Разведение стерляди
ПЛАН
.
БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СТЕРЛЯДИ
.1
Систематика
.2
Морфология
.3
Питание
.4
Распространение
.5
Размножение
.6
Жизненный цикл стерляди
.6.1
Эмбриональный период развития
.6.2
Критические стадии в эмбриогенезе
.6.3
Нарушения в эмбриогенезе
.6.4
Особенности предличиночного периода
.6.5
Личиночный период
.6.6
Мальковый период
.
ВЫБОР МЕСТА ДЛЯ ЛОСОСЕВОГО РЫБОВОДНОГО ЗАВОДА
.
ГИДРОЛОГИЧЕСКАЯ И ГИДРОХИМИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕКИ АМУР
.1
Гидрологические условия
.2
Гидрохимические особенности
.
ОПИСАНИЕ ТЕХНОЛОГИЧЕСКОГО ПРОЦЕССА ЛРЗ НА РЕКЕ АМУР
.1
Отлов производителей стерляди
.2
Выдерживание производителей стерляди
.3
Взятие зрелых половых продуктов у производителей, осеменение икры и
подготовка
её к инкубации
.4
Инкубация икры
.5
Выдерживание предличинок и подращивание личинок
.6
Выращивание молоди
.7
Уход за предличинками, личинками и молодью стерляди
.8
Выпуск молоди, мероприятия, обеспечивающие наибольшее её выживание
.
СОСТАВ ЛОСОСЕВОГО РЫБОВОДНОГО ЗАВОДА
ЛИТЕРАТУРА
разведение стерлядь икра
1.
БИОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА СТЕРЛЯДИ
.1 Систематика
Тип Chordata - хордовые
Подтип Vertebrata - позвоночные
Надкласс Gnatostomata - челюстноротые
Класс Osteichtyes - костные рыбы
Подкласс Actinopterygii - лучеперые рыбы
Надотряд Ganoidomorpha - ганоидные
Отряд Acipenseriformes - осетрообразные
Семейство Acipenseridae - осетровые
Вид Acipenser ruthenus (Lovetsky, 1828) -
стерлядь (рис. 1).


Рисунок 1 - Стерлядь [1]
.2 Морфология
Стерлядь по внешнему виду очень похожа на
осетра, но рыло более заостренное и слегка загнуто вверх. Жаберные перепонки
прикреплены к межжаберному промежутку. Рот небольшой в виде поперечной щели,
расположен на нижней части головы. Нижняя губа посередине прервана. Впереди рта
четыре усика с бахромой. Тело покрыто пятью рядами костных жучек, а между
рядами жучек покрыто очень мелкими гребенчатыми зернышками. Костные жучки
маленькие, на боках тела их 60-70. Боковой ряд жучек светлый; у молоди он имеет
вид белой полосы вдоль тела. Спинных жучек 12-17, боковых 57-71, брюшных 10-19.
Жаберных тычинок 15-26 (31). Д 37-54; А 19-31. Спинная сторона тела
тёмно-зелёная или серая, брюхо желтоватое.[2]
.3 Питание
Первое время после выхода из икры мальки
держатся в слабо заиленных воложках, расположенных ниже нерестилищ, где они
питаются мелкими олигохетами, личинками хирономид и личинками мошки. По мере
роста стерлядки начинают питаться более разнообразными организмами, и к
середине лета их пища мало отличается от пищи крупных особей. В это время
сеголетки волжской стерляди перекочевывают на пески у левого берега Волги и
питаются здесь главным образом личинками хирономид. Много мелкой стерляди
держится также на камнях у правого берега Волги. Состав пищи стерляди в очень сильной
степени зависит от тех участков, на которых она держится.
Весной, в периоды массового развития воздушных
насекомых, стерлядь питается почти исключительно личинками последних и выходит
для нагула на полои. Если во время разлива насекомых мало, основными местами
нагула служат каменистые и галечные участки русла. Стерлядь питается там
преимущественно Corophium и личинками Hydropsyche. В это время она заходит
также в затоны, где поедает в больших количествах личинок хирономид. В период
паводка обычно в составе пищи стерляди на каждом участке преобладает
какой-нибудь один организм.
Летом основными местами нагула служат каменистые
участки у правого берега Волги. Здесь состав организмов, поедаемых стерлядью,
значительно разнообразнее, чем весной; резкого преобладания одной какой-либо
формы не наблюдается. Обычны для этих участков Corophium, Hirudinea, личинки
Hydropsyche. В меньших количествах встречаются бокоплавы, личинки хирономид и
некоторые другие формы. На пески крупная стерлядь заходит редко. На этих участках
желудки стерляди обычно наполнены большим количеством личинок хирономид, а
иногда и мелкими моллюсками.[3]
.4 Распространение
Из всех осетровых стерлядь имеет самое широкое
распространение: она водится в бассейнах Черного и Каспийского морей, а также в
бассейнах рек Северного Ледовитого океана от Северной Двины на западе до Енисея
на востоке.
Стерлядь рыба речная и, как правило, в море не
встречается, однако попадались отдельные экземпляры в чисто морских водах
Черного, Азовского и Каспийского морей.
В бассейне Дуная стерлядь водится повсюду, но в верховьях редка. В среднем
течении и в низовьях многочисленна. Из Дуная стерлядь поднимается в Вааг,
Драву, Саву и её притоки, Уну, Кульпу, а также в Тису, Альт и Прут. В Днестре
стерлядь встречается главным образом на среднем течении и редко на участке в
предгорий Карпат, обитает в верхней части Дубоссарского водохранилища, в
Днестровском лимане очень редка.
Из Днепра заходит в Припять, Тетерев, Десну до
Брянска, в Псёл. Обитает преимущественно на нижних участках реки (Кременчуг -
Херсон).
В Дону водится на нижнем и среднем течении.
Обычна в Цимлянском водохранилище, особенно в верхних участках. Из притоков
Дона заходит в Донец, Оскол, Медведицу. В Волге стерлядь встречается по всей
реке от низовьев (дельты) до Твери. Особенно многочисленна она в среднем и
нижнем течениях, обитает во всех волжских водохранилищах (очень редка в
Рыбинском, Иваньковском, Угличском).
Широко распространена в притоках Волги. Редко
встречается после постройки Рыбинского водохранилища в Шексне и Мологе. В Оке
встречается до г. Орла, наиболее многочисленна стерлядь в нижнем течении.
Заходит в притоки Оки - Жиздру, Упу, Угру, Москву, Клязьму. Встречается в
Ветлуге, Суре до Пензы, в Свияге. Широко распространена в Каме и ее притоках;
Белой, Вятке до верховьев, Чусовой, Вишере, Колве, Самаре. Обитает в камских
водохранилищах. Единичные экземпляры попадаются в бассейнах Ладожского и
Онежского озер, в реках - Нева, Волхов, Свирь.
В бассейн Северной Двины стерлядь проникла в 19
веке через Северо-Екатерининский канал из Камы. В самой Северной Двине стерлядь
наиболее многочисленна в среднем и нижнем течении. В притоках Северной Двины
стерлядь широко распространена в Вычегде (от верховьев до устья), в Пинеге и
Ваге (в среднем и нижнем течении), встречается также в низовьях Лузы, Юга,
Емцы, Сысоле, Выми.
В Печоре стерлядь встречается преимущественно в
среднем течении.
Ареал распространения стерляди в Сибири
охватывает большую часть Западной Сибири (Обь-Иртышский бассейн) и почти всю
западную часть Средней Сибири (бассейн Енисея). Б. Г. Иоганзен (1946) в
Обь-Иртышском бассейне выделяет три самостоятельных стада стерляди,
различающиеся между собой по некоторым систематическим признакам, также
биологическими особенностями и определенным местам локализации этих стад в
определенных границах речных бассейнов Оби, Иртыша и их притоков. Так,
ежегодные заморы на нижней и средней Оби отделили иртышскую стерлядь от средне-
и верхнеобской. В Иртыше стерлядь распространена от его устья до Черного Иртыша
включительно. Заходит в его притоки: Ишим, Тобол, Туру, Тавду, Демьянку,
держится там преимущественно в нижнем течении. Промысловое значение на Иртыше
стерлядь имеет в Вагайском, Уватском Тарском районах, в нижнем течении реки. В
пределах верхнего течения Оби стерлядь распространена на всем участке;
встречается в нижнем течении Бии, Катуни, Чарьппа, Алея, обычна в Новосибирском
водохранилище, северная граница распространения этого стада проходит где-то в
районе устья Томи, где происходит смешивание верхнеобской стерляди с приходящей
из заморной зоны среднеобской.
Основным местом распространения стерляди на
Средней Оби является участок, заключенный между устьями Томи и Иртыша. Из Оби
стерлядь заходит в низовье Томи, обычна в нижнем и среднем течении Чулыма, в
небольшом количестве встречается в
низовьях Шегарки, Чаи, Кети, Парабели, Васюгана, Тыма, Ваха. В промысловом
количестве водится в пределах всего среднего течения Оби, но преобладает в его
незаморной зоне. В нижнем течении Оби, по мере продвижения к северу, количество
стерляди постепенно сходит на нет. Малочисленность стерляди на Нижней Оби можно
объяснить ежегодной заморностью реки, отсутствием зимовалов и нерестилищ.
Однако существует локальное стадо стерляди в р. Ляпин - в одном из верхних
притоков р. Северная Сосьва. Заходы стерляди в р. Ляпин можно объяснить,
вероятно сильным загрязнением Нижнего Иртыша в последние годы и миграцией
стерляди из Иртыша вниз по Оби, а также благоприятными условиями в реке Ляпин
для ее размножения и зимовки.
Стерлядь в бассейне р. Енисея известна, помимо
самого Енисея, в правых притоках: р. Ангаре и ее притоках (Чуне, Тасеевой,
Оке), Подкаменной Тунгуске, Нижней Тунгуске, Сисиме, Тубе, а также в левых
притоках - Абакане и р. Сыме, в последней на протяжении 250 - 300 км от устья.
По Енисею стерлядь выше Большого порога не встречается. Ниже Большого порога до
Минусинска численность ее не велика. От Минусинска и до дельты Енисея стерлядь
является промысловой рыбой, наибольшие концентрации ее находятся между Ангарой
и Курейкой. В дельте Енисея она встречается сравнительно редко. В протоках
Бреховского архипелага и в губе держится единично Восточнее бассейна Ангары,
стерлядь не встречается.[4]
Выделяют две формы стерляди: острорылую и
тупорылую. Острорылая (медленнорастущая, яровая раса) форма стерляди
встречается преимущественно в реках: Дунае Днестре, Днепре, Волге, Каме, Оке,
Иртыше, Оби, причем в Верхней и Средней Оби стерлядь представлена исключительно
острорылой формой, а амплитуда индекса длины рыла весьма незначительна (44 -
52% длины головы). Яровая раса стерляди более длиннорылая (длина рыла > 40%
длины головы), быстро растет только в первый год жизни (затем темп роста
снижается), менее упитанная и плодовитая, но созревающая раньше и нерестующая
чаще озимой стерляди. Острорылая стерлядь старше 14 лет обычно не встречается и
особенно крупных размеров не достигает. Всюду является более многочисленной и
составляет основу промысла. Тупорылая (быстрорастущая, озимая раса) форма
встречается в Дунае (очень редко), Волге, Каме, Оке, Суре, Северной Двине,
Вычегде, Иртыше, Енисее. Высокий процент тупорылых форм отмечен для Северной
Двины и еще более высокий для Вычегды от 43 до 60%. Озимая раса отличается от
яровой меньшей длиной рыла (длина рыла <38% длины головы), более упитанная и
плодовитая, нереститься позже яровой расы. [4]
Стерлядь совершает лишь небольшие передвижения,
связанные или с сезонными изменениями гидрологического режима реки, или же с
поисками пищи, нерестом и залеганием на зимовку. Во время заморов наблюдается
заход стерляди в глубокие затоны или подход ее к устьям тех притоков, вода
которых богата кислородом.[4]
.5 Размножение
Нерест происходит в период от второй половины
апреля до первой половины июня. На Волге и Каме время нереста (май - начало
июня) обычно совпадает с пиком паводка; начало нереста зависит, прежде всего от
прогревания воды до 7-10,5°С. Плодовитость составляет от 5 до 140 тыс. икринок.
Нерест неежегодный: самцы нерестятся через 2 года, самки - через 4-5 лет.
Минимальная длина зрелых самцов 28 см, самок - 34 см. Допустимое соотношение
самок и самцов при искусственном воспроизводстве в маточном стаде 5:1.
Чем выше расположены нерестилища, тем раньше
начинается икрометание. При очень быстром потеплении воды икра не успевает
созреть к тому моменту, когда среда становится благоприятной для нереста, и
размножение начинается при более высоких температурах, чем обычно (нерест может
происходить при температуре до 20°). Икра откладывается на галечных или
каменистых участках, на субстрат, попадая в расщелины между камнями.
Нерестилища располагаются у верховой части
островов или у вдающихся в реку мысов, на местах с очень быстрым течением, не
ниже 1,5-2 м/сек., где отсутствует заиление и слабо развита фауна
беспозвоночных.
У половозрелых стерлядей весной появляется
брачный наряд, в виде беловатого налета на голове.
Икринки стерлядей клейкие; диаметр их 1,9-2 мм.
Развитие икринок длится, в зависимости от температуры воды, в естественных
условиях от 6 до 11 суток. Только что выклюнувшиеся предличинки имеют длину 6-7
мм. Желток у личинки рассасывается через шесть-десять суток после выклева.
Месячные мальки достигают длины 3-4 см, а сеголетки в августе - сентябре имеют
длину от 8 до 18 и даже до 25 см.[5]
.6.1 Эмбриональный период развития
В эмбриональный период онтогенеза эмбрион
питается материнскими запасами пищи, заключенными в желтке. Такое эндогенное
питание свойственно не только эмбрионам, находящимися под оболочкой икринок, но
сохраняется определенное время и после вылупления у предличинок.
этап. Оплодотворение. Следствием осеменения
является набухание оболочек, совпадающее со значительным увеличением прочности.
Через несколько минут наблюдается поворот яйца: вегетативное полушарие
переходит вниз, а анимальное - вверх (рис.2).
Затем между анимальной областью и оболочками
создается перивителиновое пространство.
А - положение яйца до осеменения и Б -
Положение яйца после
в
первые минуты оплодотворения поворота анимальной областью вверх
Рисунок 2 - 1 стадия [6]
Рисунок анимальной области яйца (светлое
полярное пятно в центре и темные пигментные кольца вокруг него) изменяется
(рис.3): пигмент постепенно стягивается к центру анимальной области, образуя
здесь темное скопление, светлое полярное пятно исчезает или сильно уменьшается
в размере.


