Походження мутацій
Відділ
освіти виконавчого комітету
Славутицької
міської ради
Міський
методичний центр
Походження
мутацій
Виконав
Учень 11-Б
класу
Михайленко
Олександр
2013
Зміст
Вступ
1.
Теоретична
частина
1.1 Класифікація
мутацій
2.
Практична
частина
2.1 Експеримент1
2.2 Експеримент
2
3. Дослідження
Висновок
Список використаної літератури
Додатки
Вступ
Нині вивчення походження мутацій
дуже складне питання, так як на даний момент існує невелика частина
вивчених стійких змін у генотипі живих істот. З розвитком суспільства, зокрема
науки у цілому підвищується ризик до набуття певних вад і хвороб. Неможливо не
помітити, що імунітет людини ПОГІРШУЄТЬСЯ, стає вразливим до навколишнього
середовища. На противагу цьому розвиваються такі прикладні науки як медицина,
генетика й генна інженерія...Цей проект присвячений для загального ознайомлення
з властивостями, класифікації та певних статистичних даних відомих мутацій.
Мета роботи: ознайомитися з
загальнонауковим матеріалом щодо мутацій, їх впливу на живі організми.
Завдання науково-дослідницької
роботи: описати та систематизувати основні аспекти вивчення процесу мутації.
1. Теоретична
частина
У біології, мута́ції
- стійкі
зміни
генетичного матеріалу (звичайно ДНК або РНК). Мутації можуть бути викликані помилками
копіювання генетичного матеріалу протягом поділу клітини, опроміненням жорсткою
радіацією, хімічними речовинами (мутагенами), вірусами або можуть відбуватися
свідомо під клітинним контролем протягом таких процесів як, наприклад, мейоз
або гіпермутація. У багатоклітинних організмах мутації можуть бути підрозділені
на генеративні мутації, які можуть бути передані нащадкам, і соматичні мутації.
Соматичні мутації не можуть передаватися до нащадків у тварин. Рослини іноді
можуть передавати соматичні мутації своїм нащадкам безстатево або статево (у
випадку, коли брунька розвивається в соматично зміненій частині рослини).
Мутації були відкриті де Фрізом в
1900 р., спостерігаючи за мінливістю енотери (Oenothera)(рис1).
.1 Класифікація
мутацій
За ефектом на структуру
Послідовність ДНК гена може бути
змінена безліччю шляхів. Генетичні мутації мають різні ефекти на здоров'я
залежно від того, де вони відбуваються і чи змінюють вони функцію важливих
білків. Структурно, мутації можуть бути класифіковані, як:
· Невеликі мутації,
що охоплюють один або декілька нуклеотидів, зокрема:
o Точкові
мутації,(рис.2) часто викликані хімічними речовинами або помилками при
реплікації ДНК, представляють собою заміну одного нуклеотиду іншим. Найбільше
загальні - заміна пурину на пурин (A↔G) або піримідіну на піримідін (C↔T).
Така заміна може бути викликана азотистою кислотою, помилкою спарювання основ
або мутагенними аналогами основ, наприклад, 5-бромо-2-дезоксиуридином (BrdU).
Менш загальний випадок - трансверсія, або заміна пурину на піримідін або
піримідіну на пурин (C/T↔A/G). Точкова мутація може бути нейтралізована
іншою точковою мутацією, в якій нуклеотид змінюється назад до свого
оригінального стану (дійсна реверсія) або додатковою мутацією де-небудь у
іншому місці, яка приводить до відновлення функціональності гена (додаткова
реверсія). Такі зміни класифікуються як переходи або трансверсії. Приклад
трансверсії - аденін, що перетворюється на цитозин. Точкові мутації, які
відбуваються в межах області гена що кодує білки, можуть бути класифіковані на
три види, залежно від того, для чого використовуються помилкові кодони:
§ Безмовні мутації:
які кодують ту ж саму амінокислоту.
§ Міссенс-мутації:
які кодують іншу амінокислоту.
§ Нонсенс-мутації:
які кодують код зупинки (стоп-кодон) трансляції білка.
o Вставки
додають один або більше нуклеотидів до ДНК. Вони звичайно викликані мобільними
генетичними елементами, або помилками протягом копіювання елементів, що
повторюються (наприклад AT повторення). Вставки в кодуючі області гена можуть
змінювати сплайсинг мРНК або викликати "зміщення рамки" (англ.
frameshift), обидва типи мутацій можуть значно змінити продукт гена. Вставки
можуть бути звернені делецією мобільного генетичного елементу.
o Делеції
видаляють один або більше нуклеотидів із ДНК. Подібно до вставок, ці мутації
можуть викликати "зміщення рамки" гена. Вони незворотні.
