Влияние аммиачной селитры и суперфосфата на личиночное развитие озерной лягушки
Министерство
образования и науки Российской Федерации
Федеральное
государственное бюджетное образовательное учреждение
высшего
профессионального образования
Кубанский
государственный университет
Кафедра
зоологии
Дипломная
работа
Влияние
аммиачной селитры и суперфосфата
на
личиночное развитие озерной лягушки
Работу выполнила
Мамий З.А.
Специальность
020201 Биология
Нормоконтролёр,
д-р. биол. наук, профессор
Плотников Г.К.
Краснодар
2013
РЕФЕРАТ
Дипломная работа выполнена на 39 страницах машинописного
текста, в работе содержится 4 таблицы, 1 рисунок. В список использованной
литературы входит 42 источника, в том числе 27 на английском языке.
Ключевые слова: Озерная лягушка, личиночное
развитие, темп роста, Минеральные удобрения
Цель дипломной работы -
выяснить влияние минеральных удобрений на личиночное развитие (смертность, темп
линейного и весового роста головастиков) озерной лягушки Pеlophylax ridibundus
Pall.
Смертность головастиков озерной лягушки в
растворах аммиачной селитры увеличивается с 60 до 97% (в контроле смертность
29%) с возрастанием концентрации минерального удобрения. В растворе
суперфосфата самой низкой из исследованных концентраций погибли 97%, а в более
высоких 100% головастиков.
Темп линейного роста в растворах исследованных
удобрений меньше, а темп весового роста больше, чем в контроле.
Сравнение воздействия аммиачной селитры и
суперфосфата на головастиков озерной лягушки показало, что оба исследованные
удобрения оказывают сходное воздействие, вызывая их гибель (96-97%) в
концентрации 10мг/л. Концентрация аммиачной селитры 5 мг/л является
сублетальной. Более высокие концентрации суперфосфата. Различий во влиянии на
темп линейного и весового роста головастиков в растворах двух исследованных
удобрений не отмечено.
Содержание
Введение
.
Аналитический обзор
.
Материал и методы исследования
.
Влияние аммиачной селитры и суперфосфата на личиночное развитие озерной лягушки
.1
Смертность головастиков озерной лягушки в растворах аммиачной селитры и
суперфосфата
.2
Темп роста головастиков озерной лягушки в растворах аммиачной селитры и
суперфосфата
Заключение
Библиографический
список
Введение
Проблема токсического действия азотсодержащих
соединений на рыб возникла в связи с рыборазведением. Уровень азотсодержащих
соединений - аммиака, нитратов и нитритов в воде водоемов -
варьирует в широких пределах в зависимости от типа водоема, гидробиологических
условий, антропогенного воздействия и других факторов. В течение вегетационного
периода в результате жизнедеятельности водяных растений количество
азотсодержащих соединений в воде значительно уменьшается. Увеличение же уровня
этих веществ в воде водоёмов в основном связано с интенсивным использованием в
сельском хозяйстве минеральных азотсодержащих удобрений. Несоблюдение правил их
применения ведет к загрязнению почвы, откуда азотсодержащие соединения
вымываются в водоемы. Кроме того, нитраты и нитриты образуются из аммиака,
являющегося конечным продуктом обмена веществ у рыб [Велдре,
Зоома, 1990].
Интенсификация сельскохозяйственного
производства обязательно включает в себя широкое использование различных
химических веществ, среди которых первыми в списке значатся минеральные
удобрения и разнообразные пестициды (инсектициды, акарициды, гербициды,
фунгициды).
Для повышения урожайности сельскохозяйственных
культур огромное значение имеет внесение в почву элементов, необходимых для
роста и развития растений. Эти элементы вносятся в почву в виде органических
(навоз, торф и др.) и минеральных (продукты химической переработки минерального
сырья) удобрений. Производством минеральных удобрений занята химическая
промышленность. Основные группы минеральных удобрений -
азотные, калийные и фосфорные. Кроме них, растениям для нормальной
жизнедеятельности необходимы и некоторые химические элементы в малых дозах, это
микроэлементы (Fe, Cu, Mn, Mo, B, Zn, Co). Микроэлементы вносятся в почву с
микроудобрениями.
Существует практика применения удобрений в
выростных прудах рыбопитомников. Так, например, выростные пруды удобряются
аммиачной селитрой из расчета 70 кг/га. Развитие фитопланктона в прудах часто
лимитируется минеральным фосфором. Поэтому для повышения естественной
рыбопродуктивности нужно применять и фосфорные удобрения, регулярно контролируя
гидрохимический режим в прудах [Булатова, 1976].
Для уничтожения вредителей растений -
насекомых, пауков, грибов и сорных растений в сельском хозяйстве широко
используются пестициды.
И пестициды, и удобрения, вносимые в почву или
на растения, с дождевой водой попадают в различные водоемы и оказывают
определенное воздействие на всех обитающих в водоемах живых организмов.
Влияние пестицидов на животных, и в частности на
обитающих в водоемах земноводных, в последние 25 - 30 лет
изучается очень широко в разных географических точках планеты, в разных
странах, на разных видах бесхвостых и хвостатых амфибий. Большой список работ
по влиянию пестицидов на земноводных приведен в монографии Т.Ю. Песковой [2001].
Что же касается влияния удобрений на
земноводных, то здесь перечень работ гораздо короче, и эти работы вышли в свет
в основном в последние 20 - 25 лет. Преимущественно это
работы зарубежных авторов, исследований на территории России единичны.
В связи с этим и была избрана тема данной
дипломной работы.
Цель дипломной работы -
выяснить влияние минеральных удобрений на личиночное развитие озерной лягушки
Pеlophylax ridibundus Pall.
Задачи работы следующие:
. В лабораторных условиях определить смертность
головастиков озерной лягушки в контроле и растворах минеральных удобрений
(аммиачной селитры и суперфосфата) различной концентрации;
. Установить темп линейного и весового роста в
период личиночного развития озерной лягушки при тех же условиях опыта;
. Сравнить действие двух исследованных удобрений
на личиночное развитие озерной лягушки.
удобрение личиночный лягушка рост
.Аналитический обзор
Нитриты в водной среде встречаются в виде ионов
NO2 и свободной кислоты HNO3. Более токсичны они в форме свободной кислоты, так
как кислота быстрее проникает через биологические мембраны. Дозы токсичности
для рыб как нитратов, так и нитритов в воде варьируют в широких пределах.
Обнаруженные ЛД50 нитритов составляют для карпов 1,8 мг/л, гольяна -
40 мг/л по иону.
Воздействие различных загрязнителей испытывают
на себе в большей или меньшей степени все живые организмы. При этом земноводные
находятся в особом положении, так как на протяжении онтогенеза они обитают в
двух средах - водной и наземной. На личинок амфибий
воздействуют токсические вещества, попадающие в водоемы при смыве с полей, а
взрослые амфибии, питаясь отравленными беспозвоночными на суше, в воде также
испытывают воздействие токсикантов.
Большой объем выбросов в окружающую среду
отходов промышленного производства и широкомасштабное внесение синтетических
химических удобрений в почву приводят (в результате переноса с воздушными и
водными потоками) к попаданию загрязнителей в места обитания диких животных,
значительно удаленных от источников выброса загрязняющих веществ.
Резкое повышение содержания нитратов в
поверхностных и грунтовых водах вызывает опасение, что это может привести к
сокращению амфибий.
Способность длительное время сохраняться и
сравнительно легко перераспределяться в природных средах определяет условия для
широкого распространения различных химикатов.
Огромные негативные последствия хронического
воздействия низких концентраций многих ксенобиотиков на организмы животных
определяет особую важность постоянного контроля за изменением фоновых уровней
загрязнения фауны этими соединениями [Воронова, Денисова,
Пушкарь,1987].
Земноводные представляют удобный объект для
исследования результатов влияний антропогенных загрязнений, так как они
достаточно обычны в различных материковых водоемах, широко распространены, на
разных этапах онтогенеза являются то фито-, то зоофагами, доступны для
изучения, биология большинства их видов хорошо изучена, они многоплодны и
достаточно быстро воспроизводятся по времени, пригодны для индикационного
использования [Пескова, 2001].
Отмечается токсичность для амфибий химикалий,
используемых в системе спортивного рыболовства, и удобрений. Загрязнения
приводят к нарушению гормональных реакций, низкой выживаемости икры,
обнаруженных в популяциях в районе Великих Озер и реки Святого Лаврентия [Bishop,
Gendron, 1997].
Резкое повышение содержания нитратов в
поверхностных и грунтовых водах вызывает опасение, что это может привести к
сохранению амфибий.