м.к. - микропилярные канальца; пв. п -
перивителлиновое пространство; с. о. - студенистая оболочка; ж.о.1 - наружная
желточная оболочка; ж о. 11 - внутренняя желточная оболочка
Рисунок 3 - 2 стадия [6]
Некоторое время спустя после оплодотворения на
краю анимальной области появляется очень светлая белая полоска протяженностью
около половины окружности. Полоска эта постепенно расширяется и превращается в
светлый серп. Сторона яйца со светлым серпом становится затем спинной стороной
зародыша (рис.4).[6]

с - серп
Рисунок 4 - 3 ст. Стадия светлого серпа [6]
этап. Дробление. В отличие от костистых рыб, у
которых дробится только бластодиск, у осетровых делится все яйцо. Наступление
первого деления обнаруживается по появлению в области анимального полюса
небольшой светлой полоски, посередине которой проходит тонкая темная линия -
первая борозда дробления (рис.5). Яйцо делится вначале на 2 части (два бластомера),
потом каждая из них снова делится надвое (4 бластомера) и так далее. Деление
происходит быстрее на анимальном полюсе.
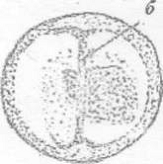
б - борозда
Рисунок 5 - 4 ст. Стадия первого деления [6]
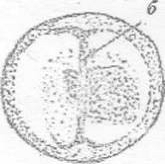
б - борозда
Рисунок 5 - 4 ст. Стадия первого деления [6]
Анимальная область разделена первой бороздой.
б1

Вид сверху Вид сбоку
б1 - борозда 1-го деления; б2 - борозда 2-го
деления
Рисунок 6 - 5 ст. Стадия второго деления [6]
Анимальная область разделена на четыре
бластомера (рис.6).

б3 - борозда третьего деления
Рисунок 7 - 6 ст. Стадия третьего деления [6]
Анимальная область разделена на 8 бластомеров
(рис.7).
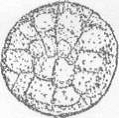
Рисунок 8 - 7 ст. Стадия четвёртого деления [6]
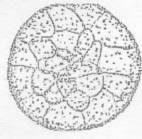
Рисунок 9 - 8 ст. Стадия пятого деления [6]
ан.б.

Вид сверху Вид сбоку
ан. б. мелкие бластомеры анимальной области;
вег. б - крупные бластомеры вегетативной области
Рисунок 10 - 9 ст. Стадия седьмого деления [6]
Борозды полностью разделяют вегетативную область
(рис.10).

Рисунок 11 - 10 ст. Стадия позднего дробления
[6]
Дробление переходное от полного радиального
дробления к частичному дискоидальному. Первые три плоскости дробления -
меридиальные. Деления в вегетативной части идут, но сильно запаздывают, и это
ведет к резкому отличию анимального полушария, состоящего из мелких клеток, от
вегетативного -крупноклеточного (рис.8-11). Это следствие неравномерного
распределения в яйце запасных веществ.
Постепенно в центре эмбриона между бластомерами
накапливается жидкость, раздвигающая их, возникает полость и эмбрион
приобретает строение полого шарика -бластулы (рис.12).
Постепенно полость бластулы (полость дробления)
увеличивается, а крыша ее утончается. На стадии ранней бластулы клетки
анимальной области еще довольно велики и их можно различить при небольшом
увеличении. Несколько позже, на стадии поздней бластулы, они настолько мелки,
что увидеть их можно только под микроскопом. Между мелкими анимальными и
относительно крупными вегетативными бластомерами имеется зона бластомеров
промежуточного размера, носящая название краевой зоны. Именно с нее начинаются
изменения, приводящие к переходу эмбриона на новый этап развития.
вег.б.
ан.б.

ан.б. - мелкие бластомеры анимальной области;
вег.б. - крупные бластомеры вегетативной области
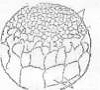
Рисунок 12 - 11 ст. Стадия ранней бластулы [6]
ан. - мелкоклеточная анимальная область; вег. -
вегетативная область; кр.- краевая зона
Рисунок 13 - 12 ст. Стадия поздней бластулы [6]
Стадия поздней бластулы (рис.13) характеризуется
тем, что в анимальной области отдельные клетки при небольшом увеличении
неразличимы; перестройка клеточного цикла в бластомерах анимальной области.[6]
этап. Гаструляция. Материал краевой зоны
вворачивается внутрь, а светлая анимальная область разрастается и покрывает
снаружи, темные вегетативные клетки. Эти процессы называются гаструляцией, а
эмбрион в описываемый период - гаструлой. Гаструляция у осетровых того же типа,
что и у костистых, но начинается несколько позже, при обрастании (эпиболии)
более ½
яйца.
Изменения начинаются с того, что на будущей
спинной стороне эмбриона, приблизительно на уровне экватора, появляется более
темная полоска (рис.14).

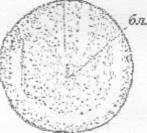
бл. - бластопор
Рисунок 14 - 13 ст. Стадия начала гаструляции
[6]
На месте этой полоски клетки уходят внутрь и
образуется узкая щель, носящая название первичного рта и бластопора. Сначала
щель бластопора короткая.

Рисунок 15 - 14 ст. Стадия ранней гаструлы [6]
Спинная губа бластопора в виде щели на небольшом
протяжении. Мало-помалу она распространяется в стороны (рис.15) и охватывает
все большую часть окружности зародыша, пока, наконец, не замкнется в кольцо,
окружая темные клетки нижней части
эмбриона (рис.16).

Рисунок 17 - 16 ст. Стадия большой желточной
пробки [6]
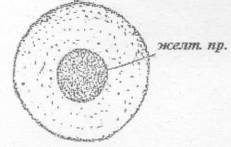
Рисунок18 - 17 ст. Стадия маленькой желточной
пробки [6]
Когда желточная пробка замыкается, происходит
закрытие бластопора (рис.19). Этот момент можно считать концом гаструляции. С
этого момента начинается основное формирование эмбриона.[6]
бл. - бластопор
Рисунок 19 - 18 ст. Стадия щелевидного
бластопора
этап. Развитие эмбриона от конца гаструляции до
начала пульсации сердца. После окончания гаструляции у эмбриона постепенно
формируются зачатки основных систем органов: нервной, пищеварительной,
кровеносной, а также мускулатуры и хорды. Эмбрион на этом этапе развития
по-прежнему неподвижен, питается за счет желтка и дышит всей поверхностью тела.
Образование нервной трубки и зачатков выделительной
системы.
Вскоре после закрытия бластопора на спинной
стороне эмбриона, в светлом слое гаструлы образуется утолщенная пластинка
(рис.20).

Рисунок 20 - 19 ст. Стадия ранней нейрулы
Начинают обозначаться нервные валики вокруг
головного отдела нервной пластинки.
н.в. - нервный валик; н.п. - нервная пластинка;
жел - желобок
Рисунок 21 - 20 ст. Стадия широкой нервной
пластинки
В ее центре проходит продольный желобок, а по
краям приподнимаются в виде подковы невысокие нервные валики. Из нервной
пластинки возникает затем нервная система (рис.21).
Более широкая передняя часть пластинки
представляет собой зачаток головного мозга, а остальная более узкая часть -
будущий спинной мозг. Постепенно срединная часть нервной пластинки углубляется,
а края, окаймленные валиками, сближаются (рис.22).
Наконец, правый и левый валики смыкаются, и из
нервной пластинки образуется нервная трубка. Преобразование эмбриона в этот
период развития называется нейруляцией, а сам эмбрион - нейрулой (рис.23).