Синдактилія(рис.4)
· Великі мутації в
хромосомній структурі, зокрема:
o Ампліфікації
(або дублювання гена) приводять до створення багатьох копій хромосомних областей,
збільшуючи дозування генів, розміщених в їхніх межах.
o Мутації, чий
ефект - зіставити разом окремі шматки ДНК, що може привести до створення
гібридних генів з новою функціональністю (наприклад
bcr-abl)(рис.5)Полідактилія. Вони включають:
§ Хромосомні
транслокації: обмін частинами генів між негомологічними хромосомами.
§ Інтерстиціальні
(проміжниі) делеції: видалення областей ДНК з єдиної хромосоми, таким чином
сполучаючи наперед віддалені гени.
§ Хромосомні
інверсії: зміна орієнтації хромосомного сегменту.
o Втрата
гетерозиготності: втрата одного алелю, шляхом делеції або рекомбінації, в
організмах які перед тим мали два.
За ефектом на функції
Мутації втрати функції приводять до
виробу гена, що має зменшену або немає функції. Коли алель має повну втрату
функції (нульовий алель), така мутація часто називається аморфною мутацією.
Фенотипи, пов'язані з такими мутаціями, частіше всього рецесивні, але є винятки
- коли організм гаплоїдний або коли зменшене дозування продукту гена мало для
підтримання нормального фенотипу (гаплонедостатність рис.6).
· Мутації отримання
функції замінюють продукт гена таким чином що він набуває нової анормальної
функції. Такі мутації звичайно мають домінантний фенотип.
· Домінантні
негативні мутації (також відомі як антиморфні мутації) мають змінений продукт
гену, який діє антагонічно до алелю дикого типу. Такі мутації звичайно
приводять до зміненої молекулярної функції (часто недіючої) і характеризуються
домінантним або напів-домінантним фенотипом. Синдром Марфана(рис.7) у людини -
приклад домінантної негативної мутації, яка проявляється як аутосомна
домінантна хвороба. За цими умовами, дефектний глікопротеїн що кодується
зміненим геном фібрілину (FBN1) протидіє продукту алелю.
· Смертельні мутації
- мутації, які приводять до фенотипу, нездібному до ефективного відтворення.
За аспектом зміненого фенотипу
Морфологічні мутації звичайно
впливають на зовнішність індивідуума. Мутації можуть змінити висоту рослини або
змінити вигляд її насіння від гладкого до грубого.
· Біохімічні мутації
приводять до пошкоджень, що зупиняють ферментний шлях. Часто, морфологічні
мутанти - прямий результат мутації завдяки ферментному шляху Триметиломінурія.
Спеціальні категорії
Умовна мутація - мутація, яка має
фенотип дикого типу за певними природними умовами і мутантом фенотип за певними
умовами. Умовні мутації також можуть бути смертельними.
Причини мутацій: Два класи мутацій -
спонтанні мутації (молекулярний розпад) і вимушені мутації, викликані
мутагенами.
Спонтанні мутації на молекулярному
рівні включають:
· Таутомерізм - В
основі замінюється розташування водневого атома.
· Депуринація -
Втрата пурину (А або G).
· Деамінація - Заміна
нормальної основи на нетипову; C → U, (може бути виправлений механізмами
ремонту ДНК), або спонтанна деамінація 5-метилцитозину (непоправна), або A →
HX (гіпоксантин).
· Транзиція - зміни
пурину на інший пурин, або пірімідіну на пірімідін.
· Трансверсія - пурин
стає пірімідіном або навпаки.
Вимушені мутації на молекулярному
рівні можуть бути викликані:
· Хімічними речовинами
o Нітрозогуанадін
(Nitrosoguanidine, NTG)
o Аналоги
основ (наприклад BrdU)
o Прості
хімічні речовини (наприклад кислоти)
o Алкілуючі
агенти (наприклад, N-етил-N-нітрозосечовина, англ. N-ethyl-N-nitrosourea, ENU).