Нам известны несколько статей по гемотоксичности
минеральных удобрений. Так, одна статья посвящена гемотоксичности удобрения
фосфата диаммония для рыбы змееголова Channa punctatus [Singh
R., Singh R., Verma, 1995]. Показано, что при концентрациях удобрения свыше
0,25 г/л и экспозиции от 24 до 96 часов отмечается снижение концентрации
гемоглобина в крови рыб, снижение числа эритроцитов и уменьшение гематокрита.
Число лейкоцитов в крови рыб возрастало по мере увеличения времени экспозиции к
веществу.
Содержание молоди рыбы катля Catla catla в
присутствии нитрита в концентрации до 4 мг/л в течение 6 часов приводило к
увеличению популяции незрелых эритроцитов до 7-24% [Effect
of sub-lethal … , 2004]. При повышении концентрации нитрита
до 8 или 10,4 мг/л этот эффект отсутствовал. Содержание молоди в присутствии
нитрита в концентрации 1-19,4 мг/л в течение 96 часов
вызывало снижение общей популяции эритроцитов на 21,2-31,8%.
Концентрация гемоглобина по мере увеличения дозы нитрита прогрессивно
снижалась. В течение первых 6 часов от начала воздействия нитрита количество
лейкоцитов падало, а в течение последующих 6 часов возрастало. Обращено
внимание на необходимость контроля за количеством нитрита, используемого в
качестве удобрения прудов для выращивания молоди рыб.
Есть статья о влиянии арсенита калия в модельных
экспериментах на организм гидробионтов на примере прудовой лягушки Rana
lessonae [Влияние арсенита калия…, 1994].
Лягушек после 6-суточной инкубации держали в аквариумах с водой, содержащей
арсенит калия в концентрации, соответствующей 4 ПДК. Изучали кислотную и
осмотическую резистентность эритроцитов, морфологические и морфометрические их
характеристики, лейкоцитарную формулу крови. Обнаружены изменения,
свидетельствующие об угнетении процессов гранулопоэза и некоторой активации
эритропоэза.
Влияние минеральных удобрений на земноводных,
судя по обзору литературы, изучали, главным образом, на ранние стадии
индивидуального развития земноводных (икру и личинки).
Наблюдая за земноводными в Германии, немецкие
экологи отметили в сельскохозяйственном регионе массовую гибель амфибий в
результате использования минеральных удобрений - азотных и
комплексных. Особенно серьезный урон понесли популяции обыкновенного Triturus
vulgaris и гребенчатого Tr. cristatus тритонов. Предлагается выделять буферные
зоны вокруг водоемов и увеличивать долю лугов на территориях, важных для
поддержания существования земноводных [Schneeweiss N.,
Schneeweiss U., 1997].
В более поздней работе тех же авторов отмечено
несколько случаев гибели земноводных в сельскохозяйственных районах в
окрестностях Берлина. Амфибии мигрировали в места размножения (март -
май) и из этих мест (август - октябрь). Установлено, что
причиной смерти были минеральные удобрения: кальциево-аммониевый нитрат
(азотистое удобрение); смесь кальциево-аммониевого нитрата и соединений фосфора
и калия; смесь, содержащая только соединения фосфора и калия. Даны рекомендации
по сохранению мест размножения амфибий в сельскохозяйственных ландшафтах [Schneeweiss
N., Schneeweiss U., 1999].
В лабораторных условиях показано, что в
скоплениях личинок серой жабы Bufo bufo 36-37 стадии развития
при высокой их двигательной активности в воде появляются экскреты, избегаемые
конспецифичными личинками. Вода с добавлением аммония в концентрациях 1 мг/л и
0,2 мг/л также вызывает реакцию избегания. Содержание аммония до 2 мг/л
характерно для всех водоемов, считающихся «чистыми», и не влияет на развитие и
рост головастиков. Сделан вывод, что повышение концентрации аммония (у личинок
это конечный продукт азотистого обмена) играет роль неспецифичного феромонного
сигнала о присутствии хищника. Избегание данного стимула увеличивает
вероятность выживания личинок амфибий. Неспецифичность такого сигнала
способствует его использованию не только во внутривидовой, но и в межвидовой
коммуникации [Мантейфель, Киселева, Марголис, 2004].
Головастики травяной Rana temporaria и остромордой R. arvalis лягушек на 37-39
стадиях развития воспринимают химические стимулы - экскреты
активно двигающихся конспецифических головастиков -
и повышение концентрации аммиака в воде не более чем на 1 мг/л и реагируют на
эти химические стимулы избеганием. Анализ экскретов личинок остромордой лягушки
выявил увеличение содержания аммиака на 0,925 мг/л. Возможно, аммиак является
по крайней мере одним из компонентов данного стимула, вызывающего у
конспецификов реакции избегания [Мантейфель, 2006].
Изучали влияние нитрата аммония, нитрата натрия
и мочевины на эмбрионов и личинок красноногой лягушки Rana aurora на
головастиков болотной квакши Pseudacris regilla и шпорцевой лягушки Xenopus
laevis. Показано, что нитрат аммония в концентрации >
105,5 мг/л и нитрата натрия в концентрации > 918 мг/л вызывали
100%-ную гибель эмбрионов. Экспозиция в более слабых растворах приводила к
уменьшению веса и длины эмбрионов и личинок. Реакция личинок двух
использованных видов на мочевину была сходной: поголовная гибель при очень
высокой концентрации 15 мг/л и снижение веса, и уменьшение длины после экспозиции
в менее концентрированных растворах. Наиболее сильным токсическим действием
обладает нитрат аммония, тогда как действие мочевины было самым слабым [Schuytema,
Nebecer, 1999].
Проведено исследование влияния мочевины на
пищедобывательное поведение, на выживаемость метаморфизировавшей молоди
наземных амфибий - западноамериканской жабы Bufo
boreas, каскадной лягушки Eleutherodactylus punctariolus, длиннопалой амбистомы
Ambystoma macrodactilum и желтобрюхого тритона Taricha granulosa.
Мочевина, или карбамид, CO(NH2)2 -
высококонцентрированное азотное удобрение. Показано, что молодь не способна
распознавать почву, обработанную удобрением в дозе 50 и 100 кгN/га, и избегать
ее, хотя избегали обработанную мочевиной туалетную бумагу. При этом
экспонирование в течение пяти дней на почве, обработанной мочевиной в дозе 100
кгN/га, приводило к снижению интенсивности питания и гибели сеголеток
бесхвостых, но не хвостатых амфибий [Juvenile amphibians
do not avoid…, 2001].
Согласно результатам других исследований, проведенных
британскими герпетологами на травяной лягушке, нитрат кальция практически не
влияет на увеличение смертности амфибий. Увеличение содержания нитрата аммония
в воде повышает смертность и число физических отклонений и приводит к
увеличению массы тела при метаморфозе [De Wijer, Watt,
Oldham, 1997].
Проведено сравнение толерантности к нитратам у
различных популяций травяной лягушки Rana temporaria [Johansson,
Rasanen., Merila, 2001]. Личинок лягушек из северных и
южных регионов Швеции содержали в присутствии нитрата натрия в концентрации до
5000 мкг/л до наступления метаморфоза. Показано, что наличие NO3 иона в высоких
концентрациях задерживало наступление метаморфоза, особенно у северной
популяции. Рост личинок северной популяции при воздействии NO3 иона также
незначительно замедлялся. Вне зависимости от концентрации нитрата выживаемость
личинок лягушек северной популяции оказалась по сравнению с южной популяцией
ниже. Авторы приходят к выводу, что нитраты в экологически значимых
концентрациях не оказывают существенного воздействия на развитие травяной
лягушки.
Провели оценку состояния популяций земноводных -
красноногой лягушки Rana aurora и бурой амбистомы Ambystoma gracile
(вылупляемость, уродства, выживаемость) в трех сельскохозяйственных и трех не
затронутых сельскохозяйственной деятельностью районах долины низовий реки
Фрейзер, Британской Колумбии (Канада). Показано, что процент вылупления у этих
видов во всех сельскохозяйственных зонах ниже, чем в районах, не затронутых
сельскохозяйственной деятельностью. Во всех сельскохозяйственных зонах были
высокие концентрации в воде аммиака (1,27 мг/л), фосфатов (до 8,14 мг/л) [Effects
of agricultural runoff …, 2002].
