Особенности биологии гольяна, обитающего в бассейне реки Кубань
Выпускная
квалификационная работа
Особенности биологии
гольяна, обитающего в бассейне реки Кубань
Введение
Реки Кубани обладают богатой, как по численности, так и по
разнообразию видового состава ихтиофауной, которая представляет значительный
интерес в научном плане. Существует множество проблем связанных с размерами
популяций (возрастной, половой), их составом, численностью, изучением
воздействия абиотических и биотических факторов на популяции. Одной из таких
проблем является уточнение систематики гольяна из бассейна реки Кубань.
Основным моментом при использовании любой методологической
концепции, как на видовом, так и на надвидовом уровне остается возможность
диагностировать таксон. Возможность диагностировать - это логическое требование
для отличия любой группы организмов от всех других групп. Поэтому базой любых
таксономических исследований остается изучение многообразия признаков (и
состояний признаков) любого свойства - от традиционных внешнеморфологических
признаков до особенностей физиологии, поведения и строения ДНК.
Основоположником зоологической системы является шведский
ученый естествоиспытатель Карл Линней. В основу систематики рыб в нашей стране
положена система Л.С. Берга. Однако в последние годы в эту систему внесены
некоторые изменения и дополнения.
В систематике животных существует определенная иерархия
систематических категорий. (Г.А.МОСКУЛРЫБЫ ВОДОЕМОВ БАССЕЙНА КУБАНИ
(ОПРЕДЕЛИТЕЛЬ)). Виды объединяют в роды, роды - в семейства, семейства - в
отряды, отряды - в классы, а классы - в типы. Но не всегда эти основные
систематические группы объединяют все органические формы, существующие в
природе. Поэтому применяют некоторые промежуточные категории (подтип, надкласс,
подкласс, надотряд, подотряд, надсемейство, подсемейство, подрод, подвид).
Исходя из этого полная современная иерархия систематических групп выглядит
следующим образом: тип ® подтип ® надкласс ® класс ® подкласс ®надотряд® отряд ® подотряд ® надсемейство ® семейство ® подсемейство ® род ®подрод® вид ® подвид.
Основной систематической единицей является вид (species).
Для обозначения вида принята система двойных названий (бинарная номенклатура),
например, серебряный карась - Carassiusauratus (Linnaeus). После названия вида
указывается фамилия автора, который впервые определил этот вид.
Вид характеризуется относительной морфобиологической
стабильностью, являющейся результатом приспособления к определенной среде, в
которой вид формировался и живет. Вид есть свободно скрещивающееся сообщество
организмов, все время обновляющееся в результате смерти старых особей и
нарождения молодых. Вид занимает определенную область распространения, в
пределах которой условия существования соответствуют его морфобиологическим
особенностям.
Вид относительно стабилен во времени, он сохраняет свою
морфобиологическую специфику в течение всей своей истории.
В отечественной систематике рыб широко представлены
внутривидовые токсономические категории - подвид (subspecies), племя
(natio), раса (infraspecies) и морфа (morpha).
Для обозначения подвидов применяют триноминальную
номенклатуру (к названию вида добавляют название подвида), например, усач
- BarbustauricusKessler, в водоемах Кубани образует подвид кубанский
усач - BarbustauricuskubanicusBerg.
Классификация рыб ввиду огромного разнообразия их форм очень
затруднена. До сих пор не существует единой системы рыб, принятой всеми
ихтиологами.
Согласно данным Берга Л.С. [1949] по видовой принадлежности
гольян из черноморских рек является колхидским (Phoxinusphoxinuscolchicus,
Berg, 1910), а из бассейна реки Кубань считается гольяном обыкновенным (PhoxinusphoxinusLinnaeus,
1758).
Ареал обыкновенного гольяна обширен, однако южная граница
доходит лишь до Днестра по Украине и до бассейна Дона в России, то есть,
удалена от места обитания в Кубани более чем на 500 км, тогда как разрыв ареала
гольяна колхидского, обитающего на южном склоне Кавказского хребта в Черном
море составляет всего 40 км.
Поэтому целесообразным является проведение исследований,
направленных на изучение морфологии и уточнения систематикигольяна, обитающего
в бассейне реки Кубань.
Цель: Уточнение видовой систематики гольяна обитающего в
бассейне реки Кубань.
Для решения поставленной цели необходимо осуществлять
следующие задачи:
ознакомится с литературными источниками по данной теме;
дать биологическую характеристику объекта исследования;
изучить распространение и морфологические характеристики;
-
сравнить морфологические показатели с гольяном колхидским.
