Дослідження зчепленого успадкування ознак та рекомбінації генів у Drosophila melanogaster
МІНІСТЕРСТВО
ОСВІТИ ТА НАУКИ УКРАЇНИ
Харківський
національний педагогічний університет
імені
Г.С. Сковороди
Факультет
заочного навчання
Спеціальність
“Біологія”
Кафедра
ботаніки
Дослідження
зчепленого успадкування ознак та рекомбінації генів у Drosophila melanogaster
Дипломна
робота
Студента 51
групи
Бєлкіна
Данили Олександровича
Науковий
керівник:
викладач
Потапенко Г.С.
Консультант
викладач
Кудряшова Л.Г.
Харків
2007 р
ВСТУП
Підставою для вибору теми дипломної
роботи став той факт, що в колекції класичного об’єкта генетики - Drosophila
melanogaster, природничого факультету ХНПУ імені Г.С. Сковороди були відсутні
лінії з мутантними генами White та Cut. Ці мутації були тільки у зчепленому
вигляді (подвійні мутанти). Крім того, аналізуючи шкільну програму з біології
для класів з різним рівнем вивчення цієї дисципліни і на власному досвіді, який
ми набули при проходженні педагогічної практики, було з’ясовне, що тема “Явище
зчепленого успадкування. Кросинговер. Хромосомна теорія спадковості.” займає
недостатнє за кількістю годин місце у навчальній програмі. А втім ця тема одна
з найскладніших у загальному курсі біології і генетики зокрема. Серед
випускників шкіл мало хто може не тільки використовувати ці знання на практиці
(розв’язування задач, аналіз результатів дослідів), але й просто відтворити
усно її зміст. А без розуміння цих важливих явищ та закономірностей сприйняття
таких розділів загальної біології, як генетика, еволюція, популяційна генетика,
селекція, мінливість, навіть цитологія, основи екології значно ускладнюється,
йде на рівні запам`ятовування, а не розуміння. Таким чином, очевидно, що тема
нашої дипломної роботи є актуальною і необхідною, вона наочно демонструє
складні природні явища такі як кросинговер, успадкування, зчеплене зі статтю і
т.д., сприяє полегшенню засвоєння цього матеріалу.
Мета:
Дослідити особливості успадкування
мутацій ”white” та “cut” (відповідно “білі очі” та “зрізані крила”) у
Drosophila melanogaster.
Встановити відстань між локусами
генів white та cut.
Отримати нові форми одиночних
мутантів white та cut.
Розробити методичні рекомендації по
даній тематиці для вчителя біології відповідно до сучасних умов освіти.
Для досягнення найбільшої
результативності нашої роботи ми поставили перед собою такі задачі:
Ознайомитися та проаналізувати
літературні дані з теми досліджень для обґрунтування та вірного пояснення
власних експериментальних даних.
Ознайомитися з особливостями
біології Drosophila melanogaster, як об’єктом генетичних досліджень, та
методикою проведення експеріменту.
Провести серію спланованих
схрещувань серед вихідного генетичного матеріалу Drosophila melanogaster і
отримати необхідні мутантні форми.
Обробити експериментальні дані за
допомогою математичної статистики та зробити висновки.
По результатам експерименту скласти
типові задачі творчого рівня для учбового процесу школярів-старшокласників,
студентів та вчителів.
Скласти план-конспект уроку на тему
“Явище зчепленого успадкування. Кросинговер” Підібрати типові задачі на ці теми
різного рівня складності, та розробити змістовні методичні пояснення для їх
розв`язування. зчеплене успадкування drosophila ген
Робота була виконана на базі
лабораторії генетики природничого факультету ХНПУ імені Г.С. Сковороди під
керівництвом.
РОЗДІЛ 1.
Літературний
огляд
.1 Хромосомна теорія спадковості.
Кросинговер та конверсія генів
Проблема спадковості і тим більше
механізму спадковості турбувала і цікавила людство та передових вчених
споконвіку. Але ця проблема була дуже глибока, складна і майже до ХХ століття
вона залишалась загадкою. Багато вчених намагалися досліджувати цю проблему,
деякі навіть підійшли дуже близько до її розгадки, але остаточно вона була
вирішена тільки школою Т. Моргана. Навіть сам великий Ч. Дарвін намагався
розібратися в неї. Але спадковість - сама слабка частина теорії Ч. Дарвіна. Він
вважав, що в статевих клітинах є інформація про ознаки, які можуть проявитись в
наступних поколіннях. Але Дарвін не визначає, які ж саме ознаки успадковуються.
Він припускав, що деякі ознаки, які
виникли під впливом зовнішніх умов середовища можуть успадковуватись. Таким
чином, Ч. Дарвін висуває теорію пангенезиса: від кожної клітини кожного органа
відокремлюються маленькі частинки (пангени або гемули), з кров’ю вони проходять
по всім органам і частинам організму, “вбираючи” в себе ознаки, які їм
притаманні і потрапляють в статеві клітини, завдяки чому всі ознаки і зміни
успадковуються. Проте сам Ч. Дарвін називав ці погляди “спекулятивним сміттям”.
В основу неодарвінізму були
покладені переконання Ф. Гальтона (F. Galton), який працював по теорії
спадковості і створив відому теорію кореня, за якої окремі особини уявляються у
вигляді ніби то бокових відгалужень на безперервному кореневищі, яке відображає
історію статевих клітин.
Саме ці погляди і були далі розроблені
Вейсманом. Теорія кореня Гальтона стала основою вчення про безперервність
зачаткової плазми. При цьому досить чітко підкреслювалась відома протилежність
між сомою та статевими клітинами, а також відокремленість статевих елементів.
Відмічалась більша стійкість зачаткової плазми (тобто спадкової субстанції), а
також її здатність дробитись на спадкові одиниці різного порядку. Уявлення про
незалежність у змінах окремих ознак Вейсман пов’язав із мозаїчною теорією
особистого розвитку. Остання вже була розроблена В. Гісом (W. His) а також,
трохи в іншому вигляді, В. Ру (W. Roux). Як і Ру, Вейсман припускав, що у
процесі дроблення, а також при подальших, більш пізніх клітинних поділах
відбувається нерівний розподіл ядерної речовини (яка вміщує в собі спадкову субстанцію),
який і визначає різну подальшу долю клітин і часток зародку в процесі
індивідуального розвитку.
За Вейсманом, спадкова субстанція,
або зачаткова плазма складається з цілої ієрархії спадкових одиниць різного
порядку, локалізованих у ядрі клітини (іданти, відповідні до хромосом; іди,
відповідні до хромомер; детермінанти і біофори, які визначають розвиток окремих
часток).
З них найбільше значення мають
детермінанти, тобто визначники відомого роду клітин, а через це і окремих
часток організму. Припущення про існування відокремлених матеріальних одиниць
(детермінантів), які визначають розвиток окремих часток і навіть окремих
клітин, природно, примушує приймати і незалежність змін цих одиниць, а отже і
окремих частин організму в процесі еволюції. Організм уявляється ніби то
мозаїкою окремих ознак, які змінюються незалежно одна від одної.
За уявленнями Вейсмана, органи,
тканини, клітини і навіть молекули змагаються за їжу. В цій боротьбі
перемагають більш сильні частини, ті, що активно функціонують, в той час як
неактивні елементи атрофуються. Ці уявлення не зовсім вірні. Про пряму боротьбу
частин не може бути і мови, і часто ми бачимо, що тканина, яка не функціонує не
тільки не має браку у поживному матеріалі, а навпаки, повністю не використовує
речовин, що поступають, так що наприклад м`язи при їх атрофії проростають
жировою тканиною.
Суть поглядів Вейсмана на джерело
мінливості можна уявити так: первісне джерело мінливості - пряма дія зовнішніх
факторів на зачаткову плазму. Через ті чи інші причини ця дія по різному
відбивається на різних детермінантах. Результати цього диференційного впливу
викликають різницю у живленні зачаткової плазми. Через це одні детермінанти
(при кращому живленні) посилюються, а інші послаблюються. Ця різниця призводить
до внутрішньої боротьби. Сильні детермінанти більш активні, інтенсивно
живляться і через це посилюються ще більше, скоріше ростуть і спрямовують
розвиток більш активних тканин та органів. У ряді поколінь ці зміни, що
визначаються розвитком спадкових одиниць, посилюються ще більше. Як наслідок,
посилення детермінантів призводить до прогресивного розвитку органів та тканин.
Навпаки, ослаблені детермінанти стають ще більш слабкими у боротьбі з іншими
детермінантами і це призводить до редукції частин та органів.
Таким чином, зачатковий відбір, що
виникає на базі випадкової зміни (яка індукована зовнішнім фактором) веде до
ортогенетичної, тобто певним чином спрямованої мінливості та еволюції в її
початкових фазах. У результаті таких процесів можуть виникнути і зачатки нових
органів, які ортогенетично досягають такого рівня, коли вони починають набувати
селекційного значення у боротьбі за існування цілих особин.
Отже, тут має місце прийняття
принципів ектогенезу для виникнення змін та ортогенезу -для пояснення їх
подальшого розвитку. Природний добір тоді має справу із вже готовими формами.
Він виступає у ролі сита, яке знищує негодні форми. В уявленнях неодарвінізму
природний добір таким чином позбувається своєї мистецької ролі.
Зачатковий добір Вейсмнана не має
нічого спільного із теорією природного добору Дарвіна і по суті веде до її
заперечення. Між тим, Вейсман гадав, що саме це і є подальший розвиток
дарвінізму, який він ставив дуже високо. Вплив Вейсмана на розвиток теоретичної
біології був дуже вагомим, але це стосується інших його поглядів. Саме деякі
інші уявлення, пов’язані із вченням Вейсмана про зачаткову плазму та її
детермінанти а також із мозаїчною теорією розвитку набагато пережили ці
гіпотези та доволі міцно увійшли у свідомість біологів.
Уявлення про спадкові одиниці як
визначники окремих ознак і властивостей організму було сприйнято генетикою.
Уявлення про організм як про мозаїку ознак, що змінюються незалежно одна від
одної було сприйнято мутаційною теорією та довгий час трималося, головним чином
серед генетиків.
Проаналізувавши праці, в яких йшлося
про відкриття статевих хромосом у трав’яних клопів (Х. Генкінг, 1981; К. Мак.
Клунг, 1894; Е. Вильсон, 1902), закономірності дроблення яєць у морських їжаків
(Т. Бовері, 1902) та результати досліджень з вивчення сперматогенезу в коника
(Brachystila magna). У. Сеттон (1902) дійшов висновку, що “об’єднання в пари
батьківських і материнських хромосом та їхнє наступне розходження в період
редукційного поділу може становити фізичну основу закону спадковості Менделя”. В
подальших дослідженнях (1903) Сеттон переконався, що поведінка хромосом у
мейозі точно повторює розподіл встановлених Менделем спадкових факторів. У 1905
році Е. Вільсон сформулював основні положення хромосомних механізмів визначення
статі. Так виникла хромосомна теорія спадковості. В її основу була покладена
думка про те, що спадкові фактори, встановлені Менделем, входять до складу
хромосом. Рух хромосом в процесі гаметогенезу та запліднення створює паралелізм
в успадкуванні менделевських спадкових факторів та хромосом. Проте ця теорія не
була до кінця розроблена і викликала сумніві щодо її наукової правомірності.
Остаточна розробка хромосомної
теорії спадковості належить Т.Г. Моргану та його співробітникам, які проводили
свої дослідження на плодових мушках-дрозофілах. Об’єктами дослідження були
комахи виду Drosophila melanogaster, які мають короткий цикл розвитку (при
температурі 25 оС він триває 9 діб). Тіло цих комашок має довжину 2-3 мм. В
природі вони живляться гниючими овочами та фруктами, в лабораторних умовах -
поживною сумішшю, до складу якої входить кукурудзяне борошно, манна крупа або
картопля, цукор, сухофрукти, дріжджі, агар-агар. В пробірці діаметром 1,5 - 2,0
см, на дно якої наливають 8 - 10 мл поживного середовища, від окремої пари мух
можна отримати 20 - 70 комах.
У дрозофіл (Рис. 1) в ядрах
соматичних клітин міститься по чотири пари гомологічних хромосом. Вони мають по
три пари аутосом і по одній парі статевих хромосом. На тілі дрозофіл порівняно
легко ідентифікуються різні фенотипові ознаки, які формуються під впливом
мутантних генів.
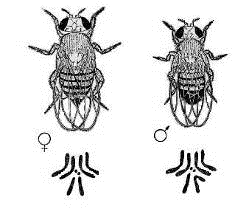
Рис. 1 Зовнішній вигляд самця і
самки дрозофіли і морфологія диплоїдних наборів їхніх хромосом
Швидке відтворення поколінь, низькі
витрати на розмноження й утримання, легкість в ідентифікації ознак - все це
робить дрозофілу зручним об’єктом генетичних досліджень.
В 1906 р. В. Бетсон і Р. Пеннет
опублікували працю, в якій повідомили, що в деяких схрещуваннях запашних
горошків розщеплення нащадків F2 не відповідає законам Менделя, однак не змогли
пояснити цього явища. Це пізніше зробив Т.Г. Морган на основі результатів
досліджень, виконаних під його керівництвом.
У дослідах на дрозофілах було
з’ясовано, що всі гени організмів цих комах виявляють тенденцію до успадкування
групами. У дрозофіли таких груп виявилось чотири. Оскільки в гаметах дрозофіли
локалізовано по чотири хромосоми, Морган зробив висновок, що кожна група генів
пов’язана з хромосомою. При будь-якому переміщенні хромосоми в мейозі, а також
при заплідненні у процесі розмноження, разом з хромосомою переносяться всі
локалізовані в ній гени. Отже, гени кожної групи виявилися зчепленими тілом
хромосоми. Тому сукупність генів, локалізованих в окремій хромосомі, Морган
назвав групою зчеплення.
Явище зчепленого успадкування було
виявлено в усіх видів еукаріотів. Щоб зрозуміти його суть, порівняємо характер
успадкування незчеплених та зчеплених генів А і В при аналізуючих схемах
схрещування. Зі схеми видно, що дигетерозиготи за незчепленими генами А і В у
F2 утворюють чотири рівновеликі групи нащадків (рис. 2 а). Але в разі зчеплених
генів А і В у F2 утворюються лише дві групи нащадків (рис. 2 б). Отже
розщеплення дигібрида в разі незчеплених генів А і В відбувається у
співвідношенні 1 : 1 : 1 : 1, а в разі зчеплених двох неалельних генів - у
співвідношенні 1 : 1.
|
не зчеплені (а)
|
зчеплені (б)
|
|
|
Р1:
|
АВАВ х аbаb
|
Р1:
|
АВАВ х аbаb
|
|
|
Гамети
|
АВ аb
|
Гамети
|
АВ аb
|
|
|
F1
|
АВаb
|
F1
|
АВаb
|
|
|
Р2:
|
АВаb х аbаb
|
Р2:
|
АВаb х аbаb
|
|
|
Гамети
|
АВАb аbаВ аb
|
Гамети
|
АВ аb аb
|
|
|
F2
|
АВАb Аbаb аВаb аbаb
|
F2
|
АВ аb
|
|
|
Відношення 1: 1: 1: 1
|
Відношення 1 : 1
|
Рис.2. Відміни в успадкуванні не
зчеплених (а) і зчеплених (б) ознак в аналізуючих схрещуваннях
В свою чергу, при дигібридному
схрещуванні з не зчепленими генами А і В утворюється покоління нащадків F2 зі
співвідношенням 9 : 3 : 3 : 1, а при схрещуванні особин зі зчепленими АВ-генами
в F2 утворюються лише дві групи нащадків у співвідношенні 3 : 1 (рис. 3).
|
не зчеплені (а)
|
зчеплені (б)
|
|
Р1:
|
ААВВ х ааbb
|
Р1:
|
ААВВ х ааbb
|
|
Гамети
|
АВ аb
|
Гамети
|
АВ аb
|
|
F1
|
АаВb
|
F1
|
АаВb
|
|
Р2:
|
АаВb х ааbb
|
Р2:
|
АаВb х ааbb
|
|
Гамети
|
АВ аb
|
Гамети
|
АВ аb АВ аb
|
|
F2
|
А_В_ А-bb аaВ_ aabb
|
F2
|
А_В_ аabb
|
|
Відношення 9: 3: 3: 1
|
Відношення 3 : 1
|
Рис.3 Відміни в успадкуванні
незчеплених (а) і зчеплених (б) АВ ознак в дигібридному схрещуванні
Таким чином, зчеплення суттєво
обмежує можливості перекомбінації генного матеріалу в генотипах організмів.