Изучалось воздействие удобрений с нитратом
аммония в воде на головастиков четырех видов амфибий: американской жабы Bufo
americanus, леопардовой лягушки Rana pipiens, трехполосой квакши Pseudacris
triseriata и крикливой лягушки Rana clamitans. В острых экспериментах (96
часов; 13,6 - 39,3 мг/л) смертность разных видов была
различной и наблюдалось значительное снижение массы тела. При хронических
воздействиях (100 дней; 2,5; 5,0; 10 мг/л) выживаемость головастиков
трехполосой квакши и леопардовой лягушки при дозе 10 мг/л была значительно
ниже, но выживаемость головастиков крикливой лягушки не изменилась. Воздействие
концентрации 10 мг/л значительно понизило число метаморфизировавших
головастиков трехполосой квакши, но не повлияло на сроки развития.
В связи с сокращением численности амфибий
исследовали влияние такого удобрения как нитрат аммония на личинок
обыкновенного тритона Triturus vulgaris [Watt, Oldham, 1995].
Личинок держали в контролируемых лабораторных условиях в четырех разных
концентрациях нитрата аммония: 50, 100, 200 и 500 мг/л. В самой низкой
концентрации у личинок не отмечено никаких отличий от контроля. В растворах 200
и 500 мг/л (время одноразовой экспозиции 24, 48 или 72 часа) животные были
заметно мельче контрольных при метаморфозе. В растворе концентрации 100 мг/л
личинки достоверно лучше питались по сравнению как с контрольными, так и с
личинками в растворах более высоких концентраций, однако достоверных размерных
отличий при метаморфозе отмечено не было. Выживаемость личинок оставалась
высокой во всех случаях. В отдельных резервуарах выдерживали дафнию (корм) в
таких же концентрациях. Изменение поведения дафнии было отмечено при
концентрациях 200 и 500 мг/л, причем в обоих случаях рачки достоверно чаще
держались в самой верхней части водяного столба.
В ходе лабораторных экспериментов оценивали
влияние загрязнения воды нитратом аммония на эмбрионы и личинок шести видов
европейских земноводных. Особенно высокой чувствительностью отличается
обыкновенная квакша Hyla arborea. Повышенная смертность у этого вида отмечается
уже на 80-й день даже при минимальной концентрации загрязнителя. У двух видов -
дискоязычной лягушки Discoglossus galganoi и обыкновенной жабы Bufo bufo -
повышение смертности отмечено на 15-й день. У остальных нитрат аммония не
вызывает изменений показателя смертности, но приводит (при высоких
концентрациях) к появлению аномалий или к замедлению роста. Таким образом,
применение удобрений может быть причиной уменьшения численности некоторых
земноводных в аграрных районах Пиренейского полуострова [Impact
of ammonium nitrate …, 2004].
В лабораторных экспериментах изучали связь между
стадией развития и чувствительностью к нитрату аммония у головастиков
дискоязычной лягушки, западной чесночницы Pelobates cultripes, камышовой жабы
Bufo calamita и средиземноморской квакши Hyla meridionalis: у двух первых видов
острые эффекты более выражены на ранних стадиях. Так, 100% только что
вылупившихся личинок западной чесночницы погибает в течение четырех часов после
воздействия реагентом, а его воздействие при той же концентрации на личинок
более старшего возраста приводит к гибели <4% особей. У камышовой жабы более
чувствительными оказываются головастики более поздних стадий развития. Отмечены
межвидовые различия по чувствительности личинок к нитрату аммония [Influence
of developmental stage … , 2006].
Нам известны работы по влиянию нитратов и
пестицида атразина (в том числе в сочетании друг с другом) на личиночное
развитие и формирование гонад у леопардовой лягушки Rana pipiens [Orton,
Carr, Handy, 2006]. Тестировали концентрации
загрязнителей, соответствующие встречающимся в природе. Не найдено влияния
тестируемых реагентов на массу и размеры тела, относительную длину конечностей
или сроки развития передних конечностей. Воздействие загрязнителей приводит к
повышению доли самок в потомстве (до 66,4%). В то же время доля интерсексов во
всех экспериментах остается на одинаково низком уровне. Соотношение полов
изменяется особенно значительно при совместном воздействии нитрата и атразина.
На квакше коралловопалой литории Litoria
caerulea изучали влияние нитратов на выживаемость и развитие головастиков.
Высокие концентрации нитратов замедляют темпы роста и дифференцировки, а также
повышают смертность. Авторы полагают, что нитраты оказывают сходное действие на
личинок амфибий в целом [Baker, Waights, 1994].
Исследовали действие нитрита на поведение и
метаморфоз каскадных лягушек Rana cascade. Экспозиция головастиков каскадных
лягушек в сублетальных дозах нитрита замедляла их трансформацию и меняла
поведение метаморфизирующих особей (они чаще выходили на мелководье и
высовывали голову из воды), но не влияла на время выхода, хотя у
экспонированных головастиков метаморфоз происходил при большей длине хвоста [Marco,
Blaustein, 1999].
Изучено действие 11 концентраций нитрита натрия
(от 0,08 до 103,4 мг/л) на личинок короткоголовой амбистомы Ambystoma texanum.
LC50 при 96часах экспозиции составила 1,09 мг/л, смертность 100% личинок -
при концентрации нитрита натрия 2,5 мг/л. Полученные данные говорят об очень
высокой чувствительности личинок этого вида амбистом к действию нитритов
(минимальная из известных 96-часовая LC50 нитритов для позвоночных -
0,7 мг/л для радужной форели) [Huey, Betinger, 1980].
В эксперименте анализировали воздействие нитрита
в концентрациях от 0,6 до 6,1мг/л на земноводных - тигровую
амбистому Ambystoma tigrinum tigrinum и лесную лягушку Rana sylvatica на разных
стадиях онтогенеза. У лесной лягушки нитрит значительно снижает успех
вылупления икринок (при 3,4мг/л нитрита) и даже при более низких концентрациях
(0,5мг/л) приводит к высокой смертности; у амбистомы такое действие вызывает
более высокая концентрация нитрита - 4,6 мг/л. При
постэмбриональном воздействии чувствительность к нитриту снижается. В связи с
этим отмечается, что изучение токсического эффекта, проводимое на более поздних
онтогенетических стадиях, может давать заниженную оценку вредных экологических
последствий загрязнения среды нитритом [Kerry, 2005].
В лабораторных условиях исследовали влияние
перхлората аммония в различных концентрациях на развитие яиц и личинок
шпорцевой лягушки Xenopus laevis. LC50 для пяти суток определена в 510 мг/л,
для 70-и суток - 223 мг/л. В зависимости от
концентрации перхлорат аммония тормозит метаморфоз, причем этот эффект
отмечается при минимальных концентрациях, более низких, чем отмечаются в
поверхностных водах. Таким образом, этот реагент представляет реальную угрозу
природным популяциям земноводных [Environmentally
relevant concentrations …, 2002].
Исследовали влияние гербицида атразина в
сублетальных концентрациях 40 и 320 мкг/д и нитрата натрия -
37 и 292 мг/л на метаморфоз у шпорцевой лягушки. Оценивали смертность и размеры
головастиков на стадии метаморфоза. Нитрат не влиял на смертность. Атразин
вызывал повышение смертности. Уменьшение размеров и массы тела и это влияние
усиливается с повышением его концентрации. Проанализированы результаты
одновременного воздействия двух агентов. Их синергический эффект при низкой
концентрации выражается в меньших, чем ожидалось, размерах тела головастиков.
При высокой концентрации одного из веществ отмечается антагонистический эффект -
размеры тела оказываются большими, чем ожидалось. Таким образом, отмечаемые в
природе концентрации атразина оказывают негативное воздействие на земноводных.
Кроме того, смеси сельскохозяйственных химикатов в сублетальных концентрациях
могут вызывать более сильный, чем предполагалось, отрицательный эффект на
популяции земноводных [Brown, Spence, 2003]
В следующей статье приведены результаты изучения
влияния бора и нитрита на успех вылупления личинок из икры амфибий. В
лабораторных условиях икру лесной лягушки Rana sylvatica, амбистомы Джефферсона
Ambystoma jeffersonianum, желтопятнистой амбистомы A. maculatum и американской
жабы Bufo americanus выращивали в среде с различным содержанием тетрабората (0,
50 и 100 мг/л) и нитрата натрия (от 10 до 49 мг/л), обычно встречающихся в
сточных водах. Нитрат натрия не оказал влияния на развитие икры. Высокие
концентрации бора привели к снижению вылупляемости у жабы и развитию уродливого
потомства у остальных трех видов. Хотя реальные концентрации бора и нитрата в
водоемах, откуда была взята икра, ниже использованных в лаборатории (1,5 и 26,7
мг/л соответственно), повышение содержания бора в нерестовых водоемах может
приводить к нарушениям в развитии икры [Laposata, Dunson,
1998].
