Биофильтрация как способ питания беспозвоночных
Министерство
образования и науки Российской Федерации
Бурятский
Государственный Университет
Педагогический
Институт
Курсовая
работа
По
дисциплине: зоология беспозвоночных
На
тему: Биофильтрация как способ питания беспозвоночных
Выполнила:
студентка II курса
гр.
07300з Батуева С.З.
Научный
руководитель:
Абашеев
Р.Ю.
Улан-Удэ 2012 год
Содержание
Введение
Понятие о биофильтрации
Способы питания беспозвоночных
Биоремедиация атмосферы
Биофильтрация гидросферы
Биофильтрация почв
Заключение
Список литературы
Приложения
Введение
В нашем индустриальном обществе проблема
загрязнения окружающей среды и переработки промышленных отходов заставляет нас
искать и пересматривать методы очистки и переработки в поисках наиболее
дешевого, экологичного и имеющего возможность широкого применения. Помимо чисто
механических и химических этапов очистки наиболее естественным, легко и дешево
воспроизводимым является метод биофильтрации.
Еще до приспособления бактерий в качестве биофильтров и
биоочистителей, до появления искусственных загрязнителей, микроорганизмы уже
эффективно выполняли очистительную роль в природе. Недавно российские ученые
исследовали образцы мха из разных болот тундры северной части России и
обнаружили метанотрофные бактерии, которые прекрасно живут в кислой среде и при
низких температурах прямо в клетках сфагнума. Полученные данные позволили
ученым утверждать, что на всей территории севера России от Чукотки и Камчатки
до Полярного Урала работает метаноокисляющий бактериальный фильтр. Этот фильтр
тесно связан с растениями сфагнума и представляет собой физически
организованную структуру, способную контролировать поток метана из торфяных
болот в атмосферу[ 4 ].
Сам термин биофильтрация еще формируется, а
пока рассматривается только как метод очистки, который использует способность
живых организмов усваивать вредные вещества или перерабатывать их в безвредные[
5 ].
Беспозвоночные составляющие большую часть
царства животных осуществляют в своей массе процесс биофильтрации в природе.
Есть такое понятие как животные - фильтраторы, к таковым, например можно
отнести из нашего региона эндемика озера Байкал - эпишура байкальская.
Нужно отметить, что процесс биофильтрации
происходит в результате питания живых организмов, в частности беспозвоночных.
То есть в процессе жизнедеятельности осуществляется круговорот вещества в
природе. Но при вмешательстве антропогенной деятельности баланс нарушается и
происходит загрязнение, которое требует осваивание биофильтрации как
технологического процесса.
Всё сказанное определяет актуальность темы курсовой работы.
Объект исследований - питание беспозвоночных.
Предмет исследований - осуществляемая биофильтрация.
Цель и задачи исследований - анализ явления процесса
биофильтрации как способа питания беспозвоночных. Для достижения цели решались
следующие основные задачи:
• определение термина биофильтрации и его особенностей;
• определение беспозвоночных и их фильтрующих способностей;
• рассмотреть примеры биофильтрации беспозвоночными в
почвенной, водной, воздушной среде обитания и в промышленных биофильтрах;
• разработать методические принципы поиска и выявить виды
беспозвоночных, обладающие высоким фильтрующим эффектом и наилучшей адаптацией
к условиям биологической очистки;
• обосновать целесообразность применения биофильтрации
совместно с существующими методами очистки промышленных отходов для повышения
их эффективности и экологической чистоты.
Для решения поставленных задач использованы методы: анализ
литературы, сравнительный, наблюдение.
Обоснована целесообразность применения
биофильтрации совместно с существующими методами очистки отходов, что позволяет
повысить их эффективность и обеспечить экологическую чистоту. Экономические
показатели при разработке биологической ступени очистки должны учитывать, что в
принципе любые загрязнители - всего лишь выбрасываемые ресурсы, среди которых
немало ценных и дефицитных.
Понятие о биофильтрации
биофильтрация отход беспозвоночный микроорганизм
Термин «биофильтрация» в настоящее время
часто встречается в повседневной жизни обычного человека и ассоциируется со
словами «очищение», «переработка», «экология». Сам процесс существовал в
природе всегда, но был выделен и тщательно проанализирован в конце XX - начале
XXI вв., когда человек впервые столкнулся с проблемой переработки отходов собственной
хозяйственной деятельности. Биофильтрация (от др.
