Влияние восстановленного глутатиона и ингибитора каталазы на пероксидную резистентность и скорость лизиса эритроцитов при действии хлорида железа
Министерство
науки и образования Украины
Харьковский
национальный университет имени В.Н. Каразина
Биологический
факультет
Кафедра
биохимии
Дипломная
работа магистра на тему
Влияние
восстановленного глутатиона и ингибитора каталазы на пероксидную резистентность
и скорость лизиса эритроцитов при действии хлорида железа
Харьков
2010 г.
РЕФЕРАТ
Работа изложена на 39 страницах, содержит 6
таблиц и 7 рисунков, 42 литературных источника (из них 32 иностранных).
Объект исследования - эритроциты человека из
донорской эритромассы (ХОЦСК, кровь (мужская) группы А(II), Rh+, консервант -
глюгицир).
Ключевые слова: гемолиз, эритроциты, пероксидная
резистентность, кинетика, каталаза, азид натрия, хлорид железа (III) ,
глутатион.
Целью работы являлось изучение активности
каталазы, пероксидного гемолиза эритроцитов и кинетики гемолиза после обработки
эритроцитов хлоридом железа (III), азидом натрия, глутатионом.
Установлено, что при ингибировании каталазной
активности азидом натрия, в том числе при действии хлорида железа скорость
гемолиза эритроцитов возрастает. Хлорид железа (III) в концентрации 0,5%
вызывал полный лизис эритроцитов человека за 5 мин инкубации с максимумом
лизиса от 1,5 до 3,5 минут инкубации вне зависимости от предварительной обработки
эритроцитов. Предварительная обработка азидом натрия увеличивала скорость
гемолиза в присутствии 0,5% FeCl3. Последовательная предварительная обработка
эритроцитов азидом натрия или глутатионом и далее 0,05% FeCl3 вызывала снижение
скорости лизиса эритроцитов хлоридом железа.
СПИСОК ОБОЗНАЧЕНИЙ
- восстановленный глутатион- хлорид железа (III)
СОД- супероксиддисмутаза- азид натрия
ВВЕДЕНИЕ
В настоящее время установлено, что
внутрисосудистый лизис эритроцитов при патологиях и действии стрессорных
факторов вызывает развитие сосудистых повреждений за счет накопления в крови
гем- и железо-содержащих продуктов гемолиза. Высвободившиеся из гема ионы Fe3+
увеличивают выработку активных форм кислорода, которые способны индуцировать
адгезию клеток крови к стенкам сосудов и вызывать окисление липопротеинов
низкой плотности. Из литературных данных известно, что содержание железа в
окисленной форме коррелирует с риском сердечнососудистых заболеваний, что
обуславливает постоянный научный интерес к изучению факторов, отвечающих за
устойчивость эритроцитов к гемолизу. Особую актуальность приобретает изучение
пероксидной резистентности клеток крови, т.к. повышенное содержание перекиси
водорода в кровяном русле отмечается при различных патологических состояниях, в
том числе инфекциях и воспалении.
Учитывая прооксидантный характер действия ионов
железа на мембраны клеток, можно предположить ключевую роль антиоксидантных
систем в защите эритроцитов от лизиса. Вместе с тем, механизмы гемолитического
действия соединений железа, а также роль каталазы и восстановленного
глутатиона, как ключевых компонентов антиоксидантной защиты, в поддержании
пероксидной резистентности эритроцитов остаются малоизученными. В связи с
вышесказанным, целью настоящей работы являлось изучение влияния хлорида железа
(ІІІ) на пероксидную резистентность и скорость лизиса эритроцитов человека в
условиях ингибирования каталазной активности и повышения содержания
восстановленного глутатиона в опытах in vitro.
Глава 1. Обзор
литературы
.1 МЕХАНИЗМЫ ПЕРОКСИДНОГО ГЕМОЛИЗА
Перекись водорода - одна из активных форм
кислорода, которая способна вызывать гемолиз эритроцитов. При этом лизис клеток
может осуществляться как при действии экзогенной перекиси водорода, так и
эндогенной, выработавшейся в клетке в избыточных количествах. Генерация
перекиси водорода может наблюдаться при различных процессах, таких как действие
гормонов, гипероксия, инфекционные заболевания. Основными клеточными мишенями
являются липиды и белки мембраны эритроцита, а также внутриклеточный белковый
компонент.
1.1.1 Влияние перекиси
водорода на липидный компонент мембраны эритроцита
Одной из причин гемолиза эритроцитов при влиянии
перекиси водорода является перекисное окисление липидов мембраны.
Непосредственными эндогенными предшественниками гидроксильного радикала,
инициирующего цепное окисление липидов, служат ионы двухвалентного железа
гемоглобина и перекись водорода (реакция Фентона)[1]. При недостатке витамина Е
и внесении Н2О2 наблюдается перекисное окисление фосфолипидов мембраны и последующий
гемолиз по коллоидно-осмотическому типу. Наиболее чувствительной к окислению
фракцией липидов в данном случае является фосфатидилэтаноламин,
Перекись водорода служит инициатором перекисного
процесса, в результате ее превращений образуются свободные радикалы, которые
инициируют перекисное окисление липидов по типу цепной реакции [2]. Под
влиянием эндогенной перекиси водорода в результате аутокаталитических реакций
также образуются перекиси и гидроперекиси липидов.
1.1.2 Влияние перекиси
водорода на белковый компонент мембран
Гемолиз эритроцитов под влиянием перекиси
водорода может быть следствием не только повреждения липидного компонента
мембран. Окисление сульфгидрильных групп белков мембраны может также приводить
к лизису красных кровяных клеток.[3] , [4]
При этом эндогенная перекись, повреждающая
белковые структуры может образовываться в результате действия на эритроцит
органических перекисей Повреждение сульфгидрильных групп гемоглобина при атаке
гидроксильного радикала может лежать в основе денатурации гемоглобина. В
дальнейшем, потерявший нативную структуру гемоглобин может связываться со
спектриновой сетью, обуславливая ее повреждение и последующий гемолиз [5],[6].
При этом пероксид водорода может быть результатом окислительно-восстановительных
превращений фенилгидразина. Фенилгидразин является прооксидантом,
метаболизируется в организме с образованием фенилдиазина, при этом генерируются
свободные радикалы, в том числе перекись водорода [7], [8]
Гемоглобин может действовать как ловушка
(scavenger) для пероксида водорода, при этом ион железа гемоглобина Fe (II)
окисляется до Fe (III) и даже до Fe (IV) с образованием соответственно таких
форм гемоглобина как метгемоглобин и феррил гемоглобин. Молекулы феррил
гемоглобина и радикалы гемоглобина могут полимеризоваться, что в последующем
ведет к гемолизу. При этом повышенное содержание перекиси водорода в кровяном
русле в физиологических условиях может наблюдаться при активности нейтрофилов в
воспалительных процессах. Под влиянием перекиси водорода могут образовываться
не только метгемоглобин и феррил гемоглобин, но и временно существующие
белковые радикалы гемоглобина и гем-белковые сшивки.
Также было показано, что при обработке
гемоглобина перекисью водорода этот белок становится более чувствительным к
действию протеиназ (трипсина, химотрипсина, эластазы и др.) [9]. Интенсивный
гемолиз наблюдается при обработке эритроцитов новорожденных перекисью водорода.