Ці агенти можуть діяти на ДНК як при реплікації, так і у інший час. На відміну,
аналог основи може тільки видозмінити ДНК якщо він включається при реплікації
ДНК. Кожний з цих класів хімічних мутагенів має певні ефекти, які приводять до
транзицій, трансверсій або делецій.
o Метилируючі
агенти (наприклад етан-метил-сульфонат, англ. ethane methyl sulfonate, EMS).
o Поліциклічні
вуглеводні (наприклад бензопірен(рис.), знайдений у вихлопах двигунів
внутрішнього згорання)
o Інтеркаляційні
агенти (наприклад, бромистий етідій, англ. ethidium bromide)
o Крослінкери
або агенти поперечного зшивання ДНК (наприклад, платина)
· Радіація або
випромінювання
o Ультрафіолетове
випромінювання (неіонізуюче випромінювання) - збуджує електрони до вищого рівня
енергії та визиває хімічні реакції інакше неможливі. Дві нуклеотидні основи ДНК
- цитозин та тимін - найуразливіші до збудження, яке може замінити властивості
спаровування цих основ.
o Іонізуюча
радіація
У ДНК існують так звані гарячі
точки, де мутації відбуваються до 100 разів частіше, ніж у решті ДНК. Гаряча
точка може знаходитися в незвичайній основі, наприклад, 5-метилцитозині.
Частота мутацій також відрізняється
для різних видів. Еволюційні біологи запропонували, що вищі частоти мутації
вигідні в деяких ситуаціях, тому що вони дозволяють організмам еволюціонувати і
тому пристосувати швидше до своїх навколишніх середовищ. Наприклад,
багатократна дія антибіотиків на бактерій і відбір стійких мутантів може
привести до відбору бактерій, які мають вищі частоти мутацій, ніж оригінальна
популяція (мутаторна лінія).
Мутації і хвороби
Зміни в ДНК, викликані мутацією,
можуть викликати помилки в послідовності білка, створюючи частково або цілком
нефункціональні білки. Щоб функціонувати правильно, кожна клітина залежить від
тисяч білків, що повинні функціонувати в правильному місці в правильний час.
Коли мутація змінює білок, який грає критичну роль в тілі, може виникнути
захворювання. Захворювання, викликане мутаціями в одному або більше генах
називається генетичним захворюванням. Проте, тільки маленький відсоток мутацій
визивають генетичні захворювання, більшість не мають ніякого впливу на
здоров'я. Наприклад, деякі мутації змінюють послідовність основ ДНК гена, але
не замінюють функцію білка, зробленого геном.
Якщо мутація присутня в зародковій
клітині, вона може дати початок нащадку, який буде носити мутацію у всіх своїх
клітинах. Це причина спадкових хвороб. З другого боку, мутація може відбуватися
в соматичній клітині організму. Такі мутації будуть присутні у всіх нащадках
цієї клітини, і певні мутації можуть примусити осередок почати безконтрольно
відтворюватися, і тому викликати рак.
Часто, мутації гена, які могли б
створити генетичну хворобу, виправляються ремонтною системою клітини. Кожна
клітина має цілий ряд шляхів, через які ферменти визнають і ремонтують помилки
в ДНК. Оскільки ДНК може бути пошкоджена або видозмінюються у багатьох
відношеннях, процес ремонту ДНК - важливий шлях, в якому тіло захищає себе від
хвороби.
Дуже маленький відсоток всіх мутацій
фактично має позитивний ефект. Ці мутації приводять до нових версій білків, які
допомагають організму і його майбутнім поколінням краще пристосовують до змін в
їхньому навколишньому середовищі. Наприклад, вигідна мутація могла б приводити
до білка, який захищає організм від нової лінії бактерій.
Мутагенез
Мутагенез (штучне створення мутацій)
може використовуватися як ефективний генетичний інструмент. Стимулюванням
мутацій певним шляхом а потім спостереженням фенотипу організму можуть бути
визначені функції генів і навіть індивідуальних нуклеотидів.
Мутації розглядаються як рушійна
сила еволюції, де менш сприятливі (або шкідливі) мутації видаляються з
генофонду природним відбором, тоді як сприятливі (вигідні) прагнуть
накопичуватися. Нейтральні мутації визначаються як мутації, чиї ефекти не
впливають на виживання видів або індивідуумів, які складають види. Вони також
можуть накопичуватися. Переважна більшість мутацій не мають ніякого ефекту,
тому що механізми репарації ДНК (ремонту ДНК) можуть виправити більшість змін
перед тим, як вони стануть постійними мутаціями, і багато організмів мають
механізми для усунення інакше постійно видозмінених соматичних клітин.
