Морфоэкологические особенности ценопопуляций можжевельника обыкновенного в Припышминских борах подзоны предлесостепи Западной Сибири
Федеральное
государственное учреждение науки
«Ботанический
сад» УрО РАН
МОРФОЭКОЛОГИЧЕСКИЕ
ОСОБЕННОСТИ ЦЕНОПОПУЛЯЦИЙ
МОЖЖЕВЕЛЬНИКА
ОБЫКНОВЕННОГО В ПРИПЫШМИНСКИХ БОРАХ ПОДЗОНЫ ПРЕДЛЕСОСТЕПИ ЗАПАДНОЙ СИБИРИ
06.03.02 -
Лесоведение, лесоводство, лесоустройство и лесная таксация
Диссертация
на соискание ученой степени
кандидата
биологических наук
Дюбанова Наталья Владимировна
Екатеринбург
- 2013
Оглавление
Введение
Глава 1. Природные условия региона
1.1 Географическое положение
1.2 Климат
.3 Рельеф и гидрология
.4 Почвенные условия
.5 Лесная растительность
Глава 2. Состояние проблемы
2.1 Ареал и экоареал можжевельника
обыкновенного
.2 Морфологические особенности
.3 Размножение и естественное
возобновление
.4 Хозяйственное использование
Глава 3. Объекты и методы
исследований
.1 Объекты изучения
.2 Принципы и методы исследований
.4 Экспериментальная часть
Глава 4. Лесотипологический ареал,
рост, морфогенез и жизненность особей
.1 Рост и жизненность
.2 Морфологические особенности
.3 Опыты по искусственному
разведению
Глава 5. Экологическая структура
ценопопуляций
.1 Численность
.2 Жизненность
.3 Возрастная структура
.4 Половая структура
.5 Семенная репродукция
.6 Естественное возобновление
Глава 6. Морфоэкологические
особенности можжевельника обыкновенного на открытых местообитаниях
.1 Морфологические параметры
можжевельника обыкновенного на сплошных вырубках
.2 Морфологические параметры
можжевельника обыкновенного на гарях
Глава 7. Анализ связей роста и
жизненности можжевельника обыкновенного со структурой и конкуренцией древостоя
.1 Связь с конкуренцией древостоя
.2 Влияние абиотических факторов
Выводы
Рекомендации
Заключение
Литература
Приложения
Введение
Актуальность темы. Можжевельник обыкновенный
(Juniperus communis L.) является одним из широко распространенных и ценотически
важных видов подлеска хвойных лесов Северной Евразии и перспективным для
интродукции и озеленения видов семейства кипарисовых. Однако информация о
распространении, морфоэкологических особенностях его структуры, роста и
семеношения в различных типах леса, особенно в Западной Сибири, недостаточна.
Почти не изучены его эколого-морфологические изменения под влиянием таких
глобальных лесоразрушающих факторов как сплошные рубки и пожары, а также
ценоэкологические особенности изменений морфологических параметров
ценопопуляций можжевельника под влиянием изменений структуры и конкуренции
древостоя-эдификатора.
Виды можжевельников рода Juniperus
широко распространены в Евразии и Северной Америке, а можжевельник обыкновенный
- в лесной зоне Русской равнины, Западной и Восточной Сибири [Ареалы, 1977].
Как и другие виды хвойных с обширным
географическим ареалом, можжевельник обыкновенный обладает широкой
экологической амплитудой и генетической изменчивостью, являясь характерным
компонентом или даже доминируя в подлеске хвойных лесов и выполняя важную роль
в их формировании.
Ранее были установлены некоторые особенности
географического и экологического (лесотипологического) ареалов, морфологической
и анатомической структуры и роста можжевельника обыкновенного в лесах Русской
равнины [Каппер О.Г., 1954; Аксенова Н.А., 1976], на Урале [Бакланова Е.Г.,
1988; Аши, 1991а; Кожевников А.П., Тишкина Е.А., Кожевникова Г.М., 2009;
Кожевников А.П., Тишкина Е. А., 2009, 2011а, 2011б], в Средней Сибири [Князева
С. Г., 2000а, 2000б, 2010; Михеева Н.А., 2002а, 2002б], и в лесах Тянь-Шаня
[Мухамедшин К.Д., 1970; Мухамедшин К.Д., Таланцев Н.К., 1982]. Однако сведения
о его распространении, морфоэкологических особенностях его структуры, роста, семеношении
в различных типах леса особенно в Западной Сибири очень скудны. Почти не
изучены эколого-морфологическая изменчивость, влияние на его развитие таких
главнейших лесоразрушающих факторов, как сплошные рубки и пожары. Почти не
изучены ценоэкологические особенности изменений морфологической структуры его
популяций под влиянием изменений структуры и конкуренции древостоя-эдификатора
под его пологом.
Цель диссертации. Целью диссертационной работы
является изучение лесотипологического распределения, морфологической структуры,
жизненности, роста и семеношения в зависимости от структуры и конкуренции
древостоя-эдификатора, а также на сплошных вырубках и гарях в преобладающих
типах сосновых лесов подзоны предлесостепи Западной Сибири (на примере
Припышминских боров).
Для достижения поставленной цели решались
следующие задачи:
. Изучение лесотипологического
распределения, численности, возрастной, высотной структуры и проективного
покрытия ценопопуляций можжевельника под пологом древостоев.
. Изучение роста, морфогенеза и
жизненности особей можжевельника под пологом древостоев и на вырубках.
. Изучение половой структуры и
семеношения ценопопуляций можжевельника.
. Количественный анализ
структурно-функциональных связей роста и жизненности можжевельника с параметрами
структуры и конкуренции древостоя-эдификатора сосны обыкновенной.
5. Изучение возрастной структуры и
жизненности ценопопуляций можжевельника после сплошной рубки.
6. Разработка рекомендаций по сохранению
природных ценопопуляций можжевельника обыкновенного и его интродукции.
Обоснованность результатов исследований.
Сформулированные в диссертационной работе научные положения, выводы и
рекомендации обоснованы соответствующими цели, задачам и объектам изучения
апробированными методическими подходами и методами, достаточным объемом,
репрезентативностью и уровнем анализа натурных фактических данных, полученных в
ходе полевых исследований. Достоверность результатов обеспечена применением
современных математических методов их анализа и оценки с применением компьютерной
техники и программ.
Научная новизна. Впервые на южной границе ареала
можжевельника обыкновенного в сосновых лесах подзоны предлесостепи западной
части Западной Сибири изучены рост, морфогенез и семеношение его особей,
возрастная и половая структура, проективное покрытие и жизненность
ценопопуляций можжевельника обыкновенного под пологом древостоев в
преобладающих типах леса. На основе микроэкосистемного подхода впервые в
лесоведении на количественном уровне проанализирована связь роста, проективного
покрытия и жизненности можжевельника со структурой, конкуренцией
древостоя-эдификатора и световым режимом под его пологом.
На защиту выносятся следующие положения:
. Лесотипологическое распределение
проективного покрытия и жизненности можжевельника обыкновенного в Припышминских
борах предлесостепи Западной Сибири.
. Параметры роста, морфогенеза особей и
проективного покрытия, возрастной, половой структуры и семеношения
ценопопуляций можжевельника под пологом древостоев и на сплошных вырубках в
преобладающих типах леса.
. Структурно-функциональные связи
параметров роста и жизненности можжевельника обыкновенного с конкуренцией
древостоя-эдификатора и уровнем ФАР.
Практическая полезность и реализация
результатов. В ходе выполнения исследований были составлены схематические карты
ареала можжевельника обыкновенного в национальном парке «Припышминские боры» на
площади 49,5 тыс. га и разработаны лесоводственные рекомендации по сохранению и
введению его в культуру.
Апробация работы и научные публикации. Основные результаты
исследований представлены и доложены на: III
Всероссийской научной конференции «Принципы и способы сохранения
биоразнообразия» (Йошкар-Ола, Пущино, 2008); I
Международной научной конференции студентов, аспирантов и молодых ученых
«Фундаментальные и прикладные исследования в биологии» (Донецк, 2009); I
Всероссийской научно-практической конференции студентов, аспирантов и молодых
ученых с международным участием «Проблемы сохранения биологического
разнообразия Волжского бассейна и сопредельных территорий» (Чебоксары, 2009);
Научно-практической конференции, посвященной 5-летию природного парка «Река
Чусовая» «Охрана, экопросвещение, рекреационная деятельность, изучение
биоразнообразия и культурного наследия на ООПТ» (Нижний Тагил, 2009); III
Всероссийской научно-практической конференции с международным участием
«Биологические системы: устойчивость, принципы и механизмы функционирования»
(Нижний Тагил, 2010); Всероссийской научно-практической конференции «Актуальные
проблемы сохранения биоразнообразия на охраняемых и иных территориях» (Уфа,
2010); II
Международной научно-практической конференции «Биодиверситиология: современные
проблемы сохранения и изучения биологического разнообразия» (Чебоксары, 2010);
Всероссийской научной конференции «Отечественная геоботаника: основные вехи и
перспективы» (Санкт-Петербург, 2011).
Основные положения диссертации изложены в 12
печатных работах, в т.ч. 2 в журналах, рекомендованных ВАК.
Личный вклад автора заключается в анализе
состояния проблемы, определении цели и основных задач программы, выборе
адекватных методов и объектов исследований, сборе, обработке и анализе их
результатов, формулировании выводов и рекомендаций по введению можжевельника в
культуру, а также в написании и оформлении диссертационной работы.
В закладке пробных площадей в период с 2003 по
2008 годы принимали участие сотрудники национального парка «Припышминские боры»
Д. В. Прядеин, А. С. Дюбанов, Л. Л. Козулин, В. Б. Вахрушевой. Работа выполнена
под руководством доктора биологических наук Петровой И. В., при консультативном
содействии кандидата биологических наук Санниковой Н. С. и доктора
биологических наук, профессора, Санникова С. Н. Всем им автор выражает
искреннюю признательность.
Структура и объем диссертации. Диссертационная
работа состоит из введения, 7 глав, выводов, рекомендаций, заключения, списка
литературы, включающего 136 работ, из которых 5 на иностранных языках.
Диссертация изложена на 125 страницах, иллюстрирована 28 рисунками, содержит 16
таблиц и 7 приложений.
Глава 1. Природные условия региона
.1 Географическое положение
Исследования проводились на территории
национального парка «Припышминские боры». Ландшафтный комплекс Припышминских
боров (площадь около 350 тыс. га) относится к числу уникальных феноменов
природы Зауралья, представляющих выдающееся народно-хозяйственное,
научно-познавательное, природозащитное, рекреационное значение.
Припышминские боры, расположенные на юго-востоке
Свердловской области, - один из крупнейших в России «компактных и живописных
массивов исключительно высокопродуктивных и генетически ценных коренных
сосновых лесов (с вкраплениями ельников) с хорошо сохранившимся ядром
естественной флоры, фауны и почв» [Санников С. Н., 1961].
По исследованиям С. Н. Санникова, массив
Припышминских боров образован системой ленточно-островных сосновых лесов
различных размеров, протянувшихся почти непрерывной полосой вдоль берегов
среднего и нижнего течения р. Пышма. Общая протяженность этой полосы боров с
запада (от устья р. Дерней) на восток (до устья р. Пышма) приблизительно равна
180 км, ширина - от 6 до 45 км. Кроме того, отдельные мелкие острова сосновых
боров встречаются в 30-50 км к западу от основного массива и имеют особые
названия: Никольский, Камышловский, Реутинский боры. Несколько особняком, в
20-30 км к северу от основного Припышминского массива, расположен довольно
крупный Бахметский бор, который по природным условиям и характеру лесной
растительности также можно относить к Припышминским борам. Все эти боры,
по-видимому, представляют собой лишь разрозненные остатки некогда обширных
девственных сосновых и елово-сосновых лесов, почти сплошь покрывавших
территорию приречных террас долины р. Пышма.