Это объясняется пониженной активностью метгемоглобинредуктазы, каталазы и
низким содержанием витамина Е в эритроцитах. [10]
Добавление экзогенной перекиси, либо
внутриклеточное ее образование в ответ на рецепторную стимуляцию влияет на
функции различных белков, включая протеинкиназы, протеинфосфатазы,
транскрипционные факторы, фосфолипазы, ионные каналы и G-белки. Т.к. перекись
водорода является небольшой, способной к диффузии молекулой, которая может
легко синтезироваться и разлагаться в ответ на внеклеточные стимулы, то она,
таким образом, соответствует всем требованиям, предъявляемым к внутриклеточным
вторичным посредникам при передаче сигнала. И на данный момент перекись
водорода рассматривается как вторичный посредник при токсических условиях. [11]
.2 АНТИОКСИДАНТНАЯ СИСТЕМА ЭРИТРОЦИТОВ
1.2.1 Антиоксидантные
ферменты
Одну из первых линий защиты клеток от
агрессивного действия свободных радикалов обеспечивают ферменты -
супероксид-дисмутаза (СОД), каталаза, глутатионзависимые пероксидазы и
трансферазы, удаляющие органические перекиси.
Супероксиддисмутаза
Различаясь по строению активного центра и структуре
полипептидной цепи, все СОД (металлоферменты) катализируют одну и ту же реакцию
дисмутации:
•ОО - +•ОО - + 2Н+ → О2 + НООН
Судьба перекиси водорода, образуемой в процессе
реакций дисмутации, различна. Частично перекись водорода разлагается при участии
каталазы, проявляющей активность почти во всех клетках организма человека,
особенно в печени, почках, эритроцитах:
Н2О2 → Н2О + О2
В присутствии ионов двухвалентного железа
перекись разлагается в реакции Фентона с образованием гидроксильного радикала
(OH•):
Н2О2 + Fe2+ → Fe3+ + OH- + OH•+ + H2O2 →
Fe2+ + OOH· + H+
Радикалы гидроксила чрезвычайно активны и
разрушают различные по структуре молекулы.
СОД существенно ускоряет дисмутацию супероксид
анион-радикала. Однако, несмотря на высокую специфичность фермента, при
определенных условиях Cu-Zn СОД может взаимодействовать с перекисью водорода и
выступать в качестве прооксиданта.
Исключительно важным моментом эффективности
ферментного звена антиоксидантной системы является сбалансированность
активности СОД, каталазы и пероксидазы. Подавление активности одного из
ферментов антиоксидантной системы может привести к избыточному накоплению
активных форм кислорода и деструкции клеток.
Каталаза
Каталаза - гемопротеин, локализуется в основном
в пероксисомах, частично - в микросомах и в меньшей мере - в цитозоле.
Разложение перекиси водорода каталазой
осуществляется в два этапа:
хлорид ингибирование железо каталазный
Fe3+-каталаза + 2 Н2О2→ окисленная
каталаза + Н2О2 → Fe3+-каталаза + Н2О2 + О2.
При этом в окисленном состоянии каталаза
работает как пероксидаза. Субстратами в пероксидазной реакции могут быть
этанол, метанол, формиат, формальдегид и другие доноры водорода.
Следует отметить, что около 0,5% кислорода,
образующегося в результате разложения перекиси водорода, возникает в
возбужденном, синглетном состоянии и таким образом в процессе разложения
перекиси водорода вновь генерируются активные формы кислорода.
Активности каталазы и СОД коррелируют между
собой, что может быть связано с переключением потока электронов с одной цепи
транспорта на другую. В этих условиях СОД и каталаза действуют как звенья одной
системы утилизации кислорода, размещенные в разных участках клетки.
Максимальная концентрация каталазы обнаружена в эритроцитах.
Глутатионзависимые антиоксидантные ферменты.
Важнейшей системой инактивации свободных
радикалов являются восстановленный глутатион и комплекс ферментов -
глутатионпероксидазы, глутатионтрансферазы и глутатионредуктазы. Глутатион
синтезируется в печени, откуда транспортируется в различные органы и ткани,
обеспечивает восстановление дисульфидных групп белков, дигидроаскорбиновой
кислоты, с участием глутатионтрансферазы образует конъюгаты в печени с
электрофильными соединениями и последующим выведением их с мочой.
Инактивация перекиси водорода в клетках
обеспечивается также глутатионпероксидазой (ГПО). Глутатионпероксидаза
катализирует реакцию восстановления гидроперекиси с помощью глутатиона,
обладает широкой субстратной специфичностью по отношению к гидроперекисям, но
абсолютно специфична к глутатиону.
В клетках млекопитающих выделяют семейство
мультифункциональных белков - глутатионтрансфераз, использующих глутатион для
конъюгации с гидрофобными соединениями и восстановление органических перекисей.
[9]. Трансферазы эффективно и специфично катализируют восстановление
белок-S-S-смешанных дисульфидов (protein-SSG). Глутатионтрансфераза способна
также реагировать с глутатионовыми радикалами (GS) и стабилизировать
дисульфидный анионный радикальный интермедиат (GRx-SSG.), т. о. способствуя превращению
GS в GSSG либо переносу GS с формированием protein-SSG. . [12]
Глюкозо-6-фосфат дегидрогеназа
Глюкозо-6-фосфатдегидрогеназа является ключевым
ферментом, который катализирует первые реакции в пентозомонофосфатном пути. В
результате работы фермента восстанавливается НАДФ+ до НАДФН., за счет которого
происходит восстановление оксисленного глутатиона. Снижение активности фермента
приводит к уменьшению НАДФН и восстановленного глутатиона
соответственно[13],[14].
Метгемоглобинредуктаза
При воздействии ксенобиотиков - окислителей
восстановление избыточных концентраций MtHb в Hb осуществляется в организме с
помощью специальных MtHb-восстанавливающих систем. Их главным компонентом
является никотинамидадениндинуклеотид-Н-цитохром b5-редуктаза (НАДН-Н-метгемоглобинредуктаза
или диафораза-1. Реакция восстановления MtHb двухступенчатая: энзиматическое
восстановление цитохрома b5 и неэнзиматическое восстановление MtHb.
Неэнзиматическое восстановление MtHb происходит
с участием аскорбиновой кислоты (12-16%) и глутатиона (9-12%). Другой
энзиматический путь превращения MtHb в Hb происходит с участием фермента
НАДНФ-Н-метгемоглобинредуктазы или диафоразы-2. Этот фермент физиологически
инертен, но может активироваться экзогенными акцепторами электронов, например,
метиленовым синим, рибофлавином. Диафораза-2 выполняет роль резервной
энзиматической системы.
У пациентов с наследственной недостаточностью
глюкозо-6-фосфатдегидрогеназы лекарственный гемолиз обычно сопровождается
выраженной метгемоглобинемией [15].
1.2.2.Эндогенные
низкомолекулярные антиоксиданты
Глутатион
Антиоксидантные свойства глутатиона определяются
как непосредственным взаимодействием с АФК и обменными реакциями с
дисульфидными связями, так и функционированием ряда ферментов глутатионового
цикла, из которых главная глутатионпероксидаза и глутатион -S - трансфераза.
Глутатион играет важную роль в антиоксидантной защите не только при
гипоксических, но и гипероксических состояниях, ограничивающих
свободнорадикальное окисление. Глутатион обеспечивает формирование
антиоксидантного потенциала в эритроцитах, кроветворных клетках, поддерживает
пул восстановительного аскорбата. [16]
Восстановленный глутатион реагирует с продуктами
супероксиданионной реакции, и, действуя в составе глутатион-зависимых ферментов
разлагает перекись водорода и перекиси липидов, а также ковалентно модифицирует
токсические ксенобиотики и эндогенные электрофилы с формированием
водорастворимых конъюгатов, которые экспортируются из эритроцита для экскреции.