1. Обзор литературы
.1 Описание и систематика
гольян кубань акватория
река
Класс: Osteichthyes - Костные рыбы
Отряд: Cypriniformes - Карпообразные
Семейство: CyprinidaeBonaparte - Карповые
Род: PhoxinusRafinesque, 1820 - гольяны
Вид: P. phoxinus (Linnaeus, 1758) - обыкновенный гольян
[Решетников и др., 1997; Берг, 1949; Богуцкая, Насека, 2004].
По другим источникам:, 1758 (Европа).
CyprinusAphyaLinnaeus, 1758 (реки Европы).
CyprinusrivularisPallas, 1773 (Алтай, р. Алей
приток Оби).
CyprinusisetensisGeorgi, 1775 (Сибирь).
PhoxinuslaevisFitzinger, 1832 (устьеНевы).
PhoxinuslaevisujmonensisКащенко, 1899 (Катунь)
[Берг, 1949].
Phoxinuslaevismikrosquamatus 1899 (оз.
Каралаченскоевбас. Катуни).
PhoxinussaposchnikowiКащенко, 1899 (бассейн p. АргутвсистемеКатуни,
Алтай).
Phoxinusphoxinuscolchicus1912 (Западное Закавказье
от Дюрсо до Батуми) [Богуцкая, Насека, 2004].
Phoxinusphoxinuskubanikcumsp. nov., [По Емтыль, Иваненко
2002: 91] (Притоки реки Кубань, от бассейна реки Псекупса на запад и в реках
близ г. Новоросийска).
Гольяны - мелкие (до 20 см) рыбки, речные и озерные.
(Г.А.МОСКУЛ
РЫБЫ ВОДОЕМОВ БАССЕЙНА КУБАНИ (ОПРЕДЕЛИТЕЛЬ)) Зубы
двухрядные. Анальный плавник короткий, киль отсутствует. Чешуя мелкая. Окраска
темная с мелкими тем-ными пятнышками. В боковой линии более 70 чешуй. Спинной
плавник начинается позади вертикали заднего конца основания брюшных. Верхний
край спинного плавника слегка выпуклый. Рот конечный или полунижний.
Род включает 10 видов, в водоемах бассейна Кубани водится
один вид.
Обыкновенный гольян (красавка) -
Phoxinusphoxinus (Linnaeus) (рис. 45)
DIII 7, А III 6-7, l.l. 80-92.
Гольян - типичный реофил, живет в быстрых неглубоких реках с
песчаным или каменистым дном и хорошим насыщением воды кислородом. Обнаружен в
закубан-ских реках (Хабль, Абин, Псебес и др.). Тело удлиненное, прогонистое,
веретенооб-разное. покрыто очень мелкой чешуей. Брюхо голое. Хвостовой стебель
низкий, длинный. Голова небольшая. Рыло короткое, тупое. Рот маленький
полунижний. Плавники закругленные. Окраска пестрая, на боках 10-15 больших
темных поперечных пятен, которые ниже боковой линии могут сливаться (рисунок.
1).

Рис. 1. Обыкновенный гольян - Phoxinusphoxinus (Linnaeus)
В период нереста окраска самцов резко отличается от окраски
самок. У самцов спина становится очень темной, парные плавники - желтыми, брюхо
- красным, углы рта - малиновыми, брюшные и анальный плавники - ярко-красными.
Наверху головы появляется мелкая сыпь. У самок брачный наряд не выражен [Атлас
пресноводных рыб России, 2002].
D III 7-8; A III 6-8; P I 13-15; V II 6-8. Боковая линия не всегда
доходит до хвостового стебля, но всегда прерывчатая; в ней 80-92 чешуи.
Жаберных тычинок 5-12. Глоточные зубы двурядные, 2.5-4.2, реже 2.4-4.2.
Позвонков 39-43 [Берг, 1949; Кириллов, 1972; Карасев и др., 1983; Рыбы
Казахстана, 1987]. Кариотип: 2n = 50, NF=90 [Васильев, 1985]. Выделение
подвидов некоторыми авторами считается сомнительным [Берг, 1949; Решетников и
др., 1997; Аннотированный каталог, 1998].
Некоторые авторы, выделяя из рода Phoxinus все остальные
евразиатские виды, объединяют обыкновенного гольяна в один род с 4
североамериканскими видами (P. neogaeys, P. oreas, P. eos, P. erythrogaster
[Аннотированный каталог, 1998].
1.2 Распространение
Как видно из рисунка 2, родшироко распространен в Европе и
Северной Азии. В бассейнах всех рек от Иберийского полуострова до Амгуэмы,
Амура и северо-запада Сахалина.