Явище зчепленого спадкування одержало назву закону Моргана, який можна
сформулювати так: гени, локалізовані в хромосомах, утворюють групи зчеплення,
кількість яких обмежена числом пар гомологічних хромосом, характерних для
відповідного виду організмів.
У дослідах на дрозофілах Морган,
крім явища зчепленого спадкування, виявив нові комбінації генів, які виникли в
результаті аналізуючого схрещування.
Сталося це так.
Коли Морган схрестив мух з ознаками
чорного тіла(b - black) і рудиментарних крил (vg - vestigial) (bbvgvg) з
особинами нормального забарвлення тіла та нормальних крил (b+b+vg+vg+), а
одержаних дигетерозиготних особин, в яких домінантні b+vg+ - гени комбінувались
з рецесивними алелями в цис-комбінації (b+vg+) / (bvg)*, схрестив з особинами
мутантної лінії материнського генотипу (bbvgvg), то від такого аналізуючого
схрещування було отримано чотири групи нащадків у співвідношенні 4,9 : 1 : 1 :
4,9 замість 1 : 1 : 1 : 1, яке очікувалось при відсутності зчеплення, або ж 1 :
1 при повному зчепленні bvg - генів. У другій серії схрещувань, де домінантний
b+ ген комбінувався з рецесивним vg - геном у транс-комбінації (b+vg) / (bvg+),
в F2 утворилось співвідношення 1 : 4,9 : 4,9 : 1 (на рис. 4 наведені схеми цих
схрещувань). Одержані числові співвідношення показали, що 83 % нащадків
відтворили батьківські комбінації bv-генів, а 17 % утворили нові комбінації цих
генів.
Аналіз закономірностей таких
розщеплень у багатьох аналогічних схемах схрещувань привів Моргана до висновку,
що нові комбінації генів, в умовах зчеплення їх, могли виникнути внаслідок
взаємного обміну гомологічними ділянками між гомологічними хромосомами в
процесі їхньої кон’югації в мейозі.
Цей висновок чітко підтвердився не
тільки на дрозофілах, але і на інших вивчених видах організмів. Явище взаємного
обміну гомологічними ділянками між гомологічними хромосомами, які входять до
складу бівалентів, дістало назву кросинговера або перехрещення хромосом, а типи
гамет, генотипи яких по-новому комбінувались між собою внаслідок кросинговера,
а також організми, що виникли від злиття цих гамет, були названі кросоверними.
Р 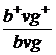 x
x 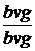
Гамети b+vg+ bvg bvg+vg bvg+  ;
; 
,5 % 8,5 % 8,5 % 41,5 % а
Р 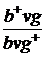 x
x
Гамети b+vg+ bvg bvg+vg bvg+ 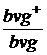 ;
;
,5 % 41,5 % 41,5 % 8,5 % б
Рис. 4. Відміни в характері
фактичного розщеплення нащадків за зчепленими bvg - генами в аналізуючих
схрещуваннях:
а - аналіз генотипу
дигетерозиготи, в якої домінантні і рецесивні гени знаходяться в цис-положенні
(b+vg+ // b vg)
б - аналіз генотипу
дигетерозиготи, в якої домінантний ген зчеплений з неалельним рецесивним геном,
утворюючи транс-комбінацію з неалельним домінантним геном (b+vg // b vg)
Ген b+ - нормальне і b -
чорне забарвлення тіла.
Ген vg+ - нормальні і vg -
редуковані крила.
Пізніше з’ясувалось, що
частота кросинговера між певними двома зчепленими генами в усіх особин виду
варіює в межах загальної для них середньої величини. Проте вона не однакова для
різних неалельних генів. Наприклад, частота кросинговера між геном С
(забарвлення алейрона) та геном sh (зморшкувате насіння) в кукурудзи в
середньому становить 3 %, а між геном С і геном wx (вокоподібний ендосперм) -
33 % . Це означає, що різні неалельні гени, локалізовані в одній і тій самій
хромосомі, мають неоднакову силу зчеплення.
Локалізація генів у
хромосомах і утворення кросоверних нащадків свідчать про те, що кросинговер
можливий лише при взаємному стиканні гомологічних хромосом. Вивчення
цитологічних механізмів мейозу показало, що таке стикання відбувається на
стадії зиготени мейозу і закінчується на стадії диплотени.
Кожна хромосома, яка вступає
в мейоз, складається з двох хроматид. Структури синаптонемального комплексу,
які формуються в процесі зближення та кон’югації пар хромосом-гомологів,
виконують функцію своєрідного шаблона, в рамках якого локалізуються хроматиди
обох гомологічних хромосом. Саме завдяки цьому молекули ферменту рН 5,2-нікази
можуть робити поперечні розрізи суворо в гомологічних точках хроматид, які
відносяться до складу двох гомологічних хромосом. Таким чином, кросоверні
розрізи зачіпають не цілу хромосому, а лише гомологічні точки двох хроматид,
котрі належать протилежним хромосомам-гомологам. Кожний розріз двох хроматид
утворює чотири відкритих кінці, які можуть знову об’єднуватись між собою за
допомогою ферментів лігаз. Якщо між собою об’єднаються відкриті кінці вихідної
хроматиди, це означатиме відновлення її вихідної цілості. Якщо ж між собою
об`єднаються кінці хроматид, що належать до двох хромосом-гомологів, утвориться
нова комбінація генів у межах хроматид відповідної групи зчеплення.
Описані явища відбуваються на
молекулярному рівні на стадії пахітени профази мейозу. В оптичному мікроскопі
їх побачити не можна. Проте результати гібридологічного аналізу в поєднанні з
цитологічним аналізом показують, що перехрещення хромосом обумовлює утворення
нових груп зчеплення, тобто нових комбінацій неалельних рецесивних та
домінантних генів у хромосомах.
Якщо взяти до уваги, що в
мейоз вступає багато клітин організму, які в нормі мають однакові каріотипи і
генотипи, стає зрозумілим, що пара гомологічних хромосом, гетерозиготних за
АБВГДЕЖ-генами, без кросинговера утворила б лише два типи гамет з вихідними
комбінаціями домінантних АБВГДЕЖ- і рецесивних абвгдеж-генів. Але процеси
кросинговера сприяють утворенню багатьох типів гамет з різними комбінаціями
домінантних і рецесивних генів. Згідно з законами Менделя кількість цих типів
гамет також має очікуватись з частотою 2n (у даному разі n означає число
гетерозиготних пар генів, які комбінуються між собою). В нашому прикладі
кількість типів гамет з новими комбінаціями домінантних і рецесивних генів
становитиме 27 = 128. Відзначимо, що в клітинах є не одна, а декілька пар
хромосом. Оскільки кожна хромосома може виявитись гетерозиготною за багатьма парами
неалельних генів, частота і кількість можливих нових комбінацій генів у групах
зчеплення суттєво зростає. Внаслідок цього створюється величезне генотипове
різноманіття нащадків, що сприяє природному добору й успішному плину
еволюційного процесу.
У 1909 році бельгійський
цитолог Ф. Янссен вивчаючи закономірності гаметогенезу в земноводних,
встановив, що після завершення кон’югації гомологічних хромосом наприкінці
профази першого мейотичного поділу, їхнє розходження супроводжується утворенням
між ними Х-подібних фігур, які були названі хіазмами. Пізніше Морган зробив
висновок, що хіазми утворюються внаслідок перехрещення, яке відбувається між
хромосомами. Численні дослідження підтвердили цей висновок Моргана.
Частота кросинговера між
генами може служити мірою відстані між ними лише в разі лінійного розміщення
генів у хромосомі. Саме віддаль між лінійно локалізованими генами вздовж
хромосоми може бути прямо пропорційною частоті кросинговера, який відбувся між
гомологічними хромосомами. Звідси випливає, що гени локалізуються в хромосомах
суворо в лінійному порядку.
Частоту кросинговера між
неалельними генами, що вивчаються, визначають методами гібридологічного
аналізу. Серед них найзручнішим є метод аналізуючих схрещувань. Розглянемо
приклад з дрозофілами. В третій хромосомі дрозофіли локалізуються такі три
гени: s- (spineles bristles) - шиповидні щетинки, g - (glass eyes) - скловидні
очі, е - (ebony) - ебенове забарвлення тіла.
Нормальних мух схрестили з
тригомозиготними рецесивними особинами за цими генами. Одержаних нащадків F1
знову схрестили з тригомозиготними рецесивними особинами за схемою аналізуючого
схрещування (рис.5).
Р:1 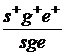 x
x 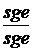
Гамети s+g+e+ sge:2 x
Гамети s+g+e+ sge sge+g+e
sge++ge +ge+ sg+e s+g+e+ - 5376- 5370 - 10746 - 88,8 % - некросоверні
нащадки+ge - 271+e+ - 257 - 528 - 4,32 % перехрест між sg - генами+g+e - 448+ -
445 - 893 - 7,32 % - перехрест між ge - генами +ge+ - 18+e - 16 - 34 - 0,28 % -
подвійний перехрест між sp i ge - генами
Всього - 12101 - 100 %
Рис. 5 Визначення частоти та
типу кросинговера між неалельними генами у Drosophila melanogaster - аналізуючи
схрещування:(spineles bristles) - шиповидні щетинки, - (glass eyes) - скловидні
очі,
е - (ebony) - ебенове
забарвлення тіла.
Аналіз одержаних результатів
показує, що фенотипові групи нащадків s+ g+ e+ / s g e та s g e / s g e -
генотипів повторюють комбінації s+ g+ e+ і s g e - неалельних генів
батьківських генотипів. Це означає, що в тригетерозиготних s+ g+ e+ / s g e -
клітинах у мейозі між s g e - генами перехрещення не відбулося. Внаслідок
редукційного поділу утворились гамети s+ g+ e+ та s g e -генотипів. Встановлено,
що частота некросоверних нащадків, а значить, і гамет, складає 88,08 %. Серед
одержаних нащадків F2 виникло 528 особин, або 4,32 %, в яких проявилась нова
комбінація між s g -генами, а саме - s +s gg ee - та ssg+ge+e - генотипів. Це
кросоверні особини за sg-генами. Крім того, утворилось 894; 7 або 7,32 %
кросоверних особин за g+e та ge+ генами, та 34; 2 або 0,28 % подвійних
кросоверів між s+ge+ та sg+e - генами. За одержаними експериментальними даними
(рис. 5) можна обчислити частоту перехрещення між гомологічними хромосомами,
гетерозиготними за s g e -генами. Наприклад, частота кросинговера між s
g-генами дорівнює 4,32 + 0,28 = 4,6 %, а між g e-генами аналогічна частота буде
7,32 + 0,28 = 7,6 %. З цієї ж схеми видно, що частота кросинговера між s e -
генами дорівнює сумі частот між s g та g e- генами. В нашому прикладі (рис. 5)
- 4,6 + 7,6 = 12,2 %.
Морган з`ясував також, що
розподіл точок на хромосомі, де відбуваються розрізи хромосомної ДНК молекулами
ферменту рН 5,2 - нікази, яким започатковується процес кросинговера,
підпорядковується закону ймовірностей. Тому теоретично очікувана частота
подвійного кросинговера між sge - генами дорівнює добутку загальних частот
кросинговера між sg- та ge-генами. Перетворивши для зручності відсотки в частки
одиниці, можна записати, що в нашому прикладі теоретично очікувана частота
подвійного кросинговера становитиме 0,046 х 0,076 = 0,0035 часток одиниці, або
0, 35 %. У досліді, згідно з одержаними даними (рис. 5), фактично відбулося 0,
28 % подвійних перехрещень, тобто на 0,07 % менше від теоретично очікуваної
частоти. Морган встановив, що розбіжність між теоретично очікуваною частотою
кросинговера і фактично одержаною обумовлена явищем інтерференції. У даному
випадку під інтерференцією розуміють зниження ймовірності сусіднього
перехрещення біля точки, в якій перехрещення відбулося. Розміри інтерференції
можна виміряти показником коїнциденції, за відношенням фактично одержаної
частоти перехрещення до теоретично очікуваної. Цей показник вимірюється в
частках одиниці або процентах. У наведеному прикладі показник коїнциденції, або
збіжності, становитиме - 0,28 / 0,35 = 0,8, або 80 %. Таким чином, частота
подвійного перехрещення між sge - генами здійснюється на 80 %, а в 20 %
випадків пригнічується.
Маючи усі вихідні дані,
можемо точніше визначити теоретично очікувану частоту перехрещення між sge -
генами. Вона становитиме:
між генами s i g - 4,32 +
0,35 % = 4,76 %
між генами g i e - 7,32 % +
0,35 % = 7,56 %
між генами s i e - 4,67 % +
7,67 % = 12,34 %.
Таким чином, за розмірами
інтерференції можна більш точно визначити теоретично очікувану частоту
кросинговера між зчепленими генами, що має важливе значення при побудові
генетичних карт. Якщо коїнциденція дорівнює одиниці, це означає, що на двох
відрізках хромосоми, які обмежують три гени, відбулись усі очікувані
перехрещення, а отже, теоретично очікувана і практично одержана частота
кросинговера збіжиться, якщо ж показник коїнциденції буде менше одиниці, це
означатиме, що частина подвійних перехрещень пригнічувалась внаслідок
інтерференції, і на даній ділянці хромосоми відбулись не всі очікувані подвійні
перехрещення. При цьому зауважимо, що зі скороченням двох відрізків хромосоми,
ймовірність перехрещення зменшується, а з збільшенням їхньої довжини - зростає.
Аналіз частот, розрахованих
за даними схемами (рис. 5) свідчить, що між кожною парою з трьох розглянутих
генів проявляється своя частота кросинговера. При цьому з’ясовується, що
фактична частота перехрещення між se- генами - це сума частот між sg i ge -
генами. Численні експерименти підтвердили даний висновок. Це дозволило А.
Стертеванту (1913) першому заявити, що в хромосомах дрозофіли локалізуються в
лінійному порядку.
Якщо вірно, що гени
локалізуються в хромосомах у лінійному порядку, а відстань між ними є функцією
частоти перехрещення, то за допомогою методів гібридологічного аналізу легко
визначити послідовність локалізації генів у хромосомах та відносні відстані між
ними. Наприклад, в одному з дослідів Моргана на дрозофілах були визначені
частоти перехрещення між зчепленими генами, які обумовлюють формування жовтого
тіла, білих фасеток очей та вилочної форми крил. Ці гени позначаються символами
у (yellow), w (white), bi (bifid). Гетерозиготними за цими генами самок Морган
схрестив з тригемізиготними самцями. Результати відповідного аналізуючого
схрещування були такими:
Частота перехрещення між y w
- генами - 1,2 %,
Частота перехрещення між w bi
- генами - 3,5 %,
Частота перехрещення між y bi
- генами - 4,7 %.
Крім того, іншими
дослідниками була виділена мутантна m-лінія низькожиттєздатних комах, котрі з
w-генами давали 5,8 % кросоверних нащадків, а з bi-геном - 2,3 %. Спробуємо всі
ці дані використати для побудови генетичної карти. (Під генетичною картою
розуміють послідовність розміщення локусів хромосоми та відстані між ними,
виражених у відсотках кросинговера).
Проведемо пряму лінію і
розділимо її на 100 однакових відрізків, тоді ціна кожної поділки дорівнюватиме
одному відсотку. Позначимо на цій прямій місце одного з генів, наприклад гена
у, а праворуч чи ліворуч від нього на відстані 1,2 % кросинговера - місце гена
w, тоді третій ген bi займе своє фіксоване місце відносно генів y i w. Так,
коли ген w поставили праворуч гена у, то ген bi зайняв місце на відстані 4,7 %
перехрещення від гена у та 3,5 % від гена w. Це відповідає частотам, які були
одержані в досліді. Відносно генів w i bi знайшов своє місце сусідній ген m, а
відносно генів bi та m буде визначено місце локалізації найближчого сусіднього
до них гена. За таким принципом будуються генетичні карти.