В западных Гатах (Индия) сельскохозяйственная
деятельность фрагментирует лесные местообитания, и большинство лесных амфибий
становятся тесно связанными с сельскохозяйственными ландшафтами. Многие из них -
эндемики, один из которых южноиндийская лягушка Nyctibatrachus major.
Головастики южноиндийской лягушки подвергались сублетальному воздействию
нитратов от 100 до 5000 мкг/л до завершения метаморфоза. Повышение концентрации
нитратов приводило к уменьшению массы тела и сокращению времени, потраченного
на питание. При 1000-5000 мкг/л головастики плавали в
толще воды или вертикально, конвульсивно двигаясь без отдыха, или были
парализованы. Описаны дегенеративные морфологические изменения [Effects
of nitrate on feeding … , 2006].
Показано, что воздействие нитрата в концентрации
20 мг/л в течение 15 дней снижало жизнеспособность головастиков лягушки-быка
Rana catesbeiana и крикливой лягушки R. сlamitans; наиболее чувствительными к
воздействию нитрата оказались головастики крикливой лягушки. В концентрации 5
мг/л нитрат снижал прирост массы у головастиков лягушки-быка [Effects
of nitrate on the tadpoles …, 2005]. Содержание
головастиков лягушки-быка и крикливой лягушки в лабораторных условиях в
присутствии нитратов в течение 42 дней приводило к снижению выживаемости, но не
тормозило прироста массы тела. Присутствие головастиков крикливой лягушки
снижало выживаемость головастиков лягушки-быка дополнительно. Влияния нитрата
на выживаемость головастиков крикливой лягушки не обнаружено вне зависимости от
внутри- и межвидовых конкурентных отношений [Effects of nitrate
on the interactions …, 2006].
Содержание головастиков крикливой лягушки в
течение 95 дней в аквариуме в присутствии нитрата аммония в концентрации 10
мг/л или карбарила в концентрации 2.5 мг/л стимулировало прирост массы тела.
При совместном воздействии обоих препаратов отмечено торможение метаморфоза
головастиков и снижение их выживаемости. Авторы обсуждают необходимость учета
комплексного воздействия ксенобиотиков на развитие амфибий в водных экосистемах
[Multiple
sublethal …, 2005].
Оценивали влияние удобрения нитрата аммония,
инсектицида карбарила и широко распространенного водного болезнетворного
организма Saprolegnia ferax на вылупление и выживаемость головастиков
лягушки-быка в лаборатории. Карбарил значительно уменьшил выживаемость и
вылупляемость головастиков, но интерактивные аффекты среди стрессоров не были
найдены [Puglis,
Boone, 2007].
Две статьи Т.И. Жуковой [2009а,б]
посвящены результатам изучения влияния минеральных удобрений на смертность
головастиков, а также на темп роста головастиков малоазиатской лягушки Rana
macrocnemis. Из контрольных головастиков 56,7% животных завершили метаморфоз. В
опытах с концентрациями нитрата аммония 0,5; 1,0 и 2,0 мг/л количество
метаморфизировавших животных меньшее, чем в контроле, а именно -
16,7 -
23,3%. Следовательно, в растворах нитрата аммония выживает в 2,4 -
3,4 раза меньше головастиков. В самом концентрированном из исследованных
растворов удобрения (3,0 мг/л) все головастики погибли на 55-й день, не
завершив метаморфоза.
Если в контроле и растворах нитрата аммония
относительно низких из использованных концентраций (0,5; 1,0 и 2,0 мг/л)
численность остающихся в живых головастиков снижалась постепенно на протяжении
всего опыта, то в растворах нитрата аммония более высокой концентрации (3,0
мг/л) наблюдали резкое падение численности между 14 и 22 днями наблюдения.
Следовательно, даже непродолжительное воздействие нитрата аммония такой
концентрации может губительно сказываться на выживаемости головастиков
малоазиатской лягушки. В исследованных растворах фосфорного удобрения аммофоса,
как и в растворах азотного удобрения, общая выживаемость головастиков
малоазиатской лягушки очень существенно снижена по сравнению с выживаемостью в
контроле - в 8,5 раз (концентрация 0,1-0,2
мг/л) и в 17 раз (концентрация 0,5 мг/л); в растворе самой высокой из
исследованных концентраций (1,0 мг/л) погибли все головастики [Жукова,
2009а].
При посадке линейные размеры головастиков были
одинаковыми в контроле и растворах обоих удобрений. На 9-й и 14-й день
наблюдения длина тела головастиков также не отличается во всех емкостях.
Практически статистически достоверные различия длины тела контрольных и
подопытных животных на протяжении эксперимента были с 22 по 37-й дни
наблюдения. Например, на 22-й день головастики были крупнее в растворе нитрата
аммония концентрации 1,0 мг/л, на 27-й день - в растворе того же
удобрения концентрации 2,0мг/л и в растворе аммофоса концентрации 0,5 мг/л.
Автор объясняет тенденцию к увеличению размеров тела головастиков в
соответствующих растворах удобрений влиянием последних на более интенсивное
развитие водорослей, которые служат пищей личинкам земноводных. Однако эти
различия длины тела в дальнейшем не везде сохраняются. В частности, на 37-й
день наблюдения различия размеров головастиков в разных вариантах опыта и
контроле, как правило, находятся в пределах статистической ошибки. Самые
крупные метаморфизирующие животные в опыте были в самой низкой и самой высокой
из исследованных концентраций нитрата аммония. Действие нитрата аммония на
развивающихся головастиков малоазиатской лягушки проявляется и в замедлении
развития.
В контроле головастики завершили метаморфоз на
55-й день наблюдения, что соответствовало 63 дню развития. В опытах с
концентрациями нитрата аммония 0,5; 1,0 и 2,0 мг/л, а также аммофоса 0,2 и
0,5мг/л метаморфоз закончился позднее - на 63-й день
наблюдения (71-й день развития). И только в самом слабом растворе аммофоса -
0,1мг/л, метаморфоз произошел одновременно с контрольными животными [Жукова,
2009б].
Таким образом, неконтролируемое применение
химических веществ приводит к сокращению численности многих видов земноводных
как бесхвостых (таких исследований подавляющее большинство), так и хвостатых.
При длительном поступлении в организмы животных низких концентраций различных
токсикантов и при их накоплении в жизненно важных органах возникают внешне
долгое время не проявляющиеся биохимические и физиологические изменения,
которые в определенный период жизни животных (миграция, размножение), а также
при стрессовых ситуациях (болезни, ухудшение погодных условий или питания)
вызывают изменение поведенческих реакций, нарушение воспроизводительной
способности, снижение жизненности потомства, что подрывает устойчивость
популяций к неблагоприятным воздействиям среды [Воронова,
Денисова, Пушкарь, 1987].
Обзор литературы показал, что воздействие
минеральных удобрений на земноводных остается мало изученным. Известные нам
работы посвящены исследованию влияния некоторых удобрений (чаще всего нитрата
аммония различных концентраций) и проведены на разных объектах, что затрудняет
сравнение полученных результатов. Этим и объясняется выбор темы данной
дипломной работы.
2. Материал и методы исследования
Исследования проводили на личинках озерной
лягушки.
В водоемах Ботанического сада КубГУ были собраны
головастики озерной лягушки. Эксперимент был начат 02.06.16г. с головастиками
восьмидневного возраста, на 37-й стадии развития [Дабагян,
Слепцова, 1975]. Эксперимент проводился до гибели головастиков
или метаморфоза.
В лабораторных условиях был поставлен опыт по выявлению
влияния на личиночное развитие озерной лягушки минеральных удобрений.
Контролем были личинки указанного вида,
содержащиеся в чистой, отстоянной водопроводной воде; растворы минеральных
удобрений всех использованных в опыте концентраций также были приготовлены на
отстоянной воде.
Исходя из литературных данных, наиболее часто
попадают в природные системы из агроценозов нитраты и фосфорные удобрения. В
наибольшей степени изучено действие нитратов на земноводных, однако, их
действие выяснялось для других видов амфибий.
Мы использовали следующие минеральные удобрения:
нитрат аммония NH4NO3 (процентное содержание усвояемого азота в этой соли выше,
чем в других нитратах или солях аммония), или аммиачная селитра; и суперфосфат,
который представляет собой смесь сульфата кальция CaSO4 с дигидрофосфатом
кальция Ca(H2PO4)2 [Глинка, 1985].
