Механизм действия ферментов
Содержание
Введение
1. Типы ферментов
2. Структура ферментов
. Механизм действия ферментов
Библиографический список
Введение
Ферменты - наиболее важный класс белковых веществ, универсальный по своей
биологической функции. Ферменты представляют собой специфические и
высокоэффективные катализаторы химических реакций, протекающих в живой клетке.
Изучение ферментов, их строения, свойств и механизма биологического действия
составляет один из главных разделов биохимии и биоорганической химии. К
настоящему времени охарактеризовано несколько тысяч ферментов, свыше тысячи из
них получены в индивидуальном состоянии. Для многих сотен белков-ферментов
выяснена аминокислотная последовательность, а самые известные из них
расшифрованы с помощью рентгеноструктурного анализа до уровня полной
пространственной структуры. Изучение любой проблемы в области познания
механизмов жизнедеятельности обязательно связано с исследованием
соответствующих ферментных систем. Кроме того, ферменты широко используются как
мощные инструменты при выяснении строения биополимеров и при генно-инженерных
разработках. Они находят широкое практическое применение в медицине и пищевой
промышленности.
Ферментативные процессы известны человеку с глубокой древности. В
частности, брожение широко использовалось греками для получения вина (открытие
этого способа приписывалось богу Бахусу). Народы многих стран издавна владели
искусством приготовления хлеба, сыра, уксуса на основе переработки
растительного и животного сырья. Однако современный этап в развитии энзимологии
относится к началу прошлого века. В 1814 г. член Петербургской Академии наук К.
Кирхгоф установил, что крахмал превращается в сахар под действием некоторых
веществ, находящихся в прорастающих зернах ячменя. Дальнейший шаг вперед в этом
направлении был сделан французскими химиками А. Пайеном и Ж. Пирсо, которые в
1833 г. показали, что термолабильный фактор, получаемый из солодового экстракта
путем осаждения спиртом, обладает способностью гидролизовать крахмал; они
назвали его диастазой.
Вскоре разгорелся спор о природе брожения, в котором участвовали
крупнейшие представители естествознания того времени. В частности, Л. Пастер
придерживался мнения, что брожение вызывается живыми микроорганизмами и,
следовательно, связано исключительно с их жизнедеятельностью. С другой стороны,
Ю. Либих и К. Бернар отстаивали химическую природу брожения, считая, что оно
связано с особыми веществами, подобными диастазе (амилазе). Й. Берцелиус в 1837
г. показал, что ферменты - это катализаторы, поставляемые живыми клетками.
Именно тогда появились термины «фермент» (от лат. fermentatio- брожение) и
«энзим» (от греч. - в дрожжах). Спор был окончательно разрешен лишь в 1897 г.,
когда немецкие ученые братья Ганс и Эдвард Бухнеры показали, что дрожжевой
бесклеточный сок (полученный при растирании дрожжей с инфузорной землей)
способен сбраживать сахар с образованием спирта и СО2. Стало ясным,
что дрожжевой сок содержит сложную смесь ферментов (названную зимазой) и эти
ферменты способны функционировать как внутри,
так и вне клеток. По словам одного из историков, появление пузырьков углекислого газа в опыте Бухнеров означало рождение
современных биохимии и энзимологии.
Попытки выделить ферменты в индивидуальном состоянии предпринимали многие
исследователи, среди которых следует упомянуть А. Я. Данилевского, Р.
Вильштеттера и др. Белковая природа ферментов была однозначно доказана в 1926
г. американским биохимиком Дж. Самнером, выделившим в кристаллическом виде
фермент уреазу из семян канавалии. В 1930 г. Дж. Нортроп получил кристаллический
пепсин, а затем трипсин и химотрипсин. С этого периода стало общепринятым
утверждение, что все ферменты являются белками.
В конце XIX в. на базе достижений в области исследования структуры
органических соединений биологического происхождения появилась возможность
изучения специфичности ферментов. В это время Э. Фишером было выдвинуто
знаменитое положение о необходимости стерического соответствия между ферментом
и субстратом; по его образному выражению, «субстрат подходит к ферменту, как
ключ к замку». В начале XX
века были заложены основы
исследования кинетики действия ферментов.
Ферменты имеют различные молекулярные массы - от 10 000 до 1000 000 и
выше. Они могут быть построены из одной полипепидной цепи, нескольких
полипептидных цепей или представлять собой сложные (иногда полиферментные)
комплексы. В состав фермента входят и небелковые компоненты, получившие
название коферентов (кофакторов), - ионы металлов, небольшие органические
молекулы типа витаминов и т.д.
Ферменты являются высокоэффективными катализаторами: они способны
увеличивать скорости реакции в миллионы и миллиарды раз. Так, например, уреаза
(при рН 8,0, 20 0С) ускоряет гидролиз мочевины примерно в 1014
раз.
Ферменты являются высокоспецифичными катализаторами. Они проявляют специфичность
в отношении типа катализируемой химической реакции, причем образования побочных
продуктов не происходит. Кроме того, они обладают выраженной субстратной
специфичностью и, как правило, высокой стереоспецифичностью.
1. Типы ферментов
Классификация ферментов. Ранее при наименовании ферментов за основу брали
название субстрата с добавлением суффикса «аза»; так появились, в частности,
протеиназы, липазы, карбогидразы. По исходному принципу обозначали ферменты,
катализирующие окислительные реакции (дегидрогеназы). Некоторые ферменты
получили специальные названия - трипсин, пепсин и др. В настоящее время принята
классификация, в которой ферменты сгруппированы в 6 классов в соответствии с
типом катализируемых реакций:
. Оксидоредуктазы (окислительно-восстановительные реакции).
. Трансферазы (реакции переноса функциональных групп) .
. Гидролазы (реакции гидролиза).
. Лиазы (реакции отщепления групп негидролитическим путем) .
. Изомеразы (реакции изомеризации).
. Лигазы (реакции синтеза за счет энергии АТР).
В пределах классов ферменты группируются в подклассы и подклассы в
соответствии с особенностями катализируемых реакций; на этой основе составлена
кодовая нумерация (шифры) ферментов и их систематические названия. Шифр
фермента состоит из четырех разделенных точками чисел: первое число означает
класс фермента, второе и третье числа - подкласс и подподкласс соответственно,
а четвертое число - порядковый номер фермента в его подподклассе. Например,
кислая фосфатаза имеет шифр 3.1.3.2; это означает, что она относится к классу
гидролаз (3.1.3.2), подклассу этих ферментов, действующих на сложноэфирные
связи (3.1.3.2), к подподклассу ферментов, гидролизующих моноэфиры фосфорной
кислоты (3.1.3.2), а порядковый номер фермента в данном подподклассе - 2 (3.1.3.2).