греч. βίος - жизнь, от лат. filtrum
- войлок) это процесс очищения воды, почвы и
воздуха, в основе которого лежит способность живых организмов ( бактерий, растений
и пр.) усваивать вредные вещества или перерабатывать их в безвредные[ 5 ].
Чаще всего говоря «биофильтрация» подразумевают метод.
С технической точки зрения биофильтрация это технология или
одна из ступеней очистки. Биологическая очистка предполагает деградацию
органической составляющей микроорганизмами (бактериями и простейшими).
Например, основными вариантами очистки сточных вод являются активный ил (аэротенки),
биофильтры и метантенки (анаэробное брожение)[9].
С биологической точки зрения, микроорганизмы питаясь и
размножаясь осуществляют биофильтрацию. Например, биологическая очистка сточных
вод представляет собой результат функционирования системы активный ил - сточная
вода, характеризуемой наличием сложной многоуровневой структуры. Биологическое
окисление составляющее основу этого процесса, является следствием протекания
большого комплекса взаимосвязанных процессов различной сложности: от элементных
актов обмена электронов до сложных взаимодействий биоценоза с внешней
средой[14].
Результаты исследований показывают, что характерной особенностью
сложных многовидовых популяций, к которым относятся и активный ил, является
установление в системе динамического равновесия, которое достигается сложением
множества относительно небольших отклонений активности и численности отдельных
видов в ту или иную сторону от их среднего уровня.
Микроорганизмы вообще обладают высокой чувствительностью к
антропогенному воздействию, и в городских условиях их состав сильно меняется.
Поэтому они являются хорошим индикатором загрязненности окружающей среды[24]. Так, по виду
микрофлоры, преимущественно обитающей (или, наоборот, отсутствующей) на данной
территории, можно определить не только степень загрязнения, но и его вид (какое
именно загрязняющее вещество превалирует на данном участке). Например,
индикаторами сильного антропогенного загрязнения является отсутствие коккоидных
форм микроводорослей из отдела Chlorophyta. Наиболее устойчивыми к загрязнению
оказались нитчатые формы синезеленых водорослей (цианобактерий Cyanophyta) и
зеленых водорослей[22] .
Вместе с тем, микроорганизмы сами являются очистителями
окружающей среды. Дело в том, что питательными веществами для многих бактерий
являются абсолютно несъедобные для высших организмов вещества. В большинстве
случаев данные вещества (такие, как нефть, метан и т.п.) являются для таких
бактерий прямыми источниками энергии, без которой они не выживут. В некоторых
других случаях такие вещества не являются для бактерий жизненно важными, но
бактерии могут их поглощать в больших количествах без вреда для себя[25].
Создавая оптимальные условия для роста микроорганизмов в
надлежащим образом спроектированных инженерных системах, скорости процессов
обработки отходов могут быть значительно увеличены, облегчая решение многих
проблем природоохранной биотехнологии. Кроме того, эта дисциплина постепенно
трансформируется от ее обычной функции к новой фазе, характеризующейся
максимальной рекуперацией ресурсов, находящихся в отходах. Каждая территория
обладает определенной техноемкостью - то есть тем количеством антропогенной
нагрузки, которую она в состоянии вынести без необратимого нарушения своих
функций. Внесение на загрязненные участки соответствующих микроорганизмов
значительно повышает этот показатель.
Решение экологических проблем зиждется, в основном, на
фундаменте биокаталитических методов из-за их относительной дешевизны и высокой
производительности, а вся подчиненная область называется природоохранной
биотехнологией, являющейся в настоящее время крупнейшей областью промышленного
применения биокатализа, принимая во внимание объемы перерабатываемых веществ.
Философия в рамках современной природоохранной биотехнологии должна быть
целостной по отношению ко всем компартментам окружающей среды, а это требует
интеграции многих научных дисциплин, и, в первую очередь, детальных знаний о механизмах
протекающих биокаталитических процессов, а также их эффективного инженерного
оформления.
К настоящему времени существует ряд биокаталитических и
инженерных подходов, для защиты трех основных компартментов окружающей среды -
почвы, воды и атмосферы. Основное загрязнение грунтов и водных поверхностей в
мире - это нефтяное загрязнение. Ряд микроорганизмов способны эффективно
утилизировать нефть и нефтепродукты, очищая любые поверхности от опасных
нефтяных пятен[25].