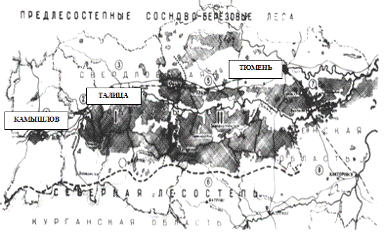
Рис. 1.
Схематическая карта лесного массива Припышминских боров (по С. Н. Санникову,
1961).
Припышминские боры - самый северный лесной
массив в системе так называемых «островных сосновых боров» Западной Сибири,
точнее Притоболья, протянувшихся меридиональной полосой на древних песчаных
отложениях вдоль долины р. Тобол от степей Северного Казахстана, через
лесостепь Курганской области до сплошных таежно-лесных массивов Свердловской и
Тюменской областей. Современная южная граница распространения таежно-лесных
массивов в Свердловской области, под влиянием деятельности человека все более
отодвигающаяся на север, находится в 60-80 км севернее Припышминских боров,
располагаясь в бассейне левых притоков р. Ница. В прошлом она, видимо,
проходила южнее.
.2 Климат
Детальный анализ климата региона Припышминских
боров как климата южной подобласти континентальной лесной западносибирской
подобласти приведен в работах С. Н. Санникова и Н. С. Санниковой (1985).
В связи с большей континентальностью и меньшим
числом пасмурных дней, среднегодовое число часов солнечного сияния здесь (около
1990) намного превосходит таковое на той же широте на Русской равнине. Общая
продолжительность вегетационного периода (со среднесуточной температурой выше
+5˚С) 160-165 дней, а с устойчивой температурой выше +10˚С - 120-126
дней. Сумма активных температур (выше + 10˚С) - 1800-2000˚С [Санников
С.Н., 1961]. Характерной чертой климата западной части Западной Сибири являются
поздние весенние (до 27 мая) и ранние осенние заморозки (с 3 сентября),
сокращающие безморозный период до 100-120 дней (в отдельные годы до 60-70 дней).
Обобщенные нами данные о продолжительности
вегетационного периода за 1995-2008 гг. представлены на рисунке 2. В работе
использованы данные метеопоста г. Талица за 14 лет (с 1995 по 2008 гг.).
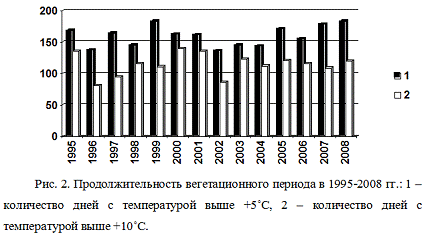
Рис. 2. Продолжительность вегетационного периода
в 1995-2008 гг.: 1 - количество дней с температурой выше +5˚С, 2 -
количество дней с температурой выше +10˚С.
Годовая сумма осадков составляет в среднем 453,5
мм (2003-2008 гг.), при этом 45-50% их выпадает в летние месяцы (рис. 3).
Минимум осадков приходится на апрель-май. Эти же месяцы, в связи с высокой
температурой и низкой относительной влажностью воздуха (47-53%),
характеризуются самым низким отношением осадков к испарению равным 0,3-0,6.
Отсюда вытекает высокая (выше 30%) вероятность засух в первой половине лета
[Санников, 1961].
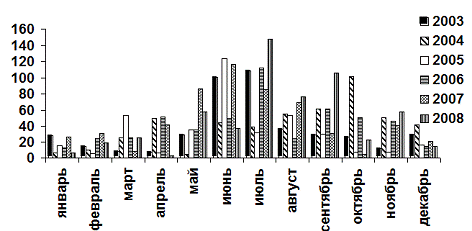
Рис. 3. Динамика выпадения осадков за 2003-2008
гг.
Устойчивый снежный покров устанавливается 12-13
ноября, а сходит (на открытых участках) в среднем 10 апреля. Мощность снежного
покрова к началу марта достигает в лесу около 80 см, но на обширных и сплошных
вырубках всего 65-70 см (запас влаги около 150 мм). На песчаных и супесчаных
почвах снеговые воды просачивается вглубь почвы, пополняя верховодки. Климатическая
характеристика района исследования приведена в таблице 1.2.1 и на
климадиаграмме (рис. 4.).
Таблица 1.2.1 - Климатическая характеристика
региона национального парка «Припышминские боры» (по данным метеопоста г.
Талица за 2003-2010 гг.)
|
Месяцы
|
Температура
(°)
|
Кол-во
осадков (мм)
|
Снежный
покров (см)
|
Относительная
влажность (%)
|
Преобла-дающие
ветры, направление
|
|
средние
|
абс.
|
|
|
|
|
|
|
max
|
min
|
|
|
|
|
|
Январь
|
-14,5
|
4
|
-38,7
|
25
|
39
|
71
|
ЮЗ
|
|
Февраль
|
-11,1
|
12
|
-34
|
17
|
50
|
70
|
ЮЗ
|
|
Март
|
-4,1
|
17
|
-28
|
21
|
45
|
58
|
Ю
|
|
Апрель
|
3,1
|
27
|
-17
|
19
|
35
|
32
|
СЗ
|
|
Май
|
12
|
33,6
|
-7
|
48
|
-
|
37
|
ЮЗ
|
|
Июнь
|
16,9
|
38
|
-1,4
|
65
|
-
|
57
|
З
|
|
Июль
|
19,2
|
36
|
2
|
63
|
-
|
64
|
З
|
|
Август
|
15,9
|
34
|
0
|
112
|
-
|
77
|
ЮЗ
|
|
Сентябрь
|
9,5
|
31,1
|
-2,5
|
49
|
-
|
59
|
ЮЗ
|
|
Октябрь
|
3,7
|
21
|
-13
|
38
|
5
|
65
|
З
|
|
Ноябрь
|
-6,8
|
12,9
|
-36
|
35
|
15
|
77
|
З
|
|
Декабрь
|
-11,1
|
6,1
|
-35
|
34
|
26
|
77
|
ЮЗ
|
|
Год
|
2,7
|
38
|
-38,7
|
526
|
-
|
62
|
ЮЗ
|
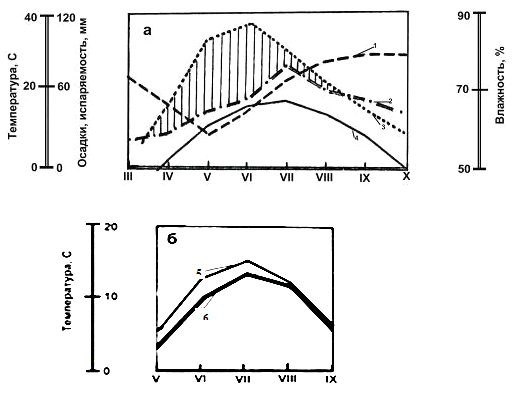
Рис. 4. Климат (а) и температура почвы (б) в
Припышминском лесном массиве (по Н. С. Санниковой, 1992): 1 - относительная
влажность воздуха, 2 - осадки, 3 - испаряемость, 4 - температура воздуха, 5 -
температура почвы на глубине 5 см, 6 - то же на глубине 20 см.
Для сравнения условного баланса влаги определен
гидротермический коэффициент Селянинова за 2008 год по данным Талицкого
метеопоста по формуле:
= УР : (0,1 х Уtа), (1.2.1)
где К - гидротермический коэффициент Селянинова;
УР - месячная сумма осадков;
Уtа - сумма активных температур за месяц.
Май: К = 49,4 : (0,1 х 282,1) = 1,8;
Июнь: К = 36,9 : (0,1 х 474) = 0,8;
Июль: К = 147,6 : (0,1 х 635,5) = 2,3;
Август: К = 76,8 : (0,1 х 502,2) = 1,5;
Сентябрь: К = 31,7 : (0,1 х 140,4) = 2,3.
Исходя из полученных гидротермических
коэффициентов, можно сделать вывод, что июль и сентябрь - с избыточным
увлажнением (К более 2), май и август - с достаточным (К от 1 до 2), а июнь -
засушливый (К менее 1).
.3 Рельеф и гидрология
Юго-восточная часть Свердловской области
расположена во внеледниковой части Западно-Сибирской низменности. Четвертичные
отложения представлены здесь аллювиальными, озерно-аллювиальными и
делювиальными отложениями. В приречных зонах почвы формируются на аллювии и
делювии третичных отложений - опок и опоковидных глин и суглинков, трепелов,
песчаников и т.д. [Фирсова В. П., 1969].
По С. Н. Санникову (1961), Припышминские боры
находятся на западной окраине Западно-Сибирской равнины вблизи ее перехода в
эрозионно-абразионную платформу Зауралья. Конфигурация массива довольно строго
повторяет очертания геоморфологического района надпойменных террас древней
долины р. Пышма. В восточной же части массива встречаются участки с
дюнно-бугристым рельефом. Высота бугров, которым приписывается эоловое
происхождение, достигает 10-15 м. Вблизи современной долины р. Пышма подобие
ландшафта дюн и песчаных бугров создают многочисленные ложбины стока, имеющие
местами характер древних оврагов, расчленивших надпойменные террасы на отдельные
отроги и островки. Однако уже на расстоянии 3-5 км от русла реки расчлененность
террас слабеет, и рельеф становится однообразно плоскоувалистым. Современный
рельеф Припышминских надпойменных террас, в основном выполненный уровнем
вторых террас, в целом равнинный. Поверхность террас представляет
ориентированные в направлении течения пра-Пышмы цепи и гряды невысоких (от 5-8
до 10-12 м) широких увалов со слабо выпуклыми, иногда почти плоскими, вершинами
и пологими склонами. Увалы, вытянутые с запада на восток, разделены широкими и
плоскодонными древними ложбинами стока, а в поперечном направлении расчленены
притоками р. Пышма. На поверхности увалов обычны многочисленные мелкие
западины, террасы и ложбины, а замкнутые суффозионные котловины заняты верховыми
болотами, реже озерами [Санников С. Н., 1961].
Гидрологические особенности территории парка
тесно связаны с местонахождением двух его дач - Талицкой и Тугулымской (прил.
1).
В районе парка р. Пышма представляет собой
типичную для Зауралья реку и ограничивает парк с севера. Река отличается
довольно энергичным половодьем. Высокий паводок отмечается через 10-12 лет.
Наибольший среднесуточный расход воды 261 м3/с, процент стока за
половодье составляет 57% годового. Летом средняя температура воды в реке: июнь
+18,6˚С, июль +21,6˚С, август +18,3˚С.
По данным А. И. Мошкина в районе Талицкой дачи
притоками р. Пышма являются небольшие речки - Урга, Ретин, Молоюр и др. На р.
Урга, в южных окрестностях находится Ургинский пруд (площадь 93 га), являющийся
государственным гидрологическим памятником природы областного значения. В
территорию национального парка не входит.
Гидрологические условия Тугулымской дачи
значительно отличаются от Талицкой дачи. Этот район приурочен к водораздельным
пространствам правого притока р. Тура - Липки и реки Пышма. Река Липка берет
начало из озера Гурино в западной части Тугулымской дачи и впадает в Туру ниже
с. Липчинского, она ограничивает парк с севера. В эту реку с территории дачи
впадают небольшие притоки Малая и Курейка, их истоки находятся в центре
Бахметского болотного массива. В юго-западной части Тугулымской дачи протекает
р. Айба - левый приток Пышмы.
Большой интерес представляет собой озеро Гурино
- самый большой водоем национального парка. Его площадь 210 га, отметка воды
93,6 м. Само озеро проточное, с юга в него впадает небольшой водоисточник, а на
севере находится исток реки Липка.
.4 Почвенные условия
Почвы коренных Припышминских боров относятся, в
основном, к подзолистому типу, что свидетельствует о том, что они формировались
под влиянием лесной растительности. Под воздействием травянистой
растительности, внедряющейся под полог хвойных лесов в результате рубок, частых
пожаров и бокового осветления, в последние десятилетия усиливается дерновый
процесс, и формируются дерново-подзолистые почвы [Чудников П. И., 1930;
Зубарева Р. С., 1960; Санников С. Н., 1961; Фирсова В. П., 1969; Санников С.
