К вопросу о методах генезисных построений развития фитостромы
К ВОПРОСУ
О МЕТОДАХ ГЕНЕЗИСНЫХ ПОСТРОЕНИЙ РАЗВИТИЯ ФИТОСТРОМЫ
Харитонцев Б.С.
Любое явление (предмет) включает общие и частные признаки. Соответственно
его изучение может идти в дедуктивном (дедукция -логическое умозаключение от
общего к частному) и индуктивном (индукция - логическое умозаключение от
единичных фактов к обобщению) направлениями (Словарь иностранных слов, 1984).
Поэтому методы исследований можно разделить на дедуктивные и индуктивные. Это в
полной мере относится и к изучению генезиса фитостромы любой территории,
например, юга Западной Сибири. Обсуждая данный вопрос следует подчеркнуть, что
фитострома (растительное население) характеризуется многими измерениями:
флорой, растительностью, флороценокомплексами, спектром жизненных форм и т.д.
Изучение каждого направления генезиса фитостромы определённой территории или
других выделов требует свои методы. Нами велись генезисные построения флоры и
растительности юга Западной Сибири, поэтому мы подробнее остановимся на методах
изучения генезиса двух измерений фитостромы. Все используемые нами методы можно
представить схемой 1.

Статистика - систематический метод - предполагает изучение видов
определённых семейств (прежде всего десяти ведущих в характеризуемой флоре) по
их экологической, ареалогической и другим характеристикам (Харитонцев, 2004)
через вычисление коэффициентов синантропизации, космополитизации,
принадлежности (таёжной, подстепной, степной, пойменной, боровой, северной,
восточной, западной), гумидности - аридности, оригинальности (эндемизма),
экологической принадлежности по формуле
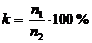 ,
,
где
k - коэффициент синантропизации и т.д.; п1 - число
видов конкретной группы в данном семействе; п2 - число всех видов данного
семейства. Сопоставляя гомологичные коэффициенты, можно составить
флорогенетическую картину определённого явления (рис. 1).
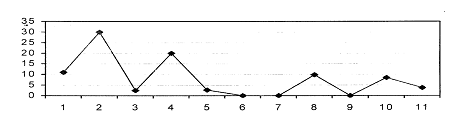
Рис.
1. Участие (%%) ведущих семейств в формировании таёжной флоры юга Западной
Сибири.
Злаки
- 1, осоковые - 2, лютиковые - 3, розановые - 4, бобовые - 5, зонтичные - 6,
губоцветные - 7, норичниковые - 8, крестоцветные - 9, гвоздичные - 10, астровые
- 1. у - процент участия, х - ведущие семейства.
Анализ
показал, что из ведущих семейств основное участие в формировании таёжной флоры
играют злаки, розановые и норичниковые. Дальнейший видовой анализ может
выявить, например, вторичные центры происхождения определённого семейства в
таёжной зоне Западной Сибири.
Систематика
- популяционный метод, предполагает изучение популяционных характеристик видов
одного рода в плане сравнения их участия в процессе флорогенеза. Исследование
популяций даёт определённую флорогенетическую информацию. Это относится, прежде
всего, к выявлению репродуктивных усилий вида весовым методом. Подобные
исследования можно проводить в таком порядке:
1. Определяются объекты. Чаще всего это
виды одного рода, например, Allium
в исследуемом регионе;
2. Изучается соотношение масс корней,
листьев, стеблей, цветков, переведённое в %%, для разных возрастных групп;
3. Из изученных видов определяют вид,
наиболее примитивный в данном перечне (по нашим наблюдениям его соотношение
масс органов совпадает с таковым для виргинильных стадий);
4. Затем, сравнивая графики соотношения
масс органов генеративных растений всех видов с графиком исходного вида, можно
составить картину их формирования в фитоценотрическом и экологическом
направлениях, исходя от исходного вида.
Метод
флорогенетической триады признаков
Различные признаки растений имеют неодинаковое флорогенетическое
значение. Из их многообразия наибольшее флорогенетическое содержание отмечено
нами для морфологических, экологических и географических признаков.