Существует еще одна уникальная и достаточно широко
распространенная группа бактерий - метанотрофы, использующие метан в качестве
единственного источника углерода и энергии. Интерес к термофильным метанотрофам
обусловлен перспективами их практического применения как в науке, так и в сфере
экологии. В биотопах в основном встречаются метанотрофные бактерии родов
Methylocystis и Methylobacter[26].
Разумеется, помимо метанотрофных и нефтеперерабатывающих
бактерий существуют и другие виды, перерабатывающие ряд других загрязняющих
веществ. Вот некоторые процессы переработки органических веществ, которые
катализируются микроорганизмами: прямое окисление пропилена в 1,2-эпоксипропан
молекулярным кислородом, прямое окисление метана в метанол, микробиальное
эпоксидирование олефинов[27], окисление газообразных углеводородов в спирты и
метилкетоны кислородом воздуха (с участием газо-ассимилирующих
микроорганизмов), эпоксидирование пропилена иммобилизованными клетками
газо-ассимилирующих микроорганизмов. При этом, если производственные процессы
переработки химических загрязнителей обычно требуют высоких температур,
биокаталитические процессы проходят в микроорганизмах при температуре, как
правило, в пределах 20-40 градусов Цельсия. И, если при химических процессах
образуется масса побочных продуктов, токсичных сами по себе (например, при
окислении пропилена в 1,2-эпоксипропан молекулярным кислородом образуются
альдегиды, угарный газ, ароматические органические вещества), то при «работе»
микроорганизмов таких веществ не образуется - они разлагаются до воды и углекислого
газа, которые выделяются аэробными бактериями[18].
В настоящее время выведены микроорганизмы, которые могут
утилизировать, то есть перерабатывать с получением для себя энергии, огромное
количество искусственных веществ - таких как, например, различные виды
пластмасс, резины и т.п.
Оценка состояния обитающих в почве организмов, их
биоразнообразия имеет важное значение при решении задач природоохранной
практики: выделении зон экологического неблагополучия, расчете ущерба,
нанесенного деятельностью человека, определении устойчивости экосистемы и
воздействии тех или иных антропогенных факторов. Микроорганизмы и их метаболиты
позволяют проводить раннюю диагностику любых изменений окружающей среды, что
важно при прогнозировании изменений окружающей среды под воздействием природных
и антропогенных факторов.
В частности, среди основных природоохранных и компенсационных
мероприятий в последнее время все чаще называют выделение местных (характерных
для данной экологической зоны) штаммов микроорганизмов, наиболее активно
утилизирующих углеводородное сырье, как основы для проведения этих
мероприятий[25].
Проведение обследований по выявлению деградированных и
загрязненных земель в целях их консервации и реабилитации, а также подбор,
разработка и проведение оптимальных комплексов природоохранных и
компенсационных мероприятий по снижению негативного антропогенного воздействия
на окружающую среду, адаптированных к локальным природным условиям и видам
воздействия[20]. Заключительным шагом является оценка состояния экосистем и
остаточных последствий антропогенного воздействия на окружающую среду после
проведения природоохранных и рекультивационных мероприятий.
В современном мире микроорганизмы активно используются для
биоремедиации. Они «работают» сами по себе или в составе различных
биопрепаратов. Разрабатываются новые и совершенствуются уже существующие
технологии очистки на основе микроорганизмов. В качестве примера можно привести
одну из недавних разработок - биокаталитическую технологию удаления
сероводорода и рекуперации элементарной серы из загрязненных газов, практически
не требующую использования реагентов[19].
Бактерии играют роль экологов в самых разных сферах
производства. С их помощью возможно проводить очистку не только трех
небиологических (гидро-, лито-, атмосферы) и так называемую «живую» (биосфера)
оболочек Земли, но и ликвидировать последствия аварий в исключительно
антропогенных зонах - например, на предприятиях. Многие микроорганизмы успешно
справляются с коррозией[10], многие могут бороться со своими «собратьями» -
бактериями патогенных видов, делая окружающую человека среду пригодной для
жизни и работы.
Способы питания беспозвоночных
Беспозвоночные (лат. Invertebrata) - термин, предложенный
Ж.Б. Ламарком как обобщающее название для насекомых и червей (следует
учитывать, что в те времена объем этих групп понимался иначе, чем сейчас).
Беспозвоночные противопоставлялись позвоночным (лат. Vertebrata) - группе,
объединявшей рыб, птиц, и млекопитающих[5].
