импульсов между фоторецепторами и биполярами, регуляцию
цветовосприятия и адаптации глаза к различной освещенности. В течение всего
периода освещения горизонтальные клетки генерируют положительный потенциал -
медленную гиперполяризацию, названную S-потенциалом (от англ. slow
-- медленный). По характеру восприятия световых раздражений горизонтальные
клетки делят на два типа:
) L-тип,
в котором S-потенциал возникает при действии
любой волны видимого света;
) С-тип, или «цветовой», тип, в котором знак
отклонения потенциала зависит от длины волны. Так, красный свет может вызвать
их деполяризацию, а синий - гиперполяризацию.
Полагают, что сигналы горизонтальных клеток передаются
в электротонической форме.
Горизонтальные, а также амакриновые клетки называют
тормозными нейронами, так как они обеспечивают латеральное торможение между
биполярными или ганглиозными клетками.
Совокупность фоторецепторов, посылающих свои сигналы к
одной ганглиозной клетке, образует ее рецептивное поле. Вблизи желтого пятна
эти поля имеют диаметр 7-200 нм, а на периферии - 400-700 нм, т.е. в центре
сетчатки рецептивные поля маленькие, а на периферии сетчатки они значительно
больше по диаметру. Рецептивные поля сетчатки имеют округлую форму, построены
концентрически, каждое из них имеет возбудительный центр и тормозную
периферическую зону в виде кольца. Различают рецептивные поля с on-центром (возбуждаются при освещении
центра) и с off-центром (возбуждаются при затемнении
центра). Тормозная кайма, как предполагают в настоящее время, образуется
горизонтальными клетками сетчатки по механизму латерального торможения, т.е.
чем сильнее возбужден центр рецептивного поля, тем большее тормозное влияние он
оказывает на периферию. Благодаря таким типам рецептивных полей (РП)
ганглиозных клеток (с on- и off-центрами) происходит обнаружение
светлых и темных объектов в поле зрения уже на уровне сетчатки.
При наличии у животных цветового зрения выделяют
цветооппонентную организацию РП ганглиозных клеток сетчатки. Эта организация
состоит в том, что определенная ганглиозная клетка получает возбуждающие и
тормозные сигналы от колбочек, имеющих разную спектральную чувствительность.
Например, если «красные» колбочки оказывают возбуждающее действие на данную
ганглиозную клетку, то «синие» колбочки ее затормаживают. Обнаружены разные
комбинации возбуждающих и тормозящих входов от разных классов колбочек.
Значительная часть цветооппонентных ганглиозных клеток связаны со всеми тремя типами
колбочек. Благодаря такой организации РП отдельные ганглиозные клетки
становятся избирательными к освещению определенного спектрального состава. Так,
если возбуждение возникает от «красных» колбочек, то возбуждение сине- и
зеленочувствительных колбочек вызовет торможение этих клеток, а если
ганглиозная клетка возбуждается от синечувствительных колбочек, то она
тормозится от зелено- и красночувствительных и т.д.

Рис. 2.3 Электроретинограмма (по Граниту) a, b, с, d -
волны ЭРГ; стрелками указаны моменты включения и выключения света
Центр и периферия рецептивного поля имеют максимальную
чувствительность в противоположных концах спектра. Так, если центр рецептивного
поля отвечает изменением активности на включение красного света, то периферия
аналогичной реакцией отвечает на включение синего. Ряд ганглиозных клеток
сетчатки имеет так называемую дирекционную чувствительность. Она проявляется в
том, что при движении стимула в одном направлении (оптимальном) ганглиозная клетка
активируется, при другом направлении движения - реакция отсутствует.
Предполагают, что избирательность реакций этих клеток на движение в разных
направлениях создается горизонтальными клетками, имеющими вытянутые отростки
(теледендриты), с помощью которых направленно тормозятся ганглиозные клетки.
Вследствие конвергенции и латеральных взаимодействий рецептивные поля соседних
ганглиозных клеток перекрываются. Это обусловливает возможность суммации
эффектов световых воздействий и возникновение взаимных тормозных отношений в
сетчатке.
Электрические явления в сетчатке. В сетчатке глаза, где локализуется
рецепторный отдел зрительного анализатора и начинается проводниковый отдел, в
ответ на действие света происходят сложные электрохимические процессы, которые
можно зарегистрировать в виде суммарного ответа - электроретинограммы (ЭРГ)
(рис. 2.3).