При заболеваниях, связанных с возросшим образованием активных форм кислорода в
результате недостатка глутатиона, восстановление нормальной концентрации
глутатиона имеет положительный терапевтический эффект. [17]
Липоевая и дигидролипоевая кислоты
Липоевая и ее восстановленная форма - дигидролипоевая
кислоты являются также компонентами антиоксидантной защиты клеток. [18],
Подобно другим низкомолекулярным тиоловым соединениям клеток (GSH,
тиоредоксину, эрготионеину) ЛК осуществляет антиоксидантную защиту от
окислительных стрессов разной природы вместе со своей восстановленной,
дитиольной формой ДЛК. [19]
ЛК выступает как синергист витаминов А, С, Е,
частично заменяя их при их недостаточности. ЛК стимулирует синтез GSH de novo,
в частности, путем оптимизации поступления и усвоения его предшественника
цистеина. Липоевая и дигидролипоевая кислоты - перехватчики активных форм
кислорода. [20]
ЛК растворима как в воде, так и в липидах, что
делает ее эффективным антиоксидантом и в жидкостях организма, и в мембранных
структурах. ЛК внутриклеточно восстанавливается в ДЛК, которая экспортируется
во внеклеточную среду, так что антиоксидантная система ЛК-ДЛК функционирует как
внутри, так и вне клетки. [21]
.3 ОКИСЛИТЕЛЬНЫЙ ГЕМОЛИЗ ПРИ ПАТОЛОГИЧЕСКИХ
СОСТОЯНИЯХ И СТРЕССОВЫХ ВОЗДЕЙСТВИЯХ.
Циркулирующий в эритроцитах кровяного русла
гемоглобин способен окисляться до метгемоглобина. Окисление ферро формы
гемоглобина (Fe(2+)) до метгемоглобина (ферри) (Fe(3+)) ингибируется самой
структурой белка, однако, это все-таки происходит. В отличие от ферро формы
гемоглобина, метгемоглобин легко высвобождает гем. [25].
При влиянии перекиси водорода происходит
расщепление порфиринового кольца и выход из кольца «свободного» окислительно-активного
железа. При этом Fe(3+) способен каталитически увеличивать выработку активных
форм кислорода. Эти активные формы кислорода могут окислять липиды, белки, ДНК;
активировать сигнальные пути в клетках и чувствительные к окислителям
провоспалительные транскрипционные факторы. Также АФК могут изменять экспрессию
белков, дестабилизировать каналы в мембране, индуцировать апоптоз клеток.
Полученные в результате окисления гема АФК индуцируют мобилизацию лейкоцитов,
тромбоцитов и эритроцитов и их адгезию к стенкам сосудов и т.д. [26].Как гем,
так и ионы железа способны взаимодействовать с липопротеинами низкой плотности,
окисляя их. Сублетальные концентрации окисленных липопротеинов при
взаимодействии с эндотелием индуцируют активность гемоксигеназы -1 и ферритина.
Подобного рода реакции могут лежать в основе геморрагических повреждений в
центральной нервной системе, и, возможно, атерогенеза, при котором гем
гемоглобина может индуцировать формирование гидроперекисей жирных кислот [27].
Свободный гем включается в состав эндотелиальных клеток, усиливая окислительные
повреждения, индуцированные перекисью водорода. При этом выявлено возрастание
перекисного окисления липидов эндотелиальных клеток. Повреждения эндотелия под
влиянием свободного гема могут усиливать сосудистые поражения при различных
клинических синдромах, включая острую почечную недостаточность после сильного
внутрисосудистого гемолиза, при давленых повреждениях и ранах, реперфузиях
после инфаркта миокарда (возможно, как результат высвобождения миоглобина
сердечной мышцы), ретролентальной фиброплазии связанной с гемопексиновой
недостаточностью новорожденных, и, возможно, атеросклерозом. [28].
Внутрисосудистый гемолиз также может быть связан с такими патологическими
состояниями как гемоглобинопатии, травмы, малярия и бактериальные инфекции.
Среди белков плазмы крови, которые защищают от окислительных повреждений,
обусловленных лизисом эритроцитов, основную роль играют гаптоглобин и
гемопексин. Гаптоглобин и гемопексин, соединяясь с высокой степенью специфичности
с гемоглобином и гемом, соответственно, осуществляют антиоксидантную защиту
путем предупреждения образования АФК. Гаптоглобин и гемопексин необходимы для
защиты от спленомегалии и фиброза печени, индуцированных внутрисосудистым
гемолизом. [29].
Окислительные повреждения эритроцитов с
формированием таких АФК как перекись водорода, гидроксильный радикал и др.
также наблюдаются при ex vivo хранении эритроцитов. При этом наблюдаются
повреждения гемоглобина, с последующим гемолизом клеток. Выработка АФК
индуцируется гипотермическими условиями хранения крови, криопротекторами,
обезвоживанием клеток. При длительном хранении крови наблюдается активное
формирование АФК, агрегация денатурировавшего гемоглобина к фосфолипидам
мембраны, и высвобождение мембранных микровезикул, содержащих гемоглобин [30].
Таким образом, окисление гемоглобина приводит к
высвобождению свободного гема в плазме крови. Гем присоединяется к гемопексину
и поглощается моноцитами и макрофагами по рецептор-зависимому пути, затем
утилизируясь в этих клетках при помощи гемоксигеназной системы.
Микросомальная гемоксигеназа катаболизирует гем
до билирубина, монооксида углерода и ионов железа Fe2+ , которые затем
включаются в состав ферритина. Временное хранение железа в составе ферритина
может ограничивать вступление железа в реакцию Фентона, таким образом, уменьшая
окислительные повреждения клеток. Этот протективный эффект увеличивается
благодаря билирубину, который является весьма эффективной ловушкой пероксильных
радикалов, в особенности, если действует вместе с токоферолом плазмы крови.
Таким образом гемоксигеназа -1 и ферритин способствуют переносу ионов железа в
ферритиновый кор, где железо менее доступно для метаболических путей. Эти
изменения сопровождаются возрастанием цитопротекции против индуцированного
железом повреждения клеток, остановкой клеточного роста путем снижения
зависимой от железа активности рибонуклеотидредуктазы, фермента, необходимого
для синтеза ДНК и подавлением синтеза гемовых простетических групп. [31].
Относительно механизма действия внеклеточных
ионов Fe3+ показано их связывание с мембраной клетки и генерация активных форм
кислорода в зоне контакта с мембраной[32].
Транспорт ионов железа Fe3+ и Fe2+ внутрь клеток
осуществляется в норме различными путями. Так, показано, что ионы Fe3+
транспортируются внутрь клеток через β3-интегрин
и мобилферрин, в то время как транспорт ионов Fe2+ облегчается переносчиком
двухвалентных металлов. В то же время, добавление к культурам клеток
восстановительных агентов изменяет связывание железа с мобилферрином и
переносчиком двухвалентных металлов. На данный момент не установлен основной
путь транспорта ионов железа внутрь клеток человека, не смотря на то, что
известны различные механизмы [33]
ГЛАВА 2. МАТЕРИАЛЫ И
МЕТОДЫ ИССЛЕДОВАНИЯ
.1 Объект исследования
Для исследования брали эритроциты человека из
донорской эритромассы (ХОЦСК, кровь (мужская) группы А(II), Rh+, консервант -
глюгицир). Эритроциты получали трехкратным отмыванием в изотоническом буфере
(NaH2PO4, Na2HPO4, 8,5% NaCl, рН 7,4). Для этого добавляли изотонический буфер
к осадку эритроцитов в соотношении 1:10 и осторожно перемешивали стеклянной
палочкой. Затем пробирки центрифугировали в течение 10 мин при 3 тыс. об/мин.