В современной систематике эта группа не признается за
полноправный таксон, поскольку она явно сформирована «по остаточному принципу»
- в неё попали все животные, не имеющие позвоночника. С точки зрения
современной систематики - это классический пример парафилетической группы (в
отличие от строго монофилетических позвоночных). По современным представлением,
животные, которых Ламарк отнёс к беспозвоночным, распределены по более, чем
двадцати равноправным группам высокого ранга - типам (наиболее крупные из
которых -членистоногие, круглые черви и моллюски). При этом позвоночные
составляют лишь один из подтипов в типе хордовых, два других подтипа которого
(оболочники и головохордовые) традиционно относят к беспозвоночным[15]. К
беспозвоночным относится порядка 97 % описанных видов животных.
Питание животных представляет собой до некоторой степени
неразработанную область исследования. Верно, что накоплен огромный материал,
например по механике фильтрационного пищеварительной системы и физиологии
пищеварения, но ответ на вопрос, почему животные питаются той или иной пищей,
предполагается очевидным - поедается то, что можно добыть и усвоить[11]. Но
почему большинство примитивных типов животных почти исключительно плотоядные?
Почему так много растительных тканей не потребляется? Почему некоторые виды
более «всеядны» чем другие? И почему некоторые болотные птицы предпочитают
мидий червям? Для ответа на эти вопросы необходимо рассмотреть филогенетические
ограничения, связанные с питанием животных (эволюционное прошлое), различные
механизмы питания(наследие прошлого), а также факторы, определяющие выбор той
или иной пищи в рамках широкого кормового спектра, и возможности влияния жертвы
на выбор хищника (экологическое настоящее[15]).
Все типы животных (может быть за исключением Uniramia) возникли в море, очень
стабильной и однородной среде. Если не считать пограничные приливно-отливные и
непосредственно прибрежные зоны, такие физические факторы, как температура,
соленость, ионный состав, насыщение кислородом, и т.д., не представляют никакой
опасности для выживания морских организмов и не создают физиологических
ограничений для их биологической активности. Поэтому жизнь обитателей моря
лимитируется всего двумя , может быть, тремя факторами, из которых большинство,
если не все связаны с питанием. Животные нуждаются в достаточном количестве
пищи, должны избегать опасности стать пищей для других и иметь определенное
пространство для охоты за пищей. Успешное выживание особей определяется только
этим. Конечно, если их генам предписано существовать в течение более
длительного периода, чем жизнь одной особи, необходимо также произвести
максимально возможное число потомков, способных в свою очередь выживать и
производить себе подобных [10]. Все анатомические, физиологические,
биохимические или онтогенетические особенности лишь механистически способствуют
повышению шансов на решение этих главных жизненных задач.
Что же могло служить пищей для первых животных, населявших
поверхность до-кембрийского морского дна? Только колониальные или одноклеточные
бактерии и протисты, которые во всех водоемах, кроме самых мелководных, также
были в основном гетеротрофами. Достаточно света для фотосинтеза проникает в
толщу воды примерно на 100 м в открытом море, а на мелководье обычно на меньшую
глубину (20-30 м в береговой зоне, а иногда лишь несколько сантиметров в очень
мутных водах с илистыми взвесями). Большая часть континентального шельфа и все
дно океана вообще не знают солнечного света. Первичная продукция органического
вещества фотосинтетическими морскими протистами образуется только в поверхностных
слоях воды, сильно удаленных от основных бентосных местообитаний. В наше время
большинство донных животных почти полностью зависят в конечном счете от
оседания сверху мертвых и уже частично разложившихся тканей и от организмов,
обусловливающих их разложение как во время прохождения через водную толщу, так
и после их попадания на морское дно.
Только в прибрежных мелководных зонах фотосинтезирующие
организмы были непосредственно доступны животным, либо отфильтровывающим их из
взвешенного в воде материала, либо заглатывающим прикрепленные формы. Если не
считать филогенетически изолированных губок, питание путем фильтрации было в
эволюции животных гораздо более поздней специализацией, чем обгладывание или
ощипывание. Вместе с тем хемосинтетическая деятельность бактерий морского дна
была, а в некоторых местах все еще остается важным источником первичной
продукции. Бактериальный фотосинтез и хемосинтез характерны для анаэробных или
микроаэробных местообитаний, а также восстановленных субстратов.