Каждый из них имеет по три подуровня (степени изменения): морфологические
- подродовой (Г), секционный (Д), видовой (Е); географический - размеры ареалов
от (А) площади флористического царства (континенты) через размеры
соответствующие площади флористических областей (Б) к ареалам совпадающим по
площади с провинциями, округами, районами (В); экологический - виды более или
менее стенотопные (Ж) - виды дуалисты или же стенотопно-эвритопные (З) - виды
экологические антогонисты, типичные эвритопы или же крайне специализированные
(И). Число подуровней каждого признака соответствует числу трёх временных
отрезков формирования флоры: третичное время - плейстоцен - голоцен. Можно предположить,
что для третичного времени характерны подуровни - Г, А, И; для плейстоцена - Д,
Б, З; для голоцена - Е, В, Ж.

Метод географо-филогенетический (метод родовых спектров) предусматривает
анализ видов политипных родов.
Нами при характеристике флорогенеза на юга Западной Сибири
проанализированы родовые спектры ряда секции Carex, а также родов Geranium, Poa, Euphorbia, Stipa, Campanula, Galium и др.
1. В качестве примера можно привести
анализрода Galium.
2. На юге Западной Сибири произрастают
следующие виды (Флора Сибири, т. 12): Galium (G.) aparine L., G.boreale L.,G.mollugo L., G.palustre L., G.physocarpum Ledeb, G.ruthenicum Willd, G.spurium, G.tinctorum (L.) Scop., G.trifidum L., G.triflorum Michaux, G.verum L..Выделим видовые признаки и из них - имеющие
флорогенетическое значение.
4а. Обращает внимание, что на территории юга Западной Сибири (равнина)
отсутствуют третичные реликты: G.paradoxa, G.odoratum и виды
близкого рода Cruciata, т. е. участие видов рода Galium во флорогенезе в основном связано с
послетретичным временем; сохранившиеся виды с третичными корнями (G.trifidum, G.triflorum и др.)
являются «таёжными гидрофильными», «степными», и др., т. е. участвовали во
флорогенезе в качестве вторичных плейстоценовых компонентов.
б. При восстановлении флорогенеза рода анализируются признаки, которые
можно разделить на адаптивно-продвигающие (способствующие новому
флорогенетическому продвижению - А. Пр.), адаптивно-расширяющие (способствующие
расширению экониши на прежнем флорогенетическом уровне - А.).
в. Формируя (зная) габитус исходной(родительской) формы мы можем
составить картину флорогенеза рода в целом.
г. Исходя из дополнительных данных для региона (характер ареала,
фитоценотическая принадлежность, экологические особенности) можно восстановить
историю рода в конкретном регионе.
Метод определения участия вида в генезисе фитобиоты
Вид А при миграции попадает на новую территорию, где формирует свою
популяцию, внедряется в аборигенные фитоценозы, а также флороценокомплексы.
Эти процессы будут иметь численные характеристики. Их величины
определяются формулами:
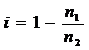 ,
,
где
i - индекс фитоценотического противодействия
аборигенных фитоценозов; п1 - число фитоценозов, куда вид внедрился, п1 - число
фитоценозов всего фитоценокомплекса.
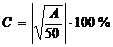 ,
,
где
с - степень заселения экоячейки; А - активность вида, определяемая как 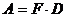 ; F - встречаемость (1-10 баллов), D - -
обилие вида (1-5 баллов).
; F - встречаемость (1-10 баллов), D - -
обилие вида (1-5 баллов).
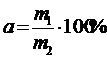 ,
,
где
а - степень внедрения вида в фитоценоз; т1 - число видов, положительно
сопряжённых с видом А, т2 - общее число всех видов в фитоценозе.
Метод
циклического развития
В
течение межледниковья и оледенения на территории юга равнины (Гричук, 1961)
последовательно сменяются термоксеротическая (теплая и сухая) - ТКс,
термогигротическая (теплая и влажная) - ТГ, криогигротическая (холодная и
влажная) - Кги, криоксеротическая (холодная и сухая) - ККс климатические
стадии. Крылов В.Г. (1961) дает схему оптимальных эколого-фитоценотических
ареалов групп типов леса. Поле ареалов групп типов леса по сочетанию
климатопов, влажности почв и их механического состава также подразделяется на 4
группы: 1) холодный умеренно-влажный климатоп, сухие почвы - доминируют
лишайниковые (1) группы типов; 2) умеренно холодный влажный климатоп, сырые
почвы (глины) - доминирует долгомошниковая группа (2) типов леса; 3) климатоп
умеренно-тепло-влажный или теплый суховатый, почвы сухие - развиваются
сухокустарниковая или низкотравная (3) группа типов леса; 4) климатоп умеренно
теплый влажный (теплый сухой) почвы сырые (глины) - преимущество получает
сложная группа (4) типов леса. Зеленомошниковая группа является центральной, связывающей
все перечисленные группы - 1, 2, 3, 4. Сфагновая и голубико-багульниковая
связывает 1 и 2 группы, травяно-болотная и крупнопапоротниковая 2 и 3 группы,
широкотравная и орляковая - 3 и 4 группы, вейниковая - 3, 4 и 1 группы.