Генетичні карти дрозофіли
мають лише пізнавальне значення, а такі карти для рослин, тварин і людини - ще
й велике практичне значення. Вони ефективно використовуються в практиці
селекції, діагностиці спадкових захворювань, генній інженерії.
Соматичний кросинговер вперше
був виявлений К. Штерном (1931) у вигляді плям на тілі гетерозиготних самок
дрозофіли за генами жовтого тіла (у) та обпалених щетинок - (sn, від singe -
обпал). Одержані від схрещування мутантних жовтих мух у з мутантними особинами
sn, в яких дуже деформовані, наче обпалені) щетинки, нащадки першого покоління
були нормальними. Проте між ними Штерн виявив мух, в яких на грудях
проявляються ділянки жовтого тіла, а поруч - ділянки однакових контурів, на
яких проявляються обпалені щетинки. Такі ділянки-близнюки створюють ситуацію
мозаїчної химерності тіла. Ці факти Штерн пояснив наявністю випадків
кросинговеру, який відбувається в соматичних клітинах. Результати соматичного
кросинговера в клітинах гетерозиготних організмів показані на рис. 6. На цьому малюнку,
щоб легше було прослідкувати за ходом процесу кросинговера, хромосоми-гомологи
умовно показані одна тонкою, а інша - жирною лініями.

Рис. 6. Схема соматичного
кросинговера
Наприклад, на схемі (рис. 6)
видно, що кожна з вихідних гомологічних хромосом складається з двох хроматид.
Нагадаємо, що в нормі дві хроматиди будь-якої хромосоми завжди мають абсолютно
ідентичну молекулярну будову. Це стосується і хромосом (рис. 6). Як ми бачимо,
(рис. 6) кросинговер між кінетохором і sn-геном обумовив утворення
дигетерозиготних за генами sn+y та sny+ хромосом (рис. 6). Обидві ці
дигетерозиготні хромосоми при мітотичному поділі орієнтують половину своїх
генотипово однакових хроматид та протилежних полюсів (рис. 6). Внаслідок цього
утворюються дві дочірні клітини, одна з яких виявляється дигетерозиготною за
sn+y, а друга - за sny+ - генами. Кожна з цих клітин у процесі росту та
розвитку дасть свій клон клітин-нащадків, які на тілі відповідного організму
утворять дві однакові за розмірами та формою плями, але одна з них проявиться
жовтим тілом (sn+sn+yy - генотип), а друга - обпаленими щетинками (snsny+y+ -
генотип).
Соматичний кросинговер ще
називають мітотичним, оскільки він має місце в клітинах, які розмножуються
мітотичним поділом. Явище соматичного кросинговера досить рідкісне, проте воно
має велике значення в індукованій мінливості у видів, які втратили статеву
стадію розвитку, але перекомбінація генетичного матеріалу в них відбувається
завдяки парасексуальному процесу. Такими, зокрема, є незавершені гриби, багато
які види прокаріот тощо. Генетична рекомбінація шляхом мітотичного кросинговера
відбуваються також у рослин-апоміктів, у рослин, які в природі розмножуються
вегетативно. Явище соматичного кросинговера виявлено у тварин, у кукурудзи,
дріжджів.
Морган постулював явище
кросинговера, виходячи з теоретичних узагальнень, які випливали з
експериментальних даних і дозволяли переконливо пояснити процес розщеплення
ознак, генотипове проявлення яких було обумовлено функцією зчеплених генів.
Реальність кросинговера була доведена К. Штерном (1931) на дрозофілах та Г.
Крайтоном і В. Мак-Клінок (1931) на кукурудзі, котрі методами
експериментального мутагенезу створили лінії з мутаційно перебудованими
хромосомами. Від схрещування нормальних особин з представниками відповідних
ліній утворились гібридні нащадки з гетероморфними хромосомами-гомологами. Якщо
в цих гетероморфних парах гомологів локалізуються алельні гени, котрі
обумовлюють чіткий фенотиповий прояв ознаки, то за характером ознак кросоверних
нащадків F2 фахівець, не вдаючись до мікроскопічних досліджень, безпомилково
опише форму хромосом, яка має бути в кожній фенотиповій групі цих нащадків.
Для наочності розглянемо
схему дослідів Штерна. У дрозофіли статеві ХУ-хромосоми морфологічно не
однакові. Наприклад, Х-хромосома акроцентрична, паличковидної форми, а
У-хромосома дещо більша, ніж Х-хромосома, метаакроцентрична. Штерну пощастило
до однієї з Х - хромосом приєднати фрагмент У-хромосоми. Внаслідок цього
виникла життєздатна Г-подібна Х - хромосома. Разом з цим друга Х - хромосома
розірвалась на дві частини, одна з яких у вигляді акроцентричного фрагмента
приєдналась до четвертої хромосоми, а друга - з наявною у неї центромерою,
продовжувала існувати як самостійна хромосома. Але саме в другій частині були
локалізовані мутантні гені cr (carnation), який обумовлював забарвлення фасеток
очей кольору гвоздики, та Bar, який обумовлював більше як удвічі зменшення
кількості фасеток очей. Уцілілі фасетки в кожному оці розміщуються своєрідною
смужкою, тому в мутантних особин формуються смугоподібні очі. Це домінантна
мутація, яку позначають символом В (Bar). У другій гомологічній, але Г -
подібній хромосомі локалізовані нормальні гени дикого типу, тобто cr+ i B+.
На схемі (рис. 7) видно, що
від схрещування самок, в яких перебудовані статеві хромосоми і фенотипово
проявляється форма смугоподібних очей, із самцями, в яких фасетки очей кольору
гвоздики, в нащадків жіночої статі без кросинговеру утворюються дві фенотипові
групи (рис. 7). Це група подвійних мутантів з очима кольору гвоздики і форми
бар. У цих особин одна Х - хромосома має нормальну, паличкоподібну форму, а
друга складається з двох частин, які в зоні стику утворюють контури, подібні до
перетяжки. Інша група некросоверних нащадків характеризується нормальними за
всіма ознаками очима, але в них одна Х - хромосома нормальна, а друга - Г -
подібна.
Що ж до кросоверних нащадків,
то вони теж характеризуються двома фенотиповими групами: в особин однієї групи
очі кольору гвоздики, але нормальної форми, в особин другої групи очі
нормального кольору, але форми бар. Нащадки з очима кольору гвоздики
характеризуються нормальними паличкоподібними Х - хромосомами, а в особин з
очима бар одна Х - хромосома нормальна, а друга - Г - подібна. Ця Г - подібна
хромосома проявляється за наявністю в неї розриву, який імітує перетяжку.
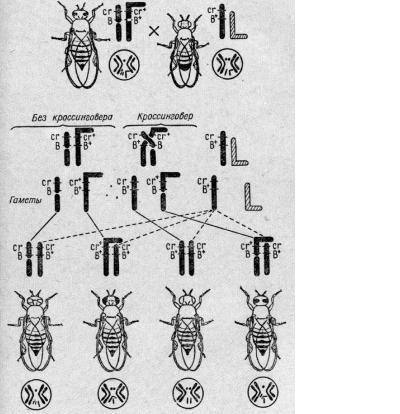
Рис. 7. Цитологічна
доказовість кросинговеру в дрозофіли К. Штерном.
Таким чином, можливість
прогнозування морфології статевих хромосом за фенотипами, або ж навпаки,
прогнозування фенотипів за морфологією статевих хромосом є переконливим доказом
того, що між зчепленими генами, які обумовлюють забарвлення фасеток очей та
характер розміщення фасеток, відбувається кросинговер.
Серед методів доведення
існування кросинговера заслуговує на увагу тетрадний аналіз у сумчастих грибів,
які характеризуються зиготичним мейозом. У цих грибів після запліднення
диплоїдна зигота вступає в мейоз. Мейотичний поділ зиготи супроводжується
формуванням сумки (аски), в якій розміщуються продукти мейозу. Після двох
мейотичних поділів у сумці утворюється чотири гаплоїдні клітини. Кожна з них ще
раз ділиться мітотично, внаслідок чого утворюється по дві ідентичні копії, з
яких формується вісім аскоспор. У сумці всі аскоспори розміщуються одним
рядочком.
У червоної хлібної плісені
Neurospora crasa виявили мутацію lys-5, яка суттєво затримує дозрівання
аскоспор. Тут дозрілі спори мають чорне, а недозрілі - біле забарвлення. Для
зручності позначимо мутацію затримки дозрівання спор через а, а ген своєчасного
дозрівання - через А. Це дає можливість простежити на схемах за переміщенням
мутантного і немутантного аллельних генів один відносно одного і зрозуміти
причини неоднакового розміщення білих і чорних спор у сумках, як це показано на
рис. 8.

Рис.8. Розміщення дозрілих (темні) і
недозрілих (світлі) аскоспор в сумках Neurospora (ліворуч). Схема різних типів
поведінки гомологічних хромосом при утворенні акоспор в сумках Neurospora
(праворуч):
- кросинговер не відбувається;
- кросинговер відбувається;
А - домінантний ген своєчасного
дозрівання спор, які темніють;
а - рецесивний ген відставання
дозрівання спор, які залишаються білими.
Зі схеми видно, що хромосома, яка
вступає в мейоз, складається з двох ниток (хроматид). Це означає, що в складі
хромосоми кожний ген, у тому числі і ген А, представлений двома копіями. Таким
чином, у гетерозиготної за геном А зиготі в двох хроматидах однієї хромосоми є
дві копії домінантного А -і дві копії рецесивного - а гена. Під записом кожної
з цих копій на рис. 8 будемо мати на увазі не тільки ген А, але і хроматиду, в
якій він локалізується. Оскільки ми простежуємо переміщення в мейозі лише двох
алельних генів, локалізованих у парі хромосом-гомологів, то й наводиться схема
лише цієї пари гомологів.
З наведеної схеми (рис. 8) видно, що
відносне розміщення дозрілих і недозрілих спор у сумках нейроспори залежить від
того, відбувся між хромосомами кросинговер чи ні. Якщо кросинговер не відбувся,
то в сумці лежать підряд чотири дозрілі, чорні і чотири недозрілі, білі спори
(рис. 8). Якщо ж кросинговер відбувся, то дозрілі і недозрілі аскоспори в асках
розміщуються в залежності від того, як були орієнтовані між собою хроматиди
гомологічних хромосом (рис. 8). Порядок розміщення аскоспор переконує, що він
може виникнути лише за рахунок перехрещення, яке відбулося між хромосомами в
складі бівалентів на стадії чотирьох ниток. При цьому перехрещення на стадії
чотирьох ниток обумовлює утворення половини кросоверних і половини
некросоверних продуктів мейозу. Якби перехрещення відбувалося між цілими
хромосомами (тобто на стадії двох ниток), то всі продукти мейозу були б
кросоверні, чого насправді не має.
Розглянуті нами схеми кросинговера,
представлені експериментальними даними, свідчать, що в аналізуючих схрещуваннях
кожній утвореній групі кросоверних особин, у яких проявились ознаки у
домінантному стані, протистоїть рівновелика група кросоверних особин, в яких та
ж сама ознака проявилась у рецесивному стані. Наприклад, від схрещування
цис-гетерозиготи з рецесивною за bvg-генами особиною (рис. 4) 8,5 % кросоверним
нащадкам, які мали нормальне сіре тіло і редуковані крила, протистояло 8,5 %
кросоверних нащадків, в яких було чорне тіло, але нормально розвинені крила.
Аналогічні дані були отримані і в разі залучення до аналізуючого схрещування
транс-гетерозиготи за цими ж ознаками (рис. 4), а також при визначенні частоти
кросинговеру між зчепленими генами (рис. 4).
Рівновеликість будь-яких двох
кросоверних груп, які характеризуються альтернативними комбінаціями одних і тих
самих ознак, а отже і альтернативними комбінаціями домінантних і рецесивних
генів, котрі обумовлюють ці ознаки, є однією з важливих закономірностей
рекомбінації генетичного матеріалу. Проте Г. Кніпом було встановлено (1926), що
внаслідок мейотичного поділу а+а -гетерозиготних зигот в нейроспори замість
очікуваного розщеплення 1а+ : 1а іноді воно відбувається у співвідношенні 1а+ :
3а або 3а+ : 1а. Крім того, хоча й дуже рідко, але можливе співвідношення 4а+ :
0а, або 0а+ : 4а замість очікуваного 1а+ : 1а, що свідчить про порушення
рівновеликості у відтворенні одного з двох алельних генів.
Порушення співвідношень у
відтворенні рівновеликих частот алельних генів у процесі мейотичного поділу
було виявлене не тільки в нейроспори, але і в дріжджових грибів та у багатьох
видів високоорганізованих організмів. У 1930 р. Г. Вінклер запропонував явище
порушення реципрокності у відтворенні частот алельних генів у мейозі називати
конверсією генів. Отже під конверсією генів слід розуміти процес переходу
домінантного гену в рецесивний стан або, навпаки, - процес переходу рецесивного
гена в домінантний стан у мейозі, який відбувається в гетерозиготних за даним
геном особин.
Молекулярні механізми конверсії
генів довгий час були предметом обговорення в наукових колах. У наш час
найбільш визначною і прийнятною є гипотеза Р. Холлідея (1964). Щоб зрозуміти
сутність гіпотези Холлідея, нагадаємо, що фермент рН 5,2-ніказа в межах одного
з бівалентів у гомологічних точках гомологічних хромосом розрізує дві нитки
ДНК, котрі належать до складу хроматид останнього. Нагадаємо також, що дві
нитки дволанцюгової спіралі молекули ДНК є протилежно полярними. З’ясовано, що
в мейозі рН 5,2-ніказа розрізує дві нитки ДНК, котрі мають однакову полярність.
Згідно ж з гіпотезою Холлідея (і в цьому полягає її сутність) системи
ферментів, які забезпечують процеси конверсії гена, розрізують дві протилежно
полярні нитки ДНК.
До складу бівалентів двома
хромосомами привноситься по дві хроматиди, які пронумеруємо від I
до IV. Те, що конверсія генів
трапляється лише у гетерозиготних особин, означає, що в одній їхній хромосомі
має локалізуватись немутантний, а в гомологічній хромосомі - мутантний алель
відповідного гена. Умовимось, що ці два алеля відрізняються між собою лише
однією парою азотистих основ, локалізованих у гомологічних точках ДНК даних
хромосом. Наприклад, пара А-Т у складі немутантного гена заміщена парою Г-Ц у
складі мутантного алеля. Для спрощення ситуації умовимося, що у процесі
кон’югації хромосом-гомологів на відповідних ділянках I
та IV хроматид ніяких змін не відбулося,
і розглянемо ці зміни лише на хроматидах II
і III.
Інша ситуація складається у разі
конверсії генів. Тут на II і III
хроматидах, у зоні гетеродуплексів, ферментами було розрізано і видалено по
фрагменту зі складу ниток ДНК протилежної полярності. Внаслідок забудови
утворених прогалин комплементарними нуклеотидами А-Г та Ц-Г гетеродуплекси в
обох хроматидах замістились Ц-Г парами. Результати описаних процесів свідчать,
що розщеплення серед хромосом нового покоління відбулося у співвідношенні 1А-Т
: 3Г-Ц. Однак якщо брати до уваги лише групу кросоверних хромосом, одержимо
розщеплення у співвідношенні 0А-Т : 2Г-Ц.
Таким чином, утворення гетеродуплексів
у складі бівалентів індукує реакції репаративного синтезу, який забезпечує
заміщення гетеродуплексів парами комплементарних нуклеотидів. Якщо ферменти
здійснюють репаративний синтез на двох нитках ДНК однакової полярності, процес
закінчується кросинговером, а якщо репаративний синтез відбувається на
матричних нитках різної полярності, це спричиняється до процесу конверсії
генів.