ЭРГ отражает такие свойства светового раздражителя,
как цвет, интенсивность и длительность его действия. ЭРГ может быть
зарегистрирована от целого глаза или непосредственно от сетчатки. Для ее
получения один электрод помещают на поверхность роговой оболочки, а другой
прикладывают к коже лица вблизи глаза или на мочку уха.
На ЭРГ, зарегистрированной при освещении глаза,
различают несколько характерных волн. Первая негативная волна а представляет
собой небольшое по амплитуде электрическое колебание, отражающее возбуждение
фоторецепторов и горизонтальных клеток. Она быстро переходит в крутонарастающую
позитивную волну b, которая
возникает в результате возбуждения биполярных и амакриновых клеток. После волны
b наблюдается медленная
электроположительная волна с - результат возбуждения клеток пигментного
эпителия. С моментом прекращения светового раздражения связывают появление
электроположительной волны d.
Показатели ЭРГ широко используются в клинике глазных
болезней для диагностики и контроля лечения различных заболеваний глаза,
связанных с поражением сетчатки.
Проводниковый отдел, начинающийся в сетчатке (первый
нейрон - биполярный, второй нейрон - ганглиозные клетки), анатомически
представлен далее зрительными нервами и после частичного перекреста их волокон
- зрительными трактами. В каждом зрительном тракте содержатся нервные волокна,
идущие от внутренней (носовой) поверхности сетчатки глаза одноименной стороны и
от наружной половины сетчатки другого глаза. Волокна зрительного тракта
направляются к зрительному бугру (собственно таламус), к метаталамусу (наружные
коленчатые тела) и к ядрам подушки. Здесь расположены третий нейрон зрительного
анализатора. От них зрительные нервные волокна направляются в кору полушарий
большого мозга.
В наружных (или латеральных) коленчатых телах, куда
приходят волокна из сетчатки, есть рецептивные поля, которые также имеют
округлую форму, но меньше по размеру, чем в сетчатке. Ответы нейронов здесь
носят фазический характер, но более выражены, чем в сетчатке.
На уровне наружных коленчатых тел происходит процесс
взаимодействия афферентных сигналов, идущих от сетчатки глаза, с эфферентными
из области коркового отдела зрительного анализатора. С участием ретикулярной
формации здесь происходит взаимодействие со слуховой и другими сенсорными
системами, что обеспечивает процессы избирательного зрительного внимания путем
выделения наиболее существенных компонентов сенсорного сигнала.
Центральный, или корковый, отдел зрительного
анализатора расположен в затылочной доле (поля 17, 18, 19 по Бродману) или VI, V2, V3
(согласно принятой номенклатуре). Считают, что первичная проекционная область
(поле 17) осуществляет специализированную, но более сложную, чем в сетчатке и в
наружных коленчатых телах, переработку информации. Рецептивные поля нейронов
зрительной коры небольших размеров имеют вытянутые, почти прямоугольные, а не
округлые формы. Наряду с этим имеются сложные и сверхсложные рецептивные поля
детекторного типа. Эта особенность позволяет выделять из цельного изображения
лишь отдельные части линий с различным расположением и ориентацией, при этом
проявляется способность избирательно реагировать на эти фрагменты.
В каждом участке коры сконцентрированы нейроны,
которые образуют колонку, проходящую по глубине через все слои вертикально, при
этом происходит функциональное объединение нейронов, выполняющих сходную
функцию. Разные свойства зрительных объектов (цвет, форма, движение)
обрабатываются в разных частях зрительной коры большого мозга параллельно.
В зрительной коре существуют функционально различные
группы клеток - простые и сложные.
Простые клетки создают рецептивное поле, которое
состоит из возбудительной и тормозной зон. Определить это можно путем исследования
реакции клетки на маленькое световое пятно. Структуру рецептивного поля сложной
клетки установить таким путем невозможно. Эти клетки являются детекторами угла,
наклона и движения линий в поле зрения.
В одной колонке могут располагаться как простые, так и
сложные клетки. В III и IV слоях зрительной коры, где
заканчиваются таламические волокна, найдены простые клетки. Сложные клетки
расположены в более поверхностных слоях поля 17, в полях 18 и 19 зрительной
коры простые клетки являются исключением, там расположены сложные и
сверхсложные клетки.
В зрительной коре часть нейронов образует «простые»
или концентрические цветооппонентные рецептивные поля (IV слой). Цветовая оппонентность РП проявляется в том, что
нейрон, расположенный в центре, реагирует возбуждением на один цвет и
тормозится при стимуляции другого цвета. Одни нейроны реагируют on-ответом на красное освещение и ofT-ответом на зеленое, реакция других -
обратная.