Поэтому неудивительно, что наиболее просто устроенные из
доживших до наших дней плоских червей и. по-видимому, их предковые формы были и
остаются потребителями в основном бактерий и протистов, связанных тем или иным
образом с донными осадками и камнями. Пожалуй, более удивительно то, что явно и
скрыто этот примитивный рацион доминировал у всех форм, которым дали начало
плоские черви, даже у перешедших к наземному существованию[27]. Численность
бактерий и протистов в морских осадках огромна, но они очень малы и часто
относительно рассеяны в пространстве, редко встречаясь плотными скоплениями,
хотя бывают и исключения, которые мы рассмотрим позже. Эти особенности имели
два очень важных последствия для образа жизни животных.
Во-первых, они должны обладать определенной подвижностью,
чтобы отыскивать новые источники пищи, когда прежние оказываются исчерпанными.
Таким образом, подвижность стала одним из условий животного образа жизни.
Во-вторых, потребитель мелких, рассеянных в пространстве организмов и сам
должен быть относительно мелким. Чем крупнее животное, тем выше его
метаболические потребности и тем больше ему требуется пищи в единицу времени.
Это можно видеть на графике зависимости между размером тела и расходом энергии,
необходимой для его существования (прил. 1). Крупное животное не сможет
прокормиться бактериями или мелкими протистами, поскольку не сумеет потребить
достаточное их количество в единицу времени.
Однако само увеличение размеров тела, по-видимому, дает
селективные преимущества по трем причинам:
) более крупные животные обычно способны произвести больше
потомства (т.е. повышается репродуктивный потенциал);
) больший размер может повысить шансы не быть съеденным
другими организмами (т. е. повышается выживаемость);
Кроме того, при одинаковой усваиваемости энергетически всегда
выгоднее поглощать крупные частицы данной пищи, чем мелкие, а для этого тоже
нужно обладать большим размером.
Итак, с учетом селективных преимуществ увеличения размеров
неудивительно, что большинство доживших до наших дней плоских червей крупнее
видов, которые питаются бактериями, и пищей им служат более крупные объекты, т.
е. животные. В наше время две наиболее значительные исходные группы животных -
билатеральные плоские черви и радиальные кишечнополостные - представлены
практически лишь плотоядными формами. Тем не менее размер тела плоских червей
ограничен скоростью диффузии поэтому преимущества, связанные с дальнейшим
увеличением тела, делают выгодными любые морфологические изменения, которые бы
сняли такое ограничение. Это обстоятельство и стремление избежать хищничества
со стороны плоских червей, возможно, и послужили двумя главными толчками,
приведшими к широкому разнообразию червеобразных форм, появившихся на ранних
этапах эволюции животных.
Однако в самых мелких морских водах в изобилии присутствуют
протисты в виде нитчатых или слоевищных, колониальных или многоклеточных
водорослей[8]. Хотя более крупные морские водоросли создают ряд особых
затруднений , нитчатые формы и ювенильные стадии макроскопических типов можно
относительно легко отделить от субстрата, к которому они прикреплены, с помощью
специального соскабливающего органа, что делает этот источник обильной и
концентрированной фотосинтезирующей ткани доступным для потребления. Моллюски,
одни из ранних эволюционных производных плоских червей, использовали именно
этот специализированный способ питания, обезопасившись как от быстрых течений,
характерных для мелких вод, так и от хищников дорсальной раковиной.
Соскабливание с помощью радулы является также эффективным средством добывания в
качестве пищи сидячих или прикрепленных животных, защищенных наружными
раковинами, оболочками или домиками, а также наземных растений, и в течение
своей последующей эволюции моллюски распространили свой исходный способ питания
на большинство доступных типов пищи.
Другие рано дивергировавшие от плоских червей животные либо
сохранили предковый кормовой рацион, либо развили механизмы для концентрации
взвесей мелких частиц, проплывающих мимо или осаждающихся из верхних слоев
воды. Во многих морских средах скорость осаждения и количество органического
материала, взвешенного в воде, практически постоянны. Значит, если консумент
расположится так, что обеспечит себе его перехват и концентрирование, он уже не
будет нуждаться в большой подвижности или даже в подвижности вообще, если не
считать одной системы органов, и, став сидячим или прикрепленным, сможет сократить
энергетические потребности и. следовательно, необходимое количество пищи на
единицу веса тела. В результате относительно крупные животные способны
существовать на рационе, состоящем в основном из бактерий, протистов и мертвого
органического вещества.