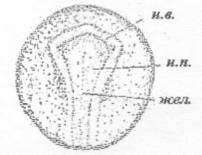
выд. - зачаток выделительной системы
Рисунок 22 - 21 ст. Стадия начала сближения
нервных валиков
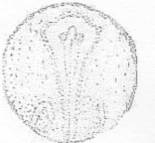
Рисунок 23 - 22 ст. Стадия поздней нейрулы
Нервные валики в туловищном отделе сближены,
зачатки выделительной системы удлинились. Когда края нервной пластинки начинают
сближаться, по бокам от нее становятся различимы неширокие светлые полоски,
которые постепенно удлиняются. Это зачатки выделительной системы.


Вид с головы Вид со спины
гол.м. - зачаток головного мозга; ш. - шов в
месте смыкания нервных валиков; выд. - зачаток выделительной системы
Рисунок 24 - 23 ст. Стадия замкнутой нервной
трубки

гл. - зачаток глаза; г.м. - передний мозговой
пузырь; з.м. - задний мозговой пузырь; с.м. - передний мозговой пузырь; ж.в. -
зачаток железы вылупления
Рисунок 25 - 24 ст. Стадия появления глазных
выростов и утолщения переднего конца зачатка выделительной системы
Изменения эмбриона после образования нервной
трубки
На стадии замкнувшейся нервной трубки (рис.24)
эмбрион имеет еще шарообразную форму, и лишь на последующих стадиях возникает
зачаток хвостового отдела, а затем, несколько позднее, начинается и обособление
головы. Постепенно задний конец тела увеличивается и образует выступ,
представляющий собой первый зачаток хвоста (рис.26).
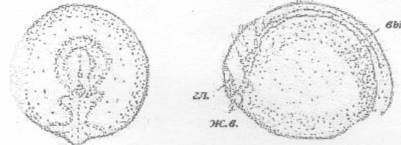
выд. - зачаток выделительной системы; гл. -
зачаток глаза; ж.в. - железа вылупления
Рисунок 26 - 25 ст. Стадия сближения боковых
пластинок и образования утолщения в области зачатка хвоста
Этот выступ постепенно удлиняется. В начале он
имеет палочковидную форму, а потом начинает уплощаться с боков. На верхней и
нижней его сторонах образуется кожная складочка - зачаток плавниковой оторочки.
Хвостовой зачаток на этих стадиях загнут вниз. Голова начинает обособляться
только к концу периода.
Начинается дифференциация головного мозга
(рис.25). В головном отделе нервной трубки вырастают три мозговых пузыря -
передний, средний и задний. В переднем мозговом пузыре в свою очередь
образуется два боковых выступа - зачатки глаз. В заднем мозговом пузыре
образуются карманообразные выросты покровного эпителия - зачатки слуховых
пузырьков. По бокам от переднего мозгового пузыря в эпителии образуются два
углубления полулунной формы - зачаток железы вылупления. Слева и справа по
бокам головного мозга образуются светлые дуги, которые загибаются вперед. Две
парные дуги -зачатки скелета челюстного аппарата. Задние дуги - зачатки
хрящевых жаберных дуг (рис.27).

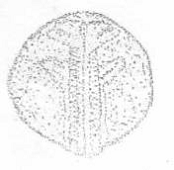
гип. - углубление в месте образования гипофиза;
гл. - зачаток глаза; Д1 - первая пара висцеральных дуг; Д2 - зачатки 2 пары
висцеральных дуг; Д3 - 3 пара висцеральных дуг
Рисунок 27 - 26 ст. Стадия слияния боковых
пластинок и начала обособления хвостового отдела эмбриона
После образования нервной трубки продолжается
расчленение среднего эмбрионального листка. Число сомитов (сегментов)
непрерывно увеличивается, и сегментация распространяется от переднетуловищного
отдела назад к хвосту. В месте срастания боковых пластинок образуется
коротенькая трубочка - зачаток сердца (рис.28).

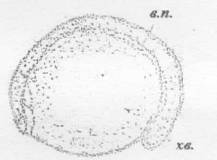
в.п. - выводной почечный проток; хв. - хвостовой
зачаток; с. - сердце
Рисунок 28 - 27 ст. Стадия короткой сердечной
трубки
Затем сердечная трубка удлиняется (рис.29) и
образует небольшой изгиб (рис.30).
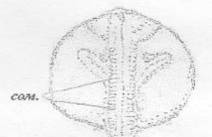

хв. - хвостовой зачаток; с. - сердце; сом. -
сомиты
Рисунок 29 - 28 ст. Стадия прямой удлинённой
сердечной трубки
С пульсации сердца начинается и
кровообращение.[6]
этап. Развитие эмбриона от начала пульсации
сердца до вылупления. В период от начала кровообращения и до вылупления
значительно изменяется внешняя форма эмбриона, голова обособляется и немного
увеличивается, хвост сильно вырастает в длину, распрямляется. Только брюшной
отдел (желточный мешок) изменяется мало; благодаря большому количеству желтка в
клетках стенки кишки он сохраняет крупные размеры. Очень заметны изменения в
хвостовом отделе. По мере удлинения хвост, будучи прижат оболочками к
желточному мешку, загибается на брюшную сторону, при этом конец его постепенно
достигает сердца, головы и заходит за голову (рис.31-32).
Продолжается развитие нервной, мышечной и других
систем эмбриона. Параллельно возрастает его подвижность. Двигаясь внутри
оболочек, эмбрион перемещает находящуюся там жидкость, что улучшает условия
дыхания. Движение эмбриона в период выклева содействует его выходу из оболочек
(рис.33).
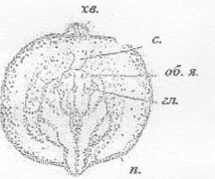
об.я. - обонятельная ямка; гл. - глаз; п. -
петля, образуемая собирающим и выводным протоками; хв. - хвост; с. - сердце
Рисунок 30 - 29 ст. Стадия образования изгиба
сердечной трубки
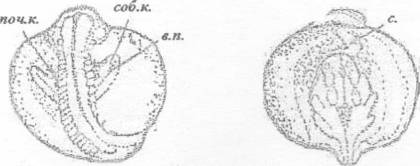
с. - сердце; в.п. - выводной почечный проток;
поч.к. - почечные канальца; соб.к. - собирающие почечные канальца
Рисунок 31 - 30 ст. Стадия на которой конец
хвоста приближается к сердцу
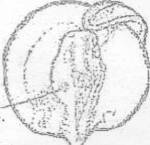
Рисунок 32 - 32 ст. Стадия на которой конец
хвоста касается головы