Факти існування груп зчеплення
показують, що якби це зчеплення було абсолютним, то утворення нових комбінацій
між неалельними генами в межах окремих груп було б неможливим. Вірніше,
комбінування генів у групах зчеплення могло б здійснюватись лише за рахунок
спонтанної мутації генів. Однак цей процес дуже повільний, і якби не було інших
механізмів утворення нових комбінацій, процес еволюції окремих генів, а отже, й
видів суттєво уповільнився б. Саме з цієї точки зору слід оцінювати роль
кросинговеру в еволюції. Виходячи з розглянутих фактів, можна стверджувати, що
вона величезна. Пояснюється це тим, що кросинговер забезпечує дієвість законів
Менделя за частотами груп генетично перекомбінованих генів. Наприклад,
кількість генотипових сортів гамет, які утворюють ди- або полігетерозиготна
особина, або ж кількість фенотипових груп нащадків F2 від ди- чи полігібридних
схрещувань дорівнює 2n. Кількість генотипових нащадків F2 дорівнює 3n, а
кількість комбінацій гамет - 4n. Для усіх випадків n означає число неалельних
генів, залучених до вивчення). В цих формулах не відтворюються лише
співвідношення частот генотипові різних сортів гамет чи особин у F2, а також
частот у межах кожного типу комбінацій гамет.
Наприклад, відповідно до законів
Менделя, тригетерозигота утворює вісім рівновеликих генотипових сортів гамет у
співвідношенні 1 : 1 : 1 : 1 : 1 : 1 : 1 : 1. Однак якщо ці гени зчеплені, то
співвідношення частот суттєво змінюються, кількість же фенотипових груп гамет
залишається сталою. Наприклад, зі схеми аналізуючого схрещування
тригетерозиготи (рис. 5) видно, що ця особина, як і очікується, утворює вісім
типів гамет. Легко обчислити, що тут співвідношення частота дорівнює 316 : 15,5
: 26 : 1 : 1 : 26 : 15,5 : 316. Покоління, що утворюються в F2 внаслідок
кросинговера, завжди успадковуються за Менделем. Саме цим забезпечується
величезна комбінативна мінливість у поколіннях нащадків, що активно стимулює
темпи еволюції видів.
.2 Хромосомні типи визначення статі.
Успадкування ознак, зчеплених зі статтю
Відповідно до співвідношень статевих
хромосом розрізняють чотири типи хромосомних механізмів визначення статі: ХУ-,
ХО-, ZW- i ZO-типи.
ХУ-тип - характеризується
гетерогаметністю чоловічих особин за Х та У-хромосомами і гомогаметністю
жіночих за Х-хромосомами. Статеві формули мають такий вигляд: ♀ = ХХ, ♂
= ХУ. Такий хромосомний тип виявлений у клопів Lygaeus, переважної більшості
видів комах, в розвитку яких немає стадії метелика, всіх видів ссавців,
дводомних видів рослин за винятком видів роду Fragaria.
ХО-тип - єднає види, в яких чоловічі
особини гетерогаметні за Х і О - хромосомами, а жіночі - гомогаметні. Статеві
формули за їхніми каріотипами мають такий вигляд: ♀ = ХХ, ♂ = ХО.
До цього типу відносяться трав’яні клопи Protrnor, цвіркуни, рослини видів роду
Dioscorea тощо.тип - за статевими хромосомами тут гетерогаметними є жіночі
особини, а гомогаметними - чоловічі. Виходячи з цього, статеві формули за
комбінаціями жіночих та чоловічих статевих хромосом мають вигляд: ♀ = ZW,
♂ = ZZ. До цього виду визначення статі відносяться деякі види риб,
переважна більшість видів комах, у циклі розвитку яких є стадія метелика, всі
птахи, багато видів амфібій та рептилій, а також види роду Fragaria.тип -
характеризується гетерогаметністю жіночої статі, та гомогаметністю чоловічої. В
даному разі статеві формули за статевими хромосомами мають такий вигляд: ♀
= ZO, ♂= ZZ. До цього типу належать метелики молі Fumea та особини ящірки
виду Lacerta vivipara, які зустрічаються лише на острові Сахалин.
У статевих хромосомах, як і в
аутосомах, локалізуються гени. Більшість їх не впливають на процеси формування
первинно- або вторинно-статевих ознак організму. Ці гени обумовлюють формування
ознак, котрі можуть належати однаковою мірою особинам і жіночої і чоловічої
статі. Своєрідність закономірностей цього успадкування визначається тим, що У
або W-хромосоми характеризуються надто низькою генетичною активність, тим часом
як гени, локалізовані в Х- або Z-хромосомах, функціонують нормально. Особини
ХХ- чи ZZ-каріотипів, незалежно від того, які гени в них локалізовані, можуть
бути гомозиготними або гетерозиготними. На відміну від цього в особин ХУ-, ХО-,
ZW-, чи ZO-каріотипів з генетично неактивною У- або W-хромосомою або без неї,
кожний локалізований в Х- чи в Z-хромосомі ген знаходиться в одному екземплярі
(без своєї алельної пари), тобто в гемізиготному стані. Тому такі гени,
незалежно від того, домінантні вони чи рецесивні, у складі генотипу завжди
дають фенотиповий прояв, оскільки кожний з них - повторимо ще раз - перебуває в
гемізиготному стані.
На закономірності успадкування
ознак, гени яких локалізовані у статевих хромосомах, впливають особливості
спадкування саме цих хромосом. Розглянемо схему схрещування самок плодової
мушки - дрозофіли (Drosophila melanogaster), фасетки очей в якої мають
нормальне червоне забарвлення, з білоокими самцями. Ген забарвлення очей
позначимо символом w (від white - білий) або w+. Це ген дикого типу, який
обумовлює червоне забарвлення очей. Він домінує над геном w і складає з ним
алельну пару в генотипі гетерозиготних за цим геном самок. У каріотиповій схемі
статеві хромосоми з мутантним геном білих очей позначимо через Хw, а з нормальним
геном дикого типу - через Х+.
Оскільки Х-хромосоми позначено
мутантним геном w та його нормальним + алеломорфом, то чітко простежується, що
батько віддає єдину Х-хромосому лише своїм дочкам, а мати віддає по одній
Х-хромосомі нащадкам як жіночої, так і чоловічої статі. Цим фактом пояснюється
те, що рецесивний ген білого забарвлення очей переходить від батька через
каріотипи дочок до половини онуків жіночої та чоловічої статі. Проте у генотипі
жіночої статі цей ген комбінується зі своїм домінантним алелем і тому
фенотипово не проявляється, тим часом як у генотипі чоловічої статі він
виявляється в гемізиготному стані і тому фенотипово проявляється. При цьому
фенотипово рецесивна ознака проявляється лише в чверті частини нащадків F2,
тобто у співвідношенні 3 : 1. Але особливістю цих співвідношень є те, що
рецесивна ознака проявляється лише в особин чоловічої статі.
Щоб переконатись у сказаному,
розглянемо таку схему схрещувань, де домінантний і рецесивний алелі
локалізовані в аутосомах. Наприклад, ген vg (від vestigial - зачаткові крила) у
дрозофіли локалізується в другій хромосомі. Якщо нормальних самок схрестити з
безкрилими самцями, до в другому поколінні розщеплення відбувається у
відношенні 3 : 1. Але в даному разі розщеплення торкається нащадків і жіночої і
чоловічої статі.
Отже, вихідні чоловічі особини, які
були залучені до схрещувань, характеризувались відсутністю крил (рецесивний ген
vg), а жіночі особини були з нормальними крилами (домінантний ген vg+), нащадки
як жіночої, так і чоловічої статі однаково розщепились у відношенні 3 : 1. Якщо
порівняти між собою розщеплення нащадків F2, котре проявилось у двох останніх
схемах схрещувань, стане видним, що характер цього розщеплення обумовлений
особливостями успадкування статевих хромосом. Ці особливості ще виразніше
проявляються в реципрокних схрещуваннях.
Аналіз схеми успадкування
рецесивного гена білих очей, якими характеризувались вихідна материнська форма
свідчить, що нащадки F2 як жіночої так і чоловічої статі розщепились за цією
ознакою у співвідношенні 1 : 1. Проте в разі локалізації гена в аутосомах
нащадки F2 як у прямих, так і в реципрокних схрещуваннях завжди розщеплюються у
співвідношенні 3 : 1. В цьому можна переконатись, якщо порівняти характер
розщеплення нащадків F2 за ознакою редукованих крил у прямих та реципрокних
схрещуваннях. Видно, що зміна напрямків схрещувань за домінантними та
рецесивними алельними генами, локалізованими в аутосомах, не змінила характеру
розщеплення нащадків F2 за цими генами.
Аналогічні закономірності
успадкування ознак, зчеплених зі статтю, проявляються і в тому разі, коли
гетерогаметною є жіноча, а гомогаметною - чоловіча стать. Наприклад, у курей
породи “Чорний лангшан” та “Зозулястий плімутрок” чорне оперення є рецесивною
ознакою, а зозулясте - домінантною. Якщо чорну курку схрестити із зозулястим
півнем, то в першому поколінні всі нащадки будуть зозулястими, а в другому -
половина самок буде чорних, а половина - зозулястих. Разом з тим, усі самці
будуть лише зозулястими. Таким чином, у видів, у яких жіноча стать є
гетерогаметною, зчеплені зі статтю ознаки успадковуються від матері через
сина-носія в половини нащадків F2 жіночої статі. А в реципрокних схрещуваннях
нащадки розщеплюються за характером домінантного та рецесивного фенотипового
прояву ознак в співвідношенні 1 : 1.
Розглядаючи схеми успадкування
статевих хромосом, маркованих мутантним геном w, ми мали можливість
переконатись, що гетерогаметний батько віддає свою єдину Х - хромосому нащадкам
жіночої статі, а гомогаметна мати передає по одній Х - хромосомі нащадкам
чоловічої та жіночої статі. Аналогічна закономірність проявляється і у видів, у
яких гетерогаметною є жіноча, а гомогаметною - чоловіча стать. Але тут,
навпаки, гетерогаметна мати віддає свою єдину Z-хромосому нащадкам лише
чоловічої статі, а гомогаметний батько передає по одній із двох своїх Z -
хромосом нащадкам і чоловічої і жіночої статі.
Перехресне успадкування статевих
хромосом при гетерогаметності як чоловічої так і жіночої статі дістало назву
крис-крос успадкування. Закономірності крис-крос успадкування проявляються не
тільки в комах, а і в амфібій, рептилій, птахів, ссавців, у дводомних видів
рослин.
З наведеного ми бачимо, що гени,
локалізовані в Х - або Z - хромосомах, у разі гетерогаметності чоловічої статі
передаються від батька лише до дочки, а в разі гетерогаметності жіночої статі -
від матері до сина. При цьому в разі гетерогаметності чоловічої статі батько
передає майбутньому синові єдину У - хромосому. Аналогічна ситуація складається
і в разі гетерогаметності жіночої статі: тут мати передає свою єдину W -
хромосому лише дочці.
В генотипі людини міститься понад 50
000 пар алельних генів. Це означає, що в гаплоїдному наборі хромосом кожний з
цих генів представлений в одному екземплярі. Якщо зазначені гени містяться в 23
хромосомах гаплоїдного набору, це означає, що в середньому в кожній з цих
хромосом має міститись понад 2000 генів. Але на сьогодні в Х - хромосомі людини
поки що виявлено місце локалізації лише біля 200 генів. Зауважимо, що будь-який
ген може бути виявлений за умови, що він мутує, а утворена мутація фенотипово
проявляється новим якісним станом відповідної ознаки або летальним ефектом.
Якщо мутантний ген локалізується в
статевій Х - хромосомі, то в складі чоловічого ХУ - каріотипу він опиняється в
гемізиготному стані й одразу характеризується фенотиповим проявом, тим часом як
у складі жіночого ХХ - каріотипу рецесивний мутантний ген як завгодно довго
може “ховатись” за своїм нормальним алеломорфом. З іншого боку, слід пам’ятати,
що кожна мутація виникає не спрямовано, під впливом випадкової дії факторів
зовнішнього та внутрішнього середовища. Тому мутантна зміна функції гена
болісно сприймається організмом. Як тільки мутантний ген опиняється в
гомозиготному або в гемізиготному стані і проявляється фенотипово, він обумовлює
або загибель організму, або його хворобливий стан.
Наприклад, у людини ген зсідання
крові мутує в новий стан, при якому в гомозиготних чи гемізиготних за цією
мутацією особин у разі поранення кров не зсідається. Ця мутація, і, відповідно,
й хвороба дістали назву гемофілія. Якщо хворий на гемофілію чоловік вступає у
шлюб зі здоровою жінкою, то свою єдину Х - хромосому, в якій локалізується ген
зсідання крові, він передає лише дочкам. Його ж сини одержують по одній Х -
хромосомі від здорової матері, а від батька по У - хромосомі, до складу якої
ген гемофілії не входить. У наступному поколінні гетерозиготні за геном
зсідання крові дочки віддають по одній Х - хромосомі з мутацією гемофілії одній
половині своїх синів, а з немутантним, нормальним геном зсідання крові - другій
половині. Згідно з таким розподілом сини, які отримали від матері Х - хромосому
з мутантним геном зсідання крові, виявляються гемофіліками, а сини, які
отримали немутантну за геном зсідання крові Х - хромосому, виявляються повністю
здоровими за ознакою гемофілії.
Отже, ознака гемофілії в людини
успадковується за крис-крос схемою. За цією схемою успадковуються й інші
рецесивні мутації, гени яких локалізовані в Х - хромосомі. Наприклад, мутація,
що обумовлює псевдогіпертрофічну м’язову дистрофію Дюшена, яка проявляється
псевдогіпертрофією м`язів ніг у хлопчиків, зниженням їхнього інтелекту та
рухливості. Є мутації локалізованих в Х - хромосомі генів, які недоумкуватості
(синдром В. О. Бер’яєссона); розвиток сімейного пігментного невуса, який проявляється
вогнищами гіперпігментації шкіри та аномаліями в розвитку нігтів, зубів,
роговиці кришталика очей; ці ж гени спричиняють відсутність потових залоз;
курячу сліпоту тощо. Описано майже 200 генів, локалізованих в Х - хромосомах,
мутації яких успадковуються за крис-крос схемою і обумовлюють аномалії розвитку
відповідних ознак.
Що стосується генів, локалізованих в
У - хромосомі, то вони представлені двома групами. Гени однієї з них
локалізуються в ділянці У - хромосоми, яка гомологічна до відповідної ділянки Х
- хромосоми; ці гени й обумовлені ними ознаки успадковуються за такою самою
схемою як і аутосомні гени. Гени другої групи, локалізовані на ділянці, яка не
має на Х - хромосомі відповідних гомологічних точок, успадковуються лише від
батька до сина. Наприклад, у людини в У - хромосомі виявлено 14 генів. З них
дев’ять локалізовано на ділянці, яка гомологічна до такої ж ділянки на Х -
хромосомі, а п’ять - на ділянці, яка не має гомологічних точок на Х -
хромосомі. Це так звані голандрічні гени, тобто гени, які в нормі передаються
тільки від батька до сина.
Явище успадкування ознак, зчеплених
статевими хромосомами, ефективно використовується в птахівництві, шовківництві
тощо.
Нерозходження статевих хромосом
порушує співвідношення, які еволюційно склалися між числом гаплоїдних наборів
аутосом та числом статевих хромосом. Фенотипово це проявляється у дрозофіли у
вигляді перерозвитку або недорозвитку первинних статевих ознак. Виникають
інтерсексуальні (проміжні у статевому відношенні) нащадки, а також надсамці і
надсамки. Ці факти свідчать, що не тільки в статевих хромосомах, але і в
аутосомах є гени, які визначають розвиток жіночої або чоловічої статі.
Кількісні співвідношення між хромосомами, які несуть статеві гени, К. Вріджес
(1939) назвав статевим індексом. Він установив, що в дрозофіли формування
первинних і вторинних статевих ознак визначається величиною статевого індексу.
Наприклад:
Х - хромосоми = 1- нормальна самка;
набори аутосом
Х - хромосома =
½ - нормальний самець;
набори аутосом
Х - хромосоми = 3/2 - надсамка;
набори аутосом
Х - хромосоми = 2/3 -
інтерсексуальна особина;
набори аутосом
Х - хромосома = 1/3 - надсамець;
набори аутосом
Х - хромосоми = ¾
- інтерсексуальна особина;
набори аутосом
Експериментально встановлені статеві
індекси для плодових мух показують, що формування чоловічої чи жіночої статі в
цих комах, безумовно, контролюється генами. Були зроблені висновки, що гени,
які локалізуються в аутосомах, обумовлюють чоловічу стать, а гени,
локалізуються в статевих хромосомах - жіночу. Між генами чоловічої і жіночої
тенденції існує рівновага (баланс). Відхилення у цьому балансі спричиняє
утворення аномалій у формуванні первинних та вторинних статевих ознак. При
цьому У - хромосома дрозофіли не впливає на формування статі, про що свідчить формування
ХО - зигот фенотипово нормальних самців, хоч вони статево стерильні.