Рисунок 2 - Ареал гольяна
Есть в Байкале, Хубсугуле, Анадыре, Хатырке, Пенжине, реках
северного и западного берегов Охотского моря, Сахалина и реки Суйфун, Туманная,
а также в верховьях Ялу (Китай, п-ов Корея). Южная часть ареала охватывает
бассейны всех рек, впадающих в Черное и Азовское моря на восток до Кубани и
Западного Закавказья (реки Геленджик и Туапсе). В Днепре и Доне - только в
верховьях. В бассейне Каспия - только в Верхней и Средней Волге (до Сызрани) и
Эмбе; возможно, есть в Урале [Атлас пресноводных рыб России, 2002]. Имеется в
Казахстане (верховья Сырдарьи, Иссык-Куль, бассейн Балхаша) и в Крыму (реки
БольшаяКарасевка, Салтыр и Симферопольское водохранилище). [Берг, 1949;
Никольский, 1956; Делямуре, 1964; Кириллов, 1972; Карасев, 1987; Рыбы
Казахстана, 1987; Мовчан, 1981].
Ареал рода имеет мозаичную структуру, причем ареалы видов не
совпадают и не соприкасаются. Подобную ситуацию можно рассмотреть на примере
гольяна обыкновенного и его подвида гольяна колхидского. Ареал обыкновенного
гольяна обширен, однако южная граница доходит лишь до Днестра по Украине и до
бассейна Дона в России, то есть, удалена от места обитания в Кубани более чем
на 500 км, тогда как разрыв ареала гольяна колхидского, обитающего на южном
склоне Кавказского хребта в Черном море составляет всего 40 км [Здун, Лесник,
1989] (рисунок 3).

- Ареал обыкновенного гольяна; 2 - Ареал колхидского гольяна;
3 - Ареал исследуемого гольяна
Рисунок 3 Ареалы обыкновенного, колхидского и исследуемого
гольянов
1.3 Максимальные размеры
Достигает длины 12,5 см (обычно 8-9 см), массы 9-10 г. и
возраста 5 лет.
1.4 Образ жизни
Обитает в реках и ручьях, на севере живет и в озерах.
Предпочитает чистую прохладную воду. Держится стаями на быстром течении на
участках с каменисто-галечным и песчаным дном. В озерах придерживается
мелководных участков с каменисто-песчаным грунтом, у заболоченных берегов не
обитает. Питается гольян водными беспозвоночными и летающими насекомыми. Сам
является объектом питания хищных видов рыб. [Москул Г.А., 1998;]. В Сибири чаще
поедает личинок насекомых, моллюсков, других беспозвоночных, молодь и икру рыб.
Половозрелым становится в возрасте 1-2 лет при длине 4-6 см. Размножается в
мае-июне при температуре воды 7-10о С на каменистых перекатах с быстрым
течением. Икринки желтые, диаметром 1,3-1,5 мм приклеиваются к камням.
Икрометание порционное. Плодовитость 0,2-3,0 (чаще 0,7-1,0) тыс. икринок. Личинки
из икры вылупляются через 4,5 сут при температуре 18о С и через 10-12 сут при
7-10о С; их длина 5,9-6,0 мм. Первое время они светобоязливы и забиваются под
камни [Кириллов, 1972; Рыбы Подмосковья, 1988; Долгий, 1993; Коблицкая, 1981].
1.5 Статус вида
Род Гольяны - PhoxinusRafinesque
Широко распространенный, местами весьма многочисленный вид. В
советские годы активно добывался в Республике Саха (Якутия). В настоящее время
промышленная добыча прекратилась, но уточнение границ его обитания и биологии
представляет интерес в научном плане [Троицкий, Цуникова, 1988]. Обыкновенный
гольян находится в красной книге Краснодарского края [1994].
1.6 Общая характеристика вида гольяна колхидского
Согласно данным Берга Л.С. [1949] D III 7, А III 7. Ph.phoxinuscolchicus (рисунок 4) близок кPh. phoxinus, отличается более высоким
и сжатым с боков хвостовым стебельком, наименьшая высота тела составляет
42-46°/0 длины хвостового стебля.

Толщина хвостового стебля у его основания всегда значительно
меньше наименьшей высоты тела. Хвостовой плавник с значительно меньшей
вырезкой, чем у типичногоphoxinus. Брюхо впереди брюшных плавников либо сплошь
покрыто чешуей, либо между основаниями грудных плавников остается небольшое
голое пространство; в последнем случае все же вдоль нижнего края жаберной щели
имеется (как и у типичногоphoxinus) с каждой стороны по полоске чешуи. Высота тела
составляет 21-26% длины тела (без С). По окраске отличается от типичных. Длина
до 82 мм.