Рисунок 33 - 35 ст. Стадия начала выклева
единичных предличинок.
После удаления оболочек эмбрион способен к
быстрому поступательному движению.[6]
.6.2 Критические стадии в эмбриогенезе
Организм на разных стадиях развития неодинаково
реагирует на одни и те же воздействия внешней среды. Стадии и этапы повышенной
чувствительности к различным внешним воздействиям называют критическими. У
весенне-нерестующих рыб непосредственно после оплодотворения следует период
высокой чувствительности, со стадии 16-32 бластомеров чувствительность
несколько падает и вновь повышается перед началом гаструляции (12-я стадия).
Гаструляция проходит при пониженной чувствительности, которая с началом
формирования эмбриона резко повышается, это совпадает с закрытием бластопора
(18-я стадия). На этой стадии чувствительность эмбрионов в начале образования
хвостового отдела (25-я стадия) и перед вылуплением.[6]
.6.3 Нарушения в эмбриогенезе
Нарушения дробления. Чаще всего встречаются
нарушения, обусловленные полиспермным оплодотворением. При полиспермном
оплодотворении все погрузившиеся в цитоплазму спермин включаются в развитие и
при первом делении дробления сразу возникает три, четыре и больше бластомеров.
Только диспермные яйца в это время еще ничем не отличаются от нормальных
моноспермных - у них в анимальной области закладывается одна борозда. Однако
при втором делении в каждом из первых двух бластомеров возникает по две
борозды, и анимальная область подразделяется на шесть бластомеров. В дальнейшем
полисперные яйца делятся одновременно с нормальными, от которых все они
отличаются наличием сверхчисленных бластомеров.
Глубокие нарушения дробления происходят при
температурных повреждениях цитоплазмы ооцитов: большой разнобой во времени
закладки отдельных борозд, искажения рисунка дробления. Иногда часть яйца
остается неразделенной (мозаичное дробление). При инкубации икры получается
высокий процент резко уродливых эмбрионов. Неразделившаяся часть яйца не только
сама не участвует в развитии эмбриона, но является механическим препятствием
для нормального перемещения клеточных пластов в процессе гаструляции. Подобные
нарушения развития особенно часто встречаются в случаях, когда при высокой
температуре самку поздно вскрыли и произошла задержка овулировавшей икры в
полости тела.[6]
Партеногенетическое дробление. Яйца, активированные
без оплодотворения, как правило, дробятся. Дробление обычно начинается со
значительным опозданием по сравнению с дроблением оплодотворенных яиц и в
дальнейшем также сильно отстает. Однако иногда, если активация произошла еще в
теле самки, дробление активизированных яиц может начаться раньше, чем
оплодотворенных, в некоторых случаях еще до получения икры, в яичнике или
полости тела самки.
Партеногенетическое дробление протекает
беспорядочно, борозды закладываются неодновременно, нередко большая область в
яйце совсем не дробится. В одних случаях дробление рано останавливается, после
закладки всего нескольких борозд, в других -продолжается до стадии нескольких
десятков бластомеров. Эта стадия - предельная, дальше партеногенетическое
развитие не идет, эмбрионы никогда не переходят к гаструляции и медленно
отмирают. Границы клеток' постепенно исчезают, и эмбрион приобретает белесую с
разводами окраску. Отмирание неоплодотворенных яиц (как активированных и
дробившихся, так и оставшихся неактивированными) происходит в то время, когда
оплодотворенные яйца из той же партии икры гаструлируют.[6]
Нарушения гаструляции. Большая часть уродств,
которые встречаются у эмбрионов, возникает в результате нарушения клеточных
перемещений в процессе гаструляции. Если обрастание темных вегетативных клеток
задерживается и эмбрион переходит к следующему периоду развития, сохраняя
желточную пробку значительного размера, то в дальнейшем у него будут
наблюдаться нарушения строения. В случаях, когда такая задержка обрастания
произошла на начальных стадиях гаструляции, эмбрион, как правило, вскоре
погибает.
Существует нарушения гаструляции и другого рода,
затрагивающие не обрастание вегетативных бластомеров, а процесс вворачивания
клеточного материала. Особенно резкие нарушения этого типа возникают у
эмбрионов с неполным дроблением (следствие неблагоприятных условий созревания).
В зависимости от того, насколько велика нераздробившаяся часть яйца и в какой
части эмбриона она оказывается, из них развиваются уроды разного строения - с
резкой степенью недоразвития передних отделов тела или с раздвоением осевых
органов.
Нарушения гаструляции наблюдаются также при
неблагоприятных условиях инкубации - слишком высокой или низкой температуре,
недостатке кислорода (результат перегрузки аппарата, недостаточного омывания
икры водой, заиливания или низкого содержания кислорода в воде). В икре
хорошего качества при благоприятных условиях гаструляция идет дружно, в конце
ее могут встречаться лишь единичные эмбрионы с большими желточными пробками
неправильной формы (такие эмбрионы обычно развиваются из полиспермных яиц).
Если же при развитии икры хорошего качества наблюдается разнобой в размерах
желточной пробки, то это свидетельствует о недостатке благоприятных условий
инкубации.[6]
Нарушения в развитии на 4 этапе. Наиболее частое
нарушение типичного развития на этом этапе заключается в том, что образование
нервной пластинки начинается при наличии желточной пробки большего или меньшего
размера. Если желточная пробка очень велика, то эмбрион развивается резко
атипично. Закладывается укороченная и искривленная нервная пластинка или же она
с самого начала оказывается разделенной на две половины, развивающиеся
независимо одна от другой. Такое раздвоение может затрагивать часть нервной
пластинки. Эмбрион с огромной обнаженной желточной пробкой обычно вскоре
погибает. При меньших размерах желточной пробки развитие может продолжаться
долгое время (до стадии вылупления), но хвост у такого эмбриона оказывается
укороченным и нередко искривленным. Если к моменту образования нервной
пластинки имеется совсем маленькая желточная пробка, то она постепенно
втягивается или отпадает, и развитие продолжается без заметных отклонений от
нормы.
При другом типе нарушений гаструляции,
затрагивающим процесс вворачивания, развиваются эмбрионы с недоразвитыми
передними отделами тела. У них закладывается укороченная нервная пластинка, а
позже отсутствуют передние отделы головы, вся голова или даже голова и часть
туловища. Иногда наблюдаются случаи удвоения, когда у эмбриона бывает две головы
или раздвоен хвост.
Нередко встречаются сложное уродство, при
котором сочетаются одновременно нарушения в развитии головы, туловища и сердца.
Голова у таких эмбрионов совсем не обособляется от желточного мешка или
обособляется в гораздо меньшей степени, чем при нормальном развитии;
заднетуловищный отдел резко укорочен. Сердце раздвоено, иногда образуется два
сердца; в некоторых случаях сердце отсутствует.
Описанное сложное уродство появляется в тех
случаях, когда эмбрионы в конце гаструляции и в период образования нервной
пластинки или же непосредственно перед возникновение зачатка сердца
подвергаются неблагоприятным внешним воздействиям.[6]
Строение атипичных эмбрионов в конце
эмбриогенезе. Если условия инкубации благоприятны, то в хорошей икре попадаются
лишь единичные уроды, в икре плохого качества их бывает до 20 - 25%. Если икра
подвергается неблагоприятным воздействиям (например, высокая температура(
дефекты могут иметь более 90% эмбрионов. Уродливые эмбрионы часто не могут
освободиться из оболочек; менее дефектные вылупляются, но, как правило, с
некоторым опозданием по сравнению с эмбрионами нормального строения.
Обычным типом нарушения развития является
недоразвитие передних отделов тела. У наименее дефектных из них передние отделы
головного мозга бывают только уменьшены. При значительном недоразвитии этих
отделов мозга вместо двух обонятельных мешков и двух глаз может развиваться
только по одному органу, располагающемуся центрально. Далее следуют эмбрионы, у
которых передний отдел головы уже полностью отсутствует; нет ни переднего, ни
промежуточного мозга, нет также обонятельных мешков и глаз; голова у них
начинается со среднего или непосредственно с продолговатого мозга, по бокам от
которого лежат слуховые пузырьки. У других эмбрионов нет и продолговатого
мозга, то есть отсутствует вся голова. Иногда у эмбрионов отсутствует
переднетуловищный отдел: от желточного мешка отходят только дефектные
заднетуловищный отдел и хвост. У большинства уродов такого типа хвост бывает
укорочен и искривлен, у многих отсутствует сердце.
Эмбрионы с недоразвитыми передними отделами тела
делают судорожные движения хвостом, но, как правило, нормально плавать не
могут. Эмбрионы без переднего отдела головы еще более дефектные неподвижны,
даже на укол иглой они не реагируют. Иногда всречаются эмбрионы с удвоением
осевых органов: две головы и два туловищных отдела на одном желточном мешке.
Распространенный тип уродства - нарушение
строения сердца, часто сопровождающееся укорочением хвоста и неполным
обособлением головы. У некоторых эмбрионов сердце имеет вид длинной и узкой
трубочки, а околосердечная полость сильно раздута (следствие патологического
накопления жидкости, называемого водянкой). У других сердце раздвоено или же
имеется два сердца, у третьих сердце полностью отсутствует.
Встречаются эмбрионы с менее значительными
нарушениями - укороченные, искривленные, с водянкой околосердечной полости,
выраженной в разной степени, а также эмбрионы с различными изъянами в
плавниковой оторочке.
Причина большинства уродств - нарушения,
возникающие на самых разных стадиях, в период созревания ооцитов и
оплодотворения. Эти нарушения, в свою очередь, обуславливают атипичное течение
процессов дробления, гаструляции, а затем и всего последующего развития.
Исключениями являются разного рода искривления, которые могут появляться на
более поздних стадиях.[6]
.6.4 Особенности предличиночного периода
Период от вылупления до начала активного
(эндогенного) питания особи имеет не меньшую продолжительность, чем
эмбриональное развитие. Своеобразие этого периода заключается в смене
эмбриональных приспособлений и функций на дефинитивные. В процессе такой смены
изменяются отношения развивающегося организма со средой, это сказывается на его
поведении (отношении к свету, субстрату, течению и др.). На протяжении этого
периода формируется ряд признаков, имеющих систематическое значение и
характеризующих взрослых рыб. Своеобразие этого периода явилось причиной
выделения в развитии рыб особого предличиночного периода, который начинается с
момента выхода эмбриона из оболочек и завершатся переходом предличинки на
активное питание. Однако до сих пор некоторые исследователи продолжают называть
предличинку свободным эмбрионом или просто личинкой, игнорируя тем самым
принципиальные отличия этого периода развития как от эмбрионального, так и от
личиночного периодов.
В предличиночный период развития интенсивно идут
процессы формообразования и дифференцировки, что обуславливает не только
существование специфических для этого периода жизни закономерностей развития,
но и возникновение в неблагоприятных условиях особых нарушений строения,
характерных для предличинок. Для совершенствования биотехники выдерживания
предличинок очень важно уметь отличать эти нарушения от тех, которые возникают
в эмбриональный период, и выяснить факторы внешней среды, которые могут вызвать
образование уродов или даже привести к гибели предличинок.
Предличиночное развитие может быть подразделено
на два этапа - от вылупления до начала активных дыхательных движений и от
начала этих движений до перехода на активное (экзогенное) питание.
этап.

в.п. - выводной почечный проток; в.э. -
впячивание покровного эпителия на месте будущих жаберных щелей; гр.п. - зачаток
грудного плавника; к.п. - кювьеров проток; м.с.
мышечные сегменты; пиг.п. - пигментное пятно в
глазу; п. прод. м. - полость продолговатого мозга; п. с. - плавниковая складка;
хр. хрусталик; ж. в. - железа вылупления; п. петля, образуемая собирающим и
выводным почечными каналами; с.ж. -сеть кровеносных сосудов желточного мешка;
кл. - зачаток клоаки; с. - сердце; об.я. обонятельная ямка; сл.п. - слуховой
пузырек.
Рисунок 34 - 36 ст. Предличинка после выхода из
оболочек в период массового вылупления
На стадии 36 (рис.34) у предличинки ротовое
отверстие и жаберные щели отсутствуют. Еще видна железа вылупления, в глазах
четкое пигментное пятно, кровь розовая.

Рисунок 35 - 37 ст. Начинают открываться ротовое
отверстие первая пара жаберных щелей, имеются чёткие зачатки грудных плавников
На стадии 37 (рис.35) появляются зачатки усиков.
Прорывается ротовое отверстие. Начинается разделение желточного мешка на
желудочный и кишечный отделы. Чётко выражены зачатки грудных плавников в виде
небольших складочек кожи. Зачатки жаберных лепестков отсутствуют. Появляется
зачаток боковой линии сейсмосенсорной системы.

Рисунок 36 - 38 ст. Стадия появления зачатков
жаберных лепестков оперкулярной жабры и лепестков на первой жаберной дуге
На стадии 38 (рис.36) становятся различимы
первые меланоциты. Энтодермальная складка, отделяющая желудок от кишечника,
неполная. Формируются первые мускульные почки в области спинного и анального
плавников. Появляются зачатки жаберных лепестков на жаберной крышке и первой жаберной
дуге. Боковая линия сейсмосенсорной системы достигает уровня кювьерова протока.

Рисунок 37 - 39 ст. Стадия складки стенки
зачатка пищеварительной системы разделяет его на 2 отдела: желудочный и
кишечный
На стадии 39 (рис.37) желудок отделён от
кишечника. Обособились спинной и анальный плавники. Боковая линия
сейсмосенсорной системы достигает уровня заднего края грудного плавника,
появляется добавочный ряд сейсмосенсорной системы.
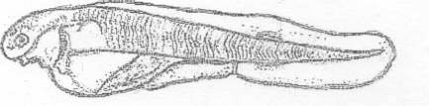
Рисунок 38 - 40 ст. Стадия появления зачатков
брюшных плавников и начала движения нижней челюсти
На стадии 40 (рис.38) различим зачаток брюшного
плавника в виде узенькой складочки кожи. Брюшные отростки мышечных сегментов в
области грудного плавника спускаются ниже его основания. Можно наюлюдать первые
нерегулярные движения нижней челюсти. Боковая линия сейсмосенсорной системы не
достигает уровня конца желудка, добавочный ряд заканчивается надгрудными
плавником.
этап.

Рисунок 39 - 41 ст. Стадия смыкания лопастей,
перегораживающих обонятельное отверстие и начала ритмичных дыхательных движений
На стадии 41 (рис.39) края обонятельных лопастей
смыкаются, но ещё не сращены. Движения нижней челюсти часты. Боковая линия
сейсмосенсорной системы заканчивается над спиральной кишкой, её добавочный ряд
заходит за заднюю границу грудного плавника, образуется короткий зачаток
спинного ряда.
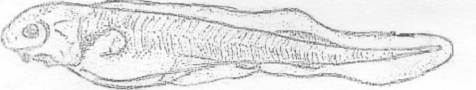
Рисунок 40 - 42 ст. Стадия срастания лопастей,
перегораживающих обонятельное отверстие, и появление зачатка пилорического
придатка кишечника
На стадии 42 (рис.40) появляется зачаток
пилорического придатка. Лопасти обонятельного органа сращены. Боковая линия
сейсмосенсорной системы достигает уровня переднего края брюшного плавника,
спинной ряд начинает изгибаться.