Промывание и центрифугирование повторяли еще 2 раза. Данная обработка позволяет
удалить консервант и лизированные клетки.
2.2 Проведение
инкубации эритроцитов в опытах in vitro
Эритроциты (гематокрит 20%) инкубировали при
+370С в изотоническом буфере без добавок (контроль) или в присутствии
соответствующих реагентов (опытные группы, табл.2.1). Инкубация в присутствии
хлорида железа (III) как интактных, так и предварительно обработанных одним из
дополнительных реагентов (азидом натрия, глутатионом, цистеином или сапонином)
эритроцитов проводилась в течение 60 минут.
Эритроциты, предварительно инкубированные в
присутствии азида натрия (для ингибирования каталазы), глутатиона или цистеина
(для модуляции тиолового статуса), или сапонина (для увеличения проницаемости
липидного бислоя), далее осаждались (10 мин при 3 тыс. об/мин) и осадок
эритроцитов добавляли в среду, содержащую FeCl3. Длительность предварительной и
итоговой инкубации, а также концентрация реагентов указаны в табл.2.1.
Табл.2.1. Виды воздействий на эритроциты in
vitro
|
Группы
|
Конечная
концентрацияреагента
|
Время
инкубации
|
Условия
прединкубации
|
|
1.
Контроль
|
-
|
60
мин
|
-
|
|
2.
FeCl3
|
0.025мг%
|
60
мин
|
-
|
|
3.
Азид натрия
|
0,01%
|
60
мин
|
0,01%
NaN3 20 мин
|
|
4.
Азид натрия и FeCl3
|
0.025мг%
FeCl3
|
60
мин
|
0,01%
NaN3 20 мин
|
|
5.
Глутатион
|
1
ммоль/л
|
60
мин
|
GSH
1мM 15 мин
|
|
6.
Глутатион и FeCl3
|
0.025мг%
FeCl3
|
60
мин
|
GSH
1мM 15 мин
|
2.3 Определение
каталазной активности
Для определения каталазной активности
использовали 2%-ную суспензию эритроцитов (после инкубации).
Об активности фермента судили по убыли перекиси
водорода в инкубационной среде, которая содержала 1,2 мл 10 мМ К-фосфатного
буфера (pH = 7,4) и 0,1 мМ ЭДТА, а также 0,015 мл 15 мМ раствора перекиси
водорода. Суспензию вносили в инкубационную смесь в количестве 0,010 мл.
Снижение оптической плотности измеряли против контрольной пробы (без
эритроцитов) при Т=37 оС на спектрофотометре Specord UV VIS при 240 нм.
Активность фермента расcчитывали с учетом
молярного коэффициента экстинкции для Н2О2 (0,0394 М 6 см -1). Выражали
активность в мкмоль Н2О2 на 1 мг белка за 1 мин [34].
2.4 Определение
содержания белка
Содержание белка определяли по методу Лоури в
модификации Миллера [36]. В 5 мл 0,1N NaOH разводили 0,02 мл суспензии
эритроцитов. Затем к 0,5 мл разведенного образца суспензии эритроцитов
приливали 0,5 мл реактива В (100 мл 10%-ного раствора Na 2CO3 в 0,5N NaOH
смешивали с 10 мл раствора Б: 0,5% CuSO4, 1% К-Na-виннокислый) и оставляли при
комнатной температуре на 10 мин. Затем приливали 1,5 мл 0,15 М реактива Фолина
и прогревали 10 мин на водяной бане при 50 0С. После охлаждения интенсивность
появившейся окраски измеряли на спектрофотометре при 638 нм против контрольной
пробы, которая содержала вместо опытного образца 0,1N NaOH. Концентрацию белка
определяли с помощью калибровочных кривых, для построения которых в качестве
стандарта использовали сывороточный бычий альбумин. [35].
2.5 Определение степени
пероксидного гемолиза эритроцитов
Метод основан на измерении интенсивности
поглощения при 540 нм.
В работе были использованы такие реактивы:
. Натрий-фосфатный изотонический буфер (NaH2PO4,
Na2HPO4, 8,5% NaCl, рН 7,4).
. Дистиллированная вода
. Перекись водорода (3%)
. Азид натрия (1 мМ)
Ход работы:
В работе использовались осадки, полученные при
предварительной инкубации. Из осадка каждой группы (контроль, хлорид железа,
глутатион, глутатион+железо ) отбирались эритроциты в 2 колбочки из темного
стекла, содержащие изотонический буфер - в одну баночку добавлялась перекись
водорода (опытная проба), во вторую - нет (контрольная проба).
В контрольных пробах к 5.5 мл изотонического
буфера добавлялось 50 мкл эритроцитов 50 мкл азида натрия, в опытных к 5 мл
изотонического буфера добавлялось 50 мкл эритроцитов, 50 мкл азида натрия , а
затем добавлялось 50 мкл перекиси водорода до конечной концентрации 0,3%
Суспензию эритроцитов инкубировали при
помешивании в термостате при 37°С с доступом воздуха. Из колбочек через 60
минут отбирали по 0.5 мл инкубационной смеси и приливали в центрифужные
пробирки соответственно к 3 мл изотонического буфера (проба №1) или к 3 мл
дистиллированной воды (проба №2). Пробы центрифугировали (10 мин при 3 тыс.
об/мин). Затем измеряли оптическую плотность надосадочной жидкости на
фотоэлектроколориметре против дистиллированной воды.
Степень гемолиза выражали в процентах (гемолиз в
пробе № 2 принимали за 100%).
Расчет % гемолиза:
(Е1/Е2)*100%, где Е1- экстинкция пробы №1 , Е2 -
экстинкция пробы №2. [36], [37].
2.6
Определение скорости лизиса эритроцитов
Кинетику гемолиза при действии хлорида железа
определяли по изменению оптической плотности при 670 нм (за счет снижения
светорассеяния за счет лизиса клеток). Эритроциты инкубировались в течение 60
минут при 37°С в термостате при помешивании в присутствии нижеперечисленных
веществ (конечные концентрации в 5 мл инкубационной среды, содержащей 0,5 мл
эритроцитов и 4,5 мл изотонического натрий-фосфатного буфера):
1) 0.05% FeCl3
2) 1 mM NaN3
) 1 mM NaN3+0.05% FeCl3
) 10 mM GSH
) 10 mM GSH+0.05% FeCl3
(предварительная обработка эритроцитов
глутатионом и азидом натрия (группы 3 и 5) происходила в течение 15 минут при
37°С в термостате при помешивании, после чего эритроциты осаждали в течение 10
мин при 3 тыс. об/мин, отмывали азид натрия, и вносили 0,5 мл эритроцитов
каждой группы в колбочки, содержащие 0,05% раствор FeCl3 в 4,5 мл изотонического
буфера и инкубировались 60 минут при 37°С в термостате при помешивании;
эритроциты 2 и 4 группы инкубировались в течение 15 минут при 37°С в термостате
при помешивании соответствующими реагентами, после чего эритроциты осаждали в
течение 10 мин при 3 тыс. об/мин, отмывали азид натрия, и вносили 0,5 мл
эритроцитов в 4,5 мл изотонического буфера и инкубировались 60 минут при 37°С в
термостате при помешивании)
Затем получали осадок эритроцитов исследуемых
групп (10 мин при 3 тыс. об/мин), из осадка брали 20 мкл эритроцитов и вносили
в кювету, содержащую 0,5% раствор FeCl3 в изотоническом буфере. В течение
каждых 30 секунд отмечали изменение оптической плотности при 670 нм против 0,5%
раствора FeCl3. Исследование производилось при помешивании пробы, при комнатной
температуре. Расчет степени гемолиза эритроцитов в каждые 30 секунд времени
происходил по формуле En *100% /∆ Eобщ , где En - оптическая плотность
пробы в данный момент времени, ∆ Eобщ - разность между начальным и
конечным значением оптической плотности всего исследования. Далее суммировались
значения степени лизиса клеток через каждые 30 секунд. Полное время лизиса
соответствовало 100% лизировавшихся эритроцитов.