Ферменты, катализирующие одну и ту же реакцию, но выделенные из разных
видов живых организмов, различаются между собой. В номенклатуре же они имеют
общее название и один кодовый номер. Различные формы того или иного фермента
нередко встречаются и у одного биологического вида. Для наименования группы
ферментов, катализирующих одну и ту же реакцию и находящихся в организмах
одного вида, рекомендуется термин множественные формы фермента. Для тех
ферментов одной группы, которые имеют генетически обусловленные различия в
первичной структуре, используют термин «изоферменты».
Оксидоредукта́зы - отдельный класс ферментов, катализирующих лежащие
в основе биологического окисления реакции, сопровождающиеся переносом
электронов с одной молекулы (восстановителя - акцептора протонов или донора
электронов) на другую (окислитель - донора протонов или акцептора электронов).
Реакции, катализируемые оксидоредуктазами, в общем виде выглядят так:
− + B → A + B−
Где A - восстановитель (донор электронов), а B - окислитель (акцептор
электронов)
В биохимических превращениях окислительно-восстановительные реакции
иногда выглядят сложнее. Вот, например, одна из реакций гликолиза:
н
+ глицеральдегид-3-фосфат + НАД+ → НАД · H + H+ +
1,3-дифосфоглицерат
Здесь в качестве окислителя выступает НАД+, а
глицеральдегид-3-фосфат является восстановителем.
Систематические названия ферментов класса образуются по схеме «донор:
акцептор + оксидоредуктаза». Однако широко используются и другие схемы
именования. Когда возможно, ферменты называют в виде «донор + дегидрогеназа»,
например глицеральдегид-3-фосфатдегидрогеназа, для второй реакции выше. Иногда
название записывается как «акцептор + редуктаза», например НАД+-редуктаза.
В частном случае, когда окислителем является кислород, название может быть в
виде «донор + оксидаза».
Согласно международной классификации и номенклатуре ферментов
оксидоредуктазы относятся к 1 классу, в пределах которого выделяют двадцать два
подкласса:
КФ 1.1 включает ферменты, взаимодействующие с CH-OH группой доноров;
КФ 1.2 включает ферменты, взаимодействующие с альдегидной или
оксо-группой доноров;
КФ 1.3 включает ферменты, взаимодействующие с CH-CH группой доноров;
КФ 1.4 включает ферменты, взаимодействующие с CH-NH2 группой
доноров;
КФ 1.5 включает ферменты, взаимодействующие с CH-NH группой доноров;
КФ 1.6 включает ферменты, взаимодействующие с НАД · H или НАДФ · H;
КФ 1.7 включает ферменты, взаимодействующие с другими азотосодержащими
соединениями в качестве доноров;
КФ 1.8 включает ферменты, взаимодействующие с сероусодержащей группой
доноров;
КФ 1.9 включает ферменты, взаимодействующие с гемовой группой доноров;
КФ 1.10 включает ферменты, взаимодействующие с дифенолами и родственными
соединениями в качестве доноров;
КФ 1.11 включает ферменты, взаимодействующие с пероксидом в качестве
акцептора (пероксидазы);
КФ 1.12 включает ферменты, взаимодействующие с водородом в качестве
донора;
КФ 1.13 включает ферменты, взаимодействующие с одиночными донорами со
встраиванием молекулярного кислорода (оксигеназы);
КФ 1.14 включает ферменты, взаимодействующие с парными донорами со
встраиванием молекулярного кислорода;
КФ 1.15 включает ферменты, взаимодействующие с супероксид-радикалами в
качестве акцепторов;
КФ 1.16 включает ферменты, окисляющие ионы металлов;
КФ 1.17 включает ферменты, взаимодействующие с CH или CH2
группами;
КФ 1.18 включает ферменты, взаимодействующие с железосерными белками в
качестве доноров;
КФ 1.19 включает ферменты, взаимодействующие с восстановленным
флаводоксином в качестве донора;
КФ 1.20 включает ферменты, взаимодействующие с фосфором или мышьяком в
качестве донора;
КФ 1.21 включает ферменты, взаимодействующие с молекулами вида X-H и Y-H
с образованием связи X-Y;
КФ 1.97 включает другие оксидоредуктазы.
Трансфера́зы - отдельный класс ферментов, катализирующих перенос
функциональных групп и молекулярных остатков от одной молекулы к другой. Широко
распространены в растительных и животных организмах, участвуют в превращениях
углеводов, липидов, нуклеиновых и аминокислот.
Реакции, катализируемые трансферазами, в общем случае выглядят так:
X + B ↔ A + B-X.
Молекула A в здесь выступает в качестве донора группы атомов (X), а
молекула B является акцептором группы. Часто в качестве донора в подобных
реакциях переноса выступает один из коферментов. Многие из катализируемых
трансферазами реакций являются обратимыми.
Систематические названия ферментов класса образуются по схеме:
«донор:акцептор + группа + трансфераза».
Или же используются чуть более общие названия, когда в название фермента
включается имя либо донора, либо акцептора группы:
«донор + группа + трансфераза» или «акцептор + группа + трансфераза».
Например, аспартатаминотрансфераза катализирует перенос аминной группы с
молекулы аспарагиновой кислоты, катехол-О-метилтрансфераза осуществляет перенос
метильной группы S-аденозилметионина на бензольное кольцо различных
катехоламинов, а гистон-ацетилтрансфераза переносит ацетильную группу с
ацетил-кофермента А на гистон в процессе активации транскрипции.
Кроме того ферменты 7 подгруппы трансфераз, переносящие остаток фосфорной
кислоты, используя в качестве донора фосфатной группы АТФ, часто называют также
киназами; аминотрансферазы (6 подгруппа) часто называют трансаминазами.