Рисунок 41 - 43 ст. Стадия появления зачатков
вторичных лепестков в первой жабре; брюшные плавники достигают края преанальной
плавниковой складки
На стадии 43 (рис.41) рострум принимает
горизонтальное положение. Брюшной плавник достигает края преанальной складки.
Появляются зачатки вторичных лепестков в первой жабре. Боковая линия
сейсмосенсорной системы достигает уровня заднего края брюшного плавника,
добавочный ряд заканчивается над спиральной кишкой, спинной ряд изогнут и
начинает расти параллельно боковой линии.
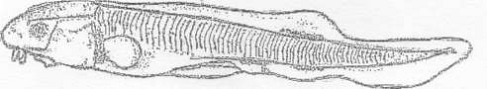
Рисунок 42 - 44 ст. Стадия, на которой края
брюшных плавников выходит за границу преанальной складки; появляется общий
зачаток спинных жучек
На стадии 44 (рис.42) основания усиков выносятся
вперёд, и концы не достигают передней границы рта. Массовый выброс пигментных
пробок. Лопасти брюшных плавников опускаются ниже края преанальной складки. В
спинной плавниковой складке появляется мезенхимная полоска (зачаток жучек).
Передняя поперечная комиссура сейсмосенсорной системы переместилась дорсально,
боковая линия заходит за уровень заднего края брюшного плавника, дополнительный
ряд не доходит до передней границы брюшного плавника.

Рисунок 43 - 45 ст. Стадия перехода на активное
питание. Предличинка способна к хватательным движениям рта, имеет раздельные
зачатки спинных жучек
На 45 стадии (рис.43) переход на активное
питание. Видны раздельные зачатки жучек в спинной плавниковой складке. Боковая
линия сейсмосенсорной системы заходит за уровень средней части спинного
плавника, добавочный ряд почти достигает уровня переднего края брюшного
плавника, спинной ряд заходит за уровень заднего края грудного плавника.
1.6.5 Личиночный период
этап - этап смешанного питания. Дыхание
становится наружно-жаберным. Появляются зачатки брюшных плавников. Рот нижний,
перед ним 4 усика. Голова большая. Рыло вытянутое, постепенно становится
заостренным. Плавниковая кайма на месте не парных плавников обособляется.
Хвостовой плавник большой, гетероцеркальный. Вдоль спинного края плавниковой
складки начинают появляться бугорки - зачатки жучек, сначала они еще не выходят
за пределы складки.
На верхней челюсти закладываются зубы. Питание
смешанное, этап длится три дня. Анальное отверстие, замкнутое у предличинки, у
личинок сформировано. Меланин, имеющийся в кишечнике эмбриона, выделен.
этап - этап экзогенного питания. Желточный мешок
полностью резорбировался, питание экзогенное.
Рот личинки при открывании выдвигается, но
ротовой трубки, типичной для более позднего, малькового этапа, еще нет.
Увеличивается число и длина жаберных лепестков, Жаберная крышка разрастается,
прикрывая собой наружные жаберные лепестки почти до трети их длины. Образовался
спиральный клапан.
.6.6 Мальковый период
Личиночные плавниковые складки резорбируются.
Вентральная поверхность туловища малька становится плоской. Имеются ряды
спинных, боковых и брюшных жучек. На голове развиваются покровные окостенения,
несущие на своей поверхности шипы. Жучки и костные пластинки образуют на теле
малька защитный колючий панцирь. Строение парных и непарных плавников
приближается к дефинитивному. Жаберные крышки разрастаются, полностью покрывая
жаберные лепестки. В отличие от личинок челюсти малька не имеют зубов. Рот
малька выдвигается в виде трубки. Зубы исчезают тогда, когда жаберные крышки
начинают функционировать как всасывающий аппарат. При этом утрачивается
необходимость удерживать добычу зубами. Питание бентосом.
2.
ВЫБОР МЕСТА ДЛЯ ОСЕТРОВОГО РЫБОВОДНОГО ЗАВОДА
При выборе места учитывается наличие
транспортных связей для сообщения с рыбоводным предприятием, размещение
промысловых участков, близость населенных пунктов для обеспечения предприятий
электроэнергией, рабочей силой, снабжения материальными ресурсами, кормами,
удобрениями.
Одним из решающих условий при выборе места для
строительства рыбоводного предприятия является наличие необходимого количества
воды соответствующего качества. В районе водозабора и на участке водоема на
расстоянии 20км выше по течению не должно быть сброса сточных вод промышленных
предприятий.
Рыбоводное предприятие будет располагаться около
г. Малая Фоминка вверх по течению на 2км (рис. 44). Недалеко от города
расположены автомобильная и железнодорожные дороги. Так же рядом находится
большой город - Орёл. Это позволит без каких-либо затруднений закупать
различное оборудование и сырье. Город обеспечит работников жильем, и другими
бытовыми услугами необходимыми для жизни (одеждой, пищей и др). Это очень
экономично для завода. Источником водоснабжения будет служить река Ока. После
выпуска, покатная молодь будет скатываться в Оку.

Х - Местоположение ОРЗ М 1:1000000
Рисунок 44 - Копия карты Орловской области [7]
3.
ГИДРОЛОГИЧЕСКАЯ И ГИДРОХИМИЧЕСКАЯ ХАРАКТЕРИСТИКА РЕКИ ОКИ
.1 Гидрологические условия
После впадения р. Москвы Ока вступает в область
обширной и сильно заболоченной Мещерской низменности [8].
В верховье, примерно на протяжении 400 км, Ока
течет на север, после впадения р. Угры поворачивает на северо-восток и
сохраняет это направление до устья. Река отличается большой извилистостью.
Долина реки в верхней части бассейна узкая и глубокая (40-60 м), овраги сильно
расчленяют ее склоны; ширина русла меняется от 20 до 80 м. Долина Оки большей
частью асимметрична. Ниже г. Серпухова долина расширяется; ширина русла реки
250-400 м. В пределах Мещерской низменности долина неясно выражена, низкие
берега реки покрыты лугами и полями. Ширина долины достигает 10-20 км. На
широкой пойме много стариц и озер. В русле реки часто встречаются мели и
перекаты.
Температура воды в реке после вскрытия сначала
медленно, а затем интенсивно повышается и примерно через месяц достигает
15-22°С. Максимум температуры наблюдался обычно в июле. Достигнув максимума,
температура воды начинала понижаться и к концу октября составляла 3-4°С. Такая
температура удерживалась до осеннего ледохода, а затем при ледоходе и ледоставе
снижалась до О°С на поверхности, сохраняя положительные значения, близкие к
О°С, в придонных слоях. Годовые колебания температуры воды распределялись по
реке равномерно [8].
Среди факторов, влияющих на величину
прозрачности воды в реке, наиболее существенными являются поступление взвешенных
частиц с водами весеннего половодья и дождевых паводков, размыв берегов и
взмучивание донных отложений при волнении, развитие фитопланктона.
Водный режим характеризуется высоким весенним
половодьем, проходящим в конце апреля, частыми летними и особенно осенними
паводками. Сток в течение года неравномерен, сток уменьшается в направлении с
севера на юг. Причиной этого является уменьшение с севера на юг количества
атмосферных осадков и возрастание потерь на испарение: на весну приходится 74%
годового стока, на лето и осень-7% и на зимний период-9%. Наивысшие уровни
наблюдаются в период весеннего половодья. Летом и зимой наблюдаются наиболее
низкие уровни воды в реках.[8]
3.2 Гидрохимические особенности
Питание преимущественно снеговое (до 65% у г.
Мурома), дождевое (более 20%) и подземное (менее 20%). Соотношение источников
питания зависит от площади водосбора реки: чем меньше бассейн, тем больше доля
снегового питания. Увеличение питания рек подземными водами, отличающимися от
поверхностных повышенной минерализацией (и особенно содержанием хлорид- и
сульфатионов), не может не сказаться на химическом составе воды рек.
Река Ока относится к рекам со средней
минерализацией - 200-500мг/л. В составе растворенных веществ преобладают ионы
HCО3 (15% экв); по преобладающему иону воды здесь принадлежат к
гидрокарбонатному классу. Наряду со средней минерализацией водам Оки
свойственно пониженное содержание анионов сильных кислот, особенно сульфатов
(9,5 % экв.). По катионному составу воды гидрокарбонатного класса относятся
почти исключительно к группе кальциевых вод. Величина жесткости является
функцией общей минерализации: чем выше минерализация, тем больше жесткость.
Жесткость измеряется в мг-экв Са (0,7 мг/л) и Mg (1,2 мг/л) в 1 л воды. Ока
относится к умеренно жестким рекам - 3-6 мг-экв. Повышение содержания солей в
меженный период невелико.
Воды Оки богаты соединениями фосфора и азота.
Общее содержание фосфора в реке в среднем составляет 200 мкг Р/л., а азота -
2,2 мгN/л. [8]
Таблица 1 - Гидрохимические показатели [8]
|
Показатели
|
Гидрохимическая
характеристика р. Оки
|
Гидрохимические
требования для стерляди
|
|
Кислород,
мг/л
|
8,5
|
7-8
|
|
Диоксид
углерода, мг/л
|
2,09
|
До
10
|
|
Водородный
показатель, pH
|
7,3-8,1
|
7-8
|
|
Щелочность,
мг-экв/л
|
1,68
|
До
2
|
|
Жесткость
общая
|
4-8
|
До
12
|
|
Окисляемость,
мгО/л
|
6-12
|
5-15
|
|
Азот,
мгN/л
|
3,2
|
4
|
|
Фосфор,
мгР/л
|
2
|
6,3
|
|
Хлориды,
мгCl/л
|
0,78
|
до
5
|
|
Сульфаты,
мгSO4/л
|
0,95
|
до
5
|
Исходя из вышесказанного следует, что по
гидрологическим и гидрохимическим показателям вода из выбранного источника
водоснабжения соответствует биологическим требованиям стерляди.
4.
ОПИСАНИЕ ТЕХНОЛОГИЧЕСКОГО ПРОЦЕССА ОРЗ НА РЕКЕ ОКА
.1 Отлов производителей стерляди
Строительство рыбоводных заводов и разведение
стерляди может стать мощным фактором в пополнении запасов ценных видов рыб рек.
Биотехнический процесс разведения стерляди весьма близок с таковым, действующим
на рыбоводных заводах, на которых разводят анадромных осетровых. Однако он
имеет свои специфические особенности.
Учитывая то обстоятельство, что в первые годы
работы стерляжьего рыбоводного завода, когда промысловые запасы стерляди в реке
еще незначительны или вообще отсутствуют, возникает проблема заготовки их
производителей. В этом случае производителей следует отлавливать в местах
наибольших их концентраций, в местах нагула.
При невозможности же заготовить необходимое
количество производителей формируют на заводе собственное маточное стадо
стерляди, принимая в нем соотношение самок и самцов 5:1. Возраст впервые
используемых производителей для взятия половых продуктов: самцы - 4 - 5 лет,
самки - 6 - 10 лет. В возрасте 4 - 5 лет созревает 50 % самцов.
Возраст, при котором самки становятся впервые
зрелыми, более растянут во времени. Их возрастной состав следующий (%):
шестигодовики - 5 - 10; семигодовики - 5 - 10; восьмигодовики - 10 - 15;
девятигодовики 15 - 25; десятигодовики - 30 - 40. В дальнейшем эти самцы
созревают ежегодно, а самки только через 2 - 4 года. Маточное стадо
производителей пополняется за счет ремонтного стада, включающего младшие
возрастные группы особей. Маточное стадо производителей стерляди содержат на
заводе в бассейнах Б.Н. Казанского [9].
Отлов производителей в естественных условиях
осуществляют весной в преднерестовый и нерестовый периоды и осенью, применяя
плавные и ставные сети, закидные невода и тралы.
Весной, когда температура воды еще не превысила
10°С, можно приступать к заготовке производителей стерляди. Их отлов лучше
проводить плавными сетями, так как ставные сети нужно часто в течение суток
перебирать (проверять улов), а в закидных неводах и тралах много производителей
травмируется. Для рыбоводных целей выбирают из орудий лова стерлядь с половыми
железами в IV стадии зрелости.
Весной у этих рыб наблюдается брачный наряд. На
их голове появляется рисунок в виде белых узких полосок эпидермиального
происхождения. Он отчетливо заметен на
теменных костях от начала рострума до конца
головы со спинной части рыбы и, особенно на бугорках около обонятельных капсул.
Признаком этой стадии зрелости половых желез у стерляди являются также
припухлость и покраснение генитального отверстия.
Самки отличаются от самцов более толстым и более
мягким брюшком, по середине которого, как правило, проходит темно-фиолетовая
полоса. Средняя масса отбираемых производителей должна составлять 1,5 - 2 кг у
самок и 1 - 1,5 кг у самцов. Минимальная масса самок должна быть не менее 0,5 -
1 кг, а самцов - 0,3 - 0,5 кг [9].
Осенью у стерляди нет брачного наряда, поэтому
ее производители с половыми железами в IV стадии зрелости отличаются от таковых
во II стадии зрелости только более толстым брюшком. Что касается особей с IV и
III стадиями зрелости половых желез, то их вообще нельзя отличить визуально,
ибо все они имеют толстое брюшко. В связи с этим всех отловленных осенью
производителей следует перевезти на завод и посадить в зимовальные пруды
площадью 0,25 га и глубиной 2,5 - 3 м. В каждом таком пруду можно содержать до
1000 производителей. После зимовки производителей осматривают и выбраковывают
особей с половыми железами в III стадии зрелости. Этих производителей можно
отсадить в нагульный пруд, где за лето их половые железы развиваются до IV
стадии зрелости. После второй зимовки на заводе они могут быть использованы
весной для рыбоводных целей.
Производителей стерляди, отловленных весной и
осенью, доставляют на завод в прорези, в которую помещают не более 20 - 30
особей/м3. Скорость движения судна с прорезью, загруженной производителями, не
должна превышать 10 км/ч. Для перевозки производителей на завод, удаленный от
места их лова на 150 - 200 км, можно использовать также живорыбную машину, в
цистерну которой помещают 50 - 100 особей. В пути следования воду в цистерне
нужно аэрировать.
Отход производителей стерляди за время перевозки
на завод обычно не превышает 5 % [9].
.2 Выдерживание производителей стерляди
Весной на заводе производителей стерляди с
половыми железами в IV стадии зрелости можно выдерживать в бассейнах Б. Н.
Казанского с терморегулирующим режимом. В каждый бассейн помещают 300 самцов
или 150 самок (рис. 45).
Длительность выдерживания производителей в
бассейнах 20 - 30 суток в зависимости от прогрева воды, поступающей на завод, и
связанных с ним сроков наступления нерестовых температур. Гидрохимические
показатели воды должны удовлетворять тем же требованиям, которые предъявляются
на рыбоводных заводах по разведению других видов осетровых рыб [9].