Азотные удобрения в зависимости от формы
соединения азота можно разделить на следующие виды: 1)аммиачные -
азот содержится в основном в виде свободного аммиака (водный аммиак, аммиачная
вода -
раствор аммиака в воде); 2)аммонийные - азот присутствует
в виде иона аммония, связанного с какой-нибудь кислотой (сульфат аммония -
(NH4)2SO4, хлорид аммония - NH4Cl); 3)нитратные,
содержащие азот в окисленной форме в виде солей азотной кислоты (нитрат натрия,
чилийская селитра - NaNO3, нитрат кальция, норвежская
селитра -
Cа(NO3)2); 4)аммонийно-нитратные - азот присутствует
одновременно в аммонийной и нитратной формах (аммиачная селитра, нитрат аммония
-
NH4NO3); 5)амидные - азот находится в органической форме
в виде амидов (мочевина, карбамид - CO(NH2)2; цианамид
кальция -
CaCN2 [Филимонов, 1976].
Азот является важнейшим элементом для развития
растений, в частности, для образования белковых веществ. Содержание азота в
составе белка колеблется в пределах 15-19%. Азот входит в
состав хлорофилла и, следовательно, принимает участие в фотосинтезе, а также
ферментов - катализаторов жизненных процессов в организме
растений и животных. В почву азот поступает в аммиачной, амидной и нитратной
формах. При недостатке углеводов растение не может использовать азот для
образования аминокислот и белков. Практикой и опытами установлено, что
аммиачный азот может быть использован растениями более эффективно, чем
нитратный, если устранено побочное влияние кислотности аммонийных солей
[Соколовский, Унанянц, 1977].
Подбирая концентрации удобрения для
эксперимента, мы исходили из норм для этих веществ, принятых в рыбоводстве [Москул,
1998].
Для азота аммонийного и нитратов технологическая
норма 0,2 - 1,0 г/м3 (мг/л), а допустимые значения -
до 2,0 и 3,0 г/м3 (мг/л) соответственно. Для фосфатов технологическая норма 0,2
-
0,5 г/м3 (мг/л), а допустимые значения - до 2,0 г/м3
(мг/л).
Принимая во внимание эти нормы, мы взяли для
опыта следующие концентрации: нитрата аммония - 5,00; 10,0
и 20,0 мг/л; для суперфосфата - 10,0; 50,0 и 100,0 мг/л.
Плотность посадки головастиков амфибий —
10 особей на 1 литр, использовали трехлитровые емкости, в каждую из которых
посадили по 30 головастиков.
Во все сосуды добавили растения (для
прикрепления личинок и их начального питания). Головастиков кормили вареными
листьями одуванчиков (свежие листья одуванчиков заливали кипятком, кипятили 3 -
4 минуты, а затем остужали). В конце эксперимента переходили на кормление
головастиков вареным желтком.
Раз в 5 - 10 дней мы
подсчитывали число погибших личинок и удаляли их, при этом уменьшали объем воды
для поддержания постоянной плотности. Растворы заменяли также 1 раз в 5 -
10 дней. Температуру воды поддерживали в пределах + 23—25°С.
С конца мая была аномально высокая для города Краснодара температура воздуха,
вода в емкостях тоже стала нагреваться (до + 28-29°С),
мы при этом применяли аэратор для насыщения воды кислородом.
Мы фиксировали сроки наступления основных стадий
развития и метаморфоза [Дабагян, Слепцова, 1975].
Общую длину тела личинок соответствующих стадий и метаморфизирующих животных
измеряли штангенциркулем, с точностью до 0,1 мм (сначала измеряли по 7 -
8 особей из каждой емкости, а затем всех оставшихся в живых особей). Личинок
взвешивали на аналитических весах.
Цифровой материал по длине тела головастиков
обработан стандартными статистическими методами [Лакин, 1990].
Достоверность различий показателей в контроле и
разных вариантах опыта определяли с помощью критерия Стьюдента. Различия
считали достоверными, если tфакт. ³ tст. при 5%-ном
уровне значимости.
Опыты продолжали до наступления метаморфоза или
до гибели всех личинок в соответствующем растворе. Всех животных, выживших и
закончивших метаморфоз, мы выпустили в естественные места обитания.
Всего в эксперимент были взяты 210 головастиков
озерной лягушки.
. Влияние аммиачной селитры и суперфосфата на
личиночное развитие озерной лягушки
.1 Смертность головастиков озерной лягушки в
растворах аммиачной селитры и суперфосфата
Данные по изменению численности головастиков
озерной лягушки в контроле и растворах аммиачной селитры (нитрата аммония) и
суперфосфата различных концентраций в процессе эксперимента приведены на
рисунке 1.
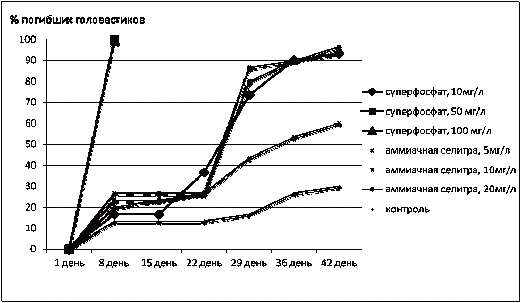
Рисунок 1 - Смертность
головастиков озерной лягушки (%) в контроле и эксперименте
В контроле гибель головастиков началась на 8-й
день наблюдения и составила 13%, этот уровень смертности оставался без
изменений до 29 дня, затем процент погибших увеличился до 19% и продолжал
возрастать до 42 дня наблюдений, когда общая смертность составила 29%. Таким
образом, в контроле 71% головастиков завершили метаморфоз.
В растворе аммиачной селитры самой низкой из
исследованных концентраций (5 мг/л) гибель головастиков началась также на 8-й
день, но была большей, чем в контроле почти вдвое -
24%. Далее на этом уровне сохранялась смертность до 22 дня, а затем резко
выросла к 29 дню до 42% и далее плавно возрастала, составив к 42 дню 60%
погибших. Таким образом, закончили метаморфоз в этом растворе селитры 40%
головастиков.
И в контроле, и в растворе нитрата аммония
концентрации 5 мг/л кривые гибели головастиков были плавными.
В растворах аммиачной селитры 10 и 20 мг/л
кривые гибели головастиков очень похожи, а именно -
на 8-й день погибли 20 и 21% головастиков, затем гибли единичные особи -
на 22-й день смертность была 24%, а далее произошел резкий подъем кривой вверх -
на 29-й день смертность составила 80 и 84%, а к 42-му дню погибли 96 и 97%.
Закончили в этих растворах метаморфоз 3 и 4% головастиков озерной лягушки.
В самой низкой из исследованных концентраций
суперфосфата 10 мг/л на 8-й день погибли 16% головастиков, этот уровень
сохранялся до 15 дня, а затем смертность резко увеличилась -
на 22-й день 38% погибших, на 29-й день 76%, а к 42-му дню погибли 96%
головастиков. Закончили метаморфоз 4% головастиков озерной лягушки.
В более концентрированных растворах суперфосфата
(50 и 100 мг/л) все 100% посаженных в растворы головастиков погибли на 8-й день
эксперимента.
Можно сделать вывод, что наименее опасен для
головастиков озерной лягушки раствор аммиачной селитры концентрации 5 мг/л
(выжили 40% животных), растворы более высоких концентраций аммиачной селитры
(10 и 20 мг/л) и наименее концентрированный из исследованных раствор суперфосфата
(10 мг/л) очень существенно влияют на выживаемость головастиков озерной лягушки
в равной степени - завершают метаморфоз 3-4%
головастиков. Более концентрированные растворы суперфосфата вызывают очень
быструю (в течение недели) и 100%-ную гибель головастиков озерной лягушки.
Из литературы известно, что нитрат аммония в
концентрации > 105,5 мг/л вызывал 100%-ную гибель эмбрионов
красноногой лягушки Rana aurora [Schuytema, Nebecer,
1999].
Изучалось воздействие удобрений с нитратом аммония в воде на головастиков
четырех видов амфибий: американской жабы Bufo americanus, леопардовой лягушки
Rana pipiens, трехполосой квакши Pseudacris triseriata и крикливой лягушки Rana
clamitans. Выживаемость головастиков трехполосой квакши и леопардовой лягушки
при дозе 10 мг/л была значительно ниже, но выживаемость головастиков крикливой
лягушки не изменилась. Воздействие концентрации нитрата аммония 10 мг/л
значительно понизило число метаморфизировавших головастиков трехполосой квакши.
Нитрат аммония оказывал токсическое воздействие на головастиков всех видов,
хотя использованные концентрации были ниже применяемых в сельском хозяйстве
Канады [Hecnar,
1995].