Согласно международной классификации и номенклатуре ферментов трансферазы
относятся ко 2 классу, в пределах которого выделяют девять подклассов:
КФ 2.1 включает ферменты, переносящие одноуглеродные группы;
КФ 2.2 - ферменты, переносящие альдегидные и кетонные группы;
КФ 2.3 - переносящие ацильные остатки (ацилтрансферазы);
КФ 2.4 - переносящие остатки сахаров (гликозилтрансферазы);
КФ 2.6 - переносящие группы атомов, содержащие азот;
КФ 2.7 - переносящие фосфор-содержащие остатки;
КФ 2.8 - переносящие группы, содержащие серу;
КФ 2.9 - переносящие группы, содержащие селен.
Гидролазы - это класс ферментов, катализирующий гидролиз ковалентной
связи. Общий вид реакции, катализируемой гидролазой выглядит следующим образом:
B + H2O → A-OH + B-H
Систематическое название гидролаз включает название расщепляемого
субстрата с последующим добавлением гидролаза. Однако, как правило в
тривиальном названии слово гидролаза опускается и остаётся только суффикс
«-аза».
КФ 3.1 сложноэфирная связьэстеразы: нуклеаза, фосфодиэстераза, липаза,
фосфатаза
КФ 3.2 сахарагликозидазы: амилаза, гиалуронидаза, лизоцим и др.
КФ 3.3 простая эфирная связь
КФ 3.4 пептидная связьпротеазы: трипсин, химотрипсин, эластаза, тромбин,
ренин и др.
КФ 3.5 непептидная углерод-азотная связь
КФ 3.6 кислотный ангидридкислотный ангидрид-гидролаза (хеликаза, ГТФаза)
КФ 3.7 углерод-углеродная связь (C-C)
КФ 3.8 галогенная связь
КФ 3.9 азотно-фосфорная связь (P-N)
КФ 3.10 азотно-серная связь (S-N)
КФ 3.11 углеродно-фосфорная связь (C-P)
КФ 3.12 дисульфидная связь (S-S)
КФ 3.13 серо-углеродная связь (C-S)
Лиа́зы (синтазы) - отдельный класс ферментов, катализирующих
реакции негидролитического и неокислительного разрыва различных химических
связей (C-C, C-O, C-N, C-S и других) субстрата, обратимые реакции образования и
разрыва двойных связей, сопровождающиеся отщеплением или присоединением групп
атомов по её месту, а также образованием циклических структур.
В общем виде названия ферментов образуются по схеме «субстрат + лиаза».
Однако чаще в названии учитывают подкласс фермента. Лиазы отличаются от других
ферментов тем, что в катализируемых реакциях в одном направлении участвуют два
субстрата, а в обратной реакции только один. В названии фермента присутствуют
слова "декарбоксилаза" и "альдолаза" или "лиаза"
(пируват-декарбоксилаза, оксалат-декарбоксилаза, оксалоацетат-декарбоксилаза,
треонин-альдолаза, фенилсерин-альдолаза, изоцитрат-лиаза, аланин-лиаза,
АТФ-цитрат-лиаза и др.), а для ферментов, катализирующих реакции отщепления
воды от субстрата - "дегидратаза" (карбонат-дегидратаза,
цитрат-дегидратаза, серин-дегидратаза и др.). В тех случаях, когда обнаружена
только обратная реакция, или это направление в реакциях более существенно, в
названии ферментов пристутствует слово "синтаза" (малат-синтаза,
2-изопропилмалат-синтаза, цитрат-синтаза, гидроксиметилглутарил-CoA-синтаза и
др.).
Примеры: гистидиндекарбоксилаза, фумаратгидратаза.
По международной классификации и номенклатуре ферментов лиазы принадлежат
к 4 классу, в пределах которого выделяют семь подклассов:
КФ 4.1 включает ферменты, которые расщепляют углерод-углеродные связи,
например, декарбоксилазы (карбокси-лиазы);
КФ 4.2 - ферменты, расщепляющие углерод-кислородные связи, например,
дегидратазы;
КФ 4.3 - ферменты, расщепляющие углерод-азотные связи (амидин-лиазы);
КФ 4.4 - ферменты, расщепляющие углерод-серные связи;
КФ 4.5 - включает ферменты, расщепляющие связи углерод - галоген,
например, ДДТ-дегидрохлориназа;
КФ 4.6 - ферменты, расщепляющие фосфор-кислородные связи, например,
аденилатциклаза;
КФ 4.99 - включает другие лиазы
Изомеразы - ферменты, катализирующие структурные превращения изомеров
(рацемизация или эпимеризация). Изомеразы катализируют реакции, подобные
следующей:→ B, где B является изомером A.
В названии фермента присутствует слово "рацемаза"
(аланин-рацемаза, метионин-рацемаза, гидроксипролин-рацемаза, лактат-рацемаза и
др.), "эпимераза" (альдоза-1-эпимераза, рибулозофосфат-4-эпимераза,
УДФ-глюкуронат-4-эпимераза и др.), "изомераза"
(рибозофосфат-изомераза, ксилозоизомераза, глюкозаминфосфат-изомераза, эноил-СоА
изомераза и др.), "мутаза" (фосфоглицерат-мутаза,
метиласпартат-мутаза, фосфоглюкомутаза и др.).
Изомеразы имеют собственную классификацию, КФ 5 и имеют следующие
подклассы:
КФ 5.1 включает ферменты, катализирующие рацемизацию (рацемазы) и
эпимеризацию (эпимеразы)
КФ 5.2 включает ферменты, катализирущие геометрическую изомеризацию
(цис-транс изомеразы)
КФ 5.3 включает внутримолекулярные оксидоредуктазы
КФ 5.4 включает трансферазы (мутазы)
КФ 5.5 включает внутримолекулярные лиазы
КФ 5.99 включает другие изомеразы, в том числе, топоизомеразы
Лигазы (синтетазы). К классу лигаз относят ферменты, катализирующие
синтез органических веществ из двух исходных молекул с использованием энергии
распада АТФ (или другого нуклеозидтрифосфата). Систематическое название их
составляют по форме «X : Y лигаза», где X и Y обозначают исходные вещества. В
качестве примера можно назвать L-глутамат: аммиак лигазу (рекомендуемое
сокращенное название «глутаминсинтета-за»), при участии которой из глутаминовой
кислоты и аммиака в присутствии АТФ синтезируется глутамин.