Рисунок 45 - Бассейны Казанского для
выдерживания производителей осетровых [10]
Заготовку производителей ведут во время
нерестовых миграций. Отловленных производителей рыбоводы осматривают и отбирают
самок и самцов в определенном соотношении, без травм, кровоподтеков, среднего
размера и массы.
Помимо визуальных признаков, при помощи которых
у производителей определяют завершенную 4 стадию зрелости половых желез, можно
также устанавливать состояние гонад самок модифицированным методом «щуповых
проб» Гончарова, отбраковывая несозревших самок и самок, дающих икру низкого
рыбоводного качества. Взятые с помощью щупа фолликулы помещают в раствор
Рингера с прогестероном 5 мкг/мл и через 18τ0 варят
в воде, разрезают и определяют, сохранилось ли у них ядро. Самок, в ооцитах
которых ядро сохраняется дольше 18τ0, не
следует использовать для рыбоводных целей.
Щуп позволяет брать гонады и у самцов. Мазок
семенника на предметном стекле рассматривается под микроскопом при среднем
увеличении и косом освещении. Наличие множества сформированных сперматозоидов
свидетельствует о возможности их нормального созревания. Отбирают самцов,
сперма которых отличного, хорошего и удовлетворительного качества по пятибалльной
шкале Персова.[1]
С наступлением нерестовых температур самок и
самцов разделяют на группы по 30 особей, а затем каждой группе рыб делают
внутримышечные инъекции сурфагона. Созревание в большинчтве случаев наступает
после однократного введения препарата дозой 100мгк/кг. Продолжительность
созревания половых клеток стерляди при использовании сурфагона на 10 - 25%
больше, чем у рыб, инъецированных препаратом гипофиза [1].
Выдерживание самок и самцов после применения
сурфагона проводят отдельно в бассейнах Б. Н. Казанского. Скорость созревания
половых продуктов у производителей различная и зависит от температуры воды и
индивидуального физиологического состояния рыбы [1].
.3 Взятие зрелых половых продуктов у
производителей, осеменение икры и подготовка её к инкубации
Проверку производителей с целью установления
готовности их половых продуктов для рыбоводных целей начинают за 2-3 ч до
предполагаемого появления первых зрелых особей. Затем этот контроль за
состоянием половых желез производителей продолжают осуществлять на протяжении
всего периода их выдерживания в бассейнах.
При опускании этих самок вниз головой икра
свободно переливается в полости тела, что можно наблюдать по перемещению
вздутия брюшка к голове.
При поднятии самок вверх головой или при легком
массировании их брюшка в сторону задней части тела из генитального отверстия
вытекает полостная жидкость с икрой. Зрелые икринки имеют на анимальном полюсе
хорошо выраженный рисунок из темных и светлых колец.
По мере появления зрелых самок с овулированной
икрой и зрелых самцов с текучей спермой их вынимают из бассейнов, помещают в
брезентовые носилки с водой и доставляют в операционное отделение при
инкубационном цехе, где берут у производителей зрелые половые продукты [1].
Сперму получают от самцов способом отцеживания.
Ее отцеживают в небольшую сухую посуду (в химический стакан, фаянсовую чашку и
др.). При взятии у самцов спермы обращают внимание на ее качество. Сперма,
выходящая из генитального отверстия самца, кремового и белого цвета, и имеющая
консистенцию сливок, является, как правило, доброкачественной, отличается
высокой концентрацией и активностью сперматозоидов. Жидкая по консистенции
сперма, синеватого оттенка или почти прозрачная является недоброкачественной.
Она содержит небольшое количество сперматозоидов с очень низкой активностью.
Такую сперму отбраковывают.
От одних и тех же самцов можно получить
несколько порций спермы. Так, после получения первой порции спермы самцов
отсаживают в бассейны, а через 2 - 4 ч у них берут вторую порцию. Таким путем
от самцов получают до 3 - 5 порций спермы. В связи с этим количество спермы,
которое можно получить от одного самца, составляет 30 - 60 мл [1].
Икру получают от самок методом надрезания
яйцевода, разработанный С.Б. Подушка. Яичники осетровых не имеют собственной
полости, и овулировавшая икра попадает непосредственно в полость тела. Пред
тем, как попасть во внешнюю среду яйца должны пройти через яйцеводы. Воронки
яйцеводов значительно удалены от генитального отверстия в краниальном
направлении. Эти анатомические особенности половой системы самок объясняют,
почему у осетровых нельзя отцедить всю овулировавшую икру сразу. Массаж брюха
самки от головы к хвосту приводит к выдавливанию икры только из яйцеводов,
после чего их стенки спадаются, и дальнейшее сцеживание оказывается
невозможным. После надреза каудального участка одного их яйцеводов
овулировавшая икра может поступать к генитальному отверстию непосредственно из
полости тела, минуя яйцеводы, и отцеживание икры можно осуществлять обычным путём.
Рыбу размещают на операционном столе брюхом
вверх, так, чтобы её хвостовой стебель свисал. Брюхо и хвостовой стебель
протирают сухим полотенцем, чтобы предотвратить попадание воды и слизи в таз с
икрой. Первоначально отцеживают икру из яйцеводов. После того, как дальнейшее
выделение икры прекращается, в генитальное отверстие, которое находится
каудальнее анального, вводят скальпель. Скальпель вводят латерально, направляя
его в правый или левый яйцевод. Делают небольшой надрез яйцевода, режущий край
скальпеля при этом должен быть направлен вверх, к брюху рыбы. После надреза
яйцевода поступление икры к генитальному отверстию возобновляется. Для удобства
отцеживания самку можно положить на бок. Отцеживание продолжается до тех пор,
пока икра свободно вытекает из полости тела. По окончании отцеживания рыбу
полезно поднять головой вверх и согнать остатки икры к генитальному отверстию.
При первом отцеживании у самки извлекают основную часть овулировавшей икры
(80-90%). Через час после первого отцеживания целесообразно провести второе.
Второе отцеживание не требует нового надрезания яйцевода и осуществляется очень
быстро [1].
Количество производителей, у которых не удается
получить доброкачественные половые продукты, составляет 10 % у самцов и 30 % у
самок (от числа заготовленных особей в реке).
Взяв от производителей половые продукты,
приступают к осеменению икры. Первоначально удаляют из таза с икрой полостную
жидкость. Затем икру взвешивают. Осеменение икры производят полусухим способом.
Для этого в таз с икрой выливают разбавленную водой сперму в соотношении 1: 150
или 1: 200 в зависимости от ее качества. На осеменение 1 кг икры расходуют 10
мл спермы. После добавления в таз спермы икру перемешивают в течение 5 мин. За
это время происходит оплодотворение.
Для определения процента оплодотворения лучше
всего брать пробу на стадии второго деления дробления 4 бластомера, стадия 5.
Время взятия пробы определяют по кривой графика, выражающей зависимость времени
наступления второго и деления дробления эмбрионов от температуры. Определяем
среднюю температуру за период, пршедший с момента осеменения икры. Находим на
горизонтальной оси графика точку, соответствующую этой температуре, и опускаем
перпендикуляр до пересечения с кривыми. Брать пробу следует в интервале времени,
ограниченном I и III кривыми, ближе к кривой II, т.е. на стадии завершенного
второго деления. Прежде чем брать пробу тщательно перемешиваем икру в аппарате
и берут 200-300 икринок. Сначала разбирают пробу, разделив дробящиеся и
недробящиеся яйца, затем пересчитываем их и вычисляем процент оплодотворения,
который совпадает с процентом дробящихся яиц. Активированные неоплодотворённые
яйца начинают дробиться с опозданием и в это время, как правило, ещё не имеют
борозд на своей поверхности. Оплодотворяемость икры составляет 95 %.
Оплодотворенную икру промывают, освобождаясь от
спермы, а затем приступают к ее обесклеиванию, которое осуществляется в
аппаратах АОИ (рис. 46).