Оценивали влияние загрязнения воды нитратом
аммония на эмбрионы и личинок шести видов европейских земноводных. Особенно
высокой чувствительностью отличается обыкновенная квакша Hyla arborea.
Повышенная смертность у этого вида отмечается уже на 80-й день даже при
минимальной концентрации загрязнителя. У двух видов -
дискоязычной лягушки Discoglossus galganoi и обыкновенной жабы Bufo bufo -
повышение смертности отмечено на 15-й день [Impact of ammonium
nitrate….., 2004].
Из контрольных головастиков малоазиатской
лягушки Rana macrocnemis 56,7% животных завершили метаморфоз. В опытах с
концентрациями нитрата аммония 0,5; 1,0 и 2,0 мг/л количество
метаморфизировавших животных меньшее, чем в контроле, а именно -
16,7 -
23,3%. Следовательно, в растворах нитрата аммония выживает в 2,4 -
3,4 раза меньше головастиков. В самом концентрированном из исследованных
растворов удобрения (3,0 мг/л) все головастики погибли на 55-й день, не
завершив метаморфоза. Даже непродолжительное воздействие нитрата аммония такой
концентрации может губительно сказываться на выживаемости головастиков
малоазиатской лягушки [Жукова, 2009а].
Другие данные получены при исследовании влияния
нитрата аммония (концентрации от 59 до 599 мг/л) на личинок обыкновенного
тритона Triturus vulgari. В самой низкой концентрации у личинок не отмечено
никаких отличий от контроля. Выживаемость личинок оставалась высокой во всех
случаях [Watt,
Oldham, 1995].
Сравнение наших и литературных данных по влиянию
нитрата аммония на смертность личинок земноводных показало, что хотя действие
указанного удобрения является специфическим для каждого вида амфибий, но для
большинства видов нитрат аммония уменьшает выживаемость личинок.
Что касается влияния суперфосфата на смертность
личинок озерной лягушки, то ее можно сравнить с литературными данными по
смертности личинок малоазиатской лягушки в присутствии фосфорного удобрения
аммофоса [Жукова, 2009а]. В исследованных
растворах аммофоса общая выживаемость головастиков малоазиатской лягушки очень
существенно снижена по сравнению с выживаемостью в контроле -
в 8,5 раз (концентрация 0,1-0,2 мг/л) и в 17 раз
(концентрация 0,5 мг/л); в растворе самой высокой из исследованных концентраций
(1,0 мг/л) погибли все головастики. Следовательно, более сильное воздействие
оказывает фосфорное удобрение по сравнению с азотным.
.2 Темп роста головастиков озерной лягушки в
растворах аммиачной селитры и суперфосфата
Изменения длины тела и массы головастиков в
контроле и растворах минеральных удобрений показаны в таблицах 1-4.
Таблица 1 - Темп линейного
роста (мм) головастиков озерной лягушки в контроле и растворе аммиачной селитры
|
День
наблю-дения
|
Контроль
|
Аммиачная
селитра, 5 мг/л
|
|
Пределы
|
X
± m
|
Прирост
за день
|
Пределы
|
X
± m
|
|
1
|
8,0- 13,0
|
12,0±1,23
|
-
|
8,0-13,0
|
10,4±0,72
|
-
|
|
8
|
10,0-16,0
|
13,8±1,12
|
0,26
|
10,0- 15,0
|
12,8±0,81
|
0,34
|
|
15
|
15,0-25,5
|
21,9±1.88
|
1,16
|
17,0-22,5
|
19,6±0,62
|
0,97
|
|
22
|
16,4-37,5
|
25,6±2,50
|
0,17
|
23,5-40,0
|
31,6±0.81
|
1,71
|
|
29
|
30,8-55,5
|
45,3±3,58
|
2,81
|
30,5-55,0
|
44,6±2,20
|
1,86
|
|
36
|
41,5-53,3
|
47,4±2,08
|
0,27
|
25,5-40,0
|
29,1±2,03
|
-2,21
|
|
42
|
35,5-44,1
|
40,2±1,95
|
-1,03
|
18,0-25,0
|
21,8±0,74
|
-1,04
|
Таблица 2 - Темп линейного
роста (мм) головастиков озерной лягушки в контроле и растворе суперфосфата
|
День
наблю-дения
|
Контроль
|
Суперфосфат,
10 мг/л
|
|
Пределы
|
X
± m
|
Прирост
за день
|
Пределы
|
X
± m
|
Прирост
за день
|
|
1
|
8,0
- 13,0
|
12,0
± 1,23
|
-
|
8,0
- 13,0
|
11,2
±0,98
|
-
|
|
8
|
10,0
- 16,0
|
13,8
± 1,12
|
0,26
|
13,0±1,00
|
0,26
|
|
15
|
15,0
- 25,5
|
21,9
± 1,88
|
1,16
|
15,0-23,0
|
19,5±1,90
|
0,93
|
|
22
|
16,4
- 37,5
|
25,6
± 2,50
|
0,17
|
17,0-28,5
|
23,7±2,35
|
0.60
|
|
29
|
30,8
- 55,5
|
45,3
± 3,58
|
2,81
|
30,5-50,0
|
39,8±3,43
|
2,30
|
|
36
|
41,5
- 53,3
|
47,4
± 2,08
|
0,27
|
30,5-62,0
|
46,1±5,24
|
0.90
|
|
42
|
35,5
- 44,1
|
40,2
± 1,95
|
-1,03
|
23,0-28,5
|
25,5±1,06
|
-2,80
|
В таблицах приведены данные по растворам
минеральных удобрений тех концентраций, при которых развитие головастиков
озерной лягушки продолжалось достаточно долго, это самые низкие из
исследованных концентраций - 5 мг/л для аммиачной селитры и
10 мг/л для суперфосфата.
При посадке линейные размеры головастиков
озерной лягушки были одинаковыми в контроле и растворах обоих удобрений (8,0-13,0
мм) -
Таблицы 1-2.
На 8-й и 15-й день длина тела головастиков также
была одинаковой в контроле и растворах исследованных удобрений, различия
находятся в пределах статистической ошибки (t = 0,98 и 0,85 при tst = 2,09). На
22-й день длина головастиков из контроля не имела статистически достоверных
различий с головастиками в растворе суперфосфата (t = 0,87 при tst = 2,09), но
в растворе аммиачной селитры головастики достоверно больше по размерам (t =
2,28 при tst = 2,09). На 29-й день контрольные головастики были одинаковыми по
длине с головастиками из обоих растворов минеральных удобрений. На 36-й день
контрольные головастики не отличались по размерам от головастиков из раствора
суперфосфата, но статистически достоверно (в 1,6 раза) превосходили
головастиков из аммиачной селитры (t = 6,30 при tst = 2,09). В последний, 42-й
день наблюдения, головастики из контроля по длине тела превышали подопытных
животных в 1,8 раза (аммиачная селитра) и в 1,6 раза (суперфосфат), t = 4,03 и
6,62 при tst = 2,09.
Прирост длины тела с 1 по 22-й день был
равномерным у головастиков из контроля и раствора суперфосфата (а в растворе
аммиачной селитры с 1 по 15-й день). Между 22 и 29-м днями отмечен максимальный
ежедневный прирост длины тела у всех головастиков. Затем между 29 и 36-м днями
прирост длины тела резко падал, а в последние 7 дней наблюдалось уменьшение
размеров тела - за счет рассасывания хвоста (метаморфоз).
Следовательно, можно констатировать влияние
исследованных удобрений на уменьшение линейных размеров головастиков озерной
лягушки в нашем эксперименте.
Таблица 3 - Темп весового роста
(мг) головастиков озерной лягушки в контроле и растворе аммиачной селитры
|
День
наблю-дения
|
Контроль
|
Аммиачная
селитра, 5 мг/л
|
|
Пределы
|
X
± m
|
Прирост
за день
|
Пределы
|
X
± m
|
Прирост
за день
|
|
1
|
44,5
- 142,1
|
99,6
± 16,29
|
-
|
60,0
- 135,0
|
101,8
± 9,02
|
_
|
|
8
|
46,0
- 155,4
|
96,5
± 17,31
|
-0,44
|
60,0
- 145,5
|
112,4
± 9,48
|
1,51
|
|
15
|
86,5
- 159,5
|
117,6
± 10,02
|
3,01
|
110,7
± 7,32
|
-0.24
|
|
22
|
120,3
- 1100,5
|
653,9
± 176,18
|
76,6
|
150,5
- 1030,0
|
726,2
± 83,11
|
87,92
|
|
29
|
155,0
- 1143,3
|
771,7
± 190,04
|
16,74
|
530,0
- 2380,0
|
1739,0
± 187,20
|
144,68
|
|
36
|
140,5
- 1430,5
|
961,5
± 199,02
|
27,11
|
1050,0
- 3150,5
|
2568,0
± 200,05
|
118,42
|
|
42
|
185,0
- 3224,5
|
1488,1
± 544,20
|
75,23
|
1100,0
- 3200,0
|
2655,0
± 227,82
|
12,43
|
По массе тела головастики озерной лягушки были
одинаковыми на 1-22-й дни наблюдений (таблицы 3-4).