Н., Санникова Н. С., 1985].
Данные анализа механического состава
дерново-сильноподзолистых почв Припышминских боров приведены в таблице 1.4.1.
Таблица 1.4.1 - Механический состав
дерново-сильноподзолистой почвы в сосняке-черничнике (%) (по В. П. Фирсовой,
1969)
|
Глубина
взятия образца (см)
|
Генетический
горизонт
|
Содержание
частиц, диаметр (мм)
|
|
|
1-0,25
|
0,25-0,05
|
0,05-0,01
|
0,01-0,005
|
0,005-0,001
|
<0,001
|
<0,01
|
|
5-8
|
А1
|
17,16
|
25,25
|
30,52
|
10,44
|
7,42
|
9,51
|
27,07
|
|
10-25
|
А2
|
21,57
|
27,65
|
30,13
|
8,18
|
7,06
|
5,41
|
20,65
|
|
30-40
|
В1
|
2,84
|
24,24
|
24,03
|
6,37
|
9,10
|
33,42
|
38,89
|
|
105-120
|
С
|
0,09
|
30,75
|
32,33
|
8,03
|
8,56
|
20,24
|
36,83
|
По исследованиям С.Н. Санникова (1961), Н. С.
Санниковой (1984) на вершинах и инсолируемых склонах бугров, наиболее широко
распространенных в восточной части массива (особенно в Тугулымской даче), под
пологом лишайниковых и бруснично-лишайниковых сосняков преобладают песчаные
сильно дренированные сухие и суховатые, бедные гумусом и илом, почвы
подзолистого типа. Химический состав тех же почв под сосняком-черничником
приведен в таблице 1.4.2.
Таблица 1.4.2 -Химический состав
дерново-сильноподзолистых почв Припышминских боров (по Р. С. Зубаревой, 1960),
тип леса - сосняк-черничник
|
Глубина
взятия образца (см)
|
Генетический
горизонт
|
Гумус
(%)
|
рН
солевой
|
Обменная
кислотность, по Соколову
|
Гидролитическая
кислотность
|
Сумма
поглощенных оснований по Кеппену
|
|
|
|
|
Н+
|
Al+++
|
H++Al3+
|
|
|
|
|
|
|
Мгэкв
на 100 г почвы
|
|
5-8
|
А1
|
3,55
|
5,2
|
0,123
|
0,227
|
0,350
|
5,20
|
6,60
|
|
10-25
|
А2
|
0,29
|
4,4
|
0,031
|
0,124
|
0,155
|
1,48
|
2,28
|
|
35-50
|
В1
|
Не
опр.
|
4,8
|
0,082
|
0,206
|
0,288
|
2,80
|
15,10
|
|
60-80
|
В2
|
Не
опр.
|
4,8
|
0,082
|
0,387
|
0,463
|
2,97
|
13,40
|
|
100-120
|
С
|
Не
опр.
|
5,0
|
0,072
|
0,381
|
0,453
|
2,31
|
9,62
|
|
|
|
|
|
|
|
|
|
|
Вершины невысоких песчаных увалов заняты
сосняками бруснично-зеленомошными и экологически близкими к ним сосняками
бруснично-вересково-зеленомошными. Почвы под ними слабоподзолистые суховатые,
относительно бедные элементами минерального питания растений. Грунтовые воды на
глубине более 2,5-3,5 м. На пологих склонах увалов под сосняками
бруснично-чернично-зеленомошными располагаются почвы
слабодерново-среднеподзолистые. И, наконец, на нижних частях склонов и обширных
плоских вершинах увалов под сосняками и ельниками чернично-зеленомошными почвы
дерново-средне- и сильноподзолистые. Верхний песчано-супесчаный слой их на
небольшой глубине (30-50 см) подстилается суглинком или глиной. В производных
сосняках злаково-мелкотравных и березовых лесах, а также в сосняках травяных
обычны типичные дерново-подзолистые почвы с мощностью гумусового горизонта до
7-8 см [Санникова Н. С., 1984].
Особое место занимают почвы под сосняками
травяными (точнее «мелкотравные» с доминированием «борового мелкотравья», реже
«разнотравья»), производными от коренных сосняков-черничников после сплошных
рубок. Приурочены они к дренированным ровным участкам, к склонам низких холмов,
реже к западинам и ложбинам на междуречьях. Производительность древостоев этого
типа леса I, реже II
бонитета [Санников С. Н., 1962]. Почвы под такими лесами иногда испытывают
влияние карбонатов, либо развиваются при близком подстилании глинистых наносов
[Фирсова В. П., 1969] обычно на границах с междуречными покровными суглинками,
но не в типичных борах на песчаных глубоких террасах. Все это создает
предпосылки для лучшего разложения органического вещества, закрепления
оснований и обеспечения растений влагой. Фотографии и описания почвенных
разрезов даны в приложении 2.
.5 Лесная растительность
По мнению С.Н. Санникова (1962, 1966),
особенности состава растительности Припышминского лесного массива и окружающих
территорий определяются его положением в подзоне предлесостепных
сосново-березовых лесов. Коренные «зональные» типы растительности на серых
лесных почвах междуречий к северу и югу от Припышминских боров представлены
березовыми и осиново-березовыми злаково-разнотравными лесами с элементами
таежного мелкотравья. В понижениях рельефа здесь расположены заболоченные
коренные березняки осоково-вейниковые. На надпойменных террасах речек, левых
притоков р. Пышма, изредка встречаются сосново-березовые и елово-березовые
мелкотравно-зеленомошные леса на дерново-подзолистых и торфянисто-подзолистых
почвах.
Типы сосновых лесов Припышминских боров изучены
достаточно полно и лучше, чем какого-либо другого лесного массива в Зауралье
[Чудников П.И., 1930, Зубарева Р.С., 1960; Картавенко Н.Т., 1960; Санников
С.Н., 1961, 1962, 1992].
Ввиду слабой расчлененности рельефа
типологический состав Припышминских боров не отличается разнообразием. Для
сосновых лесов его можно представить в виде следующей классификационной схемы
[Санников С.Н., 1962]:
А. Сосняки лишайниковые:
сосняк лишайниковый (IV
бонитета).
Б. Сосняки-зеленомошники:
сосняк бруснично-вересковый (III,
5 бонитета),
сосняк-брусничник (III
бонитета),
сосняк бруснично-черничный (I-II
бонитета),
сосняк черничник (II
бонитета).
В. Сосняки травяные:
сосняк злаково-мелкотравный (II
бонитета),
сосняк разнотравный (I
бонитета).
Г. Сосняки сложные:
сосняк с липовым подлеском (I-Iа
бонитета) встречается только в Трошковском лесничестве национального парка
(Тугулымский район).
Д. Сосняки болотно-травяные:
сосняк осоково-травяной (III-IV
бонитета).
Е. Сосняки сфагновые:
сосняк сфагновый (Vа-Vб
бонитета).
Как показали исследования С.Н. Санникова (1962),
С.Н. Санникова и Н.С. Санниковой (1985), в собственно Припышминском массиве
коренная растительность носит бореальный облик. В моховом ярусе сосняков и
ельников-зеленомошников, не расстроенных рубками и пожарами, доминируют зеленые
мхи, а в травяно-кустарничковом ярусе характерны брусника (Vaccinium
vitis-idaea
L.), черника (Vaccinium
myrtillus
L.), багульник
болотный (Ledum
palustre
L.), плаун годичный
(Lycopodium
annotinum
L.) и другие. В
составе растительности Припышминских боров преобладают сосновые леса (43%
покрытой лесом площади). На долю ельников приходится всего 2-4% площади. В
коренных типах сосновых лесов Припышминского массива преобладают сосняки
бруснично-черничные (33%), злаково-мелкотравные (27%) и черничники (20%). На
долю сосняков-брусничников приходится 13%. Незначительную часть занимают
сосняки кустарничково-сфагновые (4%), осоково-травяные (около 1%) и
бруснично-вересковые. Доли процента приходятся на сосняки
бруснично-лишайниковые, лишайниковые и сосняки с липой, встречающиеся только в
Тугулымской даче парка. В общем, в составе растительности Припышминских боров
преобладают сосновые леса. На долю ельников, сохранившихся лишь в пониженных
местообитаниях, редко затрагиваемых пожарами и не затрагиваемых рубками,
приходится всего 1,7% площади. Распределение площади сосняков Припышминского
массива по типам леса (данные лесоустройства) отражено на рис. 5.
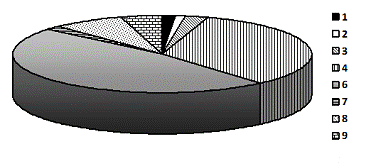
Рис. 5. Распределение площади сосняков
национального парка «Припышминские боры»: 1 - бруснично-лишайниковый; 2 -
бруснично-вересковый; 3 - брусничник; 4 - бруснично-черничный; 5 - черничник; 6
- злаково-мелкотравный; 7 - сложный (с липой); 8 - осоково-травяной; 9 -
кустарничково-сфагновый
Ниже приводится краткая
лесоводственно-экологическая характеристика наиболее распространенных и
хозяйственно-важных типов леса: сосняков лишайниковых, брусничников,
бруснично-черничных, черничников и сосняков разнотравных по С. Н. Санникову
(1962), С. Н. Санникову и Н. С. Санниковой (1985).
Сосняк лишайниковый. Занимает вершины и верхние
части склонов на высоких надпойменных террасах. Почвы подзолистые, песчаные,
сухие и бедные, на глубоких дренированных песках. Грунтовые воды на глубине
25-30 м, верховодок в ризосфере деревьев нет. Состав древостоя: 10С, полнота
0,5-0,6. Подлесок отсутствует. Подрост: группами угнетенная сосна. Напочвенный
покров лишайниково-брусничниковый, пятнами мхи (Pleurozium
schreberi (Brid.)
Mitt.) и лишайники.
Подлеска нет. В травяно-кустарничковом подъярусе встречаются прострел желтеющий
(Pulsatilla
flavescens (L.)
Mill.), гвоздика
песчаная (Dianthus
arenarius
L.), брусника (Vaccinium
vitis-idaea
L.), вейник наземный
(Calamagrostis
epigeios (L.)
Roth.), овсяница
луговая (Festuca
pratensis
Huds.).
Сосняк-брусничник. Широко распространен в районе
р. Беляковка (около 30% общей площади боров). В западной части массива
сосняки-брусничники встречаются реже (10-12%). Занимает вершины дренированных
увалов и верхние части их склонов. Почвы глубокие, суховатые, песчаные,
подзолистые со слабой верховодкой. Древостой сосновый с участием березы (Betula
pendula Roth.),
бонитет III, полнота
0,6-0,8. Подлесок - единично шиповник иглистый (Rosa
acicularis
Lindl.), ракитник
русский (Chamaecytisus
ruthenicus (Fisch.
ex Moloszcz.)
Klaskova). Подрост густой,
группами. Напочвенный покров зеленомошно-брусничниковый: мхи (Pleurozium
schreberi (Brid.)
Mitt., редко Hylocomium
splendens (Hedw.)
Schimp.), брусника (Vaccinium
vitis-idaea
L.) 10-15 см
высотой, вейник наземный (Calamagrostis
epigeios (L.)
Roth.), кошачья лапка
двудомная (Antennaria
dioica (L.)
Gaerth.), прострел
желтеющий (Pulsatilla
flavescens (L.)
Mill.), зимолюбка
зонтичная (Chimaphila
umbellata (L.)
W. Barton.),
золотарник обыкновенный (Solidago
virgaurea
L.), ястребинка
зонтичная (Hieracium
umbellatum
L.), хвощ зимующий (Equisetum
hyemale L.),
изредка багульник (Ledum
palustre
L.). Подстилка
хвойно-моховая, грубогумусная, неразложившаяся; мощность ее 2,0-2,5 см.