Рисунок 46 - Аппарат обесклеивания икры - АОИ
[1]
Икру обесклеивают водной суспензией тонкого ила
(0,5 кг ила и 10 л воды на 1 кг икры) в течение 40 мин. По окончании этого
процесса икру промывают чистой водой и переносят из аппаратов АОИ в инкубатор
«Осетр» (рис. 47), который состоит из инкубатора и сортировочного устройства,
соединенных между собой в технологическую линию (рис. 48). Количество икры,
загруженной в инкубатор должно быть не более 32 кг или 2300 тыс.шт. Количество
ящиков, в которые загружается инкубируемая икра составляет 16шт. Учет икры осуществляется
объемным методом [9].

Рисунок 47 - Инкубатор «Осётр» [1]
4.4 Инкубация икры
При температуре воды 10-16°С инкубация икры
продолжается 6 - 9 суток. В этот период необходимо постоянно поддерживать
проточность воды в аппаратах, следить за нормальным гидрохимическим режимом, не
допуская снижения насыщения воды кислородом на вытоке ниже 70 %. Кроме того,
необходимо систематически отбирать мертвые и пораженные сапролегнией икринки.
Для борьбы с сапролегнией в процессе инкубации икры применяют малахитовую
зелень или метиленовый синий из расчета 150 мг вещества на одну ванну аппарата,
содержащую около 60 л воды. Эти вещества растворяют в горячей воде, а
полученный раствор разбавляют холодной водой и выливают в аппараты с
инкубируемой икрой, подач» воды в которые должна быть приостановлена. Икру
выдерживают в течение 15 - 30 мин, а затем возобновляют подачу воды в аппараты.
Обработку икры этими веществами проводят 2 - 3 раза за время инкубации.
Для определения размеров отхода и числа уродливо
развивающихся зародышей рекомендуется брать 3 пробы. Перед взятием проб икру
нужно тщательно перемешать в аппарате, так как погибшая икра обычно отделяется.
Объем проб не меньше 200-300 икринок [9].
) На стадиях гаструляции и начала нейруляции
(ст.18-19). К концу гаструляции погибают все неоплодотворившиеся яйца, а
типично развивающиеся эмбрионы совсем не имеют желточных пробок или эти пробки
очень маленькие. Чтобы разобрать пробу, надо, прежде всего отделить погибшие,
обычно мраморного вида яйца и определить процент их от общего числа икринок в
пробе(полученное число характеризует размер отхода за счет, главным образом,
неоплодотворившихся яиц). Затем следует определить уродливых эмбрионов с
большой желточной пробкой неправильной формы. Число их зависит от качества икры
и условий осеменения, но не от условий инкубации. В остальной части пробы надо
просмотреть насколько сходно и типично строение разных эмбрионов. Если в пробе
одновременно встречаются эмбрионы с желточными пробками разного размера и
нейруляция начинается до полного втягивания желточной пробки внутрь эмбриона,
то это свидетельствует о недостаточно благоприятных условиях развития. При
улучшении условий такие отклонения могут выровняться.
) В период закладки сердца (ст.26-27). Для
определения процента отхода нужно определить дегенерирующие яйца и погибших
эмбрионов. При хороших условиях инкубации обычно процент отхода во второй пробе
равен или ненамного выше, чем в первой. У живых эмбрионов на этих стадиях легко
определить нарушение в развитии головы. Число эмбрионов с аномалиями такого
типа характеризует не условия инкубации, а качество икры.
) Перед началом вылупления (ст.35). Для учета
эмбрионов с резкими аномалиями строения в конце периода инкубации следует,
после тщательного перемешивания икры в аппарате взять 300-500 эмбрионов и
поместить их в кристаллизатор со свежей водой. В этих условиях вылупление
длится не менее суток, поэтому воду нужно менять каждые несколько часов.
Необходимо избегать перегрева воды, так как в этом случае могут возникать
параличи и эмбрионы могут быть искривленными. Когда большая часть предличинок
вылупиться, их нужно отсадить и исследовать оставшихся в кристаллизаторе
эмбрионов. Среди них всегда есть отмершие неоплодотворенные яйца, могут встречаться
погибшие эмбрионы, а также живые эмбрионы, оставшиеся в оболочках. Если
вылупление не закончилось, то часть из последних может иметь нормальное
строение. Со всех невылупившихся эмбрионов надо снять оболочки при помощи остро
отточенных пинцетов и внимательно их рассмотреть. Сосчитав общее количество
живых предличинок и эмбрионов, оставшихся в оболочках, следует вычислить, какой
процент от них составляют уроды. При хорошем исходном качестве икры и
благоприятных условиях инкубации уродов бывает очень мало.
Отход икры за период инкубации не превышает 30
%. При соблюдении биотехнических условий получения половых продуктов от
производителей, осеменения икры, подготовки ее к инкубации и проведения самой
инкубации икры количество вылупившихся предличинок достигает 85-95 % от числа
заложенных на инкубацию икринок [9].
.5 Выдерживание предличинок и подращивание
личинок
Личиночно-выростная база индустриального типа
основана на методике перевода личинок осетровых рыб на экзогенное питание в
ограниченных объемах воды.
Это помещение с кондиционерами, компрессорными
установками, воздухоразводкой, резиновыми отводами, распылителями, неокисляемым
металлом и пластмассовыми бассейнами объемом 2-3м3. Чаще всего применяются
бассейны 1,2х2,4х0,7, с донным водоспуском. На один бассейн должно быть два
распылителя. Температура в помещении должна быть 23оС. Бассейны заполняют водой
за 1-2 суток до зарыбления. После отстаивании воды, из бассейнов удаляется
осадок, затем включается аэрация. Содержание О2=7-12 мг/л; СО=3-5 мг/л; рН=7,8-8
мг/л.
Бассейны заполняются один раз и вода в них
недоливается. После полной готовности бассейна, их зарыбляют однодневными
предличинками объёмным методом учета. Плотность посадки нормативна. На 1м3 воды
45 тыс.шт. предличинок стерляди. Вылупившиеся предличинки имеют массу 7-12 мг.
Однодневные предличинки не реагируют на свет и механическое воздействие.
Равномерно распределяются по всей толще воды. Чистка бассейна спустя 2-3 дня.
Постоянное течение имитирует естественные
условия. В этих условиях предличинки находятся до перехода на экзогенное
питание и пересадки в выростные пруды. Плотность посадки в бассейны составляет
45-50 экз/л, выживаемость до 80-90% (в сетчатых садках выживаемость 65-75%).
Подращивание личинок осуществляем в выростных
прудах, куда их помещают после перехода на экзогенное питание.
Кормовая база прудов, в которые пересаживают
личинок, должна быть подготовлена заранее. Плотность посадки личинок в пруды
составляет 70-90 тыс.шт/га [10].
.6 Выращивание молоди
Молодь выращивают в прямоугольных прудах,
которые используют и на осетровых рыбоводных заводах. Их площадь - 2 - 4 га.
Средняя глубина - 2 м. Ложе прудов должно быть спланировано и свободно от
водной растительности. Подача и сброс воды производятся с противоположных
сторон. Полное освобождение пруда от воды происходит за 1 - 2 суток. Подготовку
этих прудов для выращивания молоди стерляди начинают с осени, когда их ложе
осушают, вносят в качестве органического удобрения навоз из расчета 2 - 3 т/га
и вспахивают. Весной, когда температура повышается до 7 - 13 °С, пруды заливают
водой, а через 3 - 5 суток удобряют. Если в прудах наблюдается массовое
развитие листоногих раков, оказывающих отрицательное влияние на
рыбопродуктивность, то первоначально проводят их хлорирование, а спустя 5 - 7
суток удобряют. Из органических удобрений вносят в пруды скошенную, подвяленную
растительность (100 кг/га), которую раскладывают пучками по урезу воды вдоль
дамб, а также кормовые дрожжи (10 кг/га). Из минеральных удобрений применяют
аммиачную селитру (75 - 90 кг/га) и суперфосфат (50 - 90 кг/га). В удобренные
пруды вносят культуру дафний (5 - 7 кг/га). Спустя неделю после такой
подготовки прудов в них формируется хорошая кормовая база. Биомасса
зоопланктона достигает 20 - 30 г/м3. Успешно развивается зообентос, особенно
личинки хирономид.
В водоёме молодь держится в придонных слоях
воды, иногда собираясь возле места подачи воды. В прудах молодь, обеспеченная
полноценной естественной пищей, отличается высоким темпом роста. Одновременно с
этим осуществляют контроль за сохранением нормального гидрохимического и
гидрологического режима в прудах, не допуская снижения уровня воды и содержания
в ней кислорода ниже 6 мг/л, повышение углекислоты более 10 мг/л, активной
реакции среды более 8, окисляемости более 15 мг 02/л. Все это обеспечивает
быстрый рост молоди и хорошую ее упитанность. В возрасте 40-45 суток молодь
стерляди достигает массы 3 г и ее выпускают в реку. Выживаемость молоди в
прудах составляет не менее 50 % от числа посаженных личинок. Производственный
процесс разведения стерляди на рыбоводном заводе завершается выпуском молоди в
реку [9].
4.7 Уход за предличинками, личинками и молодью
стерляди
Ежегодно перед началом рыбоводного цикла все
рыбоводные емкости нужно промыть, продезинфицировать, за каждой емкостью
закрепить необходимый рыбоводный инвентарь.
В течение всего периода выдерживания,
подращивания и выращивания необходимо:
систематически контролировать уровень и
проточность воды, освещенность, содержание в воде кислорода, углекислоты,
окисляемость, рН, температуру;
не допускать проникновения из водоисточника в
рыбоводные емкости сорных рыб (следует устраивать песчано-гравийные
биофильтры);
соблюдать оптимальную плотность посадки в
зависимости от массы;
контролировать поедаемость корма, работу фильтров.
Сбор остатков корма, ила, погибших особей
необходимо осуществлять с большой осторожностью, не допуская беспокойства и
травмирования выращиваемых объектов. При появлении инвазионных заболеваний,
вызываемых эктопаразитическими простейшими, систематически (не реже одного раза
в 10 дней) в профилактических и лечебных целях следует проводить обработку рыб
в бассейнах с раствором формалина, малахитового или бриллиантового зеленого,
фиолетового "К", поваренной соли, морской воды.
Рыбоводный инвентарь (сачки, тазы. ведра, шланги
и др.) в течение всего периода выращивания молоди ежедневно после окончания
работы дезинфицировать в растворе NаС1 (5%).
Все рыбоводные емкости (пруды, питомники,
бассейны, садки) после выпуска молоди необходимо очистить от ила, растений,
остатков корма и продезинфицировать одним из следующих растворов:
5% хлорной извести -1 ч;
% негашеной извести -2 ч;
0,5% перманганата калия - 24 ч [1].
.8 Мелиоративные мероприятия
Мелиоративные мероприятия на осетровых прудах