2. Практична
частина
.1 Експеримент
1
У ході унікального експерименту, що
тривав більше 20 років, вдалося детально простежити еволюційні зміни, що
відбувалися в популяції кишкової палички Escherichia
coli
протягом 40000 поколінь. У першій половині експерименту в
популяції фіксувалися в основному корисні мутації, що підвищували
пристосованість бактерій. Найнесподіванішим результатом виявилося сталість
швидкості накопичення корисних мутацій. До цих пір вважалося, що з постійною
швидкістю повинні накопичуватися нейтральні мутації, а не корисні, проте в
експерименті все виявилося навпаки. У середині експерименту в популяції
зафіксувалася мутація, різко підвищила темп мутагенезу. Після цього мутації
стали фіксуватися на порядок швидше, але це були в основному вже не корисні, а
нейтральні мутації.
Давня мрія біологів - безпосередньо зіставити
темпи еволюції на геномному і організмовому рівнях - нарешті починає збуватися.
До цих пір в цій області доводилося задовольнятися в основному теоретичними
міркуваннями і математичними моделями. Унікальний експеримент, розпочатий в
1988 році міжнародною командою дослідників під керівництвом Річарда Ленскі
(Richard E. Lenski)(рис.9), дав можливість з небувалою досі ступенем
детальності простежити хід еволюції як на рівні генома (темп накопичення
мутацій), так і на рівні цілого організму (темп розвитку адаптацій).
Експеримент проводиться паралельно з 12
популяціями E. coli, але в обговорюваній статті розглянута тільки одна з них.
Бактерій вирощують на мінімальному живильному середовищі (див. : minimal growth
medium), в якій лімітуючим фактором, що обмежує розмноження бактерій, є нестача
їжі (глюкози). Кожен день з пробірки з мікробами беруть 0,1 мл вмісту і
поміщають в нову пробірку з 9,9 мл свіжої живильного середовища. Періодично
частина популяції заморожують при -80 ° C і зберігають для подальшого вивчення.
Це мудро, бо аналітичні методики - зокрема, методики секвенування ("
прочитання ") геномів - зараз стрімко розвиваються і так само стрімко
дешевшають. На сьогоднішній день автори повністю отсеквенували геноми бактерій
з поколінь № 0 (предковий штам), 2000, 5000, 10 000, 15 000, 20 000 і 40 000.
Регулярно проводиться також оцінка "
пристосованості " популяції. Для цього порівнюють швидкість розмноження
" експериментальних " мікробів з контрольним міченим штамом (про
методику оцінки пристосованості см. у замітці : Досліди на черв'яках довели, що
самці - річ корисна, "Елементи", 23.10.2009).
Тривалість експерименту і розмір популяції були
достатніми для того, щоб кожна з можливих точкових мутацій (нуклеотидних замін)
в ході випадкового мутування сталася більше одного разу (розмір генома
піддослідного штаму кишкової палички - 4,6 ×
106 пар нуклеотидів).
Чим відрізняється "виникнення "
мутації від її " закріплення " (фіксації). Далеко не всяка виникла
мутація закріплюється (фіксується) в популяції. Кожна мутація спочатку виникає
тільки у одного мікроба. Щоб мутація зафіксувалася, нащадки цього мікроба
повинні витіснити всіх інших мікробів у своїй пробірці. Шкідлива мутація,
швидше за все, буде відсіяна відбором. Корисна мутація під дією відбору може
закріпитися, тобто поширитися в популяції і досягти стовідсоткової частоти, але
може і випадково загубитися, поки її носії ще не встигли як слід розмножитися.
Нарешті, нейтральні мутації, згідно з наявними теоретичними моделями, повинні
фіксуватися з постійною швидкістю, рівної швидкості мутування. Частота
народження нейтральної мутації в популяції коливається відповідно до алгоритму
випадкового блукання до тих пір, поки випадково не досягне величини 0 % (повна
елімінація) або 100 % (фіксація). Парадоксальним чином швидкість фіксації нейтральних
мутацій не залежить від розміру популяції. Це пояснюється тим, що чим більше
популяція, тим частіше в ній виникають мутації, а й тим нижча ймовірність
фіксації кожної з них. В результаті розмір популяції в рівнянні просто -
напросто скорочується.