Сосняк бруснично-черничный (ягодниковый).
Наиболее распространенный тип леса Припышминских боров, занимающий в них около
30-35% площади. Занимает пологие склоны, слабовыпуклые возвышенности с
супесчано-суглинистыми, подзолистыми почвами. Древостой сосновый с участием
березы (Betula
pendula Roth.),
единично осины (Populus
tremula L.)
и ели (Picea
obovata Ledeb.).
Возобновление сосной (Pinus
sylvestris
L.), реже - береза (Betula
pendula Roth.)
и ель (Picea
obovata Ledeb.),
приурочено к просветам в пологе. Подлесок редкий - можжевельник обыкновенный (Juniperus
communis L.), шиповник
иглистый (Rosa
acicularis
Lindl.), рябина
обыкновенная (Sorbus
aucuparia
L.). Травяной покров
состоит из вейника тростниковидного (Calamagrostis
arundinacea (L.)
Roth.), костяники
каменистой (Rubus
saxatilis
L.), герани лесной (Geranium
sylvaticum
L.), вероники
дубравной (Veronica
chamaedrys L.)
и др.
Производным типом биогеоценоза от сосняка
бруснично-черничного иногда после сплошной рубки является березняк
вейниково-мелкотравный, а вариантами - сосняк с можжевельником (Juniperus
communis
L.), сосняк
зеленомошно-мелкотравный и сосняк ягодниково-вейниково-мелкотравный.
Сосняк-черничник. Довольно распространенный тип
леса, занимающий около 20% общей площади Припышминских боров. Занимает плоские
ровные поверхности и нижние части склонов увалов и надпойменных террас. Почвы
слабо-дерново-подзолистые супесчаные, влажные; весной обильная верховодка.
Древостой 9С1Б - 8С2Б ед. Е, Пх, бонитет I-II,
5, полнота 0,6-0,8. В подлеске единично рябина (Sorbus
aucuparia
L.), ива козья (Salix
caprea L.),
шиповник (Rosa
acicularis
Lindl.). По наблюдениям
С. Н. Санникова (устное сообщение), вблизи населенных пунктов, особенно там,
где был интенсивный выпас, под пологом спелых и перестойных
сосняков-черничников встречается наиболее обильный подлесок можжевельника.
Подрост редкий, напочвенный покров зеленомошно-чернично-мелкотравный: пятнами
кукушкин лен обыкновенный (Polytrichum
commune Hedw.),
черника обыкновенная (Vaccinium
myrtillus
L.) до 40 см, вейник
наземный (Calamagrostis
epigeios (L.),
брусника обыкновенная (Vaccinium
vitis-idaea
L.) 20-25 см
высотой, ожика волосистая (Luzula
pilosa (L.)
Willd.), орляк
обыкновенный (Pteridium
aquilinum (L.)
Kuhn.), грушанка
круглолистная (Pyrola
rotundifolia
L.), плаун годичный
(Lycopodium
annotinum
L.). Подстилка 4-5
см, грубогумусная, неразложившаяся.
Производными типами леса от сосняков черничных
являются березняк злаково-разнотравный (II
бонитета), сосняк чернично-молиниевый и сосняк чернично-злаково-мелкотравный.
Сосняк разнотравный. Является довольно распространенным
типом леса, на долю которого приходится 10-15% общей площади сосняков. Занимает
пологие нижние части склонов с дерново-подзолистыми, свежими, периодически
влажными почвами. Древостой сосновый с примесью березы (Betula
pendula Roth.)
и осины (Populus
tremula L.).
Возобновление березой повислой (Betula
pendula Roth.),
редко сосной обыкновенной (Pinus
sylvestris
L.), осиной
обыкновенной (Populus
tremula L.).
Подлесок редкий - смородина красная (Ribes
rubrum L.),
малина обыкновенная (Rubus
idaeus L.),
ива пепельная (Salix
cinerea L.),
калина обыкновенная (Viburnum
opulus L.),
шиповник иглистый (Rosa
acicularis
Lindl.), шиповник
майский (Rosa
maialis Herrm.),
рябина обыкновенная (Sorbus
aucuparia
L.), черемуха
обыкновенная (Padus
avia Mill.).
Глава 2. Состояние проблемы
.1 Ареал и экоареал можжевельника обыкновенного
Первые находки семейства кипарисовых приурочены
к нижнеюрским отложениям [Решетняк Т.А., 1980]. Предковым родом этого семейства
считают Protocupressiolon
Eckhold. Родоначальником
современных можжевельников был род Juniperoxylon,
широко распространенный в засушливых горных областях северного полушария от
триаса до середины мелового периода (ныне вымерший).
Эволюция Juniperus
communis
L. проходила в
условиях достаточно влажного климата плиоцен-плейстоцена [Барзут О.С., 2007].
Местом возникновения предковых форм Juniperus
communis
L. считают Среднюю
Европу, что подтверждают палеоботанические находки на территории Польши
[Жеронкина Т.А., 1976].
Современный ареал рода Juniperus
L. простирается от
70˚с.ш. до 12˚ю.ш., охватывая все северное полушарие. Основными
центрами видового и формового разнообразия можжевельников в настоящее время
являются Тихоокеанское побережье юга Северной Америки (около 20 видов),
Средиземноморье (8 видов) и Центральная Азия (около 15 видов) [Барзут О.С.,
2007; Кожевников А.П., Тишкина Е.А., 2011б].
Вообще, можжевельник обыкновенный встречается
почти на всех типах почв лесной зоны, их механического состава и режима
увлажнения: от подзолистых до болотных, от песчаных до тяжелосуглинистых, от
сухих до избыточно-увлажненных. Многие авторы [Каппер О.Г., 1954; Мухамедшин
К.Д., 1970; Мухамедшин К. Д., Таланцев Н.К., 1982; Поплавская Л. Ф., 1982;
Мухамедшин К.Д., Сартыбаев С.К., 1988; Аши М., 1991б; Тимофеев В.В., Лантратова
А.С., Самодурова Н.С., 2001; Кожевников А.П., Тишкина Е.А., 2009] экологическим
оптимумом для можжевельника обыкновенного считают сосняки- и
ельники-черничники, обладающие повышенной трофностью почвы и обеспечивающие
растениям оптимальный световой режим, не опускающийся ниже предельного
светового барьера (7-8% от полной освещенности открытого места или не выше
0,7-0,8 единиц полноты).
По данным Е.А. Тишкиной (2009) было установлено,
что данный вид встречается в качестве подлеска в 54 типах леса, что указывает
на его большую экологическую амплитуду. Спектр типов сосновых лесов с
можжевельником шире, чем в ельниках с его участием. Разрушение экологических
связей в нарушенных биогеоценозах приводит к невозможности нормального
семенного размножения вида в пределах ареала. Находясь преимущественно в
подлеске сосняков, можжевельник мигрирует на осветленные места после рубок,
пожаров, участвует в заселении свободных участков по окраинам болот.
.2 Морфоэкологические особенности
Можжевельник обыкновенный - деревце или
кустарник, чаще высотой до 3-5 м, со сбежистым стволом, покрытым сверху
серо-бурой волокнистой корой. Молодые трехгранные побеги сначала зеленые, позже
- каштаново-коричневые. Широкая морфогенетическая изменчивость углов отхождения
ветвей от ствола способствует формированию различных типов кроны [Барзут О. С.,
2007; Кожевников А.П., Тишкина Е. А., 2011б]. Игловидные листья - колючие,
линейно-ланцетные, с низбегающим основанием, отстоящие, жесткие, длиной от 6 до
20 мм, шириной 0,7-1,5 мм, в трехчленных мутовках. На верхней стороне хвоинки
имеется неглубокий желобок с широкой белой устьичной полоской с восковым
налетом; нижняя сторона выпуклая, с тупым килем.
Древесина можжевельника с бурым ядром и
желтоватой узкой заболонью, отличается особой стойкостью против гниения,
тяжелая, обладает повышенной прочностью. Годичные слои узкие, сердцевинные лучи
не видны, смоляные ходы в древесине отсутствуют [Каппер О.Г., 1954; Мамаев
С.А., 1968б, 1972].
Характеризуя можжевельник обыкновенный, многие
авторы отмечают светолюбие, засухоустойчивость, морозостойкость,
нетребовательность этого вида к почвенным условиям [Каппер О.Г., 1954;
Мухамедшин К.Д., 1970; Мухамедшин К.Д., Таланцев Н.К., 1982; Поплавская Л.Ф.,
1982; Мухамедшин К.Д., Сартыбаев С.К., 1988; Аши М., 1991б; Тимофеев В.В.,
Лантратова А.С., Самодурова Н.С., 2001]. Однако из-за низкой
конкурентоспособности он вынужден занимать участки, свободные от других
растений: бедные песчаные, каменистые и известняковые почвы. Их корни,
внедряясь в грунт и распластываясь под землей во все стороны на десятки метров,
способны добывать воду и питательные вещества из самых бедных почв. К наиболее
заметным морфобиологическим особенностям всех можжевельников относится также
крайне медленный рост даже в оптимальных условиях существования. Можжевельники
отличаются долголетием [Денисова Г.А., Пилипенко Ф.С., 1978].
Утолщенный эпидермис хвои и глубоко погруженные
устьица - признаки ксерофитных свойств растения [Каппер О.Г., 1954].
Интенсивность транспирации листьев можжевельника в 8 раз меньше, чем у сосны
обыкновенной [Поплавская Л.Ф., 1982; Антонова Е.В., 1995]. Тем не менее, М. Аши
(1991б) подчеркивает влаголюбие можжевельника обыкновенного и чувствительность
к континентальности климата.
В статье Н.А. Михеевой (2005) охарактеризован
кариотип можжевельника обыкновенного из южной тайги Западной Сибири. До сих пор
нет единого мнения по поводу подвидовой таксономической подразделенности
можжевельника обыкновенного; ряд его форм описан как отдельные виды. Больше
всего спорных вопросов у отечественных исследователей вызывает таксономическое
положение двух видов: J.
communis и J.
sibirica. Некоторые авторы
считают J. sibirica
угнетенными особями J. communis
[Мамаев С.А., 1968б, 1969], другие утверждают, что J.
sibirica (=subsp.
J. alpina)
является горной вариацией J.
communis
L., или его экотипом
[Князева С.Г., 2000а; Коропачинский И.Ю., Встовская Т.Н., 2002]. В ряде работ
последних лет [Князева С.Г., 2000а; Ермолина П.И., 2002] можжевельник сибирский
рассматривается как подвид можжевельника обыкновенного.
Н.А. Михеевой (2005) были исследованы растения,
произрастающие в суходольных сосновых лесах Тимирязевского лесхоза Томской
области. В популяции можжевельник был представлен разнообразными жизненными
формами: колонновидное дерево, кустарник с пучковатой и узкоколонновидной
кроной, куртина с шаровидной или полушаровидной кроной.
Кариограмма можжевельника в исследованной
популяции состоит из 22 хромосом (2n
= 22). Полиплоидных особей не обнаружено. Результаты исследования показали, что
морфологические типы хромосом можжевельника обыкновенного однообразны.
Можжевельник обыкновенный встречается в обоих
полушариях: начиная с подлеска хвойных и лиственных лесов, тундровых редколесий
и заканчивая скальными породами в высокогорьях, что вызвало образование разных
жизненных форм. Это и форма высокоствольного дерева, и многоствольное дерево, и
разные формы кустов, и форма стланика. Это говорит о большой пластичности и
приспособляемости можжевеловых. Такого формового разнообразия не наблюдается у
других представителей семейства кипарисовых [Джанаева В.М., 1969; Хантемирова
Е.В., Семериков В.Л., 2009; Кожевников А.П., Тишкина Е.А., 2011б].
Juniperus
communis
L. var.
communis распространен в
подлеске хвойных и лиственных лесов, на сухих холмах и по сухим горным склонам,
на известняковых берегах рек, реже на моховых болотах.