У нейронов с концентрическими РП помимо оппонентных
отношений между цветоприемниками (колбочками) существуют антагонистические
отношения между центром и периферией, т.е. имеют место РП с двойной
цветооппонентностью. Например, если при воздействии на центр РП в нейроне
возникает on-ответ на красное и off-ответ на зеленое, то у него избирательность
к цвету сочетается с избирательностью к яркости соответствующего цвета, и он не
реагирует на диффузную стимуляцию светом волны любой длины (из-за оппонентных
отношений между центром и периферией РП).
От поля VI -
другой (дорзальный) канал проходит через средневисочную (медиотемпоральную -
МТ) область коры. Регистрация ответов нейронов этой области показала, что они
высокоселективны к диспаратности (неидентичности), скорости и направлению
движения объектов зрительного мира, хорошо реагируют на движение объектов на
текстурированном фоне. Локальное разрушение резко ухудшает способность
реагировать на движущиеся объекты, но через некоторое время эта способность
восстанавливается, свидетельствуя о том, что данная область не является
единственной зоной, где производится анализ движущихся объектов в зрительном
поле. Но наряду с этим предполагается, что информация, выделенная нейронами
первичного зрительного поля 17(V1),
далее передается для обработки во вторичную (поле V2) и третичную (поле V3) области зрительной коры.
Однако анализ зрительной информации не завершается в
полях стриарной (зрительной) коры (V1, V2, V3). Установлено, что от поля V1 начинаются пути (каналы) к другим областям, в которых
производится дальнейшая обработка зрительных сигналов.
Так, если разрушить у обезьяны поле V4, которое находится на стыке
височной и теменной областей, то нарушается восприятие цвета и формы. Обработка
зрительной информации о форме, как предполагают, происходит также и в основном
в нижневисочной области. При разрушении этой области базисные свойства
восприятия (острота зрения и восприятие света) не страдают, но выходят из строя
механизмы анализа высшего уровня.
Таким образом, в зрительной сенсорной системе
происходит усложнение рецептивных полей нейронов от уровня к уровню, и чем выше
синаптический уровень, тем строже ограничены функции отдельных нейронов.
В настоящее время зрительную систему, начиная с
ганглиозных клеток, разделяют на две функционально различные части (магна- и
парвоцеллюлярные). Это деление обусловлено тем, что в сетчатке млекопитающих
имеются ганглиозные клетки различных типов - X, Y, W. Эти клетки имеют концентрические
рецептивные поля, а их аксоны образуют зрительные нервы.
У X-клеток
- РП небольшое, с хорошо выраженной тормозной каймой, скорость проведения
возбуждения по их аксонам - 15-25 м/с. У Y-клеток центр РП гораздо больше, они лучше отвечают на
диффузные световые стимулы. Скорость проведения составляет 35-50 м/с. В
сетчатке X-клетки занимают центральную часть, а
к периферии их плотность снижается. Y-клетки распределены по сетчатке равномерно, поэтому на периферии
сетчатки плотность Y-клеток выше, чем
X-клеток. Особенности строения РП X-клеток обусловливают их лучшую
реакцию на медленные движения зрительного стимула, тогда как Y-клетки лучше реагируют на быстро
движущиеся стимулы.
В сетчатке описана также многочисленная группа W-клеток. Это самые мелкие ганглиозные
клетки, скорость проведения по их аксонам - 5-9 м/с. Клетки этой группы не
однородны. Среди них выделяются клетки с концентрическими и однородными РП и
клетки, которые чувствительны к движению стимула через рецептивное поле. При
этом реакция клетки не зависит от направления движения.
Разделение на X, Y и W системы продолжается и на уровне
коленчатого тела и зрительной коры. Нейроны X имеют фазический тип реакции (активация в виде короткой
вспышки импульсов), их рецептивные поля в большем количестве представлены в
периферических полях зрения, латентный период их реакции меньше. Такой набор
свойств показывает, что они возбуждаются быстропроводящими афферентами.
Нейроны X
имеют топический тип реакции (нейрон активируется в течение нескольких секунд),
их РП в большем количестве представлены в центре поля зрения, а латентный
период - больше.
Первичные и вторичные зоны зрительной коры (поля Y1 и Y2) различаются по содержанию Х- и Y-нейронов. Например, в поле Y1 от наружного коленчатого тела приходит афферентация как от
Х-, так и от Y-типов, тогда как поле Y2 получает афференты только от Y-типа клеток.