Скорость гемолиза рассчитывали как величину
обратную времени лизиса 50% клеток. [38-41].
2.7
Статистическая обработка результатов
Нормальность распределения
полученных данных (в каждой группе) оценивалась по критерию Шапиро-Уилка.
Учитывая нормальное распределение данных, для каждой группы рассчитывались
средние значения и стандартные отклонения.
Для оценки достоверности
различий между группами использовался t-тест Стьюдента для независимых
переменных. Уровень значимости корректировался с учетом множественных сравнений
(указан для каждой группы данных в разделе 3). [42].
Глава 3. Результаты и
их обсуждение
.1 ВЛИЯНИЕ ХЛОРИДА ЖЕЛЕЗА (III), АЗИДА НАТРИЯ И
ВОССТАНОВЛЕННОГО ГЛУТАТИОНА НА КАТАЛАЗНУЮ АКТИВНОСТЬ.
Данные об активности каталазы эритроцитов
человека, предынкубированных с азидом натрия, хлоридом железа (III), или
восстановленным глутатионом представлены на рис. 3.1

Рис. 3.1 Каталазная активность эритроцитов при
влиянии хлорида железа (III), азида натрия и восстановленного глутатиона. (*
р<0.01 по отношению к контролю)
У эритроцитов, инкубировавшихся в присутствии
азида натрия в выбранной концентрации, в том числе после их последующей
инкубации с хлоридом железа установлено снижение каталазной активности на 30% и
на 20%, соответственно. После остальных изученных воздействий каталазная
активность не отличалась от контрольных значений.
Полученные результаты согласуются с данными об
ингибировании каталазы азидом натрия. Следует отметить, что по дааным
параллельных экспериментов, установлено накопление ТБК-активных продуктов, что,
очевидно, является следствием ингибирования каталазной активности и
свидетельствует об прооксидантоном действии ионов железа. Инкубация в
присутствии хлорида железа или хлорида железа после предобработки глутатионом
не вызывала изменений каталазной активности.
.2 ПЕРОКСИДНАЯ РЕЗИСТЕНТНОСТЬ ЭРИТРОЦИТОВ
ЧЕЛОВЕКА ПРИ ДЕЙСТВИИ ХЛОРИДА ЖЕЛЕЗА (III) И ГЛУТАТИОНА
Данные о пероксидном гемолизе эритроцитов
представлены на рис. 3.2
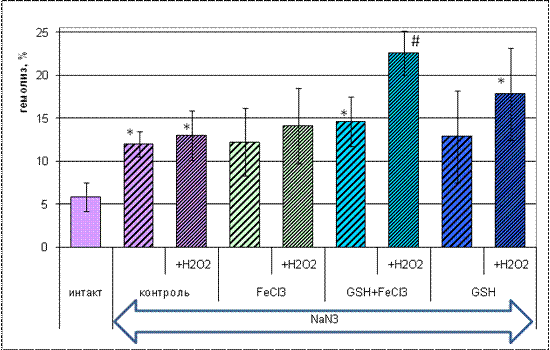
Рис 3.2 Пероксидная резистентность эритроцитов
человека под влиянием хлорида железа (III) и глутатиона. (*- p<0,01 по
отношению к интактным эритроцитам, #- p<0,01 по отношению к инкубации без
Н2О2)
В присутствии перекиси водорода ионы железа
могут вступать в окислительно-восстановительные превращения (реакция Фентона) с
формированием таких АФК, как гидроксильный радикал и радикал гидроперекиси.
Радикалы гидроксила гидроперекиси чрезвычайно
активны и разрушают различные по структуре молекулы. [1].
Ионы Fe(3+) также каталитически увеличивают
выработку активных форм кислорода, которые повреждают затем белки и липиды
эритроцитов.
Предварительная инкубация эритроцитов в
присутствии ингибитора каталазы азида натрия вызывала повышение процента лизиса
относительно интактных эритроцитов в 1,5-4 раза в зависимости от последующего
воздействия (в контрольной группе как обработанных перекисью водорода, так и не
обработанных ею - приблизительно в 2,5 раза, в группе, инкубировавшейся в
присутствии глутатиона и хлорида железа - в 3 раза) .Добавление Н2О2 в среду
инкубации вызвало дальнейшее повышение процента лизиса только эритроцитов,
предварительно инкубированных с восстановленным глутатионом (в 1,8 раз).
Предварительная обработка азидом натрия
эритроцитов, использовавшихся для изучения пероксидного гемолиза, привела к
повышению степени лизиса в контрольной группе, при инкубации с глутатионом, а
также при инкубации с глутатионом и хлоридом железа. Таким образом, в данном
исследовании также показан ингибиторный эффект азида натрия относительно
активности каталазы.
Восстановленный глутатион, очевидно, участвует в
восстановлении ионов железа до Fe2+ с их последующим вступлением в реакцию
Фентона, что может объяснить значительное снижение пероксидной резистентности
эритроцитов при совместной обработке глутатионом и хлоридом железа, в то время
как каталазная активность сохраняется на уровне контроля..
3.3 КИНЕТИКА ГЕМОЛИЗА ЭРИТРОЦИТОВ ПОД ВЛИЯНИЕМ
ХЛОРИДА ЖЕЛЕЗА (III)
Кинетическая кривая распределения эритроцитов по
стойкости отражена на рис. 3.3

Рис. 3.3. Доля лизированных эритроцитов в
зависимости от времени инкубации в присутствии 0,5% хлорида железа (III)
Концентрация хлорида железа подобрана на
основании предварительной серии экспериментов с расчетом проведения полного
лизиса эритроцитов за 5-7 минут. При этом, согласно полученным результатам
хлорид железа (III) в концентрации 0,5% вызывал полный лизис эритроцитов за 5
минут. Основная доля эритроцитов начинает лизироваться после 1,5 минут
инкубации, достигая максимума лизиса через 2,5 минуты инкубации. К 5 минутам
инкубации оптическая плотность достигает стабильного минимума, что
свидетельствует о гемолизе 100% эритроцитов.
Как видно из рис. 3.3 максимальный лизис
эритроцитов (без предобработки и после предобработки хлоридом железа)
отмечается после 1,5 минуты инкубации.