Лигазы подразделяют по типу связи, образование которой они
катализируют:O-лигазаS-лигазаN-лигазаC-лигаза
. Структура ферментов
В природе существуют как простые, так и сложные ферменты. Первые целиком
представлены полипептидными цепями и при гидролизе распадаются исключительно на
аминокислоты. Такими ферментами (простые белки) являются гидролитические
ферменты, в частности пепсин, трипсин, папаин, уреаза, лизоцим, рибонуклеаза,
фосфатаза и др. Большинство природных ферментов относится к классу сложных
белков, содержащих, помимо полипептидных цепей, какой-либо небелковый компонент
(кофактор), присутствие которого является абсолютно необходимым для
каталитической активности. Кофакторы могут иметь различную химическую природу и
различаться по прочности связи с полипептидной цепью. Если константа
диссоциации сложного фермента настолько мала, что в растворе все полипептидные
цепи оказываются связанными со своими кофакторами и не разделяются при
выделении и очистке, то такой фермент получает название холофермента
(холоэнзим), а кофактор - простетической группы, рассматривающейся как
интегральная часть молекулы фермента. Полипептидную часть фермента принято
называть апоферментом.
В литературе до сих пор употребляются и другие наименования компонентов
сложных ферментов, в частности «фермент-протеид», «белковый компонент»
(апофермент), «кофермент» (коэнзим) и «простетическая группа». Под коферментом
часто подразумевают дополнительную группу, легко отделяемую от апофермента при
диссоциации. Предполагают, что простетическая группа может быть связана с
белком ковалентными и нековалентными связями. Так, в молекуле
ацетилкоэнзим-А-карбоксилазы кофактор биотин ковалентно связан с апоферментом
посредством амидной связи. С другой стороны, химические связи между кофакторами
и пептидными цепями могут быть относительно слабыми (например, водородные
связи, электростатические взаимодействия и др.). В таких случаях при выделении
ферментов наблюдается полная диссоциация обеих частей, и изолированый белковый
компонент оказывается лишенным ферментативной активности, пока не будет
добавлен извне недостающий кофактор. Именно к подобным изолированным
низкомолекулярным органическим веществам применим термин «кофермент», типичными
представителями которых являются витамины В1, В2, В6, РР, содержащие
коферменты. Известно также, что и простетические группы, и коферменты активно
включаются в химические реакции, выполняя функции промежутоных переносчиков
электронов, атомов водорода или различных функциональных групп (например,
аминных, ацетильных, карбоксильных). В подобных случаях кофермент рассматривают
в качестве второго субстрата, или косубстрата.
Роль кофермента (Ко) в качестве переносчика, например, атомов водорода
может быть представлена в виде схемы, где SH - субстрат, КоЕ - холофермент, А -
акцептор протона:
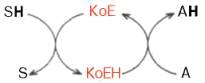
Субстрат подвергается окислению, отдавая электроны и протоны, а КоЕ -
восстановлению, принимая электроны и протоны. В следующей полуреакции
восстановленный КоЕН может отдавать электроны и протоны на какой-либо другой промежуточный
переносчик электронов и протонов или на конечный акцептор.
Коэнзим, кофактор, простетическая группа - двусмысленный биохимический
жаргон. До сих пор продолжается терминологический спор, поскольку часто
определения «коэнзим», «кофактор» и «простетическая группа» рассматриваются
через призму их роли в реакциях энзиматического (ферментативного) катализа.
Следует, однако, считаться с тем неоспоримым фактом, что во многих случаях
небелковые органические молекулы, как и ионы металлов, абсолютно необходимы
белковому компоненту при выполнении определенной биологической функции, не
имеющей отношения к биокатализу. Несомненно, имеют значение также тип и
характер связи небелкового компонента с молекулой белка. Поэтому очевидно, что
кофактором может служить любой фактор, абсолютно необходимый для выполнения
белком его каталитической или любой другой биологической роли. С другой
стороны, коферментом может быть любой небелковый фактор, который
непосредственно вовлечен в реакцию энзиматического катализа. Кофактор, который
непосредственно не участвует в акте катализа, не является коэнзимом. В то же
время простетическую группу (ковалентно связанный небелковый компонент,
необходимый для определенной функции) можно назвать коферментом, если она
непосредственно участвует в энзиматической реакции. Простетическая группа,
которая не вовлечена в акт катализа, но функционально является существенным как
для фермента, так и для некаталитического белка, может быть названа кофактором.
И наконец, кофактор и кофермент, непрочно связанные (или слабо связанные) с
ферментом или белком, тем не менее, не классифицируются в качестве
простетических групп.
Многие двухвалентные металлы (Mg2+, Мn2+, Са2+)
также выполняют роль кофакторов, хотя они не относятся ни к коферментам, ни к
простетическим группам. Известны примеры, когда ионы металлов прочно связаны с
белковой молекулой, выполняя функции простетической группы. В частности,
очищенный фермент, катализирующий окисление аскорбиновой кислоты (витамин С) в
дезоксиаскорбиновую кислоту, содержит 8 атомов меди на одну молекулу; все они
настолько прочно связаны с белковой молекулой, что даже не обмениваются с
ионообменными смолами и не отделяются методом диализа. Более того, с помощью
метода электронного парамагнитного резонанса показано участие ионов меди в
промежуточном переносе электронов. Интересно отметить, что свободные ионы меди
также наделены каталитической активностью при окислении аскорбиновой кислоты,
однако эта активность повышается во многие тысячи раз, если ионы меди
соединяются с апофер-ментом в единый комплекс - холофермент.
Получены доказательства кофакторной функции в ферментативных реакциях и
ряда других биологически активных соединений, не относящихся к витаминам:
HS-глутатиона, АТФ, липоевой кислоты, производных нуклеозидов (уридинфосфат,
цитидинфосфат, фосфоаденозинфосфосульфат), порфиринсодержащих веществ и др.
Сюда же могут быть отнесены тРНК, которые в составе ферментов
аминоацил-тРНК-синтетаз принимают активное участие в транспорте аминокислот в
рибосоме, где осуществляется синтез белка.
Следует отметить одну отличительную особенность двухкомпонентных
ферментов: ни кофактор отдельно (включая большинство коферментов), ни сам по
себе апофермент каталитической активностью не наделены, и только их объединение
в одно целое, протекающее не хаотично, а в соответствии с программой их
структурной организации, обеспечивает быстрое протекание химической реакции.
Активный центр ферментов.