Изучение передачи сигналов на разных уровнях
зрительной сенсорной системы проводят, регистрируя суммарные вызванные
потенциалы (ВП) путем отведения у человека с помощью электродов от поверхности
кожи головы в области зрительной коры (затылочная область). У животных можно
одновременно исследовать вызываемую активность во всех отделах зрительной
сенсорной системы.
Механизмы,
обеспечивающие ясное видение в различных условиях
При рассмотрении объектов,
находящихся на разном удалении от наблюдателя, ясному видению способствуют следующие
процессы.
1. Конвергенционные и дивергенционные движения глаз, благодаря
которым осуществляется сведение или разведение зрительных осей. Если оба глаза
двигаются в одном направлении, такие движения называются содружественными.
2. Реакция зрачка, которая происходит синхронно с
движением глаз. Так, при конвергенции зрительных осей, когда рассматриваются
близко расположенные предметы, происходит сужение зрачка, т. е. конвергентная
реакция зрачков. Эта реакция способствует уменьшению искажения изображения,
вызываемого сферической аберрацией. Сферическая аберрация обусловлена тем, что
преломляющие среды глаза имеют неодинаковое фокусное расстояние в разных
участках. Центральная часть, через которую проходит оптическая ось, имеет
большее фокусное расстояние, чем периферическая часть. Поэтому изображение на
сетчатке получается нерезким. Чем меньше диаметр зрачка, тем меньше искажения,
вызываемые сферической аберрацией. Конвергентные сужения зрачка включают в
действие аппарат аккомодации, обусловливающий увеличение преломляющей силы
хрусталика.
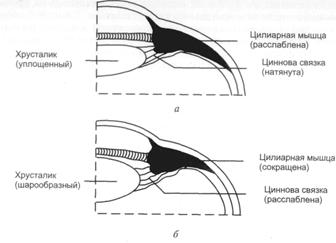
Рис. 2.4 Механизм аккомодации глаза: а - покой, б -
напряжение
Рис. 2.5 Схема хода лучей через преломляющие среды
глаза
Зрачок является также аппаратом устранения
хроматической аберрации, которая обусловлена тем, что оптический аппарат глаза,
как и простые линзы, преломляет свет с короткой волной сильнее, чем с длинной
волной. Исходя из этого, для более точной фокусировки предмета красного цвета
требуется большая степень аккомодации, чем для синего. Именно поэтому синие
предметы кажутся более удаленными, чем красные, будучи расположенными на одном
и том же расстоянии.
. Аккомодация является главным механизмом,
обеспечивающим ясное видение разноудаленных предметов, и сводится к
фокусированию изображения от далеко или близко расположенных предметов на
сетчатке. Основной механизм аккомодации заключается в непроизвольном изменении
кривизны хрусталика глаза (рис. 2.4).
Благодаря изменению кривизны хрусталика, особенно
передней поверхности, его преломляющая сила может меняться в пределах 10-14
диоптрий. Хрусталик заключен в капсулу, которая по краям (вдоль экватора
хрусталика) переходит в фиксирующую хрусталик связку (циннова связка), в свою
очередь, соединенную с волокнами ресничной (цилиарной) мышцы. При сокращении
цилиарной мышцы натяжение цинновых связок уменьшается, а хрусталик вследствие
своей эластичности становится более выпуклым. Преломляющая сила глаза
увеличивается, и глаз настраивается на видение близко расположенных предметов.
Когда человек смотрит вдаль, циннова связка находится в натянутом состоянии,
что приводит к растягиванию сумки хрусталика и его утолщению. Иннервация
цилиарной мышцы осуществляется симпатическими и парасимпатическими нервами.
Импульсация, поступающая по парасимпатическим волокнам глазодвигательного
нерва, вызывает сокращение мышцы. Симпатические волокна, отходящие от верхнего
шейного узла, вызывают ее расслабление. Изменение степени сокращения и
расслабления цилиарной мышцы связано с возбуждением сетчатки и находится под
влиянием коры головного мозга. Преломляющая сила глаза выражается в диоптриях
(Д). Одна диоптрия соответствует преломляющей силе линзы, главное фокусное
расстояние которой в воздухе равно 1 м. Если главное фокусное расстояние линзы
равно, например, 0,5 или 2 м, то ее преломляющая сила составляет,
соответственно, 2Д или 0,5Д. Преломляющая сила глаза без явления аккомодации
равна 58-60 Д и называется рефракцией глаза.