Характерная отличительная черта этого вида -
прямая форма одно- или многоствольного деревца. Распространена и форма куста.
Кроны этих форм в свою очередь тоже изменчивы [Косицын В. Н., 1999а; Харламова
С. В., 1999; Михеева Н.А., Муратова Е.Н., Ефремов С.П., 2005]. Наряду с
габитусом один из главных морфологических признаков можжевельника обыкновенного
- длинная прямая отстоящая хвоя.
В тундре, у верхней границы леса и в горах
можжевельник образует стланиковые формы. В современной литературе стланик
считают разновидностью можжевельника обыкновенного и называют Juniperus
communis
L. var.
saxatilis
Pall. [Adams
R.P.,
Pandey R.N.,
2003]. Существует множество синонимов J.
c. var.
saxatilis
Pall. в ранге формы,
подвида, разновидности и вида (sibirica,
alpina, nana,
pigmaea, montana).
О. Г. Каппер (1954) считал его формой можжевельника обыкновенного (J.
communis
f. sibirica
Rydb.). Отдельной
формой он выделял J. c.
nana Loud.,
растущий в горах и арктических областях. В то же время во многих сводках и
определителях он считается самостоятельным видом J.
sibirica
Burgsd [Ареалы…, 1977;
Деревья …, 1949; Горчаковский П.Л., Шурова Е.А., Князев М. С., 1994].
И.Ю. Коропачинский и Т.Н. Встовская (2002) считают,
что в Сибири типичный J.
communis растет только в
Томской области, а на остальной части Сибири растет J.
sibirica - вид, никогда не
образующий форму деревца. Однако, в Припышминских борах, несмотря на то, что
основную часть популяций можжевельника составляют кустовидные и стланиковые
формы, часто встречаются и экземпляры древовидной формы.
Можжевельник обыкновенный - медленнорастущий
вид, поэтому при интенсивном ведении хозяйства человеком в хорошо освоенных
лесах очень редко встречаются экземпляры, достигшие яруса древостоя.
Исследования в Белоруссии [Поплавская Л.Ф., 1982], на Дальнем Востоке
[Кожевникова З.В., 1987, 1988], в Верхневолжье [Аши М., 1991б] указывают на
среднюю высоту растений можжевельника, не превышающую двух-трех метров, и
среднегодовой линейный прирост 5-10 см. При этом авторами акцентируется
внимание на приуроченность высокоствольных древовидных форм можжевельника к
хорошо дренированным богатым почвам, которые характерны для приручейных и
прирусловых типов лесов, окраин верховых болот, закустаренных пастбищ на
торфяно-глеевых почвах. Габитус можжевельника отличается как от условий
местопроизрастания, так и в пределах выделяемых в них экоморф [Князева С.Г.,
2000а, 2000б; Михеева Н.А., 2002а, 2002б]. В долинах рек у можжевельников формируется
вертикальный, высотой более 1 м ствол, а в тундре в прибрежной зоне Белого моря
растения приобретают низкорослую стелющуюся форму.
С.Г. Князева (2000а) на основании анализа
изменчивости морфологических признаков можжевельников (длины и ширины хвои,
размеров шишкоягод), произрастающих в Сибири, выделила три разновидности
можжевельника обыкновенного - J.
communis
L. var.
communis - равнинная
разновидность, J. communis
L. var.
sibirica (Burgsd.)
Rydb. - плоскогорная
разновидность, и J. communis
L. var.
nana (Willd.)
Baumg. - высокогорная
разновидность [Хантемирова Е.В., Семериков В.Л., 2009].
А.И. Колесниковым (1974) предложена
классификация форм можжевельника обыкновенного. По характеру роста он выделяет:
Juniperus
communis
f. hibernica
Gord. (исландская,
пирамидальная); Juniperus
communis
f.
suecica
Loud.
Ait.
(шведская); J. communis
f. cracovica
Gord. (краковицкая);
J. communis f. pendula Carr. (плакучая);
J. communis f. ehiniformis Beissn. (ежевидная);
J. communis f. weckii Graeb. (Векки);
J.
communis
f.
compressa
Carr.
(сжатая);
J. communis f. depressa Pursh., Fom (J. communis canadensis Loud., J.
canadensis Burgsdorf, J. nana montana Endl.) (плоская);
J. communis f. prostrata Beissn. (распростертая);
J. communis f. hemisphaerica Parl.; J. communis f. nana Loud. (карликовая);
J. communis f. sibirica Rydb. (сибирский);
J. communis f. nord-china (северо-китайский).
По
окраске
хвои
Juniperus communis f. aurea Nochols. (золотистая);
J. communis f. aurea-spicata Rehd., aurea-variegata Hort (золотистоконечная);
J. communis f. aurea Hort. По
форме
шишкоягод
Juniperus communis f. oblonga Medw; J. communis f. globosa Medw.; J. communis
f. ovata Medw.; J. communis f. thyocarpos Asch.
Изучением форм можжевельников обыкновенного и
сибирского занимались на Тянь-Шане К.Д. Мухамедшин (1970), К.Д. Мухамедшин и
С.К. Сартыбаев (1988), В Г. Рубаник и З.И. Паршина (1975) в Казахстане; Л.Ф.
Поплавская (1982) в Белоруссии, Г.В. Вишняков (1987) в Мурманской области; С.
В. Харламова (1999) в Республике Марий Эл; В.Н. Косицын (1999, 2001) в Тверской
области; С.Г. Князева (2002) в Красноярском крае; Н.А. Михеева (2005), Е.В.
Хантемирова и В.Л. Семериков (2009), А.П. Кожевников и Е. А. Тишкина (2009,
2011) на Урале и в Сибири; О. С. Барзут (2007) в Архангельской области. В результате
проведенных исследований указанных авторов обнаружена существенная зависимость
морфологических форм можжевельника от условий природной среды.
При выделении древовидных высокоствольных,
кустовидных многоствольных, мелких кустов, стлаников и других жизненных форм
растений в основе лежит изучение системы формирования побега [Барыкина Р.П.,
Кудряшов Л.В., Класова А.И., 1963; Воробьева Т.И., 1975].
Исследователями можжевельники единогласно
заносятся в список долговечных видов, представляющих значительные перспективы
для дендрохронологических и дендроклиматологических исследований [Баранникова
Е.Е., Жмакина З.Ю., 1999; Хантемиров Р.М., Шиятов С.Г., Горланова Л.А., 1999].
Продолжительность жизни отдельных растений, по разным данным, достигает от 200
до 2000 лет и более [Каппер О.Г., 1954, Гроздов Б.П., 1960]. Естественно, что
наибольшего возраста можжевельники достигают в ненарушенных девственных лесах.
Долголетие можжевельников сильно варьирует, изменяясь в зависимости от
индивидуальных особенностей и экологических условий. Антропогенная и
хозяйственная деятельность человека, низовые пожары оказывают сильное влияние
на возрастную структуру популяций можжевельника обыкновенного, в целом занижая
ее средний возраст [Аши М., 1991б]. Повсеместная разновозрастность, как
можжевелового подлеска, так и древостоя, представленного можжевельниками,
выявлена в различных лесорастительных условиях [Аши М., 1991б; Garcia
D., Zamora
R., Hodar
J. A.,
Gomez J.
M., 1999].
Некоторыми исследователями отмечено негативное
влияние бессистемных рубок, лесных пожаров и нерегулируемого выпаса скота на
произрастание можжевельников в различных географических широтах [Коннов А.А.,
1963; Коваль И.П., 1968; Владыченский А.С., Ульянова Т.Ю., Баландин С.А.,
Козлов И.Н., 1994]. При относительно низкой промышленной ценности можжевеловые
леса выполняют важную водорегулирующую, водоохранную, почвозащитную и
противоселевую роль. В то же время в ряде работ современных авторов (в основном
зарубежных) разрабатываются способы выжигания и методы применения гербицидов
для борьбы с древесно-кустарниковой растительностью (в том числе и с
можжевельниками) при формировании желательного ботанического состава кормовых
угодий [Bunting
S. C.,
1994]. Зарастание луго-степных пастбищ можжевельником рассматривается как
сукцессионно-дигрессивный процесс, требующий предотвращения [Кумаритаев Ф.С.,
Киркосян К.Г., Габараева И.И., 1998]. Для борьбы с зарослями можжевельника в
США используют бобров. Такая практика, видимо, связана со спецификой
исторически сложившегося стиля хозяйствования, в настоящее время
ориентированного на скорейшее получение продукции, что в свою очередь требует
увеличения пастбищных площадей [Барзут О. С., 2007].
Можжевельник обыкновенный выступает как
подлесочный вид в составе сосновых и еловых лесов на Европейском Севере,
темнохвойной тайги в Сибири и на Востоке России, на западе России в лесах из
дуба, граба, бука. М. Аши (1991а) считает, что включение в состав соснового
древостоя ели и березы улучшает продуктивность можжевельника, приближая его к
экологическому оптимуму. Наиболее выражены биоценозы можжевельника с лесными
растениями (брусникой, черникой, ландышем), высшими грибами (масленком, белым
грибом) [Барзут О.С., 2007].
В процессе взаимодействия компонентов лесного
биоценоза осуществляется их взаимное влияние друг на друга [Поплавская Л. Ф.,
1982]. На участках с можжевельником в течение дня остается высокой
относительная влажность воздуха и повышена влажность почвы, что способствует
естественному возобновлению самого можжевельника и главных лесообразующих
видов.
Несомненно, древостой конкурирует с
можжевельником, оказывает влияние на его численность, рост и жизненность.
Многие авторы [Каппер О.Г., 1954; Поплавская Л.Ф., 1982; Аши М., 1991б; Барзут
О.С., 2007; Кожевников А.П., Тишкина Е.А., 2011б] экологический оптимум для
можжевельника определяют сосняками-черничниками со световым режимом не выше 0,7
- 0,8 единиц полноты. Повсеместная разновозрастность можжевелового подлеска
выявлена в различных лесорастительных условиях [Аши М., 1991а]. Распределение
растений по классам возраста или по возрастным состояниям зависит от типа и
возраста главного древостоя, а также степени его нарушенности. Крайне мало
работ в этом направлении по можжевельнику. Эта проблема успешно решается для
подроста хвойных [Санникова Н.С., 1979, 1984, 2003; Санникова Н.С., Локосова
Е.И., 2001], но остается практически неизученной по отношению к можжевельнику
обыкновенному.
.3 Размножение и естественное возобновление
Можжевельник обыкновенный - двудомное растение.
Его микроспорофиллы собраны в мелкие микростробилы, которые закладываются в
предыдущий перед опылением год в пазухах листьев на прошлогодних побегах или на
концах боковых веточек. На верхней части микроспорофиллы несут по 2-6
микроспорангиев, содержащих одноклеточную пыльцу (25,5±0,03 мкм) без воздушных
мешков [Барзут О.С., 2007; Кожевников А.П., Тишкина Е.А., 2011б]. В таком
состоянии микростробилы зимуют. При наступлении устойчивых положительных
температур (около +3˚С) начинается последующее развитие генеративных органов.
У мужских шишек удлиняется ось, обнажаются пыльники, и происходит осыпание
пыльцы, сначала с нижних микроспорофиллов, а затем - с верхних. Процесс пыления
продолжается до 7-10 дней. Женские шишки (мегастробилы) закладываются осенью на
пазушных укороченных побегах. Следующей весной мегастробилы развиваются очень
быстро. Они состоят из 3-8 перекрестно- или по три расположенных в мутовках
чешуи, над которыми возвышаются 1-3 семяпочки с микроскопическим отверстием
наверху (микропиле). В период опыления из этого отверстия выступает капелька
клейкой жидкости, которая не только удерживает пыльцу на семяпочке, но и
увлажняет ее, что способствует развитию пыльцевой трубки. Чешуи женских шишек
перед опылением расходятся, семяпочки обнажаются, становясь доступными для
попадания пыльцевых зерен. После оплодотворения чешуи мегаспорофиллов быстро
разрастаются, становятся мясистыми, затем срастаются между собой, образуя
сочную шишку (главный отличительный признак можжевельников). Шишки
можжевельников созревают либо в первый год, либо на второй или даже на третий
год после опыления.