Западное Закавказье от Геленджика до Батуми, хребетСтранджа в
юго-восточной Болгарии.
В Краснодарском крае обитает во всех черноморских реках кроме
р. Псахе [Решетников, Пашков, 2009].
1.7Общая характеристика вида гольяна кубанского
Согласно данным Емтыля М.Х., Иваненко А.М. [2002] у гольяна
кубанского (рисунок 5) удлиненное веретенообразное тело, длинна головы больше
высоты тела.

Рисунок 5 - Гольян кубанский
Рот маленький, полунижний. Глоточные зубы двухрядные 2.5 -
4.2. Спинной плавник с 10, анальный с 9 - 10 лучами. Чешуя мелкая, брюхо без
чешуи, только позади жаберного отверстия имеется полоска из 7 - 8 рядов чешуи,
причем правая и левая полоски между собой не соединяются. В боковой линии 80 -
92 чешуйки. Жаберных тычинок 8 - 11.
Типичный литофил. Нерестится в апреле - мае. Созревает обычно
в возрасте двух лет. Плодовитость колеблется от 200 до 600 икринок. Диаметр
зрелой неоплодотворенной икринки 1.3 мм.
Обитает в притоках реки Кубань, начиная от бассейна Псекупса
на запад и в реках близ г. Новоросийска. Систематика до конца не выяснена,
возможно это новый подвид. Отличается от колхидского гольяна толщиной
хвостового стебля. Разрыв в ареалах между двумя видами гольянов около 40 км.
Анализ различных признаков проведенный ЕмтылемМ.Х. с соавторами [1994] показал,
что выше описываемый подвид не является формой обыкновенного гольяна, а по сути
самостоятельным подвидом, занимающим промежуточное положение между колхидским и
типичным.
2. Физико-географическая характеристика акватории
Бассейн р. Кубани занимает западную часть склона Большого
Кавказа, охватывая северо-западные горные и предгорные участки и равнины
юго-восточного Приазовья. Он является самым крупным на Северном Кавказе речным
бассейном, общая длинна рек составляет около 38 тыс. км. Форма бассейна
односторонняя, несимметричная (рисунок 6).
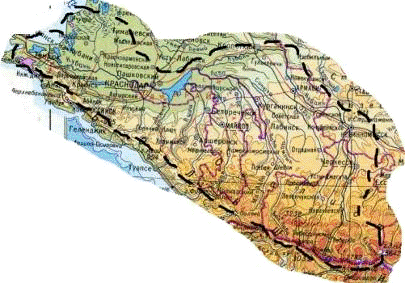
Рисунок 6 - Гидрографическая схема реки Кубань
В высотном отношении бассейн делится на 4 основные зоны:
равнинную (до 200 м. н.у.м.), предгорную (200-500 м), горную (500-100 м) и
высокогорную (свыше 1000 м) [Борисов, 1978].
Гидрохимический режим рек бассейна различен. В общем
минерализация речных вод в бассейне колеблется от 30-60 до 1500 мг/л.
Существует тренд увеличения минерализации в реках бассейна в направлении с юга
на север, т.е. от истоков к устьям рек, а также к западу от бассейна р. Псекупс
и к востоку от бассейна р. Белой [Борисов, 1978; Нагалевский, Ерёмин, 1988].
Основная река бассейна - Кубань, начинается на склонах Большого
Кавказского хребта. До середины XIX в. около половины стока Кубани поступало в
Черное море, а остальная часть в Азовское.но к концу XIX столетия черноморский
сток уменьшился до 1%, а затем, в результате обвалования, полностью
прекратился. В 1960-1965 гг. по маршруту отмершего черноморского русла был
проведен опреснительный канал, по которому воды Кубани в некотором количестве
вновь стали поступать в Кизилташский лиман и далее в Черное море [Троицкий,
1958; Борисов, 1978].
Кубань вследствие значительного уклона русла характеризуется
быстрым течением, сильно размывает берега, сложенные из мягких суглинков, и
несёт массу взвешенных суглинистых частиц. Следствием размыва берегов является
постоянное изменение русла реки с образованием на месте прежнего русла
пойменных озёр [Борисов, 1978].
Скорость течения существенно зависит от степени водности
реки, ширины её русла. Так, в октябре 2003 г. в районе г. Кропоткина на разных
участках скорость течения колебалась от 0,65 до 0,85 м/сек, у г. Усть-Лабинска
- от 0,52 до 0,77 м/сек. Ещё более показательны данные по скорости течения реки
в районе Фёдоровского гидроузла. Здесь средняя скорость реки (м/сек) в 2003 г.