При нормальной рефракции глаза лучи от далеко
расположенных предметов после прохождения через светопреломляющую систему глаза
собираются в фокусе на сетчатке в центральной ямке. Нормальная рефракция глаза
носит название эмметропии, а такой глаз называют эмметропическим. Наряду с
нормальной рефракцией наблюдаются ее аномалии.
Миопия (близорукость) - это такой вид нарушения
рефракции, при котором лучи от предмета после прохождения через
светопреломляющий аппарат фокусируются не на сетчатке, а впереди нее. Это может
зависеть от большой преломляющей силы глаза или от большой длины глазного
яблока. Близкие предметы близорукий видит без аккомодации, отдаленные предметы
видит неясными, расплывчатыми. Для коррекции применяются очки с рассеивающими
двояковогнутыми линзами.
Гиперметропия (дальнозоркость) - вид нарушения рефракции,
при котором лучи от далеко расположенных предметов в силу слабой преломляющей
способности глаза или при малой длине глазного яблока фокусируются за
сетчаткой. Даже удаленные предметы дальнозоркий глаз видит с напряжением
аккомодации, вследствие чего развивается гипертрофия аккомодационных мышц. Для
коррекции применяют двояковыпуклые линзы.
Астигматизм - вид нарушения рефракции, при котором
лучи не могут сходиться в одной точке, в фокусе (от греч. stigme - точка), обусловлен различной
кривизной роговицы и хрусталика в различных меридианах (плоскостях). При
астигматизме предметы кажутся сплющенными или вытянутыми, его коррекцию
осуществляют сфероцилиндрическими линзами.
Следует отметить, что к светопреломляющей системе
глаза относятся также: роговица, влага передней камеры глаза, хрусталик и
стекловидное тело. Однако их преломляющая сила, в отличие от хрусталика, не
регулируется и в аккомодации участия не принимает. После прохождения лучей
через преломляющую систему глаза на сетчатке получается действительное,
уменьшенное и перевернутое изображение. Но в процессе индивидуального развития
сопоставление ощущений зрительного анализатора с ощущениями двигательного,
кожного, вестибулярного и других анализаторов, как отмечалось выше, приводит к
тому, что человек воспринимает внешний мир таким, какой он есть на самом деле.
Бинокулярное зрение (зрение двумя глазами) играет
важную роль в восприятии разноудаленных предметов и определении расстояния до
них, дает более выраженное ощущение глубины пространства по сравнению с
монокулярным зрением, т.е. зрением одним глазом. При рассматривании предмета
двумя глазами его изображение может попадать на симметричные (идентичные) точки
сетчаток обоих глаз, возбуждения от которых объединяются в корковом конце
анализатора в единое целое, давая при этом одно изображение. Если изображение
предмета попадает на неидентичные (диспаратные) участки сетчатки, то возникает
раздвоение изображения. Процесс зрительного анализа пространства зависит не
только от наличия бинокулярного зрения, существенную роль в этом играют
условно-рефлекторные взаимодействия, складывающиеся между зрительным и
двигательным анализаторами. Определенное значение имеют конвергенционные
движения глаз и процесс аккомодации, которые управляются по принципу обратных связей.
Восприятие пространства в целом связано с определением пространственных
отношений видимых предметов - их величины, формы, отношения друг к другу, что
обеспечивается взаимодействием различных отделов анализатора; значительную роль
при этом играет приобретенный опыт.
При движении объектов ясному видению способствуют следующие
факторы:
) произвольные движения глаз вверх, вниз, влево или
вправо со скоростью движения объекта, что осуществляется благодаря
содружественной деятельности глазодвигательных мышц;
) при появлении объекта в новом участке поля зрения
срабатывает фиксационный рефлекс - быстрое непроизвольное движение глаз,
обеспечивающее совмещение изображения предмета на сетчатке с центральной ямкой.
При слежении за движущимся объектом происходит медленное движение глаз -
следящее движение.
При рассматривании неподвижного
предмета для
обеспечения ясного видения глаз совершает три типа мелких непроизвольных
движений: тремор - дрожание глаза с небольшой амплитудой и частотой, дрейф -
медленное смещение глаза на довольно значительное расстояние и скачки (флики) -
быстрые движения глаз. Также существуют саккадические движения (саккады) -
содружественные движения обоих глаз, совершаемые с большой скоростью.