Підсумовуючи результати своїх досліджень, Бріджес дійшов висновку про існування
генного балансу між сукупностями генів чоловічої та жіночої тенденції. Згідно з
цією гіпотезою, організм відносно статевої диференціації є біпотенціальним. Він
містить у складі свого генотипу гени, що обумовлюють розвиток як чоловічої, так
і жіночої статі, і стать того чи іншого нащадка дрозофіли обумовлюється
балансом між числом статевих хромосом та числом наборів аутосом.
Те, що стать детермінується
генетично, підтвердилось також на статевих ознаках метеликів непарного
шовкопряда Lymantria dispar. У дослідах Р. Гольдшмідта (1923) від схрещування
самок непарного шовкопряда європейської раси із самцями японської раси всі
нащадки жіночної статі виявлялись інтерсексуальними, а самці - нормальними, але
від схрещування самок шовкопряда, виявлених в районі м. Токіо, із самцями,
взятими з популяції о-ва Хокайдо, всі нащадки були лише жіночої статі, хоча половина
з них характеризувалась чоловічим ZZ - каріотипом, а половина жіночим ZW -
каріотипом.
Таким чином тенденція чоловічої
статі, яку привносять японські самці в генотипі яйцеклітин європейських самок
проявляє проміжне домінування з жіночою тенденцією цих генотипів, в результаті
чого замість жіночих особин формуються інтерсекси. Разом з цим жіноча тенденція
самок шовкопряда токійської популяції настільки сильно домінує над чоловічою
тенденцією самців з популяції о. Хокайдо, що навіть в особин із ZZ - каріотипом
формується не очікувана чоловіча стать, а жіноча.
Якщо статеві індекси в дрозофіли
показують, що в її аутосомах локалізуються гени чоловічої, а в Х - хромосомах -
жіночої статі, то в непарного шовкопряда, навпаки, гени чоловічої статі
локалізуються в статевих Z - хромосомах, а жіночої - в аутосомах.
Бріджез з’ясував, що ступінь його
прояву залежить від умов середовища. Наприклад, розвиток зигот у триплоїдних
самок в умовах підвищених температур зміщується в бік жіночої статі, а в умовах
понижених температур - в бік чоловічої статі.
Як уже зазначалось, статеві гени в
організмах дрозофіл та непарного шовкопряда локалізуються в аутосомах та
статевих Х - чи Z - хромосомах. Що ж до У- чи W - хромосом, то у відповідних
видів комах ці хромосоми не містять у собі статевих генів. Мабуть цим
пояснюється те, що у разі гетерогаметності чоловічої статі ХО - зиготи
започатковують розвиток чоловічих особин, а при гетерогаметності жіночої статі
ZО - зиготи обумовлюють розвиток самок. Проте в інших видів організмів, наприклад,
у ссавців, У - хромосома, незважаючи на низьку генетичну активність, відіграє
суттєву роль у генетичних механізмах визначення статі. Наприклад, у людини
особини з ХО-, ХХ-, ХХХ-, ХХХХ-каріотипами завжди представлені жіночою статтю,
а особини, з ХУ-, ХХУ-, ХХХУ-, ХХХХУ- каріотипами - чоловічою. Аналогічна роль
У - хромосоми проявляється в деяких дводомних видів рослин, наприклад, у
Melandrum album.
Е. Ейгвалд і К. Сілмсер (1955)
виявили в крові мишей з Х - каріотипом наявність Н - антигена, але проявляється
цей антиген лише в крові тварин з ХУ - каріотипом. У подальшому з`ясувалось, що
в У - хромосомі немає гена, котрий видавав би команду на синтез Н - антигена.
Такий ген, як було встановлено, локалізований в одній з аутосом, а У -
хромосома лише видає генетичну інформацію на синтез генних продуктів, які
депресують локалізований в аутосомі структурний ген, включаючи його в активне
функціонування. Внаслідок цього відбувається синтез Н - антигена.
Балансова гіпотеза визначення статі
цілком підтвердилась в дослідах на багатьох видах організмів. Вона дозволяє
з`ясувати бісексуальну природу, причини інтерсексуальності, гермафродитизму та
гінандроморфізму в різних видів організмів, котрі використовуються як об’єкти
відповідних досліджень.
Розглядаючи механізми визначення
статі у високоорганізованих істот, ми дійшли висновку, що формування чоловічої
чи жіночої статі обумовлене функцією двох протилежно діючих груп статевих
генів. Якщо гени однієї статі локалізуються в аутосомах, то гени протилежної
статі - в статевих хромосомах. При цьому стать формується внаслідок домінування
генів однієї статі над сумарною дією генів протилежної статі. Можливість
порушення рівноваги в балансі статевих генів пояснюється тим, що під впливом
умов зовнішнього середовища може збільшуватись або зменшуватись сила
домінування генів однієї статі відносно іншої, і саме цим фактором
обумовлюється формування інтерсексів, надсамців, надсамок чи інших аномалій.
Виникнення таких аномалій посилює дію природного добору в напрямку формування
неоднакової генетичної активності, а також різних за формою статевих хромосом.
Внаслідок цього статеві хромосоми,
на відміну від аутосом, як при спорідненому, так і при перехресному
розмноженні, не можуть перейти в гомозиготний стан за обома статями. Одна з них
(чоловіча або жіноча) в нормі завжди має залишатись гетерогаметною. Ця
особливість статевих хромосом спричинила виникнення та становлення статі.
В нижчих рослин (наприклад, у
грибів, водоростей), на відміну від інших рослин чи тварин, відсутні будь-які первинні
або вторинні статеві ознаки, а морфологічна диференціація гамет буває ледве
помітна або і взагалі відсутня. Тому практично дуже важко зробити висновок, які
з гамет слід віднести до чоловічої, а які до жіночої статі. З цієї причини в
таких організмів гамети відрізняють між собою не за ознаками жіночої або
чоловічої статі, а розподіляють по з’єднувальних або статевих групах зі знаком
“+” чи “ - “. Зрозуміло, що запліднення може відбуватися лише при злитті гамет,
які належать до різних статевих груп. Однак і тут стикаємося з цікавими
особливостями. Вони полягають у тому, що, в нижчих організмів, крім
бісексуальної потенції (як у вищих організмів), іноді проявляється
тетрасексуальна потенція, обумовлена взаємодією двох пар неалельних генів,
котрі при розмноженні вступають у різні комбінації між собою.
Наприклад, у деяких базидіальних
грибів виявлено дві пари неалельних генів А та В, які обумовлюють формування
чотирьох типів гаплоїдних спор АВ-, Ав-, аВ-, та ав-генотипів. Ці спори
проростають, започатковуючи формування гаплоїдних міцеліїв. Якщо клітини
міцеліїв АВ-генотипу з`єднуються з клітинами ав-генотипу або клітини
Ав-генотипу з`єднуються з клітинами а-генотипу, то формуються плодові тіла
АаВв-генотипу, тим часом як за інших генотипових комбінацій цих міцеліїв
(наприклад, АВ- з Ав- чи аВ- або ав- з Ав- чи аВ) плодові тіла не утворюються.
Отже утворення плодових тіл лише АаВв_ генотипу детермінується генотипами
статевих груп міцеліїв.
Слід визначити, що з’єднувальні, або
статеві, групи АВ-, Ав-, аВ- та ав-генотипів у процесі злиття клітин міцеліїв
відповідно до законів Менделя могли б утворити в складі генотипів зигот
плодових тіл 16 комбінацій домінантних і рецесивних АВ- генів. Однак із
теоретично очікуваних 16 комбінацій гамет лише чотири забезпечують утворення
дигетерозиготного АаВв-генотипу, і лише він виявляється життєздатним.
Разом з цим з`ясувалось, що у
грибів, у яких виявлена тетрасексуальна потенція, кожний з їхніх А- та В- генів
у сукупностях цих рослин буває представлений серією множинних алеломорфів,
наприклад, А1, А2, А3, … Аn або В1, В2, В3, … Вn. При цьому А1В1 міцелію,
з’єднуючись з клітинами А2В2 міцелію, утворюють плодові тіла А1А2В1В2 або
А1А3В1В2 - генотипів тощо. Таким чином, до складу генотипів плодових тіл можуть
входити гени із серії множинних алеломорфів різної сили домінування. Про різну
силу домінування статевих генів свідчать також спостереження, проведені над
іншими видами організмів. Наприклад, одноклітинні водорості хламідомонади в
природі представлені гаплофазою. Клітини будь-якого таксономічного виду [AK1]
[AK2] цих водоростей морфологічно між собою [AK3] [AK4], хоч вони відносяться
до “+” і “-” - статевих груп.
Після копуляції (злиття) “+” та “-”
статевих клітин утворюється диплоїдна зигота. Вступаючи в мейотичний поділ, вона
утворює чотири дочірні, гаплоїдні клітини. Дві з них виявляються “+”, а дві
інші - “-” статевими клітинами. Однак у даному випадку справа ускладнюється
тим, що в межах кожної статевої групи клітини можуть відрізнятися між собою
силою статевої потенції. При цьому, наприклад, “+” статеві клітини з високою
статевою потенцією можуть вступати в копуляцію і утворювати зиготу з тими “+” -
статевими клітинами, які мають низьку статеву потенцію. Аналогічна картина
спостерігається і в групі “-” статевих клітин. Таким чином, будь-яка клітина із
сукупності “+”- або “-” - статевих клітин деяких ізогамних форм хламідомонад
може поводитись і як “чоловіча” і як “жіноча” гамета залежно від того, з яким
партнером за силою потенції вона зустрілась.
У найпростіших одноклітинних
інфузорій-туфельок проявляється до восьми морфологічно однакових, але
генотипово різних статевих форм. І тут клітини однієї статевої форми можуть
зливатися з клітинами, що належать до буд-якої іншої статевої форми.
Підсумовуючи викладене, можна
зробити висновок, що однією з особливостей механізмів визначення статі в
низькоорганізованих видів організмів є їхня здатність утворювати замість двох
очікуваних по декілька статевих груп з неоднаковою статевою потенцією. При
цьому простежується чітко виражена тенденція до заміни еволюційним шляхом
полісексуальності на бісексуальність. Якщо в інфузорій та хламідомонад ця
тенденція проявляється не дуже чітко, то в деяких базидіальних грибів генетичні
механізми, які лежать в основі такої тенденції, досить очевидні. Про це
свідчать результати злиття гаплоїдних клітин міцеліїв, які належать до чотирьох
статевих груп. Щоб краще зрозуміти наведені нижче схеми, запам`ятаємо, що
кожний домінантний А- чи В- ген обумовлює формування “+” - статевих клітин, а
рецесивний - “-” - статевих. Якщо до складу генотипу привноситься “+” і “-”
статеві гени з однаковою статевою потенцією, то, взаємодіючи, вони
нейтралізують один одного, і це обумовлює розвиток статево нейтрального
плодового тіла. Однак якщо після запліднення сила статевої потенції клітин
однієї статі переважає силу статевої потенції клітин протилежної статі, то
плодові тіла не утворюються.
Як бачимо, генні мутації, які
визначають гетерозиготний та гетерогенний стан у популяціях гаплоїдних
організмів, обумовлюють виникнення в них статевих груп. Спроба цих видів
організмів перейти на диплоїдний рівень закінчується невдачею, оскільки
генотипи, незбалансовані за статевими клітинами, виявляються нежиттєздатними.
Проте зиготи, в генотипах яких сума “+” та “-” - статевих генів врівноважується,
будуть життєздатними, хоча й статево нейтральними.
Узагальнюючи накопичені наукою
факти, можна визначити, що в нижчих еукаріотів (водоростей, грибів, інфузорій)
відсутні ознаки, за якими можна було б встановити чоловічу або жіночу стать.
Тому в них гамети умовно поділяють на “+” чи “-” - статеві групи. Вони
ізоморфні. Статеві групи виникають унаслідок мутації генів у генотипах
відповідних видів організмів, однак статеві мутації в генотипах клітин
гаплоїдних організмів самі по собі (поки вони не залучені в процеси генетичної
рекомбінації) суттєво не змінюють закономірностей життєдіяльності цих
організмів. Таким чином, можливість необхідної перекомбінації генів у нижчих
еукаріотів забезпечується вже існуючими механізмами мейотичної редукції числа
хромосом, яке подвоюється в процесі злиття двох гамет “+” та “- ” - статі.
Механізми мейозу, що виникли в
процесі еволюції, забезпечували не тільки редукцію числа хромосом, але й
генетичну рекомбінацію в максимально можливих обсягах. Оскільки матеріальною
основою процесів мейозу є хромосоми, формування їх у процесі еволюції мало
супроводжуватись виникненням мітозу та мейозу. В протилежному випадку
становлення хромосом було б неможливим. Отже процеси еволюційного виникнення та
формування хромосом, а також механізмів мітозу, мейозу і статевих
взаємовідносин між гаплоїдними клітинами в популяціях відбувались паралельно
під впливом мутантних генів “+” та “-” - статі.
Слід визначити, що в
низькоорганізованих видів організмів, які належать до еукаріотів, статеві
хромосоми не виявлені, оскільки вони ще не сформувались при еволюції. Проте у
вищих організмів наявність гетероморфних статевих хромосом проявляється чітко.
Наприклад, сім аутосом і одна статева (Х- або У) - хромосома в гаплоїдних
клітинах як чоловічих, так і жіночих рослин печіночника Sphaerocarpus; по шість
аутосом та одній статевій (Х чи У) хромосомі в гаметах клопа Lygaeus; по три
аутосоми та одній статевій хромосомі в гаметах дрозофіли тощо.
Вважають, що виникнення і формування
статевих хромосом відбувається за рахунок пари гомологічних хромосом. При цьому
еволюційний розвиток проходить у напрямку поступової втрати генетичної
активності однієї з двох аутосом-гомологів зазначеної пари. В ряді випадків цей
напрямок еволюції приводить до повної втрати генетично неактивної хромосоми,
як, наприклад, у трав’яних клопів, більшості прямокрилих, в багатьох видів
нематод тощо.
Проте в процесі еволюції статевих
хромосом не завжди відбувається втрата одного з двох партнерів. Наприклад, у
ссавців, двокрилих видів комах, багатьох видів риб, дводомних видів рослин
поряд з генетично високоактивною Х - хромосомою функціонує й менш активна У -
хромосома, без якої неможливо одержати нащадків.
Формування статевих хромосом у
процесі еволюції супроводжувалось виникненням статевого диморфізму. Нагадаємо,
що в нижчих еукаріотів статеві групи формуються внаслідок мутацій у генотипах
гаплоїдних клітин. тому ці групи є ізоморфними, їхня статева потенція -
моносексуальна. При злитті двох клітин, які належать до “+” та “-” статевих груп,
виникає диплоїдна зигота з бісексуальною статевою потенцією. Зрозуміло, що у
вищих організмів вона може змінюватись і обумовлювати розвиток чоловічої або
жіночої статі (залежно від дози відповідних статевих генів, яка регулюється
наявністю однієї чи двох Х- або Z - хромосом, які ввійшли до складу зигот при
заплідненні).
Аналізуючи напрямки дії природного
добору в процесі еволюційного розвитку і становлення статі, можна зробити
висновок, що, використовуючи та вдосконалюючи генотипові й каріотипові засоби,
природа шляхом еволюційного формування статі створила досить досконалі
механізми генетичної рекомбінації та збільшила дозу кожного гена в генотипах
еукаріотів, замінивши гаплофазу диплофазою. Саме ці фактори в десятки і в сотні
разів прискорили еволюцію видів живих організмів на Землі.