При изучении механизма химической реакции, катализируемой ферментами,
исследователя всегда интересует не только определение промежуточных и конечных
продуктов и выяснение отдельных стадий реакции, но и природа тех функциональных
групп в молекуле фермента, которые обеспечивают специфичность действия фермента
на данный субстрат (субстраты) и высокую каталитическую активность. Речь идет,
следовательно, о точном знании геометрии и третичной структуры фермента, а
также химической природы того участка (участков) молекулы фермента, который
обеспечивает высокую скорость каталитической реакции. Участвующие в ферментативных
реакциях молекулы субстратов часто имеют небольшие размеры по сравнению с
молекулами ферментов, поэтому было высказано предположение, что при образовании
фермент-субстратных комплексов в непосредственный контакт с молекулой
субстрата, очевидно, вступает ограниченная часть аминокислот пептидной цепи.
Отсюда возникло представление об активном центре фермента. Под активным центром
подразумевают уникальную комбинацию аминокислотных остатков в молекуле
фермента, обеспечивающую непосредственное связывание ее с молекулой субстрата и
прямое участие в акте катализа. Установлено, что у сложных ферментов в состав
активного центра входят также простетические группы.
В активном центре условно различают так называемый каталитический центр,
непосредственно вступающий в химическое взаимодействие с субстратом, и
связывающий центр, или контактную («якорную») площадку, которая обеспечивает
специфическое сродство к субстрату и формирование его комплекса с ферментом. В
свою очередь молекула субстрата также содержит функционально различные участки:
например, субстраты эстераз или протеиназ - одну специфическую связь (или
группу атомов), подвергающуюся атаке со стороны фермента, и один или несколько
участков, избирательно связываемых ферментом.
Получены экспериментальные доказательства наличия в активном центре
химотрипсина двух остатков гистидина и остатка серина, схематически
представленных в трехмерной структурной модели предшественника этого фермента.
Выявление химической природы и вероятной топографии групп активного центра -
проблема первостепенной важности. Она сводится к определению природы
аминокислот, их последовательности и взаиморасположения в активном центре. Для
идентификации так называемых существенных аминокислотных остатков используют
специфические ингибиторы ферментов (часто это субстратподобные вещества или
аналоги коферментов), методы «мягкого» (ограниченного) гидролиза в сочетании с
химической модификацией, включающей избирательное окисление, связывание,
замещение остатков аминокислот и др.
При помощи методов ингибиторного анализа были предприняты попытки
установить закономерности состава и структуры активных центров у ферментов,
относящихся к разным группам. В частности, при использовании
диизопропилфторфосфата (ДФФ), принадлежащего к так называемым нервным ядам,
наблюдается полное выключение активного центра холинэстеразы - фермента,
катализирующего гидролиз ацетилхолина на холин и уксусную кислоту. Оказалось,
что этот ингибитор имеет близкое структурное сходство с ацетилхолином и подобно
ему взаимодействует с ОН-группой остатка серина в активном центре. Вызывая
фосфорилирование серина в активном центре ряда других ферментов, ДФФ также
инактивирует их действие:
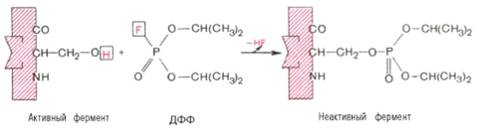
Показано, что ДФФ избирательно фосфорилирует в каждом чувствительном к
нему ферменте только один остаток серина, наделенный функциональной
активностью. Учитывая этот механизм действия ДФФ, сделаны попытки определения
природы аминокислот в окружении «каталитического» остатка серина у ряда
ферментов.
Помимо активного центра, в молекуле фермента может присутствовать также
аллостерический центр (или центры) (от греч. allos - другой, иной и steros -
пространственный, структурный), представляющий собой участок молекулы фермента,
с которым связываются определенные, обычно низкомолекулярные, вещества
(эффекторы, или модификаторы), молекулы которых отличаются по структуре от
субстратов. Присоединение эффектора к аллостерическому центру изменяет
третичную и часто также четвертичную структуру молекулы фермента и соответственно
конфигурацию активного центра, вызывая снижение или повышение энзиматической
активности. Ферменты, активность каталитического центра которых подвергается
изменению под влиянием аллостерических эффекторов, связывающихся с
аллостерическим центром, получили название аллостерических ферментов.
Отличительной особенностью ряда аллостерических ферментов является
наличие в молекуле олигомерного фермента нескольких активных центров и
нескольких аллостерических регуляторных центров, пространственно удаленных друг
от друга. В аллостерическом ферменте каждый из двух симметрично построенных
протомеров содержит один активный центр, связывающий субстрат S, и один
аллостерический центр, связывающий эффектор М2, т.е. 2 центра в одной молекуле
фермента. Получены доказательства, что для субстрата аллостерические ферменты,
помимо активного центра, содержат и так называемые эффекторные центры; при
связывании с эффекторным центром субстрат не подвергается каталитическому
превращению, однако он влияет на каталитическую эффективность активного центра.
Подобные взаимодействия между центрами, связывающими лиганды одного типа,
принято называть гомотропными взаимодействиями, а взаимодействия между
центрами, связывающими лиганды разных типов, - гетеротропными взаимодействиями.
Таким образом, в энзиматическом катализе, как и в реакции связывания
субстрата, участвует не ограниченная и небольшая часть фермента, как
предполагалось ранее, а значительно большая часть молекулы белка-фермента.
Этими обстоятельствами, вероятнее всего, можно объяснить большие размеры и
объемность трехмерной структуры молекулы фермента; эти же обстоятельства
следует учитывать в программах создания искусственных низкомолекулярных
аналогов ферментов (синзимов), обладающих свойствами нативных ферментов.
. Механизм действия ферментов
фермент биологический катализ трансаминирование
Раскрытие с помощью рентгеноструктурного анализа пространственного
строения ряда ферментов явилось надежной основой для построения рациональных
схем механизма их действия.
Установление механизма действия ферментов имеет ключевое значение для
раскрытия структурно-функциональных взаимосвязей во множестве биологически
активных систем.
Лизоцим обнаружен в различных тканях животных и растений, он находится, в
частности, в слезной жидкости и яичном белке. Лизоцим функционирует как
антибактериальный агент, катализируя гидролиз клеточных стенок ряда бактерий.
Этот полисахарид образован чередующимися остатками N-ацетилмурановой кислоты (NAM), соединенными β-1,4-гликозидной связью (полисахаридные
цепи сшиты короткими пептидными фрагментами).