Наблюдаются саккады при чтении, просматривании картин, когда обследуемые точки
зрительного пространства находятся на одном удалении от наблюдателя и других
объектов. Если заблокировать эти движения глаз, то окружающий нас мир
вследствие адаптации рецепторов сетчатки станет трудно различимым, каким он является
у лягушки. Глаза лягушки неподвижны, поэтому она хорошо различает только
движущиеся предметы, например бабочек. Именно поэтому лягушка приближается к
змее, которая постоянно выбрасывает наружу свой язык. Находящуюся в состоянии
неподвижности змею лягушка не различает, а ее движущийся язык принимает за
летающую бабочку.
В условиях изменения освещенности ясное видение обеспечивают зрачковый
рефлекс, темновая и световая адаптация.
Зрачок регулирует интенсивность светового
потока, действующего на сетчатку, путем изменения своего диаметра. Ширина
зрачка может колебаться от 1,5 до 8,0 мм. Сужение зрачка (миоз) происходит при
увеличении освещенности, а также при рассматривании близко расположенного
предмета и во сне. Расширение зрачка (мидриаз) происходит при уменьшении
освещенности, а также при возбуждении рецепторов, любых афферентных нервов, при
эмоциональных реакциях напряжения, связанных с повышением тонуса симпатического
отдела нервной системы (боль, гнев, страх, радость и т.д.), при психических
возбуждениях (психозы, истерии и т.д.), при удушье, наркозе. Зрачковый рефлекс
при изменении освещенности хотя и улучшает зрительное восприятие (в темноте
расширяется, что увеличивает световой поток, падающий на сетчатку, на свету
сужается), однако главным механизмом все же является темновая и световая
адаптация.
Темповая адаптация выражается в повышении
чувствительности зрительного анализатора (сенситизация), световая адаптация -
в снижении чувствительности глаза к свету. Основу механизмов световой и
темновой адаптации составляют протекающие в колбочках и палочках фотохимические
процессы, которые обеспечивают расщепление (на свету) и ресинтез (в темноте)
фоточувствительных пигментов, а также процессы функциональной мобильности:
включение и выключение из деятельности рецепторных элементов сетчатки. Кроме
того, адаптацию определяют некоторые нейронные механизмы и, прежде всего,
процессы, происходящие в нервных элементах сетчатки, в частности способы
подключения фоторецепторов к ганглиозным клеткам с участием горизонтальных и
биполярных клеток. Так, в темноте возрастает число рецепторов, подключенных к
одной биполярной клетке, и большее их число конвергирует на ганглиозную клетку.
При этом расширяется рецептивное поле каждой биполярной и, естественно,
ганглиозной клеток, что улучшает зрительное восприятие. Включение же
горизонтальных клеток регулируется ЦНС.
Снижение тонуса симпатической нервной системы
(десимпатизация глаза) уменьшает скорость темновой адаптации, а введение
адреналина оказывает противоположный эффект. Раздражение ретикулярной формации
ствола мозга повышает частоту импульсов в волокнах зрительных нервов. Влияние
ЦНС на адаптивные процессы в сетчатке подтверждается также тем, что
чувствительность неосвещенного глаза к свету изменяется при освещении другого глаза
и при действии звуковых, обонятельных или вкусовых раздражителей.
Цветовая адаптация. Наиболее быстрая и резкая адаптация
(снижение чувствительности) происходит при действии сине-фиолетового
раздражителя. Красный раздражитель занимает среднее положение.
Зрительное восприятие крупных
объектов и их деталей обеспечивается за счет центрального и периферического зрения - изменений
угла зрения. Наиболее тонкая оценка мелких деталей предмета обеспечивается в
том случае, если изображение падает на желтое пятно, которое локализуется в
центральной ямке сетчатки глаза, так как в этом случае имеет место наибольшая
острота зрения. Это объясняется тем, что в области желтого пятна располагаются
только колбочки, их размеры наименьшие, и каждая колбочка контактирует с малым
числом нейронов, что повышает остроту зрения. Острота зрения определяется
наименьшим углом зрения, под которым глаз еще способен видеть отдельно две
точки. Нормальный глаз способен различать две светящиеся точки под углом зрения
в 1'. Острота зрения такого глаза принимается за единицу. Острота зрения
зависит от оптических свойств глаза, структурных особенностей сетчатки и работы
нейрональных механизмов проводникового и центрального отделов зрительного
анализатора. Определение остроты зрения осуществляется с помощью буквенных или
различного вида фигурных стандартных таблиц. Крупные объекты в целом и
окружающее пространство воспринимаются в основном за счет периферического
зрения, обеспечивающего большое поле зрения.