включают в себя мелиоративные мероприятия по улучшению качества воды, борьбу с
заилением, зарастанием прудов и биологическую мелиорацию.
Мелиоративные мероприятия по улучшению качества
воды заключаются в удалении избытка закисного железа, очистке воды и аэрации.
Для удаления закисных солей железа, отрицательно
влияющих на рыбу и кормовую базу прудов, устанавливают аэрирующие
приспособления и известковые фильтры. Вредные стоки промышленных или
сельскохозяйственных предприятий подвергают полной механической и биологической
очистке на очистных сооружениях, а затем сбрасывают в водоем. Если сточные воды
с сельскохозяйственных полей содержат в своем составе гербициды и минеральные
удобрения, то их перехватывают нагорными каналами и сбрасывают в нижний бьеф
водоема или в него после предварительной очистки в отстойниках.
Нужного насыщения воды кислородом достигают с
помощью аэраторов путем разбрызгивания ее или продувания через нее воздуха.
Аэраторы устанавливают в водоподающих каналах или в прудах. Число и мощность их
определяют расчетом.
Для улучшения качества почвы рыбоводные водоемы
осушают после вегетационного сезона на осенне-зимне-весенний период, а через
5-6 лет и на летний период (выводят на летование). Между рыбоводными сезонами и
в период летования ложе водоемов распахивают и используют для посадки
сельскохозяйственных культур [1].
Заиление водоемов ухудшает условия обитания рыб.
Толщина ила ежегодно увеличивается в среднем на 5 мм. При слое 10-20 см ил
полезен, так как под действием кислорода минерализуется и обеспечивает питанием
растения и кормовых беспозвоночных. При дальнейшем увеличении слоя ила
снижается рыбопродуктивность, замедляются процессы минерализации, создаются
условия для развития врагов рыб и распространения болезней, происходит
закисание почв, ухудшается качество воды, образуется сероводород.
Очистка водоемов от ила очень трудоемка, поэтому
целесообразно принимать меры, замедляющие этот процесс. Применяются следующие
профилактические способы борьбы с заилением:
) запрещение распашки склонов на расстоянии не менее
200 м от уреза воды, склоны водосборной площади за пределами этого расстояния
вспахиваются в горизонтальном направлении - поперек склона;
) устройство нагорных каналов, отводящих
стекающую со склонов воду за пределы прудов;
) залужение прибрежной полосы водоемов;
) посадка на расстоянии не менее 10-20 м от
уреза воды лесных полос шириной 15-25 м, чередующихся с посевом полосами
луговых трав.
) устройство на водозаборных сооружениях
фильтров для очистки воды,
) закрепление оврагов плетневыми запрудами из
живых ивовых кольев от разных видов ив;
) выведение спускных водоемов на летование,
посадка сельскохозяйственных культур [1].
Настоящим бичем рыбоводных хозяйств является их
чрезмерное зарастание высшей водной растительностью. Жесткая растительность ухудшает
газовый режим, снижает содержание биогенных элементов, препятствует освещению и
прогреву воды. Все это приводит к обеднению кормовой базы, снижению
рыбопродуктивности выростных водоемов, выживаемости молоди, уменьшению ее
конечной массы. В заросших водоемах нецелесообразно применять удобрения, так
как они будут использованы растениями зарослей.
Рыбоводная практика показала, что даже
многократное выкашивание камышекосилками жесткой водной растительности не дает
высокого эффекта. Лучший эффект дает вспашка ложа прудов осенью и последующее
летование прудов. В этом случае жесткая растительность угнетается в большей
степени.
Хороший эффект подавления жесткой растительности
обеспечивает разноглубинная вспашка и посев сельскохозяйственных культур.
Например, вспашка на глубину 30-45 см с последующим посевом озимой ржи (снижает
биомассу тростника более чем на 70-74).Можно использовать яровой ячмень.
Хорошим нерестовым субстратом являются многолетние травы. Доказано, что очень
устойчив к продолжительному глубокому затоплению (40 дней) мятлик болотный,
канареечник тростниковидный. Эти культуры могут многократно использоваться
рыбами в качестве нерестового субстрата [1].
В качестве биологических конкурентов жесткой
растительности следует использовать однолетние сельскохозяйственные культуры,
которые за короткий период после осушения рыбоводных водоемов достигают
технической или полной спелости. Наиболее перспективными являются суданская
трава, гречиха и чина. Эти культуры подавляют развитие высшей водной растительности
на 80%, улучшают физико-химические свойства почвы, предотвращают заболачивание.
При этом рыбопродуктивность прудов увеличивается в 1,5-2 раза, а реализация
выращенной травы окупает все затраты на агромелиоративные работы и даже дает
прибыль. Перспективны посевы кукурузы на силос. Во время летования ложе прудов
целесообразно засевать овсом, викой, кормовой свеклой, картофелем, капустой,
огурцами, кабачками, зернобобовыми культурами.
Биологическая мелиорация. Для борьбы в водной
растительностью в рыбохозяйственных водоемах разводят нутрий, ондатр, уток,
гусей. 0дна утка за день поедает до 1 кг зеленого корма, а нутрия за сезон
уничтожает до 500 м2 зарослей. Весьма интенсивно поедает водную растительность
белый амур.
Как показала практика механический и химический
методы борьбы с "цветением" оказались малоперспективными. Больше
всего для борьбы с чрезмерным развитием фитопланктона подходят толстолобики:
белый (питается фитопланктоном) и пестрый (питается фито- и зоопланктоном).0ни
успешно поедают и усваивают сине-зеленые водоросли. Для борьбы с сорной рыбой
используют хищных рыб - щуку, судака, угря, сома, форель [1].
.9 Выпуск молоди, мероприятия, обеспечивающие
наибольшее её выживание
Важным фактором выживаемости молоди является
приобретение защитных рефлексов по отношению к хищным рыбам. У заводской молоди
можно выработать защитные рефлексы, подсаживая в выростные пруды за неделю до
выпуска молоди хищных рыб.
Ускорению выработки оборонительных рефлексов у
мелкой молоди способствует совместный выпуск ее с крупной молодью.
Поэтому выпускать молодь необходимо в местах
нагула. Здесь возможность встречи с врагами оказывается минимальной, а кормовая
база - лучшей. При выпуске следует ориентироваться на температурный и уровенный
режим в реке, состояние кормовой базы.
В период высокой воды целесообразно использовать
гибкие трубопроводы и сетные коридоры для выпуска молоди на стрежень реки. Это
позволяет существенно повысить выживаемость молоди благодаря уменьшению
вероятности встречи с речными хищниками, которые концентрируются в прибрежных
участках реки на мелководьях.
Учет молоди осуществляется устройством подсчёта
рыб УПР-1. Предназначено для автоматического подсчёта молоди рыб на основе
прерывания ультразвукового сигнала. Погрешность счёта - 10%. Быстродействие -
50тыс.шт./ч. Скорость потока водорыбной смеси - 1-3мс/с. Масса - 30кг.
Транспортировка молоди стерляди для выпуска в
реку осуществляется в полиэтиленовых пакетах. Для нормального протекания
процесса газообмена при транспортировке достаточно небольших колебаний в
пакете. Насыщение воды кислородом зависит не от силы тряски, а от её
продолжительности. При длительных остановках необходимо встряхивать пастерляди
в течении нескольких минут. При значительной разнице между температурой воздуха
и воды при транспортировке в пакете в коробки рядом со стандартным пакетом
помещают 1 или 3 кг льда, упакованный в отдельный полиэтиленовый пакет. Если во
время транспортировки ожидается резкая смена температуры, то в картонной
коробке вокруг пакета помещают теплоизоляционный материал. При выпуске рыбы
пастерляди помещают в водоём и вскрывают их после выравнивания температуры воды
в пакете и водоёме [9].
5.
СОСТАВ ОСЕТРОВОГО РЫБОВОДНОГО ЗАВОДА
ОРЗ включает в себя:
цех работы с производителями с отделениями:
) выдерживание производителей в бассейнах
Казанского;
) операционные и лабораторные пункты и склад для
инвентаря;
цех живых кормов;
инкубационный цех;
блок вспомогательных помещений (административный
центр, гараж, мастерская, склад, лаборатория);
объекты энергетического хозяйства;
цех бассейнового выращивания;
выростные пруды;
насосная станция, система водоснабжения и
очистки воды (рис. 49).
Водоснабжение осуществляется при помощи насосной
станции через водоподающую сеть. Вода подается в цеха и без очистки поступает в
водоотводящую сеть. Производится ежедневный лабораторный контроль качества
речной воды, чтобы исключить возможной действие загрязнителей. В случае
превышения показателей загрязнителей, водоснабжение с реки закрывается и вода
подается из артезианской скважины, предварительно пройдя контроль качества.
Водообеспеченеие административного здания
осуществляется из артезианской скважины. Отработанная вода проходит через
очистные сооружения и попадает в водоотводящую сеть.
Водоотвод происходит через водоотводящую сеть в
реку, предварительно пройдя систему очистки.
Водопровод рыбоводного завода оснащен фильтром -
отстойником и водонапорным баком. Водосброс обязательно впадает в реку ниже по
течению оголовка водозабора насосной станции [1].
1 - причал; 2 - цех работы с производителями; 3
- бассейны Казанского; 4 -операционный пункт; 5 - склад для инвентаря; 6 -
насосная станция; 7 - водоподающая сеть; 8 - инкубационный цех; 9 -
лаборатория; 10 - бассейны шведского типа; 11 - пруды для выращивания молоди;
12 - цех живых кормов; 13 - артезианская скважина; 14 - административное
здание; 15 - отстойник с хозяйственно-бытовыми стоками; 16 - сбросной канал; 17
- бетонированная площадка; 18 - склад; 19 - гараж, мастерская; 20 - водозабор.
Рисунок 49 - Схема ОРЗ
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
Серпунин
Г.Г. Биологические основы рыбоводства. - М.: Колос, 2009. - 384 c.
Скорняков
В.И, Апполова Т.А, Мухордова Л.Л. Практикум по ихтиологии. - М.: Агропромиздат,
1986. - 269 с.
Болдина
И.К. Некоторые особенности биологии стерляди // Труды института биологии
внутренних вод. - М., 1966. - Вып.10. - С. 32-33.
Берг
С.В. Рыбы пресных вод России. - М., 1916. - 376 с.
Афанасьев
М.В. Биология размножения стерляди в условиях незарегулированной Волги // Труды
ГОСНИОРХА. - 1980. - Вып.157. - С. 43-45.
Детлаф
Т.А. и др. Развитие осетровых рыб / Т.А. Детлаф, А.С. Гинзбург, О.И.
Шмальгаузен // - М.: Наука, 1981. - С. 224.
Карта
России - <#"727834.files/image057.gif">