На 29-й день наблюдения головастики из контроля и раствора суперфосфата не
отличались по массе тела (t = 0,56 при tst = 2,09), а в растворе аммиачной
селитры головастики были достоверно более тяжелыми -
в 2,25 раза (t = 3,60 при tst = 2,09). Эти различия сохранялись до конца
наблюдений при сравнении контрольных головастиков и головастиков из аммиачной
селитры. В 36-42-й дни были тяжелее контрольных и головастики
в растворе суперфосфата (t = 3,72 и 3,80 при tst = 2,09).
Таблица 4 - Темп весового
роста (мг) головастиков озерной лягушки в контроле и растворе суперфосфата
|
День
наблю-дения
|
Контроль
|
Суперфосфат,10
мг/л
|
|
Пределы
|
X
± m
|
Прирост
за день
|
Пределы
|
X
± m
|
Прирост
за день
|
|
1
|
44,5
- 142,1
|
99,6
± 16,29
|
-
|
42,0
- 140,0
|
96,5
± 8,37
|
-
|
|
8
|
46,0
- 155,4
|
96,5
± 17,31
|
-0,44
|
42,5
- 146,0
|
103,7
± 9,66
|
1,03
|
|
15
|
86,5
- 159,5
|
117,6
± 10,02
|
3,01
|
67,0
- 146,0
|
102,2
± 10,23
|
-0,21
|
|
22
|
120,3
- 1100,5
|
653,9
± 176,18
|
76,6
|
110,0
- 1020,0
|
634,0
± 70,10
|
75,97
|
|
29
|
771,7
± 190,04
|
16,74
|
160,5
- 1230,0
|
782,5
± 85,44
|
21,21
|
|
36
|
140,5
- 1430,5
|
961,5
± 199,02
|
27,11
|
450,0
- 2730,0
|
2223,9
± 258,82
|
205,91
|
|
42
|
185,0
- 3224,5
|
1488,1
± 544,20
|
75,23
|
452,0
- 3100,0
|
2079,2
± 184,63
|
-31,59
|
Таким образом, исследованные удобрения влияют на
повышение массы завершающих метаморфоз головастиков озерной лягушки.
По литературе, отмечена тенденция к увеличению
длины тела головастиков малоазиатской лягушки под влиянием удобрений нитрата
аммония и аммофоса [Жукова, 2009б].
Автор объясняет тенденцию к увеличению размеров тела головастиков в
соответствующих растворах удобрений влиянием последних на более интенсивное
развитие водорослей, которые служат пищей личинкам земноводных. Мы полагаем,
что это вполне возможное объяснение наблюдаемого и нами увеличения размеров
тела головастиков озерной лягушки в растворах низких концентраций аммиачной
селитры и суперфосфата.
Следовательно, происходит ускорение роста
личинок озерной и малоазиатской лягушек в присутствии нитрата аммония.
В литературе есть различные сведения о влиянии
минеральных удобрений на линейный и весовой рост земноводных. Так, содержание
головастиков крикливой лягушки в течение 95 дней в аквариуме в присутствии
нитрата аммония в концентрации 10 мг/л стимулировало прирост массы тела. [Multiple
sublethal…, 2005]. В то же время у ряда видов
европейских бесхвостых земноводных (в аграрных районах Пиренейского
полуострова) нитрат аммония приводит (при высоких концентрациях) к замедлению
роста (Impact of ammonium nitrate ……, 2004). Экспозиция в слабых растворах
нитрата аммония приводила к уменьшению веса и длины эмбрионов и личинок
красноногой лягушки Rana aurora, на головастиков болотной квакши Pseudacris
regilla и шпорцевой лягушки Xenopus laevis [Schuytema, Nebecer,
1999].
Нитрат аммония в острых экспериментах (96 часов;
13,6 -
39,3 мг/л) влиял на головастиков американской жабы, леопардовой лягушки,
трехполосой квакши и крикливой лягушки, значительно снижая массу тела и вызывая
аномалии развития [Hecnar, 1995].
В концентрации нитрата аммония 50 мг/л у личинок обыкновенного тритона не
отмечено никаких отличий от контроля. В растворах нитрата аммония 200 и 500
мг/л личинки были заметно мельче контрольных при метаморфозе. В растворе
концентрации 100 мг/л личинки достоверно лучше питались по сравнению как с контрольными,
так и с личинками в растворах более высоких концентраций, однако достоверных
размерных отличий при метаморфозе отмечено не было [Watt,
Oldham, 1995].
Таким образом, нитрат аммония на разных видах
земноводных может вызывать прямо противоположные эффекты (уменьшение или
увеличение длины и/или массы тела при метаморфозе).
Заключение
В последние 30 лет природные экосистемы
загрязняются значительными количествами минеральных удобрений, которые дождевой
водой смываются в водоемы с сельскохозяйственных полей, а также вносятся в
пруды для интенсификации искусственного разведения рыбы. Минеральные удобрения
даже в небольших дозах в водоемах отрицательно влияют на большинство
гидробионтов, в частности, на земноводных, для которых характерно размножение в
воде и обязательное обитание в воде на стадиях икринки и личинки. На основании
проведенных экспериментов можно сделать следующие выводы.
. Смертность головастиков озерной лягушки в
растворах аммиачной селитры увеличивается с возрастанием концентрации минерального
удобрения: в концентрации 5 мг/л выжили и метаморфизировали 40% головастиков, а
в концентрациях 10 и 20 мг/л только 4 и 3% соответственно. В контроле
метаморфизировали 71% головастиков. В растворе суперфосфата самой низкой из
исследованных концентраций - 10 мг/л -
закончили метаморфоз 4% головастиков, а в более высоких концентрациях (50 и 100
мг/л) все головастики погибли через 7 дней после начала опыта.
2. Метаморфоз головастиков озерной лягушки в
растворе аммиачной селитры происходит при длине тела 21,8 ±
0,74 мм, что в 1,8 раза меньше, чем у контрольных животных, а в растворе
суперфосфата при длине тела 25,5 ± 1,06 мм, что
меньше контрольных значений в 1,6 раз. Животные заканчивают метаморфоз при
массе тела, превышающей контрольные значения в 1,8 раза (аммиачная селитра) и
1,4 раза (суперфосфат). Таким образом, темп линейного роста в растворах
удобрений меньше, а темп весового роста больше, чем в контроле.
. Сравнение воздействия аммиачной селитры и
суперфосфата на головастиков озерной лягушки показало, что оба исследованные
удобрения оказывают сходное воздействие, вызывая их гибель (96-97%) в
концентрации 10мг/л. Концентрация аммиачной селитры 5 мг/л является
сублетальной. Более высокие концентрации суперфосфата. Различий во влиянии на
темп линейного и весового роста головастиков в растворах двух исследованных
удобрений не отмечено.
Библиографический список
Булатова
Н.В. К вопросу о применении удобрений в выростных прудах Билейского
рыбопитомника // Информационные материалы Института экологии растений и
животных. Свердловск, 1976. С.42 - 43.
Велдре
И.А., Зоома М.Я. Токсическое воздействие нитритов на рыб // Экология.1990, № 1.
С.71-73.
Влияние
арсенита калия на организм гидробионтов на примере лягушки прудовой в модельных
экспериментах in vivo / Гусев Н.И. [и др.] // Вестн. Удм. ун-та. 1994.
Спецвыпуск. С.96-108.
Воронова
Л., Денисова А., Пушкарь И. Оценка загрязненности фауны в мониторинге состояния
природной среды // Проблемы фонового мониторинга состояния природной среды. Л.,
1987. Вып.5. С.84-91.
Глинка
Н.Л. Общая химия . Л.:Высшая школа, 1985. 704 с.
Дабагян
Н.В., Слепцова Л.А. Травяная лягушка Rana temporaria // Объекты биологии
развития. М.:Наука, 1975. С.442 - 462.