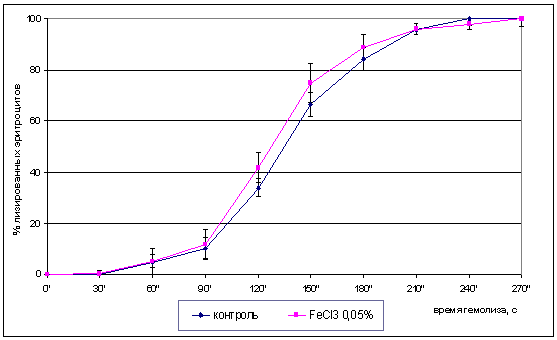
Рис 3.4. Кинетика гемолиза эритроцитов 0,5%
хлоридом железа (III). (*-p<0,05 по отношению к интактным эритроцитам)
Как видно из рис. 3,4 после предварительной
обработки хлоридом железа в негемолитической концентрации, кривая гемолиза в
присутствии 0,5% хлорида железа смещена с достоверным повышением степени
гемолиза после 120 и 150 секунд инкубации. Учитывая данные литературы о
взаимодействии хлорида железа (III) с мембраной эритроцита [] , можно предполложить
накопление железа в мембранах после первой инкубации, что снижает устойчивость
эритроцитов к гемолизу.
Установлено, что для эритроцитов контрольной
группы время лизиса 50% клеток составило 135 с, а для эритроцитов
инкубировавшихся с хлоридом железа - 130 с .
3.4 ВЛИЯНИЕ АЗИДА НАТРИЯ И ХЛОРИДА ЖЕЛЕЗА НА
КИНЕТИКУ ГЕМОЛИЗА ЭРИТРОЦИТОВ ПРИ ДЕЙСТВИИ ХЛОРИДА ЖЕЛЕЗА (III)
Данные о влиянии предынкубации эритроцитов в
растворе Fecl3 на кинетические параметры гемолиза эритроцитов, инкубированных в
растворе азида натрия представлены на рис. 3.5

Рис. 3.5 Влияние предварительной инкубации в
присутствии азида натрия и хлорида железа (0,05%) на кинетику гемолиза
эритроцитов 0,5% хлоридом железа (III) (*-p<0,05 по отношению к инкубации с
азидом натрия)
Ввиду того, что действие ионов Fe(3+)
сопровождается генерацией активных форм кислорода, возможно изменение
кинетических параметров гемолиза при инкубации эритроцитов в растворе азида
натрия, а также инкубации эритроцитов в растворе азида натрия с предварительной
обработкой этой группы 0,05% FeCl3.
Показано, что предварительная обработка хлоридом
железа в негемолитической концентрации не приводит к более быстрому гемолизу
при действии ионов железа в более высокой концентрации, в отличие от ингибитора
каталазы, который вызывает значительный рост скорости лизиса по сравнению с
контрольными эритроцитами.
Согласно полученным результатам, наблюдалось
снижение степени гемолиза в опытной группе (хлорид железа + азид натрия)
относительно контрольной (азид натрия) после 120, 150 и 180 секунд исследования
в 2,8; 1,4 и 1,2 раза соответственно. Максимум гемолиза эритроцитов в
контрольной группе наблюдался после 120 секунд проведения опыта, а в опытной -
после 150 секунд исследования.
Установлено, что для эритроцитов,
инкубировавшихся с азидом натрия время лизиса 50% клеток составило 120 с, в то
время как для эритроцитов, инкубировавшихся с азидом натрия и хлоридом железа
-138 с.
.5 ВЛИЯНИЕ ВОССТАНОВЛЕННОГО ГЛУТАТИОНА НА
КИНЕТИКУ ГЕМОЛИЗА ЭРИТРОЦИТОВ ХЛОРИДОМ ЖЕЛЕЗА (III)
Данные о влиянии предынкубации эритроцитов в
FeCl3 на кинетические параметры гемолиза эритроцитов, инкубированных в растворе
глутатиона представлены на рис. 3.6.
Согласно полученым результатам, кинетическая
кривая при действии хлорида железа на фоне восстановленного глутатиона
значительно смещена по сравнению с показателями, полученными при действии
восстановленного глутатиона. Повышение степени гемолиза наблюдалось через 120,
150 и 180 секунд инкубации, соответственно в 1,6; 1,65; 1,3 раза. Максимум
лизиса эритроцитов в группе, инкубировавшейся только с глутатионом, наблюдался
после 150 секунд проведения опыта, а в группе, инкубировавшейся с глутатионом и
хлоридом железа - после 180 секунд исследования.
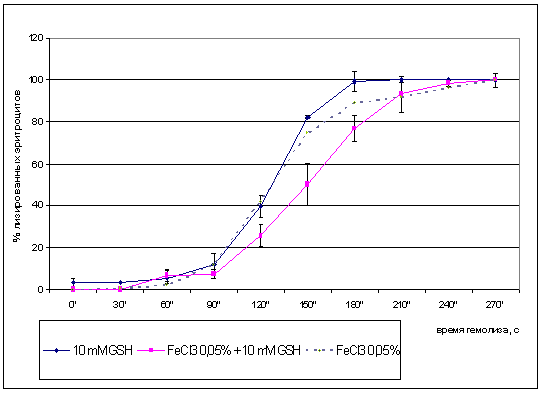
Рис. 3.6 Влияние предварительной инкубации в
присутствии глутатиона и хлорида железа (0,05%) на кинетику гемолиза
эритроцитов 0,5% хлоридом железа (III) (*-p<0,05 по отношению к инкубации с
восстановленным глутатионом) .
Глутатион выступает как мембранопроникающий
низкомолекулярный антиоксидант, обеспечивающий восстановленное состояние
сульфгидрильных групп белков. Важное значение для резистентности эритроцитов к
гемолизу имеет поддержание в восстановленной форме SH-групп белков цитоскелета,
транспортных систем и ферментов. [16]
Учитывая различные механизмы мембранного
переноса для ионов Fe2+ и Fe(3+), что известно из данных литературы [33], можно
предположить , что при действии восстановленного глутатиона, большая доля
железа поступает в эритроциты, не повреждая компоненты мембраны. Таким образом,
наблюдается более медленный лизис по сравнению с действием только хлорида
железа без обработки глутатионом.
Установлено, что для эритроцитов,
инкубировавшихся с глутатионом, время лизиса 50% клеток составило130с, а для
эритроцитов, инкубировавшихся с глутатионом и хлоридом железа - 150с.
.6 СКОРОСТЬ ГЕМОЛИЗА ЭРИТРОЦИТОВ ПРИ ДЕЙСТВИИ
ХЛОРИДА ЖЕЛЕЗА (III), АЗИДА НАТРИЯ И ВОССТАНОВЛЕННОГО ГЛУТАТИОНА
Данные о скорости гемолиза в исследуемых группах
представлены на рис. 3.7
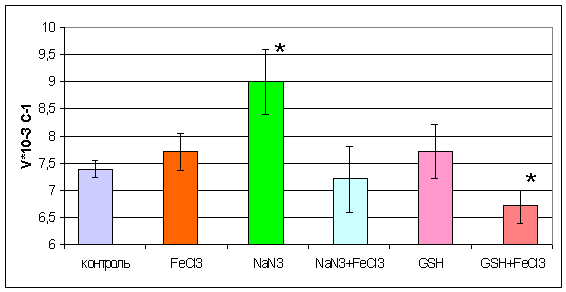
Рис. 3.7 Скорость гемолиза эритроцитов при
действии хлорида железа (III), азида натрия и восстановленного глутатиона.