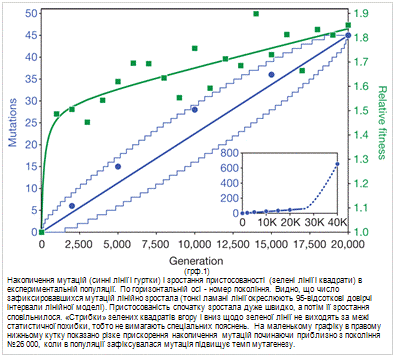
За перші 20 000 поколінь в експериментальній
популяції зафіксувалося 45 мутацій, у тому числі 29 однонуклеотидних замін і 16
інших мутацій (вставок, випадінь, інверсій, вбудовування мобільних елементів).
Найцікавіше, що швидкість накопичення мутацій на цьому етапі була строго
постійною (всі відхилення від лінійної моделі статистично недостовірні), тоді
як пристосованість спочатку росла дуже швидко, а потім її зростання
сповільнилося (грф.1).
Постійна швидкість фіксації, відповідно до
теорії, характерна для нейтральних мутацій. Однак всі 45 мутацій не могли бути
нейтральними. Ясно, що принаймні деякі з них були корисними - про це свідчить
зростання пристосованості. Отримані результати не дуже легко пов'язати і з
гіпотезою про те, що всі 45 мутацій були корисними.
Просте з можливих пояснень полягає в тому, що
серед 45 зафіксувавши мутацій більшість були нейтральними, але деякі були
корисними, причому основна маса корисних мутацій зафіксувалася незабаром після
початку експерименту. Можливості для "корисного мутування " досить
швидко вичерпалися, і надалі фіксувалися переважно нейтральні мутації.
Накопичення мутацій (сініі лінії та гуртки) і
зростання пристосованості (зелені лінії і квадрати) в експериментальній
популяції. По горизонтальній осі - номер покоління. Видно, що кількість
зафіксованих мутацій зростало лінійно (тонкі ламані лінії окреслюють 95- відсоткові
довірчі інтервали лінійної моделі). Пристосованість спочатку росла дуже швидко,
а потім її зростання сповільнилося. " Стрибки " зелених квадратів
вгору і вниз щодо зеленої лінії не виходять за межі статистичної похибки, тобто
не вимагають спеціальних пояснень. На маленькому графіку в правому нижньому
кутку показано різке прискорення накопичення мутацій починаючи приблизно з
покоління № 26000, коли в популяції зафіксувалася мутація, що підвищує темп
мутагенезу.
Автори, проте, призводять чотири серйозні
аргументи проти такого пояснення.
) У разі переважання нейтральних мутацій має
бути різко підвищено число синонімічних нуклеотидних замін (тобто таких змін
ДНК, які не ведуть до змін амінокислотної послідовності білків). Всупереч цим
очікуванням, всі без винятку зафіксованого мутації в кодують областях генів є
значущими (несинонимичної).
) У разі переважання нейтральних мутацій слід
очікувати, що в 12 експериментальних популяціях (у 11 з яких повні геноми поки
не секвеновані) за 20 000 поколінь мутації зафіксувалися в різних генах.
Навпаки, мутації в одних і тих же генах, що закріпилися незалежно в різних
популяціях, були б доказом на користь того, що мутації фіксувалися в результаті
відбору, а не генетичного дрейфу (тобто мутації були корисними). Щоб перевірити
це, автори отсеквенували у бактерій покоління № 20 000 з інших одинадцяти
експериментальних популяцій 14 генів, в яких у "головною" експериментальної
популяції закріпилися мутації. Виявилося, що в переважній більшості випадків в
інших популяціях ці гени теж змінилися.
) Якщо б більшість мутацій були нейтральними,
спостерігалася б значна Внутрішньопопуляційна мінливість за цими локусами (бо
корисні мутації під дією відбору фіксуються швидко, а нейтральні спочатку
повинні довго "випадково блукати " між нульовою і стовідсоткової
частотою). Це припущення не підтвердилося.
) За допомогою генної інженерії автори змогли
безпосередньо визначити ступінь корисності 9 мутацій з розглянутих 45. Ці
мутації штучно впроваджували в геном предкового штаму. У восьми випадках з
дев'яти пристосованість бактерій різко підвищилася. Що стосується дев'ятої
мутації, то автори думають, що вона теж корисна, але не сама по собі, а в поєднанні
з іншими мутаціями, бо точно така ж мутація закріпилася у інших піддослідних
популяцій. Для порівняння, в іншому експерименті в геном кишкової палички
вносили випадкові мутації по одній, і при цьому жодна з 26 мутацій не дала ні
найменшого виграшу в пристосованості.