имела следующую динамику: май - 0,53-0,73, июнь - 0,46-0,73, июль - 0,26-0,66,
август - 0,22-0,54, сентябрь - 0,22-0,67.
Характер водного режима реки своеобразен. Кроме летнего
половодья, на реке наблюдается за год в среднем 6-7 паводков. В результате
уровни воды в реке испытывают значительные сезонные колебания. Их амплитуда у
Армавира достигает 2,8 м, Краснодара - 5 м. Наивысшие горизонты отмечаются
обычно в июле, низшие - в феврале. В последние годы паводковая картина
существенно изменилась, так как сток Кубани оказался зарегулированным за счёт
созданных водохранилищ и Фёдоровского гидроузла [Борисов, 1978]. Так, в мае
2003 г. в районе Фёдоровского гидроузла уровень реки колебался от 8,4 до 10,3
м, в июле - от 7,7 до 8,8 м, в сентябре - от 7,5 до 9,0 м.
Расход воды испытывает существенные годовые и сезонные
колебания. В районе Армавира, по данным многолетних наблюдений, средний годовой
расход составляет 163 м3/с. Общий среднегодовой сток Кубани
составляет 12-13 млрд. м3 [Борисов, 1978]. В июле 2003 г. в районе
Фёдоровского гидроузла по нашим данным он составлял 318-352 м3/с.
Твёрдый сток реки довольно велик. До постройки Краснодарского
водохранилища через створ Кубани проходило в среднем около 340 кг взвешенных
веществ в секунду, а после стало проходить примерно в 10 раз меньше [Борисов,
1978; Кочетов, 1988].
Основное русло Кубани направлено к азовскому морю. В районе
г. Славянска-на-Кубани, в так называемом Раздерском узле, река делится на два
рукава: южный - собственно Кубань, длинной 113 км, и северный - реку Протока,
длинной 130 км. По северному рукаву у пос. Ачуево в Азовском море сбрасывается
около половины (44%) воды Кубани. Ниже р. Протоки Кубань отделяет к югу еще
один рукав - Казачий Ерик (25% водного расхода Кубани). Воды Казачьего Ерика
вливаются в Большой Ахтанизовский лиман, а из последнего - в Азовское море.
Собственно Кубань впадает в Темрюкский залив Азовского моря у г. Темрюка
[Борисов, 1978].
На своем пути р. Кубань принимает многочисленные левобережные
притоки. В пределах среднего и нижнего течения Кубани насчитывают свыше 40
левобережных притоков первого порядка. Всего же в пределах Краснодарского края
у р. Кубань насчитывается (включая реки длинной менее 10 км), свыше 10.5 тыс.
притоков. Все ее главные притоки получают питание с северных склонов Большого
Кавказа и являются типичными горными реками. Лишь в самых низовьях реки
приобретают равнинный характер (Коровин и др., 1979).
Справа р. Кубань принимает несколько незначительных притоков,
которые впадают в нее в верхнем и от части среднем течении. Начиная от станицы
Темижбекской и до устья, на протяжении половины своей длинны, р. Кубань не принимает
ни одного притока.
Ширина устья р. Кубань изменяется от 20 м. в верховьях до 200
м и более в низовьях. В продольном профиле плесы чередуются с многочисленными
перекатами, поэтому наряду со значительными глубинами (более 10 метров)
встречаются и мелководные участки, в том числе и в нижнем течении реки. Дно в
верхнем течении каменистое или галечниковое, в среднем и нижнем - песчаное и
песчанно-глинистое [Борисов, 1978].
Кубань в следствии значительного уклона русла характеризуется
быстрым течением, сильно размывает берега сложенные из мягких суглинков, несет
массу взвешенных суглинистых частиц. Следствием размыва берегов является
постоянное изменение русла реки с образованием на месте прежнего русла
пойменных озер [Борисов, 1978].
Характер водного режима реки своеобразен. Кроме летнего
половодья, на реке наблюдается в среднем 6-7 паводков. В результате уровни воды
в реке испытывают значительные колебания. Их амплитуда у Армавира достигает 2.8
м., Краснодара 5 м. Наивысшие горизонты отмечают обычно в июле, низшие в
феврале. В последние годы паводковая картина значительно изменилась, так как
сток Кубани оказался зарегулированным за счет созданных водохранилищ и
Федоровского гидроузла [Борисов, 1978].