(*-р<0,01% по отношению к контролю)
Согласно полученным данным, изменение скорости
гемолиза наблюдалось в двух исследуемых группах. При обработке эритроцитов
азидом натрия и при обработке эритроцитов восстановленным глутатионом, а затем
0,05% раствором FeCl3. Так, в первом случае наблюдалось повышение скорости
гемолиза относительно эритроцитов, не подвергавшихся предварительной обработке
в 1,22 раза. Скорость лизиса под воздействием 0,5% хлорида железа повышалась на
20% после обработки азидом по сравнению с контрольной группой, что может
свидетельствовать о важной роли каталазной активности в поддержании
устойчивости эритроцитов к данному виду гемолиза, т.к. азид ингибировал
активность каталазы (рис. 3.1) . При обработке эритроцитов глутатионом
восстановленным, а затем хлоридом железа наблюдалось снижение скорости лизиса
на 10%.
На основании полученных результатов и данных
литературы можно предположить, что снижение активности каталазы при действии
азида натрия и совместном действии азида и хлорида железа приводит к
значительному накоплению перекиси водорода и активации свободнорадикальных
процессов. Восстановленный глутатион, очевидно, участвует в восстановлении
ионов железа до Fe2+ с их последующим вступлением в реакцию Фентона, что может
объяснить значительное снижение пероксидной резистентности эритроцитов при
совместной обработке глутатионом и хлоридом железа, в то время как каталазная
активность сохраняется на уровне контроля.
Важная роль окислительных процессов в развитии
гемолиза при действии хлорида железа была выявлена и при изучении кинетики
данного процесса. Показано, что предварительная обработка хлоридом железа в
негемолитической концентрации не приводит к более быстрому гемолизу при
действии ионов железа в более высокой концентрации, в отличие от ингибитора
каталазы, который вызывает значительный рост скорости лизиса по сравнению с
контрольными эритроцитами.
ВЫВОДЫ
1. У эритроцитов, инкубированных в присутствии
азида натрия в выбранной концентрации, в том числе после их последующей
инкубации с хлоридом железа установлено снижение каталазной активности на 30% и
на 20%, соответственно. При остальных изученных воздействиях каталазная
активность не отличалась от контрольных значений.
2. Предварительная инкубация эритроцитов в
присутствии ингибитора каталазы азида натрия вызывала повышение процента лизиса
эритроцитов, обработанных азидом натрия в 2,5-3 раза относительно интактных
эритроцитов - вне зависимости от присутствия перекиси водорода. Дальнейшее
повышение степени лизиса (в 1,8 раза) при добавлении в среду инкубации перекиси
водовода отмечено только для эритроцитов, предварительно инкубированных с
восстановленным глутатионом.
. Хлорид железа (III) в концентрации 0,5%
вызывал полный лизис эритроцитов человека за 5 мин инкубации при 25оС.
Кинетическая кривая выявила S-образный характер с максимальным лизисом
эритроцитов в области от 1,5 до 3,5 минут инкубации вне зависимости от
предварительной обработки эритроцитов.
. Предварительная обработка эритроцитов
хлоридом железа в негемолитической концентрации (0,05%) не приводила к
изменению скорости лизиса хлоридом железа по сравнению с контрольными
эритроцитами, в отличие от азида натрия, обработка которым увеличивала скорость
гемолиза в присутствии 0,5% FeCl3.
. Последовательная предварительная
обработка эритроцитов восстановленным глутатионом и далее FeCl3 вызывала
снижение скорости лизиса эритроцитов хлоридом железа.
СПИСОК ЛИТЕРАТУРЫ
1. Чеснокова
Н.П., Понукалина Е.В., Бизенкова М.Н.(2007) Механизмы структурной и
функциональной дезорганизации биосистем под влиянием свободных радикалов.
Научный журнал "Фундаментальные исследования" №4, 2007 год
2. Woollard
KJ
<http://www.ncbi.nlm.nih.gov/sites/entrez?Db=pubmed&Cmd=Search&Term=%22Woollard%20KJ%22%5BAuthor%5D&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_DiscoveryPanel.Pubmed_RVAbstractPlus>,
Sturgeon S <http://www.ncbi.nlm.nih.gov/sites/entrez?Db=pubmed&Cmd=Search&Term=%22Sturgeon%20S%22%5BAuthor%5D&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_DiscoveryPanel.Pubmed_RVAbstractPlus>,
Chin-Dusting JP <http://www.ncbi.nlm.nih.gov/sites/entrez?Db=pubmed&Cmd=Search&Term=%22Chin-Dusting%20JP%22%5BAuthor%5D&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_DiscoveryPanel.Pubmed_RVAbstractPlus>,
Salem HH
<http://www.ncbi.nlm.nih.gov/sites/entrez?Db=pubmed&Cmd=Search&Term=%22Salem%20HH%22%5BAuthor%5D&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_DiscoveryPanel.Pubmed_RVAbstractPlus>,
Jackson SP
<http://www.ncbi.nlm.nih.gov/sites/entrez?Db=pubmed&Cmd=Search&Term=%22Jackson%20SP%22%5BAuthor%5D&itool=EntrezSystem2.PEntrez.Pubmed.Pubmed_ResultsPanel.Pubmed_DiscoveryPanel.Pubmed_RVAbstractPlus>(2009)
Erythrocyte hemolysis and hemoglobin oxidation promote ferric chloride-induced
vascular injury. J Biol Chem.
<javascript:AL_get(this,%20'jour',%20'J%20Biol%20Chem.');> 2009 May 8;284(19):13110-8.
Epub 2009 Mar 10
. Hagen
T. M., Ingersoll R. T., Lykkesfeldtl J., LIU J.,Wehr C. M., Vinarsky V.,
Bartholomew J. C.,* and Ames B. N.( 1999 ) (R)-a-Lipoic acid-supplemented old
rats have improved mitochondrial function, decreased oxidative damage, and
increased metabolic rate The FASEB Journal. 1999; 13:411-418
. McMillan
D.C., Powell C. L., Bowman Z. S., Morrow J. D., Jollow D.J, (2005) Lipids
versus proteins as major targets of pro-oxidant direct-acting hemolytic agens.
Toxicological sciences 88(1) :274-283 (2005)
. Kartha
V. N. R. and Krishnamurthy S. (1978) Effect of hypervitaminosis A on hemolysis
and lipid peroxidation in the rat, Journal of Lipid Research Volume 19, 1978
. Wallace
William J $, . Houtchensg Robert A, . Maxwelll John C, and Caughey Winslow S.
Mechanism of Autooxidation for Hemoglobins and Myoglobins .The Journal of
biological chemistry,1982, issue of may 10, p.4966-4977
. Hagen
T. M., Ingersoll R. T., Lykkesfeldtl J., LIU J.,Wehr C. M., Vinarsky V.,
Bartholomew J. C.,* and Ames B. N.( 1999 ) (R)-a-Lipoic acid-supplemented old
rats have improved mitochondrial function, decreased oxidative damage, and
increased metabolic rate The FASEB Journal. 1999; 13:411-418
. Yamaguchi
T, Terada S.(2003) Analysis of high-pressure-induced disruption of human
erythrocytes by flow cytometry Cell Mol Biol Lett. 2003;8(4):1013-6
. Проданчук
Г.Н., Балан Г.М. Токсические метгемоглобинемии: механизмы формирования и пути
оптимизации лечения. Клінічна токсикологія
. Rhee
S. G., Chang T-S. , Bae Y. S., Lee S-R. and Kang S. W.;(2003) Cellular
Regulation by Hydrogen Peroxide J Am Soc Nephrol 14:S211-S215, 2003
. Starke
D. W., Chock P. B., and Mieyal J. J.( 2003) Glutathione-Thiyl Radical
Scavenging and Transferase Properties of Human Glutaredoxin (Thioltransferase).