Таким чином, протягом перших 20 000 поколінь в
популяції фіксувалися переважно корисні мутації, причому їх фіксація йшла з
постійною швидкістю. Уповільнення зростання пристосованості, мабуть, було
пов'язано з тим, що середня ступінь корисності мутацій поступово знижувалася.
Найбільш радикальні адаптивні зміни відбулися протягом перших 2000 поколінь, а
потім, ймовірно, відбувалася більш тонка оптимізація фенотипу.
До цих пір мова йшла тільки про першу половину
експерименту. У другій його половині еволюційна динаміка піддослідної популяції
радикально змінилася. Справа в тому, що після 26 000 поколінь зафіксувалася
мутація в гені mutT. Цей ген кодує білок, що бере участь у репарації ("
лагодження ") ДНК. В результаті частота мутування різко зросла. Як
наслідок, більш ніж на порядок зросла і частота фіксації мутацій. Протягом
другої половини експерименту зафіксувалося 609 мутацій - в 13,5 разів більше,
ніж за перші 20 000 поколінь.
Аналогічні за своєю функцією мутації, що
збільшили темп мутагенезу, закріпилися і в кількох інших експериментальних
популяціях. З цього випливає, що зростання темпів мутагенезу дав бактеріям
адаптивне перевагу. Це, між іншим, суперечить поширеною ідеї про те, що в
стабільних умовах організмам було б вигідно знизити темп мутування до нуля - і
цього не відбувається тільки через технічну неможливість забезпечити абсолютну
точність копіювання ДНК.
У " основний" популяції ця мутація
з'явилася не пізніше, ніж в поколінні № 26 500 (з трьох перевірених мікробів
цього покоління мутація є в одного і відсутній у двох). Починаючи з покоління №
29000 мутація стала переважати в популяції і, мабуть, незабаром після цього
зафіксувалася, тобто досягла частоти 100%.
" Корисність " мутації, що підвищила
темп мутагенезу, могла полягати лише в тому, що вона підвищила ймовірність
виникнення нових корисних мутацій після того, як більшість можливостей для
"корисного мутування " вже було вичерпано. Але в якості побічного
ефекту інтенсифікація мутагенезу неминуче повинна була привести до зростання числа
шкідливих і нейтральних мутацій.
Виходячи зі сказаного вище, слід було чекати, що
тепер більшість фиксирующихся мутацій будуть нейтральними, а не корисними. Як
ми пам'ятаємо, швидкість фіксації нейтральних мутацій в популяції дорівнює
швидкості мутагенезу незалежно від розміру популяції. Швидкість мутування у
предкового штаму E. coli була низькою, і тому в перші 20 000 поколінь
нейтральних мутацій фіксувалося дуже мало. Мутація гена mutT збільшила
швидкість мутування приблизно в 70 разів (від 1,6 ×
10-10 до 1,1 × 10-8 на пару
нуклеотидів на покоління). Різноманітні статистичні тести, застосовані
авторами, підтвердили припущення про те, що більшість з 609 " пізніх
" мутацій були нейтральними.
Чомусь у статті не сказано, як змінилася
пристосованість бактерій у другій половині експерименту.
Біологи активно використовують результати
порівнянь геномів для реконструкції шляхів і темпів еволюції організмів. Досі
при цьому доводилося задовольнятися в основному математичними моделями,
заснованими на різних більш-менш правдоподібних припущеннях. Тепер нарешті
з'являється можливість експериментальної перевірки цих моделей. Перші
результати виявилися багато в чому несподіваними. Наприклад, мало хто очікував,
що постійний темп накопичення корисних мутацій може супроводжуватися
замедляющимся зростанням пристосованості ; або що можливі такі різкі скачки в
співвідношенні темпів фіксації нейтральних і корисних мутацій. Основний
висновок полягає в тому, що кількісні співвідношення між різними аспектами
еволюційного процесу (нейтральністю і адаптивністю, впливом дрейфу і відбору,
темпами змін на рівні генотипу і фенотипу) можуть бути істотно більш складними,
неоднозначними і мінливими, ніж передбачалося раніше.
Тобто можна зробити висновок. Що багато чого
замовчується. Неможливо знехтувати припущенням, що достовірна повна версія
висновків навмисно приховується. Неможливо визначити намір. Але результати
досліджень, можуть стати витратним матеріалом, для створення нової біологічної
зброї.