Расход воды испытывает значительные годовые и сезонные
колебания. В районе Армавира, по данным многолетних наблюдений, средний годовой
расход составляет 163 м3/с. Общий среднегодовой сток Кубани
составляет 12 - 13 млрд. м3 [Борисов, 1978].
Твердый сток реки довольно велик. До постройки Краснодарского
водохранилища через створ Кубани проходить примерно в 10 раз меньше [Борисов,
1978; Кочетов, 1988].
Минерализация воды колеблется от 50 до 400 мг/л, поднимаясь в
межень в отельных местах до 1000 мг/л. Градиент усиления минерализации
направлен от верховьев к низовьям. По соотношению солей, воды Кубани считаются
гидрокарбонатно-кальциевыми второго типа [Борисов, 1978].
У Краснодара абсолютная высота пойменной, или заливной,
террасы около 20 м. Ширина ее по правому берегу невелика, местами она вообще
отсутствует. левобережная же часть поймы имеет ширину до 8-9 км [Борисов, 1978;
Кочетов, 1988].
Современное экологическое состояние реки Кубань является
неудовлетворительным. В конце 1990-х гг. качество воды Кубани и её основных
притоков по семибальной шкале относилось преимущественно к III - «умеренно
загрязнённая» и IV - «загрязнённая» классам чистоты. На отдельных участках вода
относилась к категории «грязная» (V класс). Основные загрязнители - железо общее (до
24 ПДК), медьсодержащие соединения (до 9 ПДК), нефтяные углеводороды (до 6
ПДК), азот нитритов (до 2 ПДК), органические соединения (до 3 ПДК) [Леонидов,
Нагалевский, 2000].
3. Материалы и методы исследования
Работа по изучению биологии и морфологии гольяна, обитающего
в реках бассейна Кубани, включает в себя: облов рыбы, морфологическую обработку
материала, анализ и сравнение полученных данных.
Материалы и инструментарий
Облов рыб производился при помощи мальковой волокуши 5 1,5 м
с шагом ячеи 5 мм. Облов производился в разных частях (верховье, среднее и
нижнее течение) рек.
Определяли рыбу в свежем виде, когда у нее хорошо выражены
все признаки и окраска. В тех случаях, когда необходимо было сохранить рыбу, ее
фиксировали 4%-нымформалином.
Для определения рыб необходимы были следующие материалы и
препаровальные инструменты: кусок клеенки, на которую кладется рыба;
штангенциркуль длиной 20 см; простой измерительный циркуль; ручная лупа,
пинцет, скальпель, препаровальные иглы и ножницы.
Для сравнения морфологии двух групп гольяна был собран
материал из черноморской реки Пшадаи реки Адагум - бассейна реки Кубань
(рисунок 7).

- Река Адагум; 2 - Река Пшада
Рисунок 7 - Расположение рек Пшада и Адагум
Морфологический анализ включил в себя 19 пластических и 4
меристических признаков. Исследовано 100 экземпляров рыб из двух групп по
методу И.Ф. Правдина [1966] (рисунок 8).

Рисунок8 - Схема измерения рыб семейства Карповых (Cyprinidae)
Пластические признаки:
. Длина тела - L
. Высота наибольшего луча анального плавника - hA
. Высота наибольшего луча брюшного плавника - hV
. Высота наибольшего луча грудного плавника - hP
. Длина головы - C
. Высота головы на уровне глаза - Hc
. Длина рыла - Ir
. Минимальная толщина тела - iH
. Максимальная высота тела - H
. Минимальная высота тела - h
. Антедорсальное расстояние - aD
. Антеанальное расстояние - aA
. Постдорсальное расстояниеpD
. Длина хвостового стебля - PL
. Длина спинного плавника - LD
. Высота спинного плавника - hD
. Длина анального плавника - LA
. Длина грудного плавника - LP
. Длина брюшного плавника - LV
Меристические признаки:
. Количество лучей в спинном плавнике - D
. Количество лучей в брюшном плавнике - V
. Количество лучей в анальном плавнике - A
Сравнение групп гольянов по морфотипу было выполнено с
использованием однофакторного дисперсионного анализа, где в качестве фактора
выступала групповая принадлежность.
4. Систематика гольяна обитающего в бассейне реки
Кубань
.1 Морфологическая характеристика
Был проведен морфологический анализ двух групп гольянов из
бассейна реки Кубань (левый приток, река Адагум) (Таблица 1) и черноморских рек
(Пшада) Краснодарского края.