В 1949 р. М.Л. Барр і Е. Г. Бертрам,
вивчаючи морфологію нейронів у котів, виявили відмінність між цими клітинами в
самців та самок. Було встановлено, що в нейронах самок на внутрішньої поверхні
ядерної оболонки локалізуються грудочки речовини, які активно забарвлюються
ядерними барвниками, а в нейронах самців вони відсутні. Численні дослідження
показали, що грудочки з`являються від конденсації, тобто від щільної
компактизації хромосомної нитки в одній з двох Х - хромосом. Тому ці грудочки
дістали назву статевого хроматину. Інша назва - тільця Барра (на честь їхнього
першовідкривача).
Було встановлено, що статевий
хроматин проявляється не тільки в нейронах, але й в інших типах клітин.
наприклад, у людини він присутній в клітинах слизових оболонок тощо. Крім
людини та кішок статевий хроматин притаманний і багатьом іншим видам ссавців.
При чому він проявляється в нормі лише в особин жіночої статі. Цікаво
відзначити, що в чоловіків, хворих на синдром Клайнфельтера (ХХУ - каріотип) у
нейронах та клітинах слизових оболонок статевий хроматин чітко проявляється, а
в клітинах жінок, хворих на синдром Тернера (ХО - каріотип), він відсутній.
Якщо до складу каріотипу людини
входять ХХХ - хромосоми (жінки, іноді з розумовою відсталістю) або ХХХУ -
хромосоми (чоловіки з глибоким ураженням синдромом Клайнфельтера), то в ядрах
клітин цих людей проявляється по два тільця Барра. Таким чином, наявність чи
відсутність У - хромосоми не впливає на конденсацію “зайвих” Х - хромосом у
складі каріотипу. Наявність чи відсутність певної кількості тілець Барра в
клітинах людей має діагностичне значення, оскільки за цим показником можна
швидко і безболісно виявити кількість Х - хромосом у складі каріотипів
аутосомно диплоїдних особин. Бо й справді, кількість статевих Х - хромосом у
складі каріотипу особини жіночої статі дорівнює кількості тілець Барра, які
проявляються в окремих клітинах цієї особини, полюс одиниця. Наприклад, якщо в
клітинах людини налічується три тільця Барра, то в каріотипі цієї особини міститься
3 + 1 = 4 Х - хромосоми. Якщо ж ця особина чоловічої статі, то крім чотирьох Х
- хромосом в її каріотипі має міститись і У - хромосома.
Отже, можна вважати, що відсутність
тілець Барра в клітинах особин жіночої статі свідчить про наявність у складі її
каріотипу (0 + 1) лише Х0 статевих хромосом. Якщо ж в клітинах чоловічого
організму виявлено одне тільце Барра, це означає, що в складі його каріотипу (1
+ 1) містяться ХХУ - статеві хромосоми, і так далі.
Природно виникає питання, чому в
здорових клітинах здорового жіночого організму конденсується одна з двох Х -
хромосом? На думку багатьох вчених це явище спричинене необхідністю дозової
компенсації генів. Згадаємо, що в організмів гетерогаметної статі на диплоїдний
набір аутосомних генів припадає лише гаплоєдний набір генів, локалізованих в Х
- або Z - хромосомах. Разом з цим в організмів гомогаметної статі на диплоїдний
набір генів, локалізованих в аутосомах, припадає диплоїдний набір генів,
локалізованих у двох Х - хромосомах. Можна вважати, що за цієї причини в
процесі еволюційного розвитку статі природний добір діяв у напрямку формування
рівновеликого загального генетичного балансу між аутосомними генами та генами,
локалізованими в статевих хромосомах особин як чоловічої, так і жіночої статі.
Одним з механізмів досягнення такого стану і є конденсація, яка обумовлює
припинення функціонування однієї з двох Х- або однієї з двох Z - хромосом. На
думку Г. Меллера (1931), в Х - хромосомі є ген-компенсатор, котрий в каріотипах
ХХ - особин, будучи в подвійному числі, пригнічує активність генів однієї з
двох Х - хромосом. Цим самим досягається однаковість взаємодії доз аутосомних
генів з генами, локалізованими в Х - хромосомах особин жіночої і чоловічої
статі.
РОЗДІЛ 2.
Об’єкти
та методика досліджень
Об’єктом наших досліджень був
класичний об’єкт генетики - дрозофіла. На сьогодні лише один організм серед
еукаріотів - плодова мушка (Drosophila melanogaster Mg.) - вважається досить
добре генетично вивченою. Вперше як об’єкт генетичних досліджень її було використано
у 1910 році Томасом Морганом та його учнями. За даними Р. Рефа та Т. Кармена
(1986) сучасний каталог мутацій Drosophila melanogaster Mg. налічує 500
сторінок, на яких описано більше ніж 2000 мутацій.
Основні особливості Drosophila
melanogaster Mg., які роблять її універсальним об`єктом генетичних досліджень:
Короткий життєвий цикл (час розвитку
від яйця до імаго при оптимальних умовах - 10 діб).
Висока плодючість (150 особин від
однієї самки).
Висока кількість спадкових рас
(мутацій), знайдених у культурах в лабораторних умовах.
Мала кількість хромосом (4 пари).
Генетична вивченість (складені
генетичні карти).
У наш час Drosophila melanogaster Mg
- космополіт. На території нашої країни дрозофіла живе як у природних, так і в
лабораторних умовах.
Систематичне положення
Царство - Animalia (тварини)
Підцарство - Metazoa
(багатоклітинні)
Тип - Arthropoda (членистоногі)
Підтип - Trachiata (трахейні)
Клас - Insecta (комахи)
Підклас - Entognata (покриточелюсні)
Ряд - Diptera (двокрилі)
Підряд - Brachycera (коротковусі)
Родина - Drosophilidae (дрозофільні)
Рід - Drosophila (дрозофіла)
Вид - Drosophila melanogaster
melanogaster Mg (плодова чи оцтова муха) - муха довжиною до трьох міліметрів, з
сірим забарвленням тіла та червоними очима. Це комаха з повним циклом
перетворення. Цикл розвитку при оптимальній температурі, що складає 23 -25 оС,
від яйця до імаго - 10 діб:
Ембріональний розвиток - 2 доби
Розвиток личинки - 4 доби
Розвиток лялечки - 4 доби.
Тривалість життя дорослої мухи в
лабораторних умовах становить 3 -4 тижні і залежить від температури, вологості,
їжі, густоти населення та ін. Статева зрілість самців і самок настає на другий
день після вилуплення комахи. У природі дрозофіла живиться ферментуючими
фруктами та овочами. В лабораторних умовах Drosophila melanogaster Mg вирощують
на поживному середовищі такого складу:
На 100 мл води потрібно взяти 1,2 -
1,5 г агар-агару, 12 - 15 г дріжджів, 5 г цукру, 5 г манної крупи.
Методика приготування поживного
середовища.
Розчинити агар-агар у воді та
довести до кипіння
Долити до цього розчину попередньо
розчинені у воді дріжджі
Кип’ятити 35 - 40 хвилин,
перемішуючи
Досипати до розчину, перемішуючи,
цукор та манну крупу
Кип’ятити цей розчин 10 - 15 хв.
Поживне середовище розділити по
пробірках
Після того, як середовище затвердіє,
залити його дріжджами (60 г сирих та 18 г сухих)
Усі маніпуляції з дрозофілами
виконують під наркозом. В кожну пробірку з поживним середовищем саджають по дві
самки і два самця. Вивчення батьківських форм та одержання гібридів, а також
порівняння між собою різних мутаційних рас проводиться за допомогою бінокуляра
МБС - 9 із збільшенням у чотири рази. Для підрахунку результатів необхідно мати
нескладне обладнання: морилку, столик з матовим склом, загострене пір’я,
калькулятор і т. ін.
Мета наших експериментів - збагатити
кафедральну колекцію класичного генетичного об`єкту - Drosophila melanogaster
Mg двома лініями мух, які мають у генотипі мутантні гени “white” та “cut”.
Для досягнення цієї мети нами були
поставлені такі задачі:
провести серію спланованих
схрещувань (до F4), в результаті яких відбувається генетична рекомбінація;
при аналізі нащадків провести добір
необхідних мутантних особин;
шляхом схрещування мутантних особин
F2 з гетерозиготними генотипами “white” і “cut” отримати такі ж форми, але у
гомозиготному стані;
провести статистичну обробку
дослідних даних та підтвердити справедливість отриманих результатів.
У дослідах були використані зчеплені
рецесивні мутації першої хромосоми Drosophila melanogaster Mg (х -
хромосома)(w) - білі очі(ct) - крила вкорочені, з вирізками
Вивчені нами мутації відкриті до
1916 р і описані Т. Морганом. Мутація White (w) - локус 1,5 - білі очі
відноситься до перших зчеплених зі статтю генів, відкритих Т. Морганом у 1910
році. Мутація Cut (ct) - локус 20,0 - “зрізанокрилі”; задній край крил косо
зрізаний і загострений.
У роботі ми визначаємо частоту
кросинговеру (rf) між досліджуваними генами за формулою:
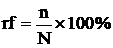 , де
, де
rf (recombination freqency) -
величина кросинговера;- кількість кросоверних особин у досліді;- загальна
кількість особин
Відстань між генами на карті
вимірюється в так званих одиницях карти. Кожна така одиниця відповідає частоті
рекомбінацій 1 %. Таким чином, якщо частота рекомбінацій між двома генами
становить 5 %, то відстань між ними дорівнює 5 одиницям карти.
Похибку rf визначаємо за
формулою:
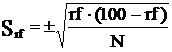 %, де
%, де
rf - частота кросинговера (в
%)- загальна кількість особин у досліді
Результати генетичного
аналізу статистично обробляємо за допомогою крітерія хі - квадрат. Хі - квадрат
розраховуємо за формулою:
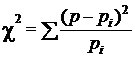 ,де
,де
р - емпірична частота;-
теоретична частота
РОЗДІЛ 3.Результати та
обговорення
.1 Вивчення взаємодії
рецесивних зчеплених генів white i cut
Аналіз сумісного успадкування
декількох ознак проводять з метою визначення локалізації генів, контролюючих ці
ознаки, належності їх до одних чи різних груп зчеплення, виявлення типів
взаємодій між генами. Вивчення сумісного успадкування декількох ознак - перший
етап роботи, який передує визначення локалізації генів у певній групі зчеплення
й картування хромосом.
При схрещуванні самок
дрозофіли з білими очима та зрізаними крилами з нормальними самцями (дикий тип)
в F1 всі самки (351 особин) виявилися диким типом, а самці мали ознаки матерів
- білі очі та зрізані крила. Вже в F1 ми спостерігали успадкування ознак по
типу кріс-крос. На основі цього можна зробити висновок, ознаки, що вивчали,
обумовлені генами, локалізованими в Х - хромосомі, тобто зчеплені зі статтю.
Дані по дням спостереження подані у таблиці 1.
Таблиця 1
Динаміка виходу гібридів F1
|
№ пробір.
|
Дні аналізу
|
|
1
|
2
|
3
|
4
|
5
|
Всього
|
|
♀
|
♂
|
♀
|
♂
|
♀
|
♂
|
♀
|
♂
|
♀
|
♂
|
|
|
1
|
5
|
1
|
14
|
12
|
12
|
7
|
4
|
12
|
5
|
9
|
81
|
|
2
|
26
|
4
|
21
|
10
|
23
|
14
|
9
|
4
|
6
|
139
|
|
3
|
30
|
17
|
12
|
9
|
1
|
4
|
3
|
1
|
5
|
9
|
91
|
|
4
|
4
|
3
|
4
|
8
|
4
|
3
|
14
|
12
|
5
|
8
|
65
|
|
5
|
17
|
16
|
6
|
8
|
4
|
9
|
9
|
13
|
12
|
13
|
107
|
|
6
|
21
|
8
|
21
|
12
|
10
|
4
|
1
|
4
|
1
|
5
|
87
|
|
7
|
17
|
16
|
9
|
9
|
4
|
5
|
8
|
12
|
6
|
12
|
98
|
|
Всього
|
120
|
65
|
87
|
68
|
58
|
46
|
48
|
76
|
38
|
62
|
351 ♀ 317 ♂
|
|
♀+ ♂
|
351 ♀ + 316 ♂ = 668
|
Те що в F1 спостерігається
одноманітність серед самок і самців, це дозволяє нам зробити такі висновки:
батьківські форми за генотипом гомозиготні (самки гомозиготні, а самці
гемізиготні).
Проводячи аналіз гібридів F1, ми
звернули увагу на те, що в перші дні спостерігається більший вихід самок, ніж
самців. Експериментальні дані по F1 представлені в таблиці 2.
Таблиця 2
Вихід самок і самців Drosophila
melanogaster (гібриди F1)
|
№ п/п
|
w+w+ct+ct+ ♀
|
wct ♂
|
Всього
|
|
особин
|
%
|
особин
|
%
|
|
|
1
|
120
|
64,9
|
65
|
35,1
|
185
|
|
2
|
87
|
56,1
|
68
|
43,9
|
155
|
|
3
|
58
|
55,8
|
46
|
44,2
|
104
|
|
4
|
48
|
38,7
|
76
|
61,3
|
124
|
|
5
|
38
|
38,0
|
62
|
62,0
|
100
|
|
Всього
|
351
|
|
317
|
|
668
|
Аналіз даних, які подані в таблиці
1, показав, що вихід самок і самців по днях коливається. В перші дні вихід
самок більший, ніж самців. В подальшому вихід самок зменшується, а самців -
збільшується. Таку особливість виходу дорослих самок і самців, вірогідно, можна
вважати генетичною особливістю мух.
Самки в F1 гетерозиготні, тобто
мають у своєму генотипі один домінантний алель. Самці гемізиготні за двома
рецесивними алелями. Рецесивні алелі виникли внаслідок мутацій. Більшість
мутацій характеризується зниженою життєздатністю порівняно з особинами дикого
типу. Кількість самок (гетерозиготи) більше кількості самців (гемізиготи за
двома рецесивними алелями).
Як вже сказано вище, в F1
спостерігається кріс-крос успадкування. В такому випадку розщеплення за статтю
теоретично можна очікувати у співвідношенні 1 : 1. В таблиці 2 наводяться дані
по самках і по самцях. Результати оброблені методом Хі- квадрат і достовірні з
вірогідністю 95 %. Отримано такі результати та розрахунки наведені у таблиці 3:
Таблиця 3.
Відповідність експериментальних
даних теоретично очікуваним
|
Дані
|
Стать
|
|
Жіноча
|
Чоловіча
|
|
Експериментальні дані р
|
351
|
317
|
|
Теоретичні дані рі
|
334
|
334
|
|
р - рі
|
17
|
- 17
|
|
(р - рі)2
|
289
|
289
|
|
(р - рі)2 / р
|
0,43
|
0,43
|
|
χ2
|
0,43 + 0,43 = 0,86
|
χ2емпір
= 0,86
χ2 теор.
= 3,8
Р = 0,95= 1
χ2 теор.>
χ2 емпір.
Висновок: розщеплення у F1 у
співвідношенні 1 :1 доведено. Ці дані статистично достовірні.
Проаналізувавши гібриди F1
дрозофіли, можна зробити такі узагальнення:
Гени, які зумовлюють ознаки, що
вивчались, (колір очей, форма крил) зчеплені зі статтю.
Розщеплення за статтю 1 : 1 доведено
за допомогою критерію χ2.
Батьківські форми за генотипом:
самки гомозиготні по рецесивним алелям, а самці гемізиготні по домінантним
алелям.
. Для отримання F2 нам потрібно
схрестити самок із F1 (червоні очі, круглі крила): Хw+ct+Хwct
з самцем із F1 (білі очі, зрізані
крила): ХwctY
Жіночі особини дають два
некросоверних та два кросоверних гамет:
|
w
|
w+
|
w
|
w+
|
|
ct
|
ct+
|
ct+
|
ct
|
|
некросоверні
|
кросоверні
|
Чоловічі особини дають 2 види гамет,
тільки некросоверні, бо кросинговер в них не проходить:
Самки за фенотипом розділились на 4
класи. Генотипи та фенотипи самок:
1.
XwctXwct 2. Xw+ct+Xwct 3. Xwct+Xwct 4. Xw+ctXwct
білі
очі червоні очі білі очі червоні очі
зрізані
крила округлі крила округлі крила зрізані крила
476 115
(к) 108 (к)
всього - 1163 особин у досліді
Самці також утворюють 4 класу
фенотипів. Генотипи та фенотипи самців:
1.