2.2 Експеримент
2
Американські біологи встановили, що
вірусам для повної мутації потрібно не 100 років, а пару тижнів
Учені з'ясували, що сучасні віруси
здатні на надшвидкі мутації. Раніше вважалося, що нові стратегії боротьби з
вірусами стають актуальні лише через приблизно 100 років з моменту розробки
вакцини. Вважалося, що саме після такого періоду часу вірус мутує так сильно,
що старий варіант вакцини на нього вже не діє.
Проте в останньому номері наукового
журналу Science група вірусологів з Державного університету Мічігану
опублікувала нові дані про еволюцію вірусів, в яких мовиться, що вірусам для
повної мутації може бути досить двох тижнів, щоб інфікувати клітку абсолютно
іншим шляхом, повністю змінивши механізм біологічної дії, передає
CyberSecurity.ru.
У статті мовиться, що раніше схожим
способом діяв гучний вірус H5n1, якому вдавалося швидко змінювати свій
біологічний код, переміщаючись від одного ссавця до іншого. Кілька років тому,
коли біологи заявили, що H5n1 здатний на надшвидкі мутації, основне наукове
співтовариство заявило, що процес мутації вірусів займає роки, а не тижні або
місяці. Тепер же американські учені на практиці підтвердили, що віруси можуть
мутувати в лічені тижні.
Дослідники Мічігану говорять, що
вони вивчили вірус, відомий як лямбда(рис. Він не є небезпечним для людей,
оскільки здатний інфікувати тільки бактерії Escherichia coli, проте біологи
зацікавилися низкою особливостей вірусу, що дозволяють йому швидко
еволюціонувати, щоб весь час заражати організм господаря новим шляхом.
Зазвичай лямбда проникає всередину клітини
традиційним способом - через клітинну мембрану, прикріпляючись до певних
з'єднань усередині клітини, крім того, він здатний упроваджувати свій код в
організм бактерії.
Разом з тим, учені провели
експеримент, в якому Escherichia coli були позбавлені потрібних вірусу
з'єднань, проте, на подив експертів, всього через 15 днів лямбда підібрав
"ключі" і до модифікованої бактерії, прикріпляючись до нових
молекулярних з'єднань і успішно інфікуючи бактерію за новим алгоритмом.
У статті в журналі Science учені
говорять, що вони не чекали такої швидкої здібності вірусу до мутацій, причому
учені проводили аналогічні дослідження в 24 ізольованих біологічних середовищах
і у всіх віруси підібрали ключі до інфікування бактерій.
Пізніше біологи досліджували геном
лямбди і дійшли до висновку, що новий механізм інфікування зажадав зміни всього
в чотирьох генах, що відповідали за розміщення вірусу в іншому організмі.
У статті учені кажуть, що лямбда є
одним з простих вірусів, тоді як є куди складніші зразки, здатні до набагато
складніших мутацій.
Наприклад, було встановлено, що
вищезазначений H5n1 був здатний до швидкої мутації, але діяв по-різному на
різні біологічні види, що робить його універсальним вірусом, здібним до швидкої
і дуже небезпечної дії на організми.
Відзначимо, що кілька років тому
було встановлене, що віруси здатні заражати один одного.
Всіма відомими на сьогодні вірусами
є мікроскопічні частинки, здатні інфікувати клітки живих організмів.
Віруси є облігатними паразитами,
тобто вони не здатні розмножуватися поза клітиною. В даний час відомі віруси,
що розмножуються в клітках рослин, тварин, грибів, бактерій та інших вірусів.
Віруси містять тільки один тип
нуклеїнової кислоти: або ДНК, або РНК.
3. Дослідження
Якщо зробити графік життєдіяльності
людини, то він буде мати форму вітки гіперболи, причому спадної. Це є лише
підтвердженням того, що стійкість імунітету людини до будь-яких хвороб спадає
із плином часу та розвитком індустрії. Алергія
- яскравий представник. З розвитком науки і суспільства, людство створило
безліч факторів, що вплиають на погіршення генофонду націй.
Людина
декілька століть назад відрізнятиметься від сучасної станом здоров'я. Алергії
не було у минулому. Вона з’явилась відносно недавно.
В
останні десятиліття відзначений виражене зростання захворюваності алергією.
Існують різні теорії пояснючі цей феномен:
Теорія
впливу гігієни ("hygiene hypothesis"), висунута в 1989 David P.