Бактериальный полисахарид является весьма сложным нерастворимым
соединением, в связи с чем в качестве субстратов лизоцима часто используют
хорошо гидролизуемые олигосахариды, образованные остатками NAG.
Лизоцим белка куриных яиц образован одной полипептидной цепью, содержащей
129 аминокислотных остатков; его молекулярная масса составляет 14 600. Высокая
стабильность фермента обеспечивается наличием четырех дисульфидных мостиков.
Информация об активном центре и типе каталитического процесса была
получена Д. Филипсом в 1965г. на основе рентгеноструктурных исследований
лизоцима и его комплексов с ингибиторами. Молекула лизоцима имеет форму
эллипсоида с осями 4,5*3*3 нм; между двумя половинами молекулы находится «щель»,
в которой происходит связывание олигосахаридов. Стенки щели образованы в
основном боковыми цепями неполярных аминокислот, обеспечивающими связывание
неполярных молекул субстрата, и включают также боковые цепи полярных
аминокислот, которые способны образовывать водородные связи с ациламинными и
гидроксильными группами субстрата. Размер щели позволяет разместиться молекуле
олигосахарида, содержащей 6 остатков моносахаридов. Методом
рентгеноструктурного анализа установить характер связывания субстрата, например
гексасахарида NAG6, не удается. В то же время комплексы фермента с
ингибитором трисахаридом NAG3 стабильны
и хорошо изучены. NAG3 связывается в щели на поверхности
фермента, образуя водородные связи и ван-дер-ваальсовы контакты; при этом он
заполняет только половину щели, в которой могут связаться еще три
моносахаридных остатка. Невосстанавливающий конец (сахар А) оказывается у
начала щели, а восстанавливающий конец (сахар С) - в центральной ее части;
остатки сахаров А, В и С имеют конформацию кресла. Построение модели
фермент-субстратного комплекса было основано на предположении о том, что при
связывании субстрата NAG6
реализуются те же взаимодействия, что и при связывании NAG3. На модели фермента внутри щели были размещены три
остатка сахара (обозначаемые как остатки D, E и F); каждый последующий сахар
присоединялся таким образом, чтобы его конформация была такой же (насколько это
возможно), как и у первых трех сахаров. В составе модельного комплекса все
остатки сахаров реализуют эффективные нековалентные взаимодействия с боковыми и
пептидными группами аминокислотных остатков, образующих щель.
Предложена следующая схема каталитического процесса при гидролизе
олигосахарида. Неионизированная карбоксильная группа Glu-35 выступает в качестве донора протона, поставляя его
гликозидному атому кислорода между атомом С(1) сахара D и атомом С(4) сахара Е
(стадия общего кислотного катализа); это приводит к разрыву гликозидной связи.
В результате остаток сахара D
переходит в состояние карбкатиона с положительно заряженным атомом углерода С(1)
и принимает конформацию полукресла. Отрицательный заряд карбоксилатной группы Asp-52 стабилизирует карбкатион. Остаток
NAG2 (сахара E+F) диффундирует из области активного
центра. Затем в реакцию вступает молекула воды; ее протон переходит к Glu-35, а ОН- -группа к атому
С(1) остатка D
(стадия основного катализа). Остаток NAG4
(сахара А+В+С+D) уходит из области активного центра,
и фермент возвращается в исходное состояние.
Рибонуклеаза (РНКаза) поджелудочной железы быка гидролизует
межнуклеотидные связи в РНК около пиримилиновых звеньев, которые при этом
остаются этерифицированы по 3’-положению. Фермент наряду с другими нуклеазами
широко используется при анализе структуры РНК.
РНКаза образована одной полипептидной цепью, содержащей 124
аминокислотных остатка, а ее молекулярная масса равна 13 680; в молекуле
имеется четыре дисульфидные связи. РНКаза является первым ферментом, для
которого была установлена первичная структура.
На основании результатов исследования ренатурации рибонуклеазы К. Афинсен
впервые четко сформулировал представление о том, что пространственное строение
белка определяется его первичной структурой.
В 1958 г. Ф. Ричардс показал, что в определенных условиях субтилизин
расщепляет в РНКазе пептидную связь Ala-20 - Ser-21. Образующиеся фрагменты были
названии S-пептидом (остатки 1-20) и S-белком (остатки 21-124); за счет
нековалентных взаимодействий фрагменты образуют комплекс, названный РНКазой S. Этот комплекс обладает почти полной
каталитической активностью нативного фермента; в изолированном виде S-пептид и S-белок неактивны. Далее было установлено, что синтетический
пептид, идентичный по последовательности фрагменту S-пептида, содержащий остатки с 1 по 13, восстанавливает
активность S-белка, однако более короткий пептид,
содержащий остатки с 1 по 11, такой способностью не обладает. Полученные данные
позволили сделать заключение о том, что соответствующие остатки His-12 или Met-13 (или оба этих остатка) входят в активный центр фермента.
При исследовании влияния рН на активность РНКазы была выяснена важная
роль функциональных групп белка с рК 5,2 и 6,8; это позволяло предполагать
участие в каталитическом процессе остатков гистидина.
При карбоксилировании РНКазы иодацетатом при рН 5,5, т.е. в условиях, при
которых преимущественно происходит модификация остатков гистидина, наблудалась
полная утрата активности; модифицированный фермент содержит 1 моль
карбоксиметильных групп на 1 моль белка. В результате образуются две
монокарбоксиметиленовые формы фермента. В одной форме карбоксиметилированным
является остаток His-12, а в другой
- His-119. Преимущественно модифицировался
His-119.
Эти данные позволили предположить, что His-12 и His-119
находятся в активном центре и что модификация одного из них препятствует
модификации другого.
В результате рентгеноструктурных исследований было выяснено
пространственное строение РНКазы S и
комплекса РНКазы S с ингибиторами.
Молекула имеет форму почки, активный центр локализован в углублении, где
находятся остатки His-12, His-119 и Lys-41.
Гидролиз происходит в результате сопряженного действия остатков His-12 и His-119, осуществляющих кислотно-основной катализ. На
приведенной схеме указаны стадии каталитического процесса:
1. Субстрат находится в активном центре; His-12, His-119
и Lys-41 расположены около отрицательно
заряженного фосфата.