Погодные условия сказываются на семеношении
можжевельников. Вымываемая дождем пыльца резко сокращает число оплодотворенных
мегастробилов, чрезмерно сухая погода высушивает гуттационной секрет на
микропиле [Барзут О.С., 2007], а понижение температуры до 12-13˚С
приостанавливает развитие пыльцевой трубки. Оптимум опыления приходится на
теплую (23-24˚С) ветреную погоду.
Уже к концу первого года шишки достигают своих
окончательных размеров, но все еще остаются зелеными. К концу лета второго года
они становятся мягкими, сине-черными с сизоватым восковым налетом. Обычно на
одной и той же особи можжевельника зеленые шишки первого года соседствуют со
зрелыми двулетними.
У J.
communis
L. 1-3 семени в
шишкоягоде, редко - 4. Созревание семян заканчивается к осени второго года.
Зрелые семена коричневые, твердые, легко отделяющиеся от смолистой мякоти
шишки. Анатомо-морфологическое созревание семян можжевельников наступает на 2-3
месяца раньше шишек, в которые они заключены [Кожевникова З. В., 1988; Иглина
П.И., 1998]. К моменту созревания шишек семенная кожура уплотняется и это
приводит к наступлению глубокого физиологического покоя, задерживая прорастание
семян на два, три и даже на четыре года. В результате опытов по проращиванию
семян из зрелых и зеленых шишек О. С. Барзут (2007) особой разницы в сроках и
проценте всхожести не заметила. Осыпание созревших шишек начинается с конца
лета и продолжается до глубокой осени.
Наиболее раннее семеношение у можжевельника
обыкновенного по литературным данным отмечено в пятилетнем возрасте у
окультуренных растений. В естественных условиях возмужалость наступает чаще
после 10 лет и определяется условиями местопроизрастания. Наилучший урожай
шишек получается со средневозрастных деревьев. Урожайные годы чередуются с
неурожайными с периодичностью 3-5 лет, что связано с трехлетним циклом развития
шишек. Выход семян из шишек по массе варьирует от 15 до 70% [Кожевникова З.В.,
1987; Барзут О.С., 2007]. Полнозернистость семян отмечена от 23 до 72%, причем
в малоурожайные годы она снижается, а в годы обильных урожаев повышается. Масса
одной шишки и урожайность изменяются в зависимости от экологических условий
[Олешко Г.И., Донцов А.А., 1991; Косицын В.Н., 2001]. Семена можжевельников
покрыты прочной и крепкой оболочкой, надежно защищающей зародыш внутри семени.
Проглоченное животным семя беспрепятственно проходит сквозь их пищеварительный
тракт, оставаясь неповрежденным, способным к прорастанию [Барзут О.С., 2007;
Кожевников А.П., Тишкина Е.А., 2011б]. Судя по тому, что шишкоягодами
можжевельника питаются многие виды птиц и мышевидных грызунов, вероятно, они и
являются главными агентами распространения семян можжевельника. По
предположению С.Н. Санникова (устное сообщение), еще одним агентом
распространения можжевельника могут выступать копытные млекопитающие - коровы,
овцы, лоси, косули, нередко объедающие побеги можжевельника и могущие разносить
семена на своих копытах. Выявление этих факторов - ключевые вопросы изучения проблемы
экологии естественного возобновления и расселения популяций можжевельников.
Принято считать, что можжевельник растет
медленно, однако, исследования Д.Н. Данилова (1941), А.П. Шиманюка и В.А. Жанет
(1958), Л.Ф. Поплавской (1982), М. Аши (1991б), D.C.
Chojnacky (1996) показали,
что образующая ствола можжевельника обыкновенного подчиняется общим
закономерностям, присущим древесным породам. В течение жизненного цикла
растений наблюдаются периоды усиления текущего прироста по высоте (до 50 лет),
затем их постепенное снижение. Улучшение условий местопроизрастания
положительно сказывается на интенсивности хода роста, вызывая всплески
приростов даже в более зрелом возрасте. По данным Л. Ф. Поплавской (1982) рубки
ухода не оказывают существенного влияния на изменение показателя роста
можжевелового подлеска. Механическое уничтожение растений при рубках леса,
нарушение мохового покрова лесозаготовительными машинами и, как следствие,
плохое прорастание семян, являются отрицательными факторами распространения можжевельника.
В условиях культуры скорость роста побегов
можжевельника увеличивается более чем в 2 раза по сравнению с самосевом
[Кожевникова З.В., 1988; Кожевников А.П., Тишкина Е.А., 2011б]. Интенсивно
семеносящие женские экземпляры испытывают большие затраты пластических веществ
на формирование семян и, поэтому, отстают в росте от мужских растений этого же
возраста.
Размножение можжевельника обыкновенного в
естественных условиях происходит как семенным, так и вегетативным путем.
Указания разных авторов о преобладании одного из них можно объяснить различными
экологическими условиями исследуемых участков. З. В. Кожевникова (1988)
отмечает в природных ценозах преобладание семенного размножения. Тем не менее,
вегетативное размножение характерно для влажных типов лесов, где происходит
укоренение приземных ветвей под благоприятным моховым покровом [Аши М., 1991б].
Длительный период покоя семян, особые условия их прорастания, значительный
процент пустозернистых семян, зараженность их вредителями создают трудности при
семенном размножении и в природной среде, и при искусственном выращивании
сеянцев.
Искусственное разведение можжевельников возможно
семенами, черенками, отводками и прививкой. Последний способ пока не имеет
широкого использования, но представляет собой перспективное направление
исследований [Барзут О.С., 2007]. Эффективность различных способов и приемов
выращивания сеянцев можжевельника обыкновенного испытывали О.Т. Истратова
(1968), Т.А. Жеронкина и В.Г. Рубаник (1976), Л.Ф. Поплавская (1982), З. В. Кожевникова
(1987), О.С. Барзут (2007); А.П. Кожевников и Е.А. Тишкина (2011б).
Наиболее целесообразным в ранних работах был
признан посев свежесобранными семенами из незрелых буроватых шишек [Жеронкина
Т.А., Рубаник В.Г., 1976; Поплавская Л.Ф., 1982], при этом всхожесть достигает
90%. Опыты О. С. Барзут (2007) не показали каких-либо различий в посевах
свежесобранными семенами из зеленых шишек и семенами после стратификации из
созревших шишек. Всхожесть и время до появления всходов были примерно
одинаковыми. Отмечено положительное действие замачивания семян в хвойном
экстракте сосны: 10 грамм хвои на 1 литр воды [Поплавская Л.Ф., 1982].
Для нарушения состояния покоя семян, полученных
из зрелых синих шишкоягод, были предложены и термическая обработка, и длительная
стратификация. Чаще других способов предлагается воздействие на семена в
течение одного месяца высокими температурами (от +24 до +27°С) и далее низкими
(от +1 до +5°С) в течение трех месяцев [Жеронкина Т.А., Рубаник В.Г., 1976].
Иногда авторы рекомендуют только холодную
стратификацию. Во всех случаях посева семенами отмечена очень низкая их
всхожесть: от 0 до 35% [Жеронкина Т.А., Рубаник В.Г., 1976; Бакланова Е.Г.,
1988]. Воздействие раствором KMnO4,
концентрированной серной кислотой, кипятком, стимуляторами роста, гибберелином
на прорастание семян не оказывают положительного влияния. Всходы появляются
через 2-3 года так же, как и при посеве шишкоягодами, а всхожесть не превышает
3-5% [Барзут О.С., 2007].
Оптимальная норма высева семян для можжевельника
обыкновенного составляет 50 штук на 1 погонный метр [Поплавская Л. Ф., 1982].
Глубина заделки семян - 1-2 см. Посев в гряды с междустрочным расстоянием 10 см
обеспечивает нормальное развитие сеянцев на протяжении двух лет, при условии
содержания гряд во влажном состоянии. Возможен посев в открытый грунт и с
междустрочным расстоянием 20 см и глубиной заделки семян 1-1,5 см [Барзут О.С.,
2007]. От пересыхания, вымывания дождем и поливами рекомендуется мульчирование
посевов торфом или опилками слоем 2-3 см [Поплавская Л.Ф., 1982]. Прорастание
семян и начало роста корней происходит уже при температуре 0°С, тогда как
развитие надземной части растений запаздывает на 1-1,5 месяца. Искусственно
выращенные сеянцы имеют более развитую корневую систему и надземную часть по
сравнению с сеянцами, выросшими в природных условиях [Барзут О.С., 2007;
Кожевников А.П., Тишкина Е.А., 2011б].
Учитывая трудности семенного возобновления
можжевельников, предпочтение отдается вегетативному способу размножения.
Использование его позволяет получить укорененные черенки достаточно хорошего
качества более быстрым путем. Кроме того, указанный посадочный материал
наследует признаки маточного растения, отличается более высокими
биометрическими показателями надземной и подземной части, по сравнению с
сеянцами [Кожевникова З.В., 1987; Кожевников А.П., Тишкина Е.А., 2011б].
Наряду с березой, осиной и сосной можжевельник
составляет группу растений, в первую очередь поедаемых копытными [Русанов Н.
С., Сорокина Л.И., 1984]. В годы истощения кормовой базы лосей, косуль, кабанов
наблюдается уничтожение ими побегов можжевельника от 60 до 100%. В Верхневолжье
зарегистрировано до 1% экземпляров можжевельника, объеденных зайцем-беляком
[Аши М., 1991б].
В ювенильном возрасте у можжевельников
наблюдается полегание сеянцев (Fusarium
трех видов: discolor,
elegans, martiella),
источник заражения - почва или шишкоягоды [Чуб А.В., 1980].
Шютте можжевеловое (Laphodermium
juniperinum
De Not.)
появляется в начале лета на прошлогодней хвое [Галасьева Т.В., Соколова Э.С.,
1989]. Бурое шютте, или бурая снежная плесень (Herpotrichia
nigra Hart.)
встречается в горных и северных районах на ели, сосне, кедре и можжевельнике
[Соколова Э.С., Семенкова И.Г., 1981].
Д.Б. Беломесяцева (2003) провела изучение
видового состава грибов, входящих в консорцию можжевельника обыкновенного.
Всего выявлено 206 видов, относящихся к 133 родам, 49 семействам, 22 порядкам,
5 классам.
Возбудителями гнилей стволов и ветвей
можжевельника являются трутовик настоящий (Fomes
fomentarius (L.:
Fr.) Fr.)
и, реже, стереум шерстистый (Stereum
hirsutrum
Willd.) [Гаршина Т.Д.,
1968; Ролл-Хансен Ф., Ролл-Хансен Х.,. 1998].
Множество грибов вызывают у можжевельников
корневую гниль: (Phacolus
phacolus), трутовик
Швейница (Polyporus
schweinitzii) [Журавлев И. И.,
Селиванова Т.Н., Черемисинов Н.А., 1979].
Кроме выше перечисленных болезней, которым
подвергаются можжевельники, существует немалое количество насекомых-вредителей,
повреждающих разные части растений: Megastigmus
bipunctatus
Swed., Megastigmus
fidus Nic.
[Тропин И.В., Ведерников Н.М., Крангауз Р.А., 1980; Долгий М.М., 1993]. Галлы
можжевельника [Гусев В.И., Римский-Корсаков М.Н., 1951] возникают в результате
повреждения веретеновидной можжевельниковой галлицей (Oligotrophus
panteli Kieff.)
и обыкновенной можжевельниковой галлицей (Oligotrophus
juniperinus
L.).
Древесина повреждается можжевельниковым лубоедом
(Phloeosinus
bicolor Brulle),
древесным толстощупиком (Serropalpus
barbatus
Schall). Гусеницы
плодовой можжевельниковой моли (Argyresthia
praecocella
Zell.) и
можжевельниковый ягодный клещик (Eriophyes
quadrisetus
Tomas.) наносят вред
шишкоягодам можжевельников [Корнилов В. П., 1990].