XwctY 2. Xw+ct+ Y 3. Xwct+Y 4.
Xw+ctY
білі
очі червоні очі білі очі червоні
очі
зрізані
крила округлі крила округлі крила зрізані крила
475 109
(к) 104 (к)
всього - 1146 особин у досліді
Отримані дані приведені у таблиці 4
Таблиця 4.
Результати аналізу успадкування
мутацій white i cut (F2)
|
♀ Жіночі особини
|
♂ Чоловічі особини
|
|
XwctXwct
|
476
|
XwctY
|
475
|
|
Xw+ct+Xwct
|
464
|
Xw+ct+Y
|
458
|
|
Xwct+Xwct (к)
|
108
|
Xwct+Y(к)
|
104
|
|
Xw+ctXwct (к)
|
115
|
Xw+ctY(к)
|
109
|
|
Разом
|
1163
|
|
1146
|
|
Всього чоловічих та жіночих особин у досліді
|
2309
|
n = 108 + 115 + 104 + 109 = 436=
2309
 = 18,8 % (морганид)
= 18,8 % (морганид)
Третій етап нашої роботи -
отримання F3 .
Проводимо експеримент за
двома варіантами:
отримання мутації white
отримання мутації cut
) Отримання мутації white
Із F2 відбираємо самок і
самців з генотипами: +Xwct (білі очі, округлі крила) Xwct+Y (білі очі,
округлі крила) та проводимо між ними схрещування
Для отримання гомозиготних
самок з генотипом та фенотипом Xwct+Xwct+ - білі очі (white), округлі крила
(cut+) у пробірку сажаємо по одній самці і одному самцю, з цими генотипами. В
тій пробірці, де серед нащадків не буде розщеплення, будуть гомозиготні жіночі
особини, та дигемізиготні самці.
) Отримання мутації cut
Із F2 відбираємо самок та
самців з генотипами та фенотипами:+ctXwct (червоні очі, зрізані крила) Xw+ctY
(червоні очі, зрізані крила) та проводимо схрещування для отримання
гомозиготних самок з генотипом та фенотипом Xw+ctXw+ct - червоні очі (white+),
зрізані крила (cut).
Отримані експериментальної
дані у дипломній роботі дозволяють зробити такі висновки:
між генами white i cut
просліджується неповне зчеплення, так як в результаті схрещування виникають
кросоверні особини.
Експериментально визначена
відстань між генами white i cut, яка дорівнює 18,8 морганід, що дуже близько до
значення між генами на хромосомній карті Drosophila melanogaster 18,5 морганід.
За допомогою гібридологічного
метода зроблено розчеплення хромосомної мутації по Х - хромосомі white - cut.
Отримані дві одинарні мутації
- white - білі очі та cut - зрізані крила.
Розроблені методичні
рекомендації що до використання дипломної роботи у шкільному курсі біології
РОЗДІЛ 4.Можливості
використання матеріалів даної дипломної роботи у школі
.1 Місце даної теми у
шкільних програмах з біології
Проаналізувавши шкільну
програму з біології для старших класів, ми з’ясували, що викладання курсу
генетики передбачається в одинадцятому класі. Для класів з поглибленим
вивченням біології на генетику приділяється 18 годин, причому розподіл годин
здійснює сам вчитель.
На наш погляд на тему
“Зчеплене успадкування. Кросинговер. Хромосомна теорія спадковості” можна
відводити дві і, як максимум, три години, щоб не зменшити значення інших дуже
важливих тем. А у звичайних класах генетиці приділяється така ж кількість
годин, але тема “Зчеплене успадкування. Кросинговер. Хромосомна теорія
спадковості” обмежена часом і їй можна виділити максимум дві години, бо вона
розглядається в сукупності з іншими важливими темами, які об’єднані загальною
назвою “Універсальні властивості організмів”. Також до цієї теми без додаткового
часу входить практична робота № 2 “Розв’язування типових задач з генетики”.
Таким чином, на наш погляд,
часу, що приділяється генетиці взагалі і на тему “Зчеплене успадкування.
Кросинговер. Хромосомна теорія спадковості” зокрема, недостатньо. Тому якість
засвоєння цього важливого матеріалу у значній мірі залежить від майстерності,
ерудиції та методичної грамотності вчителя.
На підставі проведеного
аналізу та враховуючи безпосередній досвід роботи у школі, ми спробували дати
власні методичні розробки цього складного і дуже цікавого матеріалу.
4.2 Методичні розробки уроків
для вчителя біології старших класів
1. Розробка уроку
Тема: “Явище зчепленого
успадкування. Кросинговер. Хромосомна теорія спадковості”.
Задачі уроку: сформувати
поняття про зчеплене успадкування, кросинговер, групу зчеплення, генетичні
карти хромосом, хромосомну теорію спадковості, виховувати матеріалістичний
світогляд, розвивати вміння виділяти головне, уважно слухати, порівнювати,
систематизувати і робити висновки
Тип уроку: урок формування
нових знань.
Хід уроку
|
I.
Мотивування та актуалізація опорних знань учнів Діти, ми вже з вами
познайомилися із закономірностями успадкування ознак, які були відкриті Г.
Менделем та науково визнані його послідовниками. Ці закономірності відображені
в трьох законах Г. Менделя та правилі чистоти гамет. Але з розвитком
генетики, яка на початку ХХ століття дуже зацікавила не тільки вчених, а й
усіх передових людей, стали накопичуватися факти, які не вкладалися в ці
закономірності. Не було ніякої чіткості в розподілі ознак в наступних
поколіннях і пояснити це явище вчені на той час були не в змозі. Противники
генетики були дуже раді такій ситуації, це давало їм підстави не сприймати цю
науку і вважати її необ’єктивною. Але прогрес припинити неможливо і наукова
школа американського біолога Томаса Гента Моргана захопилася проблемами
спадковості та мінливості. Працювали вчені з дуже вдалим об`єктом для
дослідження цих явищ - дрозофілою меланогастер (Drosophyla melanogaster).
Вчені цієї школи змогли не тільки усі ці питання пояснити, а й розвинути
генетичні знання у цьому напрямку. II. Пояснення нового матеріалу Т.
Морган зі своїми колегами Бріджесом та Стертевантом на підставі результатів
своїх дослідів сформували хромосомну теорію спадковості. Без цієї теорії
розвиток генетики був би неможливий.
|
Розповідь вчителя Розповідь
вчителя
|
|
Якщо Мендель працював на рослинних об’єктах,
то Морган вибрав об’єкт тваринного походження. Він виявився дуже вдалим, так
як має всього 4 пари хромосом, короткий життєвий цикл (два тижні), високу
плодючість (кількість нащадків), його легко розводити в лабораторних умовах.
Основні положення хромосомної теорії спадковості такі: Гени розташовані в
хромосомах; різні хромосоми містять неоднакову кількість генів; кожна з
негомологічних хромосом має свій унікальний набір генів. Гени розташовані
вздовж хромосом у лінійному порядку. Кожен ген займає в хромосомі певну
ділянку (локус); алельні гени заповнюють однакові локуси гомологічних
хромосом. Усі гени однієї хромосоми утворюють групу зчеплення, завдяки чому
відбувається зчеплене успадкування деяких ознак; сила зчеплення між двома
генами обернено пропорційна відстані між ними. Зчеплення між генами,
розташованими в одній хромосомі, порушується внаслідок кросинговеру, під час
якого гомологічні хромосоми обмінюються своїми ділянками. Кожен біологічний
вид характеризується певним каріотипом (повною кількістю і формою хромосом).
Вчені дуже давно звернули увагу на те, що кількість спадкових ознак організму
значно перевищує число хромосом гаплоїдного набору. Це означає, що в кожній
хромосомі розміщено багато генів. Тож разом із ознаками, які успадковуються
незалежно, також поряд існують такі, що успадковуються зчеплено одна з одною,
бо вони визначаються генами, розташованими в одній хромосомі. Такі гени
утворюють групу зчеплення. Кількість груп зчеплення в організмів певного
|
Запис у робочий зошит
учнями Запис у робочий зошит учнів
|
виду дорівнює кількості
хромосом у гаплоїдному наборі (наприклад, у дрозофіли 1n = 4, у людини 1 n =
23). Явище зчепленого успадкування Морган встановив у досліді, як показано на
малюнку 1. 
|
Мал 1. Ілюстрація дослідів Т. Морганана, що
ілюструє явище кросинговеру (на організменному рівні)
|
|
|
На підставі цих даних Морган припустив, що
гени, які визначають забарвлення тіла і форму крил, розташовані в одній
хромосомі і називаються зчепленими. Вони успадковуються разом. Але у природі
існує явище, яке порушує це зчеплення (за рахунок процесів, що відбуваються
при утворенні гамет в мейозі, а саме в профазі I, коли
гомологічні хромосоми можуть обмінюватися ділянками) і називається
кросинговером, або перехрестом хромосом. Це явище розповсюджено в природі і
має дуже велике значення для комбінативної мінливості, що сприяє природному
добору. Наслідки кросинговеру Морган побачив на рівні організму. А
користуючись новітніми методами досліджень, таким, наприклад, як метод
маркування хромосом радіоактивними ізотопами атомів, можна прослідкувати ці
явища і на рівні хромосом, і навіть на генному рівні (малюнок 2). 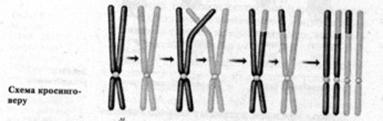 Мал. 2.
Ілюстрація кросинговеру на рівні хромосом. Враховуючи
велике значення кросинговеру для живих організмів, величезною працею вчених
були встановлені такі його закономірності: сила зчеплення між двома генами,
розташованими в одній хромосомі, обернено пропорційна відстані між ними (чим
ця відстань більше, тим частіше відбувається кросинговер); частота
кросинговеру між двома зчепленими генами являє собою відносно сталу величину
для кожної конкретної пари генів. Мал. 2.
Ілюстрація кросинговеру на рівні хромосом. Враховуючи
велике значення кросинговеру для живих організмів, величезною працею вчених
були встановлені такі його закономірності: сила зчеплення між двома генами,
розташованими в одній хромосомі, обернено пропорційна відстані між ними (чим
ця відстань більше, тим частіше відбувається кросинговер); частота
кросинговеру між двома зчепленими генами являє собою відносно сталу величину
для кожної конкретної пари генів.
|
Запис у робочий зошит Запис у робочий
зошит
|
|
|
На підставі цих закономірностей було зроблено
висновок, що гени розташовані в хромосомі по її довжині один за одним (в
лінійному порядку). За допомогою цих встановлених положень генетики навчилися
складати генетичні карти хромосом, які дають нам уявлення про послідовність
розташування генів у хромосомі. Для деяких об’єктів, таких як дрозофіла,
томати, кукурудза, миші, і навіть для людини вже складені повні генетичні
(хромосомні) карти. Це дуже важливо для розвитку таких сучасних напрямків
біології як генна інженерія, культура тканин, медична генетика та інші
напрямки. III.
Узагальнення та систематизація знань Ким була запропонована хромосомна теорія
спадковості і які її основні положення? Що таке група зчеплення? Що таке
зчеплення генів? Або які гени називаються зчепленими? Дайте визначення
кросинговеру та назвіть його основні закономірності. Що таке генетичні карти
хромосом? Який принцип їх складання? На підставі пройденого навчального
матеріалу можна зробити такі висновки: усі гени, розташовані в одній
хромосомі, утворюють групу зчеплення, внаслідок чого ознаки, які визначаються
нами, успадковуються зчеплено. Сила зчеплення між двома генами, розміщеними в
одній хромосомі, обернено пропорційна відстані між ними. Кількість груп
зчеплення в організмів певного виду дорівнює числу хромосом у його геномі.
Зчеплення між генами порушується у результаті кросинговеру (перехресту
хромосом). Частота кросинговеру для кожної пари зчеплених генів відносно
стала та пропорційна відстані між ними.
|
Запис у робочий зошит Фронтальна
бесіда Розповідь вчителя
|
|
|
Явище кросинговеру має велике значення у
природі та для науки. Підрахування величини кросинговеру між генами дає
підстави для побудови генетичних карт хромосом. IV. Домашнє
завдання § 42, питання після параграфу опрацювати.
|
|
|
II. 2. Розробка уроку
Тема: “Вирішення задач з генетики”
Задачі уроку: формувати уміння
застосовувати теоретичні знання з генетики на практиці (для вирішення
генетичних задач); виховувати матеріалістичний світогляд
Тип уроку: урок формування умінь та
навичок
Хід уроку
I. Перевірка домашнього завдання
|
Розповісти про хромосомну теорію спадковості,
назвати її основні положення. Ким була заснована? Що ви можете розповісти про
зчеплене успадкування? Які гени називають зчепленими? Що таке група
зчеплення? Що таке кросинговер? Який біологічний зміст цього процесу? Який
принцип покладений в основу побудови генетичних карт хромосом?Фронтальна бесіда
|
|
|
II.
Мотивування та актуалізація опорних знань учнів Діти, ми вже з вами вже
вивчили закономірності успадкування ознак, які теоретично і практично
відобразив у своїх законах Г. Мендель. Зараз я хочу, щоб ми разом навчилися
застосовувати теоретичні знання з розділу зчепленого успадкування ознак на
практиці. Ми з вами побачимо, як впливає явище кросинговеру на формування
нових фенотипів та генотипів нащадків. Ця тема була і буде дуже актуальна не
тільки для рослин і тварин, а й для людини. Адже процес кросинговеру має дуже
велике значення для комбінативної мінливості, що сприяє природному добору
(збільшення різноманіття генотипів та фенотипів). III.
Формування умінь та навичок учнів Задача № 1 У кукурудзи зморшкуватість
насіння, карликовість рослин та блискучість паростків контролюється трьома
рецесивними генами (а, b, с). В аналізуючому схрещуванні отримано таке
розщеплення: А_В_С_ - 522; А_В_сс - 133; А_bbС_ - 113; А_bbсс - 117; ааВ_С_ -
120; ааВ_сс - 112; ааbbС_ - 137; ааbbсс - 530. Поясніть розщеплення та
знайдіть генотипи цих рослин. Розв’язування Вводимо позначення алелей за
умовою: а - зморшкуватість насіння; А - норма; b - карликовість рослин; В -
норма; с - блискучість паростків; С - норма.
|
Розв’язує учень за допомогою
вчителя та класу
|
|
Згідно закономірностям незалежного
успадкування, які відображені у законах Г.Менделя, розщеплення повинно бути,
як при типовому аналізуючому схрещуванні: Р ♀ АаВbСс х ♂ ааbbсс Þ 1
: 1 : 1 : 1 : 1 : 1 (якщо успадкування кожної ознаки не залежить від інших).
Проте в наших дослідах фактичне розщеплення не збігається з теоретично
передбаченим. Тому ми можемо припустити, що тут має місце зчеплене
успадкування. При утворенні гамет у таких організмів в результаті
кросинговеру утворюються поряд з некросоверними гаметами гамети, що мають
нові комбінації генів (кросоверні). Про їх наявність ми можемо судити по
характеру розщеплення, тобто по фенотипам нащадків та по їх кількості.
Доводячи, що в нашій задачі має місце випадок зчепленого успадкування, вводимо
новий запис генотипів батьківських форм (такий, який запропонував Т. Морган).