Совпадение (количественное и качественное) числа климатических стадий
плейстоценовых ритмов и числа климатопов наводит на мысль об их синхронном
протекании (возникновении) в плейстоцене.
Сам эзогенез ассоциаций проходил в плакорных условиях. При этом возможны
два варианта механизма эзогенеза (на примере темнохвойных формаций):
б) зеленомошниковая группа является основой лишь одной производной группы
соответствующей климатической стадии, от которой произошла группа следующей
климатической стадии, от нее - третьей и т.д. Графически это отражено на рис
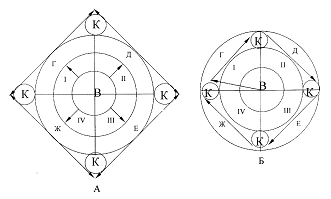
Рис. 2. Механизмы эзогенеза групп типов леса темнохвойной тайги:
генезис
фитострома флора таёжный
А - вариант радиально-генетический, Б - вариант спирально-генетический; I,II,III,IV - климатические стадии ритма
плейстоцена; в - зеленомошниковая группа типов леса, г, д, е, ж - производные
группы, к - переходные группы.
В случае А-эзогенеза должны существовать прочные флористические связи по
схеме В-Г, В-Д, В-Е, В-Ж. При Б-эзогенезе характер связей иной и выражается
отношением В-Г-Д-Е-Ж. Вполне возможно, что в процессе фитоценогенеза в
плейстоцене имели место эти два варианта. Возникнув в процессе первого ритма,
все группы ассоциаций темнохвойной тайги существовали, перемещаясь по правилу
предварения Алехина в пространстве и времени по спирали и одновременно от ритма
к ритму изменяясь (и, прежде всего за счет изменения фитоценотической роли
видов, населяющих их).
Метод узловых
сообществ основан на использовании коэффициента Жаккара. Он позволяет из всего
перечня сообществ внутри формации выделить сообщество, имеющее наибольший
коэффициент сходства с остальными. Его можно считать узловым, давшим начало
всем остальным внутри данной формации.
Метод морфолого-ареалогический (метод флорул)
Флорула - понятие, введённое во флористику А.И. Толмачёвым (1974),
используемое при географическом и генетическом анализе флоры.
Нами данное понятие используется во флорогенетическом значении. Флорула -
коадаптированная группа видов, возникших в одном генетическом центре,
вследствие чего её виды имеют сходный уровень морфологических признаков (внутри
своего семейства) и гомологичный принцип формирования ареала. К примеру, виды
третичных флорул по своим морфологическим особенностям занимают изолированное
положение в своих семействах, а ареал - близкий к космополитному или
дизъюнктивный или точеный.
Анализируя виды флоры юга Западной Сибири, нами выделено 44 флорулы. Из
них третичных флорул - 15, плейстоценовых - 16, голоценовых - 13. Исходя из
особенностей ареалов видов, входящих в состав флорул, для каждой из них было
определено направление проникновения на территорию юга Западной Сибири и
характер перемещений её видов в характеризуемом регионе.
Вопросы генезиса фитостромы - наиболее сложные среди биологического
цикла. Они требуют комплексного и всестороннего исследования.
Литература
1. Гричук М.П. Основные черты изменения
растительного покрова Сибири в течение четвертичного периода / М.П. Гричук //
Палеогеография четвертичного периода СССР. - М.: 1961. - С. 190-205.
2. Крылов Г.В., Салатова Н.Г. Леса
Западной Сибири / Г.В. Крылов, Н.Г. Салатова. - Новосибирск, 1950. - 421 с.
3. Словарь Иностранных слов. - М.: Рус.
яз., 1984. - 608 с.
4. Толмачев А.И. Введение в географию
растений / А.И. Толмачев. - Л.: Издат. ЛГУ, 1974. - 244 с.
Харитонцев
Б.С. Ареалогия и генезис флоры юга Западной Сибири / Б.С. Харитонцев. -
Тобольск: Издательство ТГПИ, 2004. - 117 с.