2. В результате действия His-12 как основания, акцептирующего протон от 2’-ОН-группы рибозы, и His-119 как кислоты, отдающей протон
атому кислорода фосфата, образуется сначала промежуточный комплекс, а затем
2’,3’-циклический фосфат.
. На место ушедшего продукта поступает вода, отдающая протон His-119, а ОН- - фосфату,
одновременно протон от His-12
переходит к кислородному атому рибозы, образуется второй продукт, а фермент возвращается
в исходное состояние.
Химотрипсин секретируется в форме профермента - химотрипсиногена
поджелудочной железой позвоночных животных; активация профермента происходит в
двенадцатиперстной кишке под действием трипсина. Физиологическая функция химотрипсина
- гидролих белков и полипептидов. Химотрипсин атакует преимущественно пептидные
связи, образованные карбоксильными остатками тирозина, триптофана, ценилаланина
и метионани. Он эффективно гидролизует также сложные эфиры соответствующих
аминокислот. Молекулярная масса химотрипсина равна 25 000, молекула содержит
241 аминокислотный остаток. Химотрипсин образован тремя полипептидными цепями,
которые связаны дисульфидными мостиками.
Функциональные группы активного центра химотрипсина идентифицированы с помощью
необратимых ингибиторов. Остаток Ser-195
был модифицирован диизопропилфторфосфатом и фенилметилсульфофторидом, а остаток
His-122 - N-тозил-L-фенилаланин-хлорметилкетоном.
Двухстадийность процесса химотрипсинового гидролиза была обнаружена при изучении
кинетики гидролиза п-нитрофенилацетата.
Характерной чертой рассматриваемого процесса является образование
ковалентного интермедиата - ацилфермента. Ацилируемая каталитическая группа
была идентефицирована - остаток Ser-195.
Механизм катализа, осуществляемого ферментом, был предложен еще до установления
пространственной структуры белка, но позднее был уточнен. В частности,
исследования с помощью 18Н2О позволили доказать
образование ацилфермента при гидролизе пептидов.
Трехмерная структура с разрешением 0,2 нм была установлена методом
рентгеноструктурного анализа Д.Блоу. в 1976г. Молекула имеет форму эллипсоида с
осями 5,4*4*4 нм. Результаты кристаллографических исследований подтвердили
предположения о том, что остатки Ser-195
и His-57 сближены. Гидроксильная группа Ser-195 находится на расстоянии ≈0,3
нм орт атома азота имидазольного кольца His-57. Наиболее интересным оказалось то обстоятельство,
что атом азота в положении 1 кольца находится на расстоянии ≈0,28 нм от
атома кислорода карбоксильной группы боковой цепи Asp-102 и занимает положение, благоприятное для
образования водородной связи.
Следует отметить, что химические исследования не могли выявить участия Asp-102 в функционировании активного
центра, поскольку этот остаток погружен вглубь молекулы.
В настоящее время считается, что три остатка Asp-102, His-57
и Ser-195 образуют систему переноса
заряда, которая играет решающую роль в процессе катализа. Функционирование
системы обеспечивает эффективное участие His-57 в катализе в качестве кислотно-основного
катализатора и повышает реакционную способность Ser-195 к карбоксильному углероду атакуемой связи.
Ключевым элементом катализа является перенос протона от Ser-195 к His-57. Одновременно происходит атака атомом кислорода
серина карбонильного атома углерода субстрата с образованием сначала
промежуточного тетраэдрического соединения (1), а затем ацилфермента (2). На
следующей стадии происходит деацилирование. Молекула воды поступает в систему
переноса заряда, а ион ОН- одновременно атакует карбонильный атом углерода
ацильной группы ацилфермента. Как и на стадии ацилирования, образуется
промежуточное тетраэдрическое соединение (4). Затем His-57 поставляет протон атому кислорода Ser-195, в результате чего
высвобождается ацильный продукт; он диффундирует в раствор, а фермент
возвращается в исходное состояние.
Карбоксипептидаза А секретируется в виде профермента поджелудочной
железой позвоночных животных. Образование активного фермента происходит в
тонком кишечнике при участии химотрипсина. Фермент последовательно отщепляет от
пептидной цепи остатки С-концевых аминокислот, т.е. является экзопептидазой.
Карбоксипептидаза А образована одиночной полипептидной цепью, содержащей
307 аминокислотных остатков; молекулярная масса равна 34 470. Аминокислотная
последовательность белка была установлена в 1969 г. Р. Бредщоу.
Выяснение механизма действия фермента оказалось возможным только после
проведения рентгеноструктурных исследований. Пространственная структура
фермента и его комплекса с дипептидом Gly-Tyr (модель субстрата) была установлена
У. Липскомбом. Молекула фермента имеет форму эллипсоида с осями 5,0*4,2*3,8 нм;
активный центр находится в углублении, переходящем в глубокий неполярный
карман. В зоне активного центра локализован ион цинка (его лигандами являются
боковые цепи остатков Glu-72,
His196, His-69 и молекула воды), а также функциональные группы,
участвующие в связывании субстрата и катализе, - остатки Arg-145, Glu-270 и Tyr-248.
При сравнительном анализе структур фермента и его комплекса с Gly-Tyr была получена важная информация о строении
фермент-субстратного комплекса. В частности, установлено, что при образовании
комплекса гидроксильная группа Tyr-248
перемещается на 1,2 нм по отношению к своему положению в свободном ферменте
(т.е. примерно на 1/3 диаметра молекулы).
Согласно схеме каталитического процесса, карбоксилатная группа Glu-270 активирует молекулу воды,
находящуюся в сфере реакции, оттягивая от нее протон; образующийся ион ОН-
осуществляет нуклеофильную атаку на карбонильный углерод расщепляемой связи.
Одновременно гидроксильная группа Tyr-248, находящаяся около атома азота расщепляемой пептидной связи, отдает
ему протон. В результате атакуемая пептидная связь расщепляется и образующиеся
продукты уходят из зоны активного центра. Приведенная схема иллюстрирует общий
основный катализ.
Аспартатаминотрансфераза катализирует обратимую реакцию
трансаминирования.

Ферментативная реакция трансаминирования была открыта А.Е. Браунштейном и
М.Г. Крицман в 1937г. при изучении ферментного препарата из мышцы голубя. В
последующих исследованиях было показано, что реакции трансаминирования широко
распространены в живой природе и играют важную роль в сопряжении азотистого и
энергетического обмена.