Некоторые из этих ранних производных плоских червей извлекали
пищевые частицы из водной взвеси, пропуская ее через некое фильтрационное
приспособление (потребители суспензий); другие собирали эти частицы и связанные
с ними скопления микроорганизмов уже после их оседания на поверхность субстрата
(потребители осадков); третьи перехватывали частицы во время их пассивного
оседания еще до того, как они достигали дна (потребители оседающих частиц)[3].
К потребителям суспензий относятся губки, лофофоровые, беспозвоночные
хордовые, ряд представителей моллюсков, кольчатых червей и членистоногих, в том
числе некоторые формы, которые позже в ходе филогенеза перенесли с собой этот
способ питания в толщу воды вплоть до зоны фотосинтеза[2]. Пищеварительные
адаптации потребителей суспензий описаны в этой главе ниже (разд. 9.2.5), а
здесь мы остановимся лишь на некоторых общих чертах их питания. Всем им
свойственны фильтр определенного типа и либо особое положение участков захвата
пиши, позволяющее перехватывать естественный (и постоянный) водный поток, либо
приспособления, создающие собственный ток воды через фильтр.
У губок одни и те же клетки- хоаноциты-создают ток воды за
счет биения своих жгутиков и улавливают из нее пищевые частицы, задерживающиеся
на воротничке из микроворсинок вокруг жгутика[1]. Мелкий размер отверстий
образуемого микроворсинками фильтра позволяет задерживать частицы, размер
которых меньше, чем у бактерий. Вполне очевидно, что всем потребителям
суспензий выгодно фильтровать один и тот же объем воды не более одного раза, и
у губок это обеспечивается ее пропусканием через множество мелких пор,
распределенных по всей поверхности тела, и выведением через одно или несколько
крупных отверстий[3]. Образующийся более сильный поток относит профильтрованную
воду от тела губки (у колониальных и планктонных огнетелок из оболочников с
аналогичной системой общий для всех зооидов выводимый поток создает, кроме
того, реактивную силу, способствующую движению колонии сквозь толщу воды[2].
В отличие от этого у разнообразных потребляющих суспензии
потомков плоских червей для фильтрационного питания адаптировались или
образовались особые органы-щупальца на голове или вблизи от нее у аннелид,
мезосомальный лофофор у форонид и их производных, сильно увеличенные ктенидиальные
жабры у двустворчатых моллюсков и т.д.[1]. Однако принцип действия здесь один и
тот же. Биение ресничек в определенных участках фильтрующего органа создает
поток воды, в то время
как другие ресничные поверхности осуществляют захват и
транспорт пищевых частиц ко рту с помощью слизи. Хотя захватываются лишь
частицы определенного размера, имеются дополнительные ресничные зоны либо на
самом фильтре, либо на каком-нибудь вспомогательном органе, в которых частицы
сортируются на потребляемые и отсеиваемые, часто в виде псевдофекальной массы.
Эквивалентная система есть и у беспозвоночных хордовых животных [4].
Однако у членистоногих фильтраторов реснички отсутствуют в
связи с наличием экзоскелета, и вместо них функцию фильтра выполняют щетинки на
различных конечностях[1]. Поток воды, несущий пищевые частицы, создается как бы
плавательными движениями фильтрующих или других придатков. Фильтры из щетинок
обычно не такие мелкие, как ресничные, поэтому захватывают только относительно
крупные частицы.
Все животные, использующие щупальцеподобные структуры для
питания суспензиями (как и многие другие фильтраторы), являются прикрепленными
или сидячими, а их ловчие щупальца расположены в принципе
радиально-симметрично, обычно вокруг рта. Такая же функциональная система наблюдается
у первично радиальных книдарий [2]. Тело обычно защищено раковиной или трубкой
или же помещается в норке, сделанной в субстрате. Это позволяет животному и/или
его нежному фильтрующему аппарату втягиваться под оболочку, обеспечивающую
безопасность от хищников. При таком полускрытом образе жизни терминальный анус
неизбежно привел бы к прохождению фекальных масс вдоль всей поверхности тела
перед выносом их во внешнюю среду. Поэтому у видов,обитающих в трубках или
эквивалентных приспособлениях, наблюдается заметная тенденция к образованию
U-образного кишечника с расположением ануса, а часто и выводных отверстий
экскреторных органов вблизи переднего конца тела на пути выходящего потока воды
[3].По тем же причинам сходной анатомической структурой обладают и многие
сидячие потребители осадков [5].В других случаях у норки бывает по крайней мере
два отверстия, и ее обитатель создает через нее однонаправленный поток воды или
же, что гораздо реже, животное внутри трубки согнуто пополам так, что рот и
анус оказываются у одного ее выхода.