J. Biol. Chem., Apr 2003; 278: 14607 - 14613
. Игонина
Н. А. (2007) Особенности оказания медицинской помощи пострадавшим от
воздействия метгемоглобинообразователей
. Гармонов
С.Ю., Евгеньев М.И., Зыкова И.Е. (2003) Перспективные методы оценки генетически
детерминированной химической чувствительности организма человека. Химическая и
биологическая безопасность. 2003. № 11-12
. Мустаев
О. З. (2006) Влияние пропофола и кетамина на процесс липидпероксидации крови и
его коррекция альфа-токоферолом у больных при холецистэктомии
. Frei
B, Stocker R, and Ames B N (1988) Antioxidant defenses and lipid peroxidation
in human blood plasma PNAS December 1, 1988 vol. 85 no. 24 9748-9752
. Raftos
J. E.,*, S Whillier S. and Kuchel P. W.; (2010) Glutathione synthesis and
turnover in the human erythrocyte: Alignment of a model based on detailed
enzyme kinetics with experimental data; jbc.M109.067017 May 24, 2010, doi:
10.1074
. Барабой
В. А., (2005) α-липоевая
-дигидролипоевая кислоты-активная биоантиоксидантная и биорегуляторная система.
Укр. біохім. журн., 2005, т. 77, № 3
. Smith
A.R.1; Shenvi S.V.1; Widlansky M.1; Suh J.H.1; Hagen T.M (2004) Lipoic Acid as
a Potential Therapy for Chronic Diseases Associated with Oxidative Stress.
Current Medicinal Chemistry, Volume 11, Number 9, May 2004 , pp. 1135-1146(12)
. Hagen
T. M., Ingersoll R. T., Lykkesfeldtl J., LIU J.,Wehr C. M., Vinarsky V.,
Bartholomew J. C., and Ames B. N.( 1999 ) (R)-a-Lipoic acid-supplemented old
rats have improved mitochondrial function, decreased oxidative damage, and
increased metabolic rate The FASEB Journal. 1999; 13:411-418
. Packer
L, Witt EH, Tritschler HJ. (1995) alpha-Lipoic acid as a biological
antioxidant.Free Radic Biol Med. 1995 Aug; 19(2):227-50
. Smith
A.R.1; Shenvi S.V.1; Widlansky M.1; Suh J.H.1; Hagen T.M (2004) Lipoic Acid as
a Potential Therapy for Chronic Diseases Associated with Oxidative Stress.
Current Medicinal Chemistry, Volume 11, Number 9, May 2004 , pp. 1135-1146(12)
. Барабой
В. А., (2005) α-липоевая
-дигидролипоевая кислоты-активная биоантиоксидантная и биорегуляторная система.
Укр. біохім. журн., 2005, т. 77, № 3
. Coleman
M. D, Tolley H. L, and Desai A. K (2001) Monitoring antioxidant effects using
methaemoglobin formation in diabetic erythrocytes† .The British Journal of
Diabetes & Vascular Disease, Jan 2001; 1: 88 - 92
. Balla
J, Balla G, Lakatos B, Jeney V, Szentmihályi K(2007)Heme-iron in the
human body; Orv Hetil. 2007 Sep 9;148(36):1699-706.
. Belcher
JD, Beckman JD, Balla G, Balla J, Vercellotti G. (2010)Heme degradation and
vascular injury; Antioxid Redox Signal. 2010 Feb;12(2):233-48
. Jeney
V, Balla J, Yachie A, Varga Z, Vercellotti GM, Eaton JW, Balla G; (2002)
Pro-oxidant and cytotoxic effects of circulating heme; Blood. 2002 Aug
1;100(3):879-87.
. Balla
G, Vercellotti G, Eaton JW, Jacob HS; (1990) Heme uptake by endothelium
synergizes polymorphonuclear granulocyte-mediated damage; Trans Assoc Am
Physicians. 1990;103:174-9.
. Tolosano
E., Fagoonee S., Hirsch E., Franklin G.;( 2002 ) Enhanced splenomegaly and severe
liver inflammation in haptoglobin/hemopexin double-null mice after acute
hemolysis; Blood. 2002 Dec 1;100(12):4201-8. Epub 2002 Jul 25.
. Kanias
T, Acker JP(2010) Biopreservation of red blood cells--the struggle with
hemoglobin oxidation; FEBS Journal, Volume 277, Number 2, January 2010 , pp.
343-356(14)
. P.M.
Schwartsburd (2001), Self-cytoprotection against stress: feedback regulation of
heme- dependent metabolism Cell Stress Chaperones. 2001 January; 6(1): 1-5.)
. Baysala
E; Sullivanb S. G.; Sterna A.(1989) Binding of Iron to Human Red Blood Cell
Membranes; Free Radic. Res Commun. 1989(1):55-9
. Conrad
M. E. , Umbreit J. N. 1, Moore El. G. ;Separate pathways for cellular uptake of
ferric and ferrous iron (2000) Am J Physiol Gastrointest Liver Physiol 279:
G767-G774, 2000; Vol. 279, Issue 4, G767-G774, October 2000
. Scott
M.D., Lubin B.H., Zuo L. Erythrocyte defense against hydrogen peroxide:
preeminent importance of catalase// J. Lab. Clin. Med. - 1991. - Vol.118. -
P.7-16
. Miller
G.L. Protein determination for large numbers of samples// Anal. Chem.- 1959.-
V.31, №5.- P.964-966
. Карагезян
К.Г., Овакимян С.С., Гимоян Л.Г. и др.(2000) Изменение показателей пероксидной
резистентности эритроцитов и показателей липидного обмена плазмы крови при
развитии ишемического инсульта// Укр. биох. журн. - 2000. - т.72, №6. -
с.74-78.
. Snyder
L., Fortier N., Trainor J. et al.(1971-1977) Effect of hydrogen peroxide
exposure on normal human erythrocyte deformability, morphology, surface
characteristics, and spectrin-hemoglobin cross-linking// J.Clin. Invest.-
1985.-V.76.- P.1971-1977
. Черницкий
Е.А., Сенькович О.А., Слобожанина Е.И.(1999) Параметры гемолиза
додецилсульфатом натрия как индикатор структурного состояния мембран
эритроцитов// Биофизика.- 1999.- Т.44, вып.1.- С.66-69
. Woollard
K.J., Sturgeon S., Chin-Dusting J.P.F., Salem H.H., and Jackson S.P.
Erythrocyte hemolysis and hemoglobin oxidation promote ferric chloride-induced
vascular injury // J. Biol. Chem.- 2009.- V. 284.- P.13110 - 13118
. Mueller
S., Riedel H.-D., and Stremmel W. Direct Evidence for Catalase as the
Predominant H2O2 -Removing Enzyme in Human Erythrocytes// Blood.- 1997.- Vol.
90, N12.- P. 4973-4978
. Low
F.M., Hampton M.B., Peskin A.V., and Winterbourn C.C. Peroxiredoxin 2 functions
as a noncatalytic scavenger of low-level hydrogen peroxide in the erythrocyte//
Blood.- 2007.- V.109.- P.2611-261
. Атраментова
Л.О., Утевська О.М. Статистичнi методи в бiологii: пiдручник. - Х: ХНУ iменi
Каразiна , 2007.- 288с.