В цьому запису горизонтальні риски символізують гомологічні хромосоми а
літери однакові - алельні гени: Р ♀ А В С х ♂ а b с а b с а b с
(рисочками показані можливі перехрести кросинговеру) Як ми з вами вже
сказали, батьківські форми утворюють кросоверні та некросоверні гамети: ♀
1) АВС некросоверні (обміну ділянками хромосом не відбулося) 2) аbс 3) Аbс
гамети кросоверні, тобто вони утворилися після 4) аВС обміну гомологічних
ділянок хромосом:
|
|
|
5) аВс 3,4 - обмін А; 5,b - обмін В; 6) АbС а
b 7) аbС 7,8 - обмін С 8) АВс с. ♂ Чоловічий організм дає тільки один
тип гамет (авс). Навіть, якщо при мейозі і відбудеться кросинговер, то його
наслідки не будуть помітні, бо нових комбінацій генів не утвориться. В
результаті схрещування батьківських форм, тобто злиття гамет, які вони
утворюють (материнського організму - 8 типів гамет, батьківського - 1 тип
гамет), ми отримуємо такі генотипи гетерозиготних рослин: АВС - 522; 5) аВс -
112; Авс авс Авс - 530; 6) АвС - 113; Авс авс Авс - 117; 7) авС - 137; Авс
авс АВС - 120; 8) АВс - 133. Авс авс. Так як за умовою задачі нормальне
листя, нормальний ріст та неблискучість паростків мають 522 екземпляри
рослин, а всі рецесивні ознаки мають 530 рослин, тобто переважна більшість
від цих рослин, наше припущення вірне. Таким чином, при утворенні жіночих
гамет в даному прикладі пройшов множинний кросинговер (між генами Аа та Вb;
Вb та Сс), внаслідок чого утворилось 6 типів кросоверних гамет, які після
запліднення утворили 6 нових фенотипів, кількість яких значно менша, ніж
некросоверних. Ці міркування відповідають даним експериментальної ситуації.
|
|
|
Задача № 2 Провели аналізуюче схрещування
рослин томату, гетерогенних за рецесивними мутаціями, які обумовлюють
карликовість рослин та опушення плоду. Нормальні алелі цих генів визначають
високий ріст та гладкість плодів. Отримані слідуючи результати: високі
гладенькі - 161; високі опушені - 5; карлікові гладенькі - 5; карликові
опушені - 118. Аналогічне аналізуюче схрещування іншої рослини томату,
гетерозиготної за тими ж алелями, показало інші результати: високі гладенькі
- 7; високі опушені - 138; карликові гладенькі - 165; карликові опушені - 4.
Чому ж ці два схрещування показали різні результати? Знайдіть частоту
рекомбінацій та генетичну відстань між двома генами. Розв’язування: Вводимо
позначення алелей за умовою: А - високий ріст; а - карликовість; В -
гладенькі плоди; b - опушені плоди. Аналіз розщеплення нащадків (перша
рослина). Нащадки від аналізуючого схрещування дигетерозиготи дали чотири
фенотипічні класи. Якщо б гени А та В успадковувались незалежно, тобто були б
розташовані в різних парах хромосом, то розщеплення було б в співвідношенні
1:1:1:1. Переважання форм з окремим фенотипом дає нам підстави припустити, що
тут йдеться про зчеплене успадкування. Нащадки рекомбнантного типу з новим,
порівняно з батьківськими формами, фенотипом (високі, опушені та карликові
гладенькі) могли утворитись тільки за рахунок кросинговеру між генами А та В.
|
Інший учень розв`язує за допомогою вчителя та
класу
|
Записуємо генотипи
батьківських форм (за Т. Морганом): Р ♀ АВ х ♂ ав ав ав ♀ 1)
АВ; 2) ав - некросоверні гамети. 3) Ав; 4) аВ - кросоверні гамети (кросинговер
одинарний, бо розглядається тільки дві ознаки, а ми в даному випадку
припускаємо, що кожна ознака контролюється одним геном, який може бути
виявлений у двох станах: домінантному та рецесивному. У результаті аналізуючого
схрещування отримали: АВ - високі гладенькі (161); ав Ав - карликові опушені
(118); ав Ав - високі опушені (5); ав Ав - карликові гладенькі (5). ав З
результатів чисельних дослідів, які протягом багатьох років на різноманітних
об’єктах проводили генетики різних країн, відомо, що кросоверних особин серед
усіх нащадків завжди значно менше, ніж некросоверних. А критерієм істини є, як
вчить філософія, практика. Тому ми вважаємо, що рослини з фенотипами високі,
опушені та карликові, гладенькі є кросоверними (їх за умовою тільки по п’ять
рослин). Далі знаходимо частоту рекомбінації (величину кросинговера): 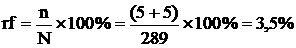
|
N = (161 + 5 + 5 + 118) = 289 - загальна
кількість особин
|
|
Висновок: гени А та В
локалізовані в одній хромосомі на відстані 3,5 % кросинговера. На підставі
попередніх міркувань про те, що кросоверних особин завжди менше, між
некросоверних, припускаємо, що у другому випадку високі рослини з гладенькими
плодами та карликові рослини з опушеними плодами, яких значно менше від інших,
утворилися від запліднення кросоверних материнських гамет. Тоді ми можемо записати
генотип та гамети материнської та батьківської рослини. 1. Тепер ми можемо
записати генотипи батьківських форм: Р ♀ 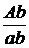 х ♂
х ♂ 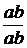
Жіноча рослина утворює 4 типа
гамет:
♀ 1) Аb; 2) аВ - некросоверні
гамети.
) АВ; 4) аb - кросоверні
гамети.
. Серед нащадків були (за
умовою):
) - високі
опушені (138);
) 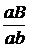 - карликові
гладенькі (165);
- карликові
гладенькі (165);
) 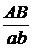 - високі
гладенькі (7);
- високі
гладенькі (7);
) - карликові
опушені (4).
Далі знаходимо частоту
рекомбінації (величину кросинговеру):
Висновок: Гени локалізовані
в одній хромосомі на відстані 3,5 % кросинговеру; На результати схрещування
впливає положення генів в гомологічних хромосомах, тобто положення, коли обидва
домінантних гена розташовані в одній хромосомі, а обидва рецесивних гена - в
другій - називається цис-положення 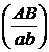 , а транс - положення - в одній
хромосомі знаходяться домінантні алелі одного гена і рецесивні іншого гена, а в
другій хромосомі (гомологічній) - навпаки
, а транс - положення - в одній
хромосомі знаходяться домінантні алелі одного гена і рецесивні іншого гена, а в
другій хромосомі (гомологічній) - навпаки 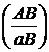 .
.
IV. Перевірка засвоєння умінь та
навичок учнів.
Варіант I
. Перший рівень
(репродуктивний)
а) Дайте визначення
кросинговеру.
Яке його біологічне значення?
б) Хто є засновником
хромосомної теорії спадковості? Назвіть її основні положення?
. Другий рівень:
Задача. Жіноча рослина
кукурудзи має рецесивні ознаки забарвлення, зморшкуватості та крохмалистості
насіння. Чоловіча ж рослина гетерозиготна по кожній з цих ознак, так що насіння
у неї безкольорові, гладенькі та воскові. Нащадки від цього схрещування були
отримані таким чином:
Забарвлені зморшкуваті
крохмалисті - 40
Забарвлені зморшкуваті
воскові - 9
Забарвлені гладенькі
крохмалисті - немає
Забарвлені гладенькі воскові
-1
Безкольорові зморшкуваті
крохмалисті - 1
Безкольорові зморшкуваті
воскові - немає
Безкольорові гладенькі
крохмалисті - 9
Безкольорові гладенькі
воскові - 40.
В одній чи декількох
хромосомах знаходяться гени вказаних ознак і чи можна спостерігати у даному
випадку явище перехресту?
. Третій рівень
Задача. Що можна сказати на
прикладі даних попередньої задачі про порядок розташування в хромосомі
фігуруючих там генів та про відстань між ними?
При яких умовах можливе
утворення нащадків із забарвленим гладеньким та крохмалистим насінням?
Варіант II
. Перший рівень
а) Як ви розумієте поняття
“зчеплене успадкування”? Що таке “зчеплені гени”, “група зчеплення”?
б) Що ви можете розповісти
про генетичні карти хромосом?
. Другий рівень
Задача. Жіноча особина мушки
дрозофіли має рецесивні ознаки сірого тіла, червоного кольору очей та
нормальної форми крил. Чоловіча особина гетерозиготна по кожній з цих ознак,
тобто має чорне тіло, білі очі та вкорочені крила. Нащадки були отримані таким
чином:
Сірі, червоні очі, нормальні
крила - 80
Сірі, білі очі, нормальні
крила - 19
Сірі, білі очі, нормальні
крила - немає
Сірі, білі очі, вкорочені
крила - 2
Чорні, червоні очі, нормальні
крила - 2
Різнорівнева самостійна
робота по варіантах (варіанти записані на дошці)
|
|
|
Чорні, червоні очі, вкорочені крила - немає
Чорні, білі очі, нормальні крила - 18 Чорні, білі очі, вкорочені крила - 80 В
одній чи декількох хромосомах знаходяться гени вказаних ознак і чи можна
сказати у даному випадку що відбувається перехрест? 3. Третій рівень Задача.
Що можна сказати на прикладі даних попередньої задачі про порядок
розташування в хромосомі фігуруючих там генів та про відстань між ними? При
яких умовах можливе утворення особин, що мають чорне тіло, червоні очі та
вкорочені крила? V. Домашнє завдання Задача. Самку дрозофіли, гетерозиготну
по рецесивним генам чорного кольору тіла маленьких крил, схрестили із самцем,
який має чорне тіло та маленькі крила. Від цього схрещування отримали: 249
мух, що мали чорне тіло та нормальні крила 20 мух, що мали сіре тіло та
нормальні крила 15 мух, які мали чорне тіло та маленькі крила 216 мух, які
мали сіре тіло та маленькі крила. Чи вважаєте ви, виходячи з цих даних, що ці
дві пари генів зчеплені або ні? Якщо ви вирішили, що вони зчеплені, то
вкажіть, яке з двох тверджень, що приведені нижче, правильно описує їх
зчеплення у самки: а) гени чорного тіла та маленьких крил знаходяться в одній
хромосомі, а гени сірого тіла та нормальних крил - в її гомолозі. Під час
мейозу відбулися кросинговери
|
|
|
б) гени чорного тіла та нормальних крил лежать
в одній хромосомі, а гени сірого тіла та маленьких крил - в її гомолозі. Під
час мейозу відбулися кросинговери. Напишіть генотипи усіх особин. Задача № 2
У дрозофіли ген червоних очей домінує над геном пурпурових очей, а ген довгих
крил домінує над геном вкорочених крил. Самку, гетерозиготну за обома
ознаками, схрестили з самцем, який має пурпурові очі та вкорочені крила. В F1
отримали: 109 особин з червоними очима та довгими крилами 114 особин з
червоними очима та вкороченими крилами 122 особини з пурпуровими очима та
довгими крилами 116 особин з пурпуровими очима та вкороченими крилами Чи
вважаєте ви, виходячи з цих даних, що ці дві пари генів зчеплені або ні? Якщо
ви вирішили, що вони зчеплені, то вкажіть, яке з двох тверджень, що приведені
нижче, правильно описує їх зчеплення у самки: а) гени червоних очей та довгих
крил знаходяться в першій хромосомі, а гени пурпурових очей та вкорочених
крил - в її гомолозі. Під час мейозу відбулися кросинговери б) гени червоних
очей та вкорочених крил знаходяться в першій хромосомі, а гени пурпурових
очей та довгих крил - в її гомолозі. Під час мейозу відбулися кросинговери.
Напишіть генотипи усіх особин.
|
|
ВИСНОВКИ
Керуючись знаннями про механізми
комбінативної мінливості можна шляхом гібридологічного методу з наступним спрямованим
добором створювати бажані комбінації генів: з’єднувати або роз’єднувати їх,
тобто отримувати подвійні або одинарні мутантні форми. Такі маніпуляції
зроблені нами з генами white та cut у Drosophila melanogaster.
Внаслідок наших практичних експериментально
отримані дві нові форми - одиночні мутанти white та cut Drosophila
melanogaster.
Отримати такі константні форми можна
через чотири покоління, причому треба враховувати особливості успадкування
ознак, зчеплених зі статтю.
Відсутність розщеплення при
аналізующому схрещуванні свідчить про зчеплення генів white та cut. Наявність
кросоверних особин дає змогу підрахувати відстань між генами, яка визначається
величиною кросинговеру. приділяється
Встановлено відстань між генами
white та cut, яка складає 18,8 сантіморган (що співпадає з літературними
даними).
Проаналізувавши шкільну програму з
біології нами з’ясовано, що час, який приділяється на генетику, зокрема на тему
“Зчеплене успадкування. Кросинговер. Хромосомна теорія спадковості”,
недостатній для розуміння такої складної теми. Тому якість розуміння та
засвоєння матеріалу, що викладається, у значній мірі залежить від
професіоналізму та майстерності вчителя. Якість засвоєння матеріалу залежить
також від наявності наочного матеріалу та практичних навичок, які набувають
учні під час уроків біології.
Наші уроки, присвячені темі
зчепленого успадкування, особливо урок “Вирішення задач з генетики”, спрямовані
на поліпшення навчального процесу. Вони складені з урахуванням нової системи
оцінки знань за 12 - бальною системою: питання та задачі складені різнорівневої
складності від більш простих до складних. Це дає змогу виявити реальні знання
школярів та рівень їх розуміння проблеми. Це дає підстави для обґрунтованого
об’єктивного індивідуального підходу та ставлення до кожного учня, а без цього
неможлива сучасна педагогіка.
Для ілюстрації явища зчепленого
успадкування у живих істот, нами спланований та проведений лабораторний
експеримент на дрозофілах. Для шкільних умов цей дослід може бути використаний
тільки при проведенні факультативних занять, або у позакласний час.
Реальні дані, отримані у досліді,
лягли в основу власноруч сформульованої, розв’язаної та поясненої задачі. Ця
задача ілюструє можливість оперування зчепленими генами, можливість розривати
зчеплення завдяки кросинговеру. Така задача може бути запропонована як задача
творчого рівня.
ЛІТЕРАТУРА
Айала
Ф.А., Кайгер Дж. “Современная генетика” в 3х томах, М.: Мир, 1987, 240 с.
Барна
В.І., Барна М.М. “Збірник задач і розв’язків з біології”, Мандрівець, 1996, с.
32 - 74.
Ватти
К.В., Тихомирова М.М. “Руководство к практическим занятиям по генетике”, М.:
Просвещение, 1972, 123 с.
Гершензон
С.М. “Основы современной генетики”, К.: Наукова думка, 1979, 247 с.
Гуляев
Т.В., Мальченко В.В. “Словарь терминов по генетике, цитологии, селекции,
семеноведению”, М.: Мир, 1975, 135 с.
Дубинин
Н.П. “Общая генетика”, М.: Наука, 1976, 347 с.
Зайцев
Г.Н. “Методика биометрических рассчетов”, М.: Наука, 1973, 254 с.
Захаров
И.А. “Генетические карты высших организмов”, Л.: Наука, 1979, 156
Инге-Вечтомов
С.Т. “Генетика с основами селекции”, М.: Высшая школа, 1989, 264 с.
Кемп
П., Армс К. “Введение в биологию”, М.: Мир, 1988, с. 415 - 423.
Кучеренко
М.Є., Вервес Ю.Г., Болан П.Г. та ін. “Загальна біологія” Підручник для учнів 10
- 11 -х класів серед. загальноосвіт. шк. К.: Генеза, 1998, 145 с.
Лобашев
М.Е. “Генетика с основами селекции”, М.: Просвещение, 1970, 342
Лобашев
М.Е. “Генетика”, Лен. Университет, 1967, 127 с.
Медведев
Н.Н. “Практическая генетика”, М.: Наука, 1966, 237 с.
Програма
з біології для 6 - 11 класів середньої загальноосвітньої школи (2001).
Програма
для профільних класів, ліцеїв, гімназій з поглибленим вивченням біології та
хімії (6 - 11 класи), 1996.
Реф
Р., Кармен Т. “Эмбрионы. Гены. Эволюция”, М.: Мир, 1989, 345 с.
Соколовская
Б.Х. “120 задач по генетике (с решениями), М.: Наука, 1992, С. 15 - 17.
Стрельчук
С.І., Демідов С.В. та ін. “Генетика з основами селекції”, Киіїв,
Фітосоціацентр, 2000, С. 80 - 90.
Тимчук
Н.Ф., Тимченко С.Л. “Хімія клітини та спадковість”, Методичний посібник,
Балаклія, 1995, 35 с.
Ткачук
З. Ю., Морозов М.М., Пилипчук О.Я. “Основи загальної генетики”, К.: Вища школа,
1995, С. 89 - 94.
Ткачев
А.Т., Надточий А.С. “Решение генетических задач”, Харьков. Методические
рекомендации, 1988, 45 с.
Уотсон
Дж. “Молекулярная биология гена”, М.: Мир, 1978, 346 с.
Шмальгаузен
И.И. “Проблемы дарвинизма”, Л.: Наука, 1969, С. 156 - 161.