Жукова
Т.И. Влияние минеральных удобрений на смертность головастиков малоазиатской
лягушки (Rana macrocnemis Boul.) // Актуальные вопросы экологии и охраны
природы экосистем южных регионов России и сопредельных территорий. Краснодар,
2009а. С.55-57.
Жукова
Т.И. Влияние минеральных удобрений на темп роста головастиков малоазиатской
лягушки (Rana macrocnemis Boul.) // II Всероссийская научно-практическая
конференция. Махачкала, 2009б. С.
Лакин
Г.Ф. Биометрия. М.: Высшая школа, 1980. 293 с.
Мантейфель
Ю.Б. Личинки травяной и остромордой лягушек Rana temporaria и R. arvalis
избегают зоны повышенного содержания аммиака // Доклады РАН, 2006. 411, №1.
С.134-136.
Мантейфель
Ю.Б., Киселева Е.И., Марголис С.Э. повышение концентрации аммония как
неспецифичный феромонный сигнал, вызывающий избегание у личинок амфибий
//Водные экосистемы и организмы-6:Труды 6 научной конференции. М., 2004ю С.58.
Москул
Г.А. Приусадебное и фермерское рыбоводство. Краснодар. 1998. 134 с.
Пескова
Т.Ю. Влияние антропогенных загрязнений среды на земноводных Волгоград: Изд-во
ВГПУ. 2001. 156 с.
Соколовский
А.А., Унанянц Т.П. Краткий справочник по минеральным удобрениям. М.: Химия,
1981. 376с.
Филимонов
Д.А. Азотные удобрения. М.: Россельхозиздат. 1976. 40 с.J.M., Waights V.
Effects of nitrate on tadpoles of the tree frog (Litoria caerulea) // Herpetol.
J. 1994. 4. N 3. Р.106 - 108. РЖ Биология 1995 1И5093.Ch.,
Gendron А. Impacts of chemical pollutants on amphibian populations // Herpetol.
97: Abstr. 3rd World Congr. Herpetol., Prague, 2-10 Aug.,
1997. Prague, 1997. P.1-2. РЖ Биология 98.11-04И5.61.S. K.,
Spence K.M. Effects of subletal concentrations of atrazine and nitrate on
metamorphosis of the African clawed frog // Environ. Toxicol.
and Chem. 2003. 22, N3. P.627-635.of agricultural
runoff on native amphibians in the Lower Fraser River Valley, British Columbia,
Canada / De Sola Sh.R. [et al.] // Environ. Toxicol. and Chem. 2002.
21. №2. Р.353 -360. РЖ Биология 03.05-04И5.67.of nitrate on
feeding and resting of tadpoles of Nyctibatrachus major (Anura, Ranidae) / S.V.
Krishnamurthry [et al.] //Ecotoxicol.
2006. 12, N 3. P. 123-127. РЖ
Биология
09.04-04И5.84.of
nitrate on the interactions of the tadpoles of two ranids (Rana caresbeiana and
R. clamitans) / G.R. Smith [et al.]//
Aquat. Ecol. 2006. 40, N 1. P.125-130. РЖ
Биология
07.12-04И5.25.of
nitrate on the tadpoles of two ranids (Rana caresbeiana and R. clamitans) /
G.R. Smith [et al.] //
Arch. Environ. Contam. and Toxicol. 2005. 49, N 4. P.559-562.
РЖ Биология 07.04-04И5.40.of sub-lethal nitrite on selected hematological
parameters in fingerling Catla catla (Hamilton) / Ch. Pratar [et
al.]
// Aquacult. Res. 2004. 35, N 9. P.874-880.relevant
concentrations of ammonium perchlorate inhibit development and metamorphosis in
Xenopus laevis / W.L. Goleman [et al.] // Environ. Toxicol. and Chem. 2002. 21.
№ 2. Р.424 - 430. РЖ Биология 03.05-04И5.71.S.J. Acute and
chronic toxicity of ammonium nitrate fertilizer to amphibians from southern
Ontario // Environ. Toxicol. and Chem. 1995. 14. №
12. Р.2131
-
2137. РЖ
Биология
97.04-04И5.153.D.W.,
Betinger T.L. Toxicity of nitrite to larvae of the salamander Ambystoma texanum
// Bull. Environ. Contam. and Toxicol. 1980. 24. №6. Р.909
-
912. РЖ
Биология
1981. 8И53.of
developmental stage on sensitivity to ammonium nitrate of aquatic stages of
amphibians / M.E. Ortiz-Santaliestra [et al.] // Environ. Toxicol. and Chem.
2006. 25. N 1. P.105 - 111. РЖ
Биология
06.10-04И5.93.
Impact
of ammonium nitrate on growth and survival of six European amphibians / M.E.
Ortiz [et al.] // Arch. Environ. Contam. and Toxicol. 2004. 47. N 2. P.234 -
239. РЖ Биология 06.04-04И5.115.amphibians do not avoid potentially lethal
levels of urea on soil substrate / A.C. Hatch [et al.] // Environ. Toxicol.
and Chem. 2001. 20. N 10. Р.2338
-
2335. РЖ
Биология
02.11-04И5.110.M.,
Rasanen К.,
Merila J. Comparison of nitrate tolerance between different populations of the
common frog, Rana temporaria // Aquat. Toicol. 2001. 54. N 1-2. Р.1-14.
РЖ
Биология
04.03-04И5.72.
Kerry
I. Ontogenetic delays in effects of nitrite exposure on tiger salamanders
(Ambystoma tigrinum tigrinum) and wood frogs (Rana sylvatica) // Environ.
Toxicol. and Chem. 2005. 24. N 3. P.1523 - 1527. РЖ Биология
06.05-04И5.91.M.M., Dunson W.A. Effects of boron and nitrate on hatching
success of amphibian eggs // Arch. Environ. Contam. and Toxicol. 1998. 35. № 4.
Р.615 -
619. РЖ Биология 99.09-04И5.114.A., Blaustein A.R. The effects of nitrite on
behavior and metamorphosis in cascades frogs (Rana cascade) / А. Marco //
Environ. Toxicol. and Chem. 1999. 18. N 5. Р.946 - 949. РЖ
Биология 00.11-04И5.93.sublethal chemicals negatively affect tadpoles of the
green frog, Rana clamitans / M.D. Boone [et al.]
// Environ. Toxicol. and Chem. 2005. 24. N5. Р.1267 -
1272. РЖ Биология 07.07-04И5.56.F., Carr J.A., Handy R.D. Effects of nitrate
and atrazine on larval development and sexual differentiation in the northern
leopard frog Rana pipiens // Environ. Toxicol. and Chem. 2006. 25. N1. P.65-71.
РЖ Биология 06.10-04И5.91.H,J,, Boone M.D. Effects of fertilizer, an
insecticide, and a pathogenic fungus on hatching and survival of bullfrog (Rana
catesbeiana) tadpoles // Environ. Toxicol. and
Chem. 2007. 26. N 10. Р.2198
-
2201. РЖ
Биология
08.05-04И5.40.N.,
Schneeweiss U. Mortality of amphibians as a consequence of mineral fertilizing
// Herpetol. 97: Abstr. 3rd World Congr. Herpetol., Prague, 2-10
Aug., 1997. Prague, 1997. P.185. РЖ Биология 98.12-04И5.88.
Schneeweiss
N., Schneeweiss U. Mortality of amphibians as a consequence of mineral
fertilizing // Bull. Chicago Herpetol. Soc. 1999. 34. №10. Р.228
-
231. РЖ
Биология
00.05-04И5.85.
Schuytema
G.S., Nebecer A. V. Effects of ammonium nitrate, sodium nitrate and urea on
red-legged frogs, Pacific treefrogs, and African clawed frogs // Bull. Environ.
Contam. and Toxicol. 1999. 63, N3. P.357-364.R., Singh R.,
Verma R.V. Haematotoxicity of fertiliser Di-ammonium phosphate to fish Channa
punctatus // Himalayan J. Environ. and Zool. 1995. 9,
№1. Р.
17-19.
РЖ
Биология
98.01-04И4.137.P.J.,
Oldham R.S. The effect of ammonium nitrate on the feeding and development of
larvae of the smooth newt, Triturus vulgaris (L.), and the behaviour of its
food source, Daphnia //Freshwater Biol. 1995. 33, N 2. Р.319
-
324. РЖ
Биология
96.03-04И5.132.P.
De, Watt P.J., Oldham R.S. Amphibian decline and aquatic pollution: Effects of
nitrogenous fertiliser // Herpetol. 97: Abstr. 3rd
World Congr. Herpetol., Prague, 2-10 Aug., 1997.
Prague, 1997. P.53. РЖ Биология 99.02- 04И5.122.