Поле зрения - пространство, которое можно видеть
фиксированным глазом. Различают отдельно поле зрения левого и правого глаз, а
также общее поле зрения для двух глаз. Величина поля зрения у людей зависит от
глубины положения глазного яблока и формы надбровных дуг и носа. Границы поля
зрения обозначаются величиной угла, образуемого зрительной осью глаза и лучом,
проведенным к крайней видимой точке через узловую точку глаза к сетчатке. Поле
зрения неодинаково в различных меридианах (направлениях). Книзу - 70°, кверху -
60°, кнаружи - 90°, кнутри - 55°. Ахроматическое поле зрения больше
хроматического в силу того, что на периферии сетчатки нет рецепторов,
воспринимающих цвет (колбочек). В свою очередь, цветное поле зрения неодинаково
для различных цветов. Самое узкое поле зрения для зеленого, желтого, больше для
красного, еще больше для синего цветов. Величина поля зрения изменяется в
зависимости от освещенности. Ахроматическое поле зрения в сумерках
увеличивается, на свету уменьшается. Хроматическое поле зрения, наоборот, на
свету увеличивается, в сумерках уменьшается. Это зависит от процессов
мобилизации и демобилизации фоторецепторов (функциональной мобильности). При
сумеречном зрении увеличение количества функционирующих палочек, т.е. их
мобилизация, приводит к увеличению ахроматического поля зрения, в то же самое
время уменьшение количества функционирующих колбочек (их демобилизация) ведет к
уменьшению хроматического поля зрения (П.Г. Снякин).
Зрительный анализатор имеет также механизм для различения
длины световой волны - цветовое зрение.
Цветовое
зрение, зрительные контрасты и последовательные образы
Цветовое зрение - способность зрительного анализатора
реагировать на изменения длины световой волны с формированием ощущения цвета.
Определенной длине волны электромагнитного излучения соответствует ощущение
определенного цвета. Так, ощущение красного цвета соответствует действию света
с длиной волны в 620-760 нм, а фиолетового - 390-450 нм, остальные цвета
спектра имеют промежуточные параметры. Смешение всех цветов дает ощущение
белого цвета. В результате смешения трех основных цветов спектра - красного,
зеленого, сине-фиолетового - в разном соотношении можно получить также
восприятие любых других цветов. Ощущение цветов связано с освещенностью. По
мере ее уменьшения сначала перестают различаться красные цвета, позднее всех -
синие. Восприятие цвета обусловлено в основном процессами, происходящими в
фоторецепторах. Наибольшим признанием пользуется трехкомпонентная теория
цветоощущения Ломоносова - Юнга - Гельмгольца-Лазарева, согласно которой в сетчатке
глаза имеются три вида фоторецепторов - колбочек, раздельно воспринимающих
красный, зеленый и сине-фиолетовые цвета. Комбинации возбуждения различных
колбочек приводят к ощущению различных цветов и оттенков. Равномерное
возбуждение трех видов колбочек дает ощущение белого цвета. Трехкомпонентная
теория цветового зрения получила свое подтверждение в электрофизиологических
исследованиях Р. Гранита (1947). Три типа цветочувствительных колбочек были
названы модуляторами, колбочки, которые возбуждались при изменении яркости
света (четвертый тип), были названы доминаторами. Впоследствии методом
микроспектрофотометрии удалось установить, что даже одиночная колбочка может
поглощать лучи различной длины волны. Обусловлено это наличием в каждой
колбочке различных пигментов, чувствительных к волнам света различной длины.
Несмотря на убедительные аргументы трехкомпонентной
теории в физиологии цветового зрения описаны факты, которые не находят
объяснения с этих позиций. Это дало возможность выдвинуть теорию противоположных,
или контрастных, цветов, т.е. создать так называемую оппонентную теорию
цветного зрения Эвальда Геринга.
Согласно этой теории, в глазу и/или в мозге существуют
три оппонентных процесса: один - для ощущения красного и зеленого, второй - для
ощущения желтого и синего, третий - качественно отличный от двух первых
процессов - для черного и белого. Эта теория применима для объяснения передачи
информации о цвете в последующих отделах зрительной системы: ганглиозных
клетках сетчатки, наружных коленчатых телах, корковых центрах зрения, где
функционируют цветооппонентные РП с их центром и периферией.
Таким образом, на основании полученных данных можно
полагать, что процессы в колбочках более соответствуют трехкомпонентной теории
цветоощущения, тогда как для нейронных сетей сетчатки и вышележащих зрительных
центров подходит теория контрастных цветов Геринга.