Достаточно внушительный перечень возможных
болезней и потенциальных вредителей можжевельников наряду с отсутствием сведений
о патологическом состоянии естественных можжевеловых насаждений на территории
России позволяет говорить о совершенной неизученности этого вопроса и в
Свердловской области.
Глобальным катастрофическим фактором
трансформации структуры, функций и динамики лесного покрова являются стихийные
и антропогенные пожары [Санников С.Н., 1973, 1992; Санникова Н.С., 1978, 1984].
В настоящее время более или менее широко изучено
и освещено в литературе влияние пожаров на факторы местообитания (почвенные,
микроклиматические и др.) и компоненты лесных фитоценозов
(травяно-кустарничковый ярус, подрост и древостой главных хвойных видов в
Западной Сибири [Санников С.Н., 1992; Санников С.Н., Санникова Н.С., Петрова
И.В., 2012].
К настоящему времени по естественному послепожарному
возобновлению видов хвойных (сем. Pinaceae)
в лесах Западной Сибири накопилось несколько сотен опубликованных работ. Между
тем, данные по естественному возобновлению можжевельника обыкновенного на гарях
почти отсутствуют. Стимулирующее влияние устойчивых пожаров, выжигающих верхний
слой грубого гумуса, на естественное возобновление сосны обыкновенной и других
хвойных видов установлено во многих частях их ареала [Санников С.Н., Санникова
Н.С., Петрова И.В., 2012].
По С.Н. Санникову (1973), стимулирующее влияние
пожаров на естественное возобновление сосны обыкновенной связано со следующими
пирогенными факторами: удаление верхнего слоя гумуса и мохового покрова;
повышение режима увлажнения субстрата для прорастания семян, улучшение
теплообеспеченности минеральных горизонтов; перевод части зольных элементов и
азота из труднодоступных корням самосева форм в легкоусвояемые минеральные
формы; смещение pH
обожженного субстрата из пессимальной области в более благоприятные пределы;
активизация микрофлоры, ускоряющей разложение органики и улучшающей условия
минерального питания самосева; уничтожение или подавление конкурирующих
растений нижних ярусов (видимо, в том числе и можжевельника обыкновенного).
По мнению Е.А. Тишкиной (2009), популяции
можжевельника сохраняются в составе лесных фитоценозов во многом благодаря
распространению его семян птицами и использованию фактора «фитоценотической
защиты». Разнообразие форм можжевельника, его возрастной спектр, характер
расселения под пологом средневозрастного и спелого древостоя являются
показателями стабильности этого вида. Нарушают эту стабильность на особо
охраняемых природных территориях (ООПТ), таких как национальный парк
«Припышминские боры», только пожары, после которых восстановление популяций
можжевельника происходит крайне медленно.
Леса таежной зоны издавна подвергаются более или
менее частому воздействию лесных пожаров, которые являются непреходящим
эволюционно-экологическим фактором [Санников С.Н., 1973, 1985; Санников С.Н.,
Санникова Н.С., 1985]. В отличие от других агентов среды, изменяющихся более
или менее постепенно и ритмично, огонь действует на лес внезапно,
кратковременно и крайне интенсивно как «катастрофа» для всего сообщества,
вызывая глубокое и длительное прямое или косвенное преобразование всех
компонентов биогеоценоза. Под влиянием огня уменьшается кислотность почв,
увеличивается содержание оснований, подвижных форм фосфора и калия, усиливается
интенсивность микробиологических процессов, уменьшается количество
органического вещества и возрастает содержание воднорастворимых веществ
[Санников С. Н., 1992].
Влияние пожаров на структуру биогеоценозов и
условия среды в сосняках-зеленомошниках предлесостепного Припышминского лесного
массива разносторонне и детально изучено на Талицком стационаре Института
экологии растений и животных УНЦ АН СССР [Санников С. Н., 1973, 1992; Санников
С.Н., Санникова Н.С., 1985; Санникова Н.С., 1978, 1984, 1992]. Установлено, что
пожар любой интенсивности уничтожает лишайниковый, зеленомошный покров и
надземные части травяно-кустарничкового яруса, способствуя вегетативному
возобновлению и росту популяций относительно пожароустойчивых «лугово-лесных»
трав, а также пирогенных политриховых мхов, бокальчатых и трубчатых кладоний
[Санников С.Н., 1973, 1992].
Влияние пожаров на ценопопуляции можжевельника
обыкновенного существенно иное.
.4 Хозяйственное использование
Нерегулируемое всестороннее использование
можжевельника на хозяйственные нужды населением, возможно, явилось причиной
сокращения можжевеловых зарослей и, в частности, крупных древовидных
экземпляров этого растения.
Крестьяне России и скандинавских стран
заготавливали можжевеловые колья и жерди, используя их для изгородей, мастерили
игрушки и детские кроватки. Прочная, но легко обрабатываемая древесина, шла на изготовление
клюк, клюшек для хоккея, посохов, курительных трубок, клепок для бочек,
кухонной утвари. Имеются сведения о применении можжевеловых корней и ветвей в
кораблестроении. Детали корабля, «сшитые» с их помощью отличались особой
долговечностью и прочностью крепления. Еще в начале XX века можжевеловым
лапником покрывали крыши своих построек жители Швеции и Норвегии [Мамаев С.А.,
Петухова И.П., 1965]. Благодаря гибкости и прочности ветки можжевельника
используются рыболовами на самодельные удилища. В ряде северных стран древний
промысел - плетение корзин из можжевеловых ветвей.
Издавна используемая в строительстве, мебельном
и карандашном производстве, древесина можжевельника теперь не имеет
промышленного значения. Из-за преобладания низкорослых экземпляров ее
используют лишь для изготовления токарных и резных изделий. Добытая из корней и
ветвей можжевельника смола перерабатывается в белый лак, так называемый
немецкий сандарак [Каппер О.Г., 1954; Денисова Г.А., Пилипенко Ф.С., 1978].
Шишкоягоды можжевельника сладковато-пряные,
ароматные, содержат до 20 - 30% сахаров (фруктозы, глюкозы, сахарозы), по
сахаристости не уступают винограду. Их широко используют в пищевой
промышленности. Отмечено стимулирующее действие можжевельника на «созревание»
соленой рыбы с одной стороны, и с другой - затормаживание процесса окисления
жиров [Денисова Г.А., Пилипенко Ф. С., 1978].
Фармацевтическое и медицинское применение
можжевельника обусловлено наличием эфирного масла [Денисова Г.А., Пилипенко
Ф.С., 1978]. Можжевеловое масло токсически влияет на рост грибов и бактерий, в
частности, на Aspergillus
niger L.,
Bacterium
coli штамма 113-3 и Staphylococcus
aureus [Поплавская Л.Ф.,
1982]. В корнях можжевельника также найдены эфирные масла, смолы, сапонины,
таннин, воск, дубильные, красящие и гуммозные (содержащие камедь) вещества,
пектины. Хвоя содержит до 0,27% витамина С, а пик содержания в ней эфирных
масел приходится на летний период [Денисова Г.А., Пилипенко Ф.С., 1978;
Антонова Е.В., 1995].
Лечебные свойства растения описаны еще Вергилием
в I веке до нашей эры, когда во время эпидемий холеры жилища окуривали дымом от
тлеющих ветвей можжевельника [Тульчинская В.П., Юргелайтис Н.Г., 1987]. В
современной медицине наибольший эффект достигнут от использования шишкоягод как
мочегонного, дезинфицирующего мочевыводящие пути, противовоспалительного,
отхаркивающего, желчегонного, обезболивающего, жаропонижающего и улучшающего
пищеварение средства. Витаминные чаи из можжевеловой хвои показаны при
аллергии, простудных заболеваниях, авитаминозах. При туберкулезе, бронхите,
язвенной болезни желудка, кожных заболеваниях используются корни можжевельника.
Фитованны из частей растения рекомендованы как успокаивающее и очищающее кожу
средство. Из хвои некоторых можжевельников получены вещества (лигнаны),
обладающие противоопухолевой активностью. Однако шишки можжевельника
противопоказаны при острых заболеваниях почек, язвенной болезни желудка и
двенадцатиперстной кишки, острых гастритах и колитах [Мухамедшин К.Д., 1970;
Денисова Г.А., Пилипенко Ф.С., 1978; Барзут О.С., 2007].
Обестерпененное эфирное масло вводят в
парфюмерные отдушки для пудры, мыла, шампуней, кремов [Барзут О.С., 2007].
Ресурсы семенной продукции. Шишкоягоды
можжевельника представляют интерес в нескольких ракурсах: а) как естественный
источник самоподдержания популяции; б) как сырье в пищевой промышленности и
медицине; в) как кормовая база лесных зверей и птиц; г) как семенной материал
для целей искусственного разведения.
О наличии сырьевой базы для заготовок шишкоягод
можжевельника обыкновенного можно судить по величине площади зарослей данного
растения. Продуктивность зарослей во многом зависит от типа местопроизрастания.
Уточнение данных о наличии и величине зарослей сырьевых растений, оценка их
продуктивности позволяют осуществить предварительный подсчет эксплуатационных
ресурсов. Минимальные размеры зарослей можжевельника, подлежащих учету в Литве
- 0,02 га, при минимальном проективном покрытии можжевельника 20% [Будрюнене Д.
К., 1979]. Продуктивными считают заросли можжевельника при относительной
полноте древостоя до 0,5. В свою очередь, согласно проективному покрытию их
делят на три категории: заросли I
категории - проективное покрытие составляет 5-10%, пригодны для рекреации;
заросли II категории -
проективное покрытие - 11-30%, они могут быть объектом, как для рекреации, так
и для промысловых заготовок; в III
категорию выделяют самые густые (проективное покрытие более 30%) и самые ценные
заросли, на которые необходимо обращать внимание при планировании хозяйственных
мероприятий. Резервными заросли определяют при полноте древостоя от 0,6 до 1,0,
при изменившихся условиях (улучшении освещения) они могут перейти в категорию
продуктивных. В Свердловской области к промысловым зарослям относят заросли,
эксплуатационный запас сырья в которых не менее 50 кг/га, при этом
эксплуатационный запас рассчитывается в объеме 90% от «биологического объема»
[Олешко Г.И., Донцов А.А., 1991]. По мнению В.Н. Косицина (1999),
эксплуатационный запас необходимо рассчитывать в размере 75% от «биологического
объема».
Урожайность шишкоягод можжевельника колеблется в
очень широких пределах, изменяясь в зависимости от экологических условий.
Авторы, изучавшие этот вопрос, указывают на значительные запасы можжевельника,
произрастающего на открытых участках: от 13 до 960 кг/га воздушно-сухих плодов
в Пермской области [Олешко Г.И., Просовский М.А., Белоногова В.Д., 1987]; от 22
до 170 кг/га воздушно-сухих плодов в Свердловской области [Олешко Г.И., Донцов
А.А., 1991]; от 150 до 160 кг/га воздушно-сухих плодов в Кировской области
[Киселева Т.М., Шабалина И.А., 1996]. В условиях покрытых лесом земель семенная
продуктивность можжевелового подлеска снижается, составляя, например, в лесных
насаждениях Литвы - от 6 до 12 кг/га [Будрюнене Д.К., 1979]; в Белоруссии до 34
кг/га [Поплавская Л.Ф., 1982].