У некоторых аннелид, включая Nereis di-versicolor и
Chaetopterus. аппендикуларий из> оболочников, некоторых брюхоногих
моллюсков, эхиурид и т. д. фильтрующий аппарат является не частью тела, а
секретиру&-мой структурой в виде сети или мешка из слизистых нитей, через
которые пропускается поток воды, создаваемый движением специальных образований,
служащих у других животных локомоторными органами. Через определенные периоды
времени эта сеть вместе с собранной на ней пищей съедается, и животное
секретирует новый фильтр [27]. У некоторых кораллов, других брюхоногих
моллюсков и личинок насекомых независимо образовались системы захвата пищи,
сходные с этой, но использующие в основном естественные, а не специально
создаваемые потоки воды.
Сбор пищи с поверхности субстрата потребителями осадков не
требует таких сложных приспособлений, как при питании суспензиями, и,
действительно, некоторые черви просто заглатывают поверхностный слой субстрата
с помощью совершенно неспециализированной ротовой области. Тем не менее в ряде
случаев, особенно у сипункулид, голотурий и эхиурид, а также у некоторых
аннелид и полухордовых около ротового отверстия развиты особые лопасти,
щупальца или растягивающийся хоботок, покрытые ресничками и способные двигаться
по субстрату или в нем, что позволяет более эффективно собирать пищевые частицы
вблизи норок, где живут такие формы .[2] У потребителей осадков с лопастями или
щупальцами вокруг рта есть черты, общие с потребителями суспензий (как
отмечалось выше в связи с U-образным кишечником), и некоторые из них могут
использовать оба этих способа питания. Однако две проблемы касаются только
потребителей осадков: а) органический материал часто составляет лишь малую
часть последних; б) основная часть самого органического материала часто состоит
из твердых и неперевариваемых остатков, накапливающихся именно из-за
использования их не животными, а только бактериями[1].
В связи с этим, за исключением мелководий, где на поверхности
субстрата могут обитать бентосные фотосинтезирующие протесты, потребители
осадков, вероятно, зависят от бактерий, способных превращать недоступную иначе
органику в съедобный материал, и от тех протистов и интерстициальных животных,
которые сами зависят от этих бактерий. Так или иначе, они вынуждены либо
поглощать большие количества отложений (поэтому переоткладывают бентосные
осадки и являются мощными агентами биотурбации), либо проводить большую работу
по их сортировке и отбору пищи. Щупальца многих потребляющих осадки червей
способны, например, вытягиваться на большие расстояния от норки, где обитает
животное. За исключением особо "плодородных" участков, потребители
осадков не могут достигать плотности населения, характерной для потребителей
суспензий, у которых лимитирующим фактором часто является пространство, а не
пищевые ресурсы[10]. Поскольку во многих случаях в конечном счете приходится
полагаться на продуктивность бактерий, а не на скорость оседания детритных
частиц, а продуктивность бактерий в свою очередь может ограничиваться не только
поступлением углерода (например, дефицитом питательных веществ в
интерстициальных водах), скорость роста потребителей осадков часто ниже, чем у
потребителей суспензий.
Последняя группа животных, питающихся веществом, поступающим
из поверхностных вод,- потребители оседающих частиц, например некоторые
стебельчатые иглокожие с телом, прикрепленным к морскому дну, но значительно
поднимающимся над ним, чтобы перехватить частицы детрита до того, как они
"разбавятся" донными отложениями [3]."Перехват" производят
несколько радиально расположенных рук, несущих гидравлически управляемые и
покрытые слизью сосочки; затем по ресничным бороздкам пища направляется к
ротовому отверстию, расположенному в центре образованного основаниями рук
круга.