В восприятии цвета определенную роль играют и
процессы, протекающие в нейронах разных уровней зрительного анализатора
(включая сетчатку), которые получили название цветооппонентных нейронов. При
действии на глаз излучений одной части спектра они возбуждаются, а другой -
тормозятся. Такие нейроны участвуют в кодировании информации о цвете.
Наблюдаются аномалии цветового зрения, которые могут
проявляться в виде частичной или полной цветовой слепоты. Людей, вообще не
различающих цвета, называют ахроматами. Частичная цветовая слепота имеет место
у 8-10% мужчин и 0,5% женщин. Полагают, что цветослепота связана с отсутствием
у мужчин определенных генов в половой непарной Х-хромосоме. Различаются три
вида частичной цветослепоты: протанопия (дальтонизм) - слепота в
основном на красный цвет. Этот вид цветослепоты впервые был описан в 1794 году
физиком Дж. Дальтоном, у которого наблюдался этот вид аномалии. Людей с таким видом
аномалии называют «краснослепыми»; дейтеранопия - понижение восприятия
зеленого цвета. Таких людей называют «зеленослепыми»; тританопия - редко
встречающаяся аномалия. При этом люди не воспринимают синий и фиолетовый цвета,
их называют «фиолетовослепыми».
С точки зрения трехкомпонентной теории цветового
зрения каждый из видов аномалии является результатом отсутствия одного из трех
колбочковых цветовоспринимающих субстратов. Для диагностики расстройства
цветоощущения пользуются цветными таблицами Е. Б. Рабкина, а также специальными
приборами, получившими название аномалоскопов. Выявление различных
аномалий цветового зрения имеет большое значение при определении
профессиональной пригодности человека для различных видов работ (шофера,
летчика, художника и др.).
Возможность оценки длины световой волны, проявляющаяся
в способности к цветоощущению, играет существенную роль в жизни человека,
оказывая влияние на эмоциональную сферу и деятельность различных систем
организма. Красный цвет вызывает ощущение тепла, действует возбуждающе на
психику, усиливает эмоции, но быстро утомляет, приводит к напряжению мышц,
повышению артериального давления, учащению дыхания. Оранжевый цвет вызывает
чувство веселья и благополучия, способствует пищеварению. Желтый цвет создает хорошее,
приподнятое настроение, стимулирует зрение и нервную систему. Это самый
«веселый» цвет. Зеленый цвет действует освежающе и успокаивающе, полезен при
бессоннице, переутомлении, понижает артериальное давление, общий тонус
организма и является самым благоприятным для человека. Голубой цвет вызывает
ощущение прохлады и действует на нервную систему успокаивающе, причем сильнее
зеленого (особенно благоприятен голубой цвет для людей с повышенной нервной
возбудимостью), больше, чем при зеленом цвете, понижает артериальное давление и
тонус мышц. Фиолетовый цвет не столько успокаивает, сколько расслабляет
психику. Создается впечатление, что человеческая психика, следуя вдоль спектра
от красного к фиолетовому, проходит всю гамму эмоций. На этом основано использование
теста Люшера для определения эмоционального состояния организма.
Зрительные контрасты и
последовательные образы. Зрительные ощущения могут продолжаться и после того, как прекратилось
раздражение. Такое явление получило название последовательных образов.
Зрительные контрасты - это измененное восприятие раздражителя в зависимости от
окружающего светового или цветового фона. Существуют понятия светового и
цветового зрительных контрастов. Явление контраста может проявляться в
преувеличении действительной разницы между двумя одновременными или
последовательными ощущениями, поэтому различают одновременные и
последовательные контрасты. Серая полоска на белом фоне кажется темнее такой же
полоски, расположенной на темном фоне. Это пример одновременного светового
контраста. Если рассматривать серый цвет на красном фоне, то он кажется
зеленоватым, а если рассматривать серый цвет на синем фоне, то он приобретает
желтый оттенок. Это явление одновременного цветового контраста.
Последовательный цветовой контраст заключается в изменении цветового ощущения
при переводе взгляда на белый фон. Так, если долго смотреть на окрашенную в
красный цвет поверхность, а затем перевести взор на белую, то она приобретает
зеленоватый оттенок. Причиной зрительного контраста являются процессы, которые
осуществляются в фоторецепторном и нейрональном аппаратах сетчатки. Основу
составляет взаимное торможение клеток, относящихся к разным рецептивным полям
сетчатки и их проекциям в корковом отделе анализаторов.
Список литературы:
Смирнов В.М.,
Будылина С.М. Физиология сенсорных систем и высшая нервная деятельность