Установлено, что можжевеловые насаждения
выделяют в 15 раз больше фитонцидов, чем лиственные породы и в 6 раз больше,
чем другие хвойные породы [Денисова Г.А., Пилипенко Ф.С., 1978; Поплавская
Л.Ф., 1982]. Можжевельник обыкновенный отнесен к I классу зимостойкости,
устойчивость против неблагоприятных факторов городской среды средняя, имеет
высокие декоративные качества, хорошо переносит стрижку, отличается
долговечностью и медленным ростом. В зеленом строительстве вид рекомендован для
создания посадок на газонах, для групп и солитеров перед зданиями, для
декоративных живых изгородей, выполняющих защитную роль [Рубаник В.Г., Паршина
З.И., 1975; Денисова Г.А., Пилипенко Ф.С., 1978; Мухамедшин К.Д., Сартыбаев
С.К., 1988; Кожевников А.П., Тишкина Е.А., 2011б]. Однако, несмотря на
естественный обширный ареал, растение не нашло широкого применения в озеленении
северных городов и населенных мест России. Так считал Орлов Ф.Б. (1962), такое
же положение в этом вопросе сохранилось и до настоящего времени.
Неоднозначность мнений по поводу устойчивости
можжевельника обыкновенного к рекреационным нагрузкам и атмосферному
загрязнению можно объяснить слабой изученностью этого вопроса. Учитывая
санитарно-гигиеническое значение можжевеловых насаждений, их высокую
сохранность на третьей и даже четвертой стадиях дигрессии, ряд авторов [Антипов
В. Г., 1979; Поплавская Л.Ф., 1982] отмечают можжевельник обыкновенный как
газоустойчивый вид и рекомендуют его к использованию в зеленом строительстве.
В более поздних работах [Бакланова Е.Г., 1988;
Аши М., 1991б] отмечается, что можжевельник обыкновенный в сильной степени
подвергается воздействию загрязнений в воздушной среде, вызывающему
суховершинность растений на расстоянии от 0,5 до 2 км от загрязнителя и
культивирование его возможно в относительно экологически чистых районах
городов.
Можжевельник выполняет почвозащитную,
водорегулирующую и противоселевую функции. Установлены его почвоулучшающие
свойства в результате повышения биологической активности почвы [Поплавская Л.
Ф., 1982].
Научное значение растения в реконструкции
климатов прошлого подтверждено его использованием в споропыльцевом анализе и
дендрохронологических исследованиях [Баранникова Е.Е., Жмакина З.Ю., 1999].
Глава 3. Объекты и методы исследований
.1 Объекты изучения
Объектами исследований являлись естественные
ценопопуляции можжевельника обыкновенного, произрастающего в преобладающих
типах сосновых лесов национального парка «Припышминские боры» - в черничнике,
бруснично-черничном и злаково-мелкотравном (далее в тексте - «сосняк
травяной»). Предполагая, что в этих типах сосняков формируются оптимальные
условия для произрастания можжевелового подлеска, подбирались средневозрастные
древостои со средней полнотой (0,6-0,7), но варьирующей на пробной площади, что
позволяет выявить влияние изменений структуры и конкуренции древостоя на
структуру, рост и жизненность можжевельника. Лесоводственно-таксационная
характеристика пробных площадей приводится в таблице 3.1.1.
Нами принята следующая схема соподчиненных
единиц классификации биотопов (по С.Н. Санникову, 1992): тип лесного массива
(сосновые леса на песчаных надпойменных террасах) - тип леса, представленный
коренным или условно-коренным типом биогеоценоза - тип биотопа (участки леса
под пологом древостоя, гари или сплошные вырубки в пределах одного типа
биогеоценоза).
Под типом леса в соответствии с представлениями
генетической (динамической) школы лесной типологии [Колесников Б.П., 1956;
Санников С.Н., 1992] мы понимаем серию (ряд) лесных биогеоценозов,
характеризующихся общностью условий местопроизрастания, главной лесообразующей
и сопутствующих древесных пород, а также состава нижних ярусов фитоценоза и
представляющих различные стадии восстановительно-возрастных смен лесных
биогеоценозов в пределах одного типа лесорастительных условий.
Таблица 3.1.1 - Лесоводственно-таксационная
характеристика пробных площадей
|
Лесничество,
№ квартала /выдел Тип леса: cосняки
|
Древостой
|
Относительная
полнота
|
Полнота
(м2/га)
|
Запас
(м3/га)
|
Почва
|
|
состав
|
Возраст
(лет)
|
Высота
(м)
|
Диаметр
(см)
|
|
|
|
|
|
Ургинское
л-во, 16/25 бруснично-черничный
|
10С*
|
70
|
23
|
24
|
0,7
|
172,6
|
390
|
Дерново-подзолистая
песчаная на супеси, подстилаемой средним суглинком
|
|
Ургинское
л-во, 16/27 бруснично-черничный
|
10С
|
60
|
21
|
22
|
0,8
|
122,9
|
380
|
Дерново-подзолистая
среднесуглинистая средней супеси, подстилаемой суглинком
|
|
Ургинское
л-во, 16/28 бруснично-черничный
|
10С
|
65
|
21
|
24
|
0,5
|
118,2
|
235
|
Дерново-подзолистая
среднесупесчаная на супеси с линзами галечника, подстилаемой средним
суглинком
|
|
Ургинское
л-во, 16/29 бруснично-черничный
|
10С
|
75
|
24
|
26
|
0,8
|
152,8
|
210
|
Дерново-подзолистая
среднесуглинистая на супеси, подстилаемой средним суглинком
|
|
Ургинское
л-во, 26/1 травяной
|
10С+Б
|
150
|
28
|
44
|
0,6
|
85,4
|
340
|
Дерново-подзолистая
среднесуглинистая на супеси, подстилаемой тяжелым суглинком
|
|
Ургинское
л-во, 26/18 травяной
|
6Б
3С 1Ос
|
60
|
23
24 23
|
20
24 22
|
0,7
|
130,6
|
272
|
Дерново-подзолистая
среднесуглинистая на супеси, подстилаемой тяжелым суглинком
|
|
Ургинское
л-во, 26/19 травяной
|
7Б
2Ос 1С
|
50
|
18
19 21
|
16
22 24
|
0,8
|
172,6
|
240
|
Дерново-подзолистая
среднесуглинистая на супеси, подстилаемой тяжелым суглинком
|
|
Талицкое
л-во, 37/6 черничник
|
7С
3Б
|
120
75
|
27
25
|
28
24
|
0,8
|
199,4
|
468
|
Дерново-подзол
средне-мощный иллювиально-железистый, песчаный на песке
|
|
Талицкое
л-во, 37/7 черничник
|
9С
1Б
|
120
75
|
28
25
|
30
24
|
0,8
|
212,7
|
513
|
Дерново-подзол
средне-мощный иллювиально-железистый, песчаный на песке
|
|
Талицкое
л-во, 37/13 черничник
|
10С
|
85
|
27
|
24
|
0,8
|
196,5
|
486
|
Дерново-подзол
средне-мощный иллювиально-железистый, песчаный на песке
|
*Примечание: С - сосна обыкновенная, Б - береза
повислая
.2 Принципы и методы исследования
В соответствии с целью работы в исследованиях
применялись следующие основные методические принципы популяционных
лесоэкологических исследований [Санников С.Н., Санникова Н.С., 1985]:
1. Лесотиполого-экологический -
сравнительное количественное изучение параметров среды, структуры и
возобновления ценопопуляций можжевельника обыкновенного на вырубках и гарях в
различных типах леса.
2. Микроэкосистемный - сопряженный
количественный анализ хорологических изменений структуры и функций
можжевельника в зависимости от структуры и функций древостоя-эдификатора
[Санникова Н.С., 1992].
Ниже рассматриваются методы исследований в плане
основных вопросов программы.
Измерение фотосинтетически активной радиации
солнца (ФАР) проводилось на всех трех пробных площадях (ПП) и 600 учетных
площадках (УП) в пасмурный день (облачность 10 баллов) в 12-14 часов с помощью
люксметра Ю-16 на высоте 1,3 м. Интенсивность относительной ФАР выражалась в
процентах от освещенности (в люксах) на ближайшем открытом месте.
Рост и морфогенез особей. Полевые исследования
проводились маршрутным методом с использованием методов, принятых в
лесоведении, лесной таксации и лесоустройстве с учетом рекомендаций по изучению
годичного прироста [Сукачев В.Н., 1938; Сукачев В.Н., Зонн С.В., Мотовилов
Г.П., 1957; Сукачев В.Н., Зонн С.В., 1961; Программа…, 1966]. На каждой из трех
наиболее репрезентативных основных пробных площадей закладка учетных лент
(трансект) проводилась вдоль визиров шириной 10 м. Длина ленты устанавливалась
с таким расчетом, чтобы на ней было в среднем не менее 100 экземпляров
можжевельника обыкновенного. Таким образом, площадь учетной ленты на пробной
площади составила 2000 м2. Вдоль натянутой мерной ленты производился
учет всех экземпляров можжевельника, произрастающих в этом условно ограниченном
пространстве, определялись проективное покрытие крон, биометрические показатели
и другие характеристики растения. Измеряли диаметр кроны куста, и на основании
данных по сумме проекций крон всех экземпляров можжевельника и площади учетной
трансекты определялось среднее проективное покрытие можжевельника на пробной
площади. У каждого отдельно растущего экземпляра измерялись прирост осевого
побега можжевельника за три последних года, фиксировалась жизненная форма
растения: куст, дерево, стланик. Отмечалась форма ствола: прямой, слегка
изогнут в виде латинской буквы «L»,
изогнут в виде русской буквы «С», изогнут в виде латинской буквы «S»,
«V»-образный [Барзут
О.С., 2007].
По жизнеспособности различались здоровые,
слабо-, средне- и сильно угнетенные, а также механически поврежденные, больные
(усыхающие) и мертвые экземпляры. Признаками здоровых растений, как и у
подроста сосны, считались прогрессирующий или, по крайней мере, стабилизированный
прирост осевого побега можжевельника длиной не менее 5% от его общей высоты,
нормально развитая зеленая хвоя, отсутствие существенных повреждений грибами и
насекомыми [Санникова Н.С., 1984].
Жизненность популяций можжевеловых насаждений
определялась как отношение среднего прироста осевого побега куста за три
последних года к общей высоте куста.
Возраст экземпляров можжевельника определялся
путем подсчета числа годичных колец на поперечном срезе базальной части ствола
на уровне гипокотиля на отшлифованной поверхности спила под микроскопом МБС-2.
Учет особей можжевельника проводился по ступеням высоты с градацией 0,5 м:
растения с высотой до 50 см, от 51 до 100 см, от 101 до 150 см, от 151 до 200
см, от 201 до 250 см и более 250 см. Средний возраст растений для каждой
ступени высоты определялся по среднему количеству годичных слоев на спилах,
взятых у шейки корня с трех-пяти модельных экземпляров.
Оценка урожайности можжевельника осуществлялась
по результатам сбора шишкоягод с трех модельных ветвей, расположенных в
верхней, средней и нижней частях кроны женского растения [Корчагин А.А., 1960;
Олешко Г.И., Донцов А.А., 1991], подсчетом общего количества плодоносящих
ветвей и умножением на него среднего числа и массы шишкоягод с модельной ветви
(в расчете на один куст).
Микроэкосистемный анализ связей с
древостоем-эдификатором. Принятый нами ценопопуляционно-микроэкосистемный
подход [Санникова Н.С., 1992] требует сопряженного изучения изменений
показателей популяции можжевельника, окружающего древостоя (в зоне его влияния)
и важнейших факторов внешней среды: жизнеспособности и роста можжевеловых
насаждений в зависимости от размещения и роста окружающего древостоя, а также
от состояния субстрата, толщины подстилки, проективного покрытия, освещенности
и других факторов.
Сопряженный анализ структуры древостоя,
можжевелового подлеска и факторов среды выполнялся на 100 круговых учетных
площадках в каждом типе леса: сосняках черничнике, бруснично-черничном и
травяном. Проведен сплошной перечет деревьев, и измерялось расстояние их от
куста можжевельника, который находился на круговой учетной площадке радиусом,
равным максимальной длине главных латеральных корней 60-80-летних деревьев
сосны (в среднем - 10 м), с измерением диаметра каждого дерева на высоте 1,3 м.
Таким радиусом охватываются все деревья, корни которых могут конкурировать с
корнями изучаемого куста можжевельника, находящегося в центре круга.