Strachan - стверджує, що перехід до дотримання норм гігієни запобігає контакту
організму з багатьма антигенами, що викликає недостатню завантаження імунної
системи (особливо у дітей). Оскільки наше тіло сконструйовано так, що воно
повинно постійно протистояти певному рівню загроз, імунна система починає
реагувати на нешкідливі антигени. [1 ] Теорія впливу гігієни була розроблена
для пояснення причин, за якими діти з англійських багатодітних сімей набагато
рідше страждали такими видами алергії, як алергічний риніт або екзема, ніж
єдині діти в сім'ї. Зростаюче
споживання продуктів хімічної промисловості. Багато хімічні продукти можуть
виступати як у ролі алергенів, так і створювати передумови для розвитку
алергічних реакції допомогою порушення функції нервової та ендокринної системи.
Однак,
незважаючи на численні спроби пояснити різке зростання захворювання алергією
впливом техногенного середовища, до цих пір не було дано пояснення, чому одні й
ті ж фактори на одних людей чинять такий вплив, а на інших - ні. Ніякого
взаємозв'язку захворювання алергією із загальним станом здоров'я також не було
виявлено. За
прогнозами вчених генетиків та алергологів, вже у 21-му столітті через 20-50
років матимемо 100% вроджену алергію. Отримаємо
новий вид людини.
Висновок
Умови навколишнього середовища
можуть впливати на генотип, що як наслідок може проявитись у фенотипі. Це є
запорука еволюції. Еволюція не припиняється.
Природно створено такий закон:
"Будь-яка система має намір зменшити енергію".
Тому для комфорту організму властиве
несвідоме його удосконалення, що необхідне для життя за певних умов.
Людина, розвиваючись, втручаючись у
"справи природи", значно ризикує втратити свою живу сутність, й
перейти до наступного щаблю еволюції…
Посилаючись на вищезгадане у
науково-дослідницькому проекту зауважимо, що мутації є головним джерелом
еволюції. Завдання сучасної генної інженерії є цілковите дослідження фіксованих
мутацій. Викорінення "шкідливих", та запровадження
"корисних" мутацій.
Вчені-генетики встановили. що
чоловіча ХУ- хромосома постійно деградує. Це пов’язано з накопиченням
негативних чинників. Серед них зменшення вмісту кисню й підвищення вмісту
шкідливих речовин та пристрасті до спиртного. Лише 5 грам спирту можуть
призвести до зміни в здатності до розмноження. Жінки ризикують залишитись без
чоловіків
Саме запровадження
"корисних" мутацій та викорінення шкідливих звичок є запорукою
збереження людської раси від вимирання.
Список використаної літератури
мутація генетичний
пристосованість популяція
1) Способность к сложному
коллективному поведению может возникнуть благодаря единственной мутации,
"Элементы", 25.05.2006.
) Микробиологи утверждают:
многоклеточность - сплошное жульничество, "Элементы", 06.04.2007.
) Начальные этапы
видообразования воспроизведены в эксперименте на дрожжах, "Элементы",
06.06.2007.
) Амёбы-мутанты не позволяют
себя обманывать, "Элементы", 06.10.2009.
) Опыты на червях доказали, что
самцы - вещь полезная, "Элементы", 23.10.2009.
Додатки

(рис.1)Енотера
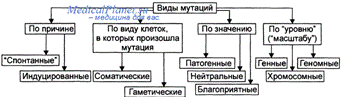
(табл.1)Класифікація мутацій

(рис.2) Результат точкової мутації -
прогерія
(рис.3)Кишкова паличка
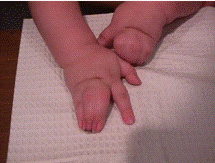
(рис.4)Синдактилія
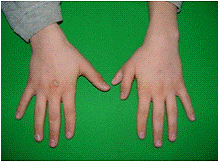
(рис.5)Полідактилія
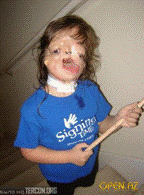
Гаплонедостатнісь Джуліани. Синдром
Колінза Критчера (рис.6)
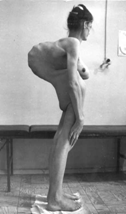
(рис.7)Синдром Морфана
(рис.8) Бензопірен

(рис.9) Річард Ленскі. Керівник
науково-дослідницького проекту вивчення мутацій бактерії -збудника кишкової
палички.
(Рис.10) Лямбда-вірус