В 1945 г. было установлено, что пиридоксаль-5’-фосфат (ПЛФ) является
коферментом аминотрансфераз. Молекула ААТ является димером, образованным идентичными
субъединицами. В сердечной мышце исследованных позвоночных имеются два
изофермента - цитоплазматическая (цААТ0 и митохондриальная (мААТ)
аминотрансферазы.
Первичная структура цААТ из сердечной мышцы была установлена в 1972г.
Ю.А. Овчинниковым и А.Е. Брайнштейном. Полипептидная цепь белка содержит 412
аминокислотных остатков; молекулярная масса равна 46 000.
Общая теория пиридоксалевого катализа была разработана А.Е. Браунштейном
и М.М. Шемякиным в 1952-1953 гг., а несколько позднее - Д.Е. Мецлером и Е.Е.
Снеллом. Согласно этой теории, каталитическое действие пиридоксалевых
ферментовобусловлено способностью альдегидной группы пиридоксальфосфата
образовывать при взаимодействии с аминами, в том числе с аминокислотами,
альдимины (шиффовы основания).
В образующейся фосфопиридоксилденаминокислоте имеется система сопряженных
двойных связей, по которой происходит смещение электронов от α-углеродного атома облегчает разрыв
связей, образованных этим атомом.
Современные представления о механизме ферментативного трансаминирования,
разработанные А.Е. Браунштейном и его сотрудниками, являются развитием
рассмотренной выше теории. В исходном состоянии альдегидная группа
пиридоксальфосфата образует альдиминную связь с ε-аминогруппой остатка Lys-258 активного центра (I). При связывании аминокислоты
образуется комплекс Михаэлиса (II), а
затем альдимин между пиридоксальфосфатом и субстратом (III). В результате последующих превращений через
промежуточные стадии (IV) и
(V) образуется оксокислота (VI). Этим заканчивается первая
полуреакция трансаминирования. Повторение этих же стадий в «обратном
направлении» с новой оксикислотой составляет вторую полуреакцию, завершающую
каталитический цикл трансаминирования.
Миоглобин и гемоглобин
Эти два белка часто называют дыхательными ферментами. Взаимодействие их с
субстратом - кислородом выснено детально, прежде всего на основе
рентгеноструктурного анализа высокого разрешения. Трехмерная структура
миоглобина была определениа Дж. Кендрью в 1961г., а трехмерная структура
гемоглобина - М. Перутцем в 1960 г.
Молекула миоглобина имеет компактную форму - 4,5*3,5*2,5 нм,
полипептидная цепь образует 8 спирализованных участков, обозначаемых буквами от
А до Н. Она специализированным образом уложена вокруг большого плоского
железосодержащего кольца гема. Гем - это комплекс порфирина с двухвалентным
железом.
Полярные цепи пропионовой кислоты гема находятся на поверхности молекулы,
остальная часть гема погружена в глобулу. Связь гема с белком осуществляется за
счет координационной связи между атомом железа и атомом гистидина,
локализованного в спирали F; это
так называемый проксимальный гистидин. В гемовом кармане в составе спирали Е
локализован другой важный остаток гистидина - дистальный гистидин; он находится
с противоположной стороны от атома железа на большем расстоянии, чем
проксимальный гистидин. Область между железом гена и дистальным гистидином в
дизоксимиоглобине свободна, и липофильная молекула О2 может
связываться с железом гема, занимая шестое координационное положение.
Уникальной особенностью миоглобина, а также гемоглобина, является их
способность обратимо связывать О2 без окисления гемового Fe2+ в Fe3+. Это оказывается возможным, поскольку в гидрофобном гемовом
карман, из которого вытеснена вода, создается среда с низкой диэлектрической
проницаемостью.
При связывании О2 с атомом железа последний перемещается
примерно на 0,06 нм и оказывается в плоскости порфиринового кольца, т.е. в
энергетически более выгодном положении. Предполагают, что это перемещение
обусловлено тем, что ион Fe2+ в дезоксимиоглобине находится в
высокоспиновом состоянии и радиус его является слишком большим, чтобы он мог
разместиться в плоскости порфиринового кольца гема. При связывании же О2
ион Fe2+ переходит в низкомпиновое состояние и
его радиус уменьшается; теперь ион Fe2+ может переместиться в плоскость порфиринового кольца.
Гемоглобин - основной компонент эритроцитов крови, осуществляющий
доставку кислорода от легких к тканям, а углекислоты - из тканей в легкие.
Гемоглобины разных видов отличаются по форме кристаллов, растворимости,
сродству к кислороду. Это обусловлено различиями в аминокислотной
последовательности белков; гемовый компонент одинаков у гемоглобинов всех видов
позвоночных и некоторых беспозвоночных животных.
Гемоглобин человека представляет собой тетрамер, состоящий из четырех
субъединиц, двух α-субъединиц и двух β-субъединиц, содержащих по 141 и 146
аминокислотных остатков соответственно. Между первичными структурами α-
и β-субъединиц существует значительная
гомология, сходны также и конформация их полипептидных цепей.
Молекула гемоглобинаимеет сферическую форму, диаметр которой равен 5,5
нм. Четыре субъединицы упакованы в форме тетраэдра.
Данные рентгеноструктурного анализа показали, что оксигенирование
гемоглобина сопровождается рядом изменений. При низком разрешении установлено,
что в этом случае структура становится более компактной (атомы Fe
β-цепей сближаются
примерно на 0,6-0,7 нм), субъединицы поворачиваются друг относительно друга и
оси второго порядка на 10-15о. Результаты исследования при высоком
разрешении свидетельствуют о том, что особенно значительные изменения
происходят в области αβ-контактов.
К настоящему времени на основе рентгеноструктурных исследований и ряда
других методических подходов достигнуты значительные успехи в выяснении механизма
действия ферментов с заданными свойствами на основе достижений в области генной
инженерии. Это открывает широкие возможности для проверки справедливости
современных представлений о механизме действия ферментов и создания
фундаментальной теории ферментативного катал.
Библиографический список
1. А. Ленинджер Основы биохимии. - Москва ”Мир”, 1985.
. Ю.А. Овчинников. Биоорганическая химия. - Москва
“Просвещение”, 1987.
. Т.Т. Березов, Б.Ф. Коровкин. Биологическая химия. - Москва
“Медицина”, 1990.