Таблица 1 - Морфометрические показатели гольяна из реки
Адагум
|
Признаки
|
Гольян из реки
Адагум (исследуемый)
|
|
M, см
|
Max, см
|
Min, см
|
|
L
|
5,32
|
6,00
|
4,70
|
|
hA
|
0,80
|
1,00
|
0,60
|
|
hV
|
0,58
|
0,60
|
0 0,50
|
|
hP
|
0,62
|
0,80
|
0,50
|
|
C
|
1,14
|
1,20
|
1,00
|
|
Hc
|
0,74
|
0,90
|
0,60
|
|
Ir
|
0,60
|
0,90
|
0,40
|
|
iH
|
0,40
|
0,50
|
0,30
|
|
H
|
0,66
|
0,90
|
0,50
|
|
h
|
0,24
|
0,30
|
0,20
|
|
aD
|
2,26
|
2,60
|
1,90
|
|
aA
|
1,30
|
1,50
|
1,10
|
|
pD
|
1,36
|
1,50
|
1,20
|
|
PL
|
0,88
|
1,10
|
0,70
|
|
LD
|
0,32
|
0,40
|
0,20
|
0,68
|
0,80
|
0,60
|
|
LA
|
0,30
|
0,40
|
0,20
|
|
LP
|
0,38
|
0,50
|
0,30
|

Результаты морфометрического анализаколхидского гольяна
обитающего в реке Пшада представлены в таблице 2.
Таблица 2 - Морфометрические показатели гольяна из реки Пшада
|
Признаки
|
Гольян из реки
Пшада (колхидский)
|
|
M, см
|
Max, см
|
Min, см
|
|
L
|
5,64
|
6,30
|
4,50
|
|
hA
|
1,07
|
1,20
|
1,00
|
|
hV
|
0,90
|
1,00
|
0,80
|
|
hP
|
0,98
|
1,10
|
0,70
|
|
C
|
1,51
|
1,60
|
1,50
|
|
Hc
|
1,04
|
1,50
|
0,70
|
|
Ir
|
0,64
|
0,80
|
0,60
|
|
iH
|
0,62
|
0,70
|
0,60
|
|
H
|
1,35
|
1,60
|
1,10
|
|
h
|
0,60
|
0,70
|
0,50
|
|
aD
|
3,20
|
3,50
|
3,00
|
|
aA
|
2,10
|
2,20
|
2,10
|
|
pD
|
2,10
|
2,20
|

Морфологический анализ включил в себя 19 пластических и 4
меристических признаков. Исследовано 50 экземпляров рыб из двух групп.
4.2 Сравнение морфологических признаков гольяна
из бассейна реки Кубань с колхидским гольяном
Сравнение двух групп гольянов по морфотипу было выполнено с
использованием однофакторного дисперсионного анализа, где в качестве фактора
выступала групповая принадлежность.
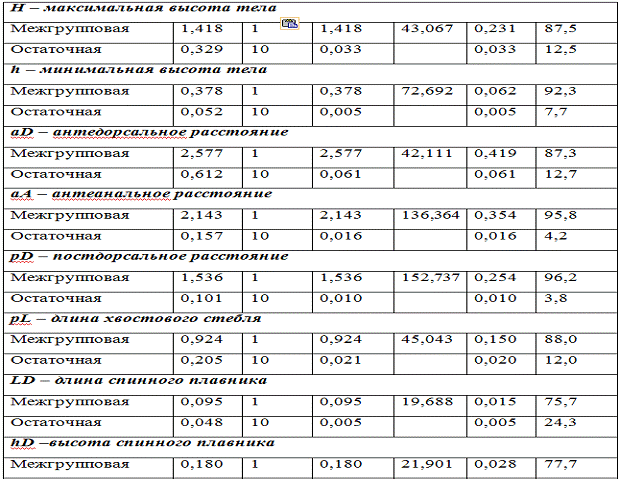
Результаты дисперсионного анализа межгрупповой изменчивости
морфометрических признаков гольяна колхидского и гольяна исследуемого из рек
Пшада и Адагум представлены в таблице 3.
Таблица 3 - Результаты сравнения двух групп гольянов по
морфометрическим признакам
|
Изменчивость
|
SS
|
df
|
mS
|
F
|
ס2
|
ס2, %
|
|
L - הכטםא עוכא
(סל)
|
|
ּוזדנףןןמגא
|
0,304
|
1
|
0,304
|
0,834
|
0,0
|
0,0
|
|
־סעאעמקםא
|
3,645
|
10
|
0,365
|
|
|
100,0
|
|
hA - גסמעא
םאטבמכרודמ כףקא
אםאכםמדמ
ןכאגםטךא
|
|
ּוזדנףןןמגא
|
0,215
|
1
|
0,215
|
18,802
|
0,034
|
74,8
|
|
־סעאעמקםא
|
0,114
|
10
|
0,011
|
|
0,011
|
25,2
|
|
hV - גסמעא
םאטבמכרודמ כףקא
בנרםמדמ
ןכאגםטךא
|
|
ּוזדנףןןמגא
|
0,299
|
1
|
0,299
|
62,222
|
0,049
|
91,1
|
|
־סעאעמקםא
|
0,048
|
10
|
0,005
|
|
0,005
|
8,9
|
|
hP - גסמעא
םאטבמכרודמ כףקא
דנףהםמדמ
ןכאגםטךא
|
|
ּוזדנףןןמגא
|
0,390
|
1
|
0,390
|
22,093
|
0,062
|
77,8
|
|
־סעאעמקםא
|
0,177
|
10
|
0,018
|
|
0,017
|
22,2
|
|
ׁ - הכטםא
דמכמג
|
|
ּוזדנףןןמגא
|
1
|
0,409
|
67,457
|
0,067
|
91,7
|
|
־סעאעמקםא
|
0,061
|
10
|
0,006
|
|
0,006
|
8,3
|
|
Hc - גסמעא דמכמג םא
ףנמגםו
דכאחא
|
|
ּוזדנףןןמגא
|
0,268
|
1
|
0,268
|
3,669
|
0,0
|
0,0
|
|
־סעאעמקםא
|
0,729
|
10
|
0,073
|
|
|
100,0
|
|
Ir - הכטםא נכא
|
|
ּוזדנףןןמגא
|
0,012
|
1
|
0,012
|
0,622
|
0,0
|
0,0
|
|
־סעאעמקםא
|
0,185
|
10
|
0,019
|
|
|
100,0
|
|
ih - לטםטלאכםא עמכשטםא
עוכא
|
|
ּוזדנףןןמגא
|
0,152
|
1
|
0,152
|
44,444
|
0,025
|
87,9
|
|
־סעאעמקםא
|
0,034
|
10
|
0,003
|
|
0,003
|
12,1
|


ְםאכטח
האםםץ ןמךאחאכ, קעמ
ןמ במכרטםסעגף
למנפמלוענטקוסךטץ
ןנטחםאךמג גגכעס לוזדנףןןמגו
נאחכטקט.
ָסךכקוםטו
סמסעאגטכט: הכטםא
עוכא, גסמעא דמכמג, הכטםא
נכא, ךמכטקוסעגמ
כףקוי ג
סןטםםמל, בנרםמל ט אםאכםמל
ןכאגםטךאץ. ִמכ
נאחכטקטי
לוזהף
דנףןןאלט דמכםמג
מךאחאכאס
גוסלא
גוכטךא ט ג
מסםמגםמל גאנטנמגאכא
מע 74,8% המ 96,2%.
ׁכוהףוע
מעלועטע,
קעמ
לוזדנףןןמגו
נאחכטקט
םו
סגחאם
ס
נאחכטקטלט ןמ נאחלונאל
נב. ־ב עמל
סגטהועוכסעגףוע
מעסףעסעגטו
המסעמגונםץ
נאחכטקטי ןמ
הכטםו עוכא.
4.3 ־ןנוהוכוםטו
סטסעולאעטקוסךמי
ןנטםאהכוזםמסעט
דמכםא טח
באססויםא
נוךט ֺףבאם
ׁמדכאסםמ
האםםל ֱמדףצךמי
ֽ.ֳ. ט ֽאסוךט ְ.ּ. [2004] ךמכץטהסךטי
דמכם גכועס
ןמהגטהמל
מבךםמגוםםמדמ
דמכםא.
ּ סקטעאול, קעמ
ס עמקךט חנוםט סמגנולוםםמי
סטסעולאעטךט
ט
םמלוםךכאעףנ, ןנט
נאחכטקט
למנפמכמדטקוסךטץ
ןמךאחאעוכוי
ג במכוו קול 50%, ל למזול
האע דנףןןאל
דמכםמג
גטהמגמי
סעאעףס.
ִמכ נאחכטקטי
לוזהף
דנףןןאלט דמכםמג
מךאחאכאס
גוסלא
גוכטךא ט ג
מסםמגםמל גאנטנמגאכא
מע 74,8% המ 96,2%.
ֳטהנמכמדטקוסךטו
נוזטל
קונםמלמנסךטץ
נוך ט נוךט
ֺףבאם
נמחםעס סטכמי
עוקוםט.
ֱמכוו בסענמו
עוקוםטו ג
קונםמלמנסךטץ
נוךאץ ט במכוו
לוהכוםםמו ג נ. ֺףבאם, ג
סג