Судя по выжившим представителям древних групп, на очень
ранних этапах истории многоклеточных животных уже, вероятно, существовал еще
один совершенно особый способ питания. У некоторых морских обитателей
мелководья в поверхностных тканях тела находятся симбиотические
оксифотобактерии (прохлорофиты и циано-бактерии) или фотосинтезирующие протисты
(одноклеточные динофлагеллаты или хлорофиты). Питание некоторых плоских червей
и книдарий, по-видимому, полностью зависит от этих симбионтов, хотя в
большинстве случаев они не способны их переваривать. Такие бескишечные
турбелларий, мягкие кораллы и медузы постоянно (сидячие формы) или временно
(подвижные формы) находятся на полном солнечном свету. Их симбионты
фотосинтезируют обычным способом, но как источник минерального питания частично
используют неорганические азот и фосфор образующиеся из органических соединений
в ходе метаболизма хозяина, а в качестве источника углерода - углекислый газ,
выделяющийся в процессе его дыхания. По-видимому животное-хозяин каким-то
образом делает клеточные стенки симбионта более тонкими и/или более
проницаемыми, чем в норме, и поэтому часть веществ, образующихся за счет
фотосинтеза (сахара, жиры и аминокислоты), диффундирует в ткани хозяина, где
ассимилируется: продукты их распада затем снова путем диффузии попадают в клетки
симбионта. Таким образом животное "доит" этих автотрофов, плотность
которых может достигать 30 ООО в 1 мм-1 ткани хозяина[10]. Другие виды морских
животных хотя и частично, но также зависят от фотосинтезирующих симбионтов.
Аналогичным образом некоторые глубоководные погонофоры
целиком или отчасти зависят от симбиотических хемоавто-трофных бактерий,
использующих восстановленные соединения серы и метан, выделяющиеся из жерл и
фумарол морского дна. Недавно было обнаружено, что различные олигохеты и двустворчатые
моллюски, связанные с другими, также богатыми сульфидами, морскими
местообитаниями, получают около 50% фиксированного углерода от бактерий,
окисляющих серу[27]. Тот факт, что симбионты участвуют в питании представителей
большинства типов животных, причем особенно широко в группах, считающихся
наиболее близкими к предковым формам, дает основание предполагать, что, хотя
симбиотическое происхождение многоклеточное™ и спорно[15], своему возникновению
и успеху древние группы животных могли быть в значительной степени обязаны
симбиозу и, вероятно, особенно тогда, когда пищевые ресурсы были ограничены (и
в самом деле, хлоропласты зеленых водорослей, по-видимому, произошли от других,
когда-то эндосимбиотических, эукариотных водорослей, а если это так, симбиотическое
объединение отдельных эукариотных организмов могло дать начало некоторым другим
эукариотам).
Способы питания и типы пищи, описанные выше, были, по всей
вероятности, в течение сотен миллионов лет единственными доступными свободно
живущим животным, и в наше время обитатели морей кормятся
в основном живыми и мертвыми бактериями, протистами и друг
другом. В конце концов возникшие в море формы заселили сушу, и, хотя наземные
бактерии, протисты и другие животные могли обеспечить им практически такой же рацион
(с поправкой на неэффективность потребления суспензий в наземных условиях), в
новых местообитаниях они столкнулись с новым, обильным, однако совершенно
особым источником пищи - наземными бриофитами и сосудистыми растениями.
Серьезная проблема, вставшая перед их потенциальными потребителями, заключалась
в том, что большая часть биомассы растительного организма-грубые
неперевариваемые клеточные стенки из целлюлозы и почти инертных химически
структурных полимеров типа лигнина. Предки большинства животных никогда не
нуждались в целлюлазах или сходных ферментах для усвоения пищи, и первые
наземные формы, во-видимому, оказались абсолютно неприспособленными для
использования этого обильного, но химически стойкого источника пищи. Только в
меловом периоде растительноядность широко распространилась среди наземных
животных. Однако небольшое число морских видов уже до некоторой степени решило
очень близкую проблему. Крупные водоросли морских мелководий также содержат
сложные утлеводы, обеспечивающие устойчивость к сильным движениям воды. Эти
макроскопические формы так же потенциально пригодны в пищу, как и большинство
тканей наземных растений, но потреблять этот в целом непривлекательный корм,
пока он живой, способны только две группы морских животных.
Первая из них- это некоторые морские ежи. Большинство
потребителей морских водорослей нуждаются в обязательном предварительном
расщеплении их прочных полисахаридов бактериями до усвояемой формы. К таким
животным относятся классические потребители осадков. Бактерии, особенно
анаэробы, способны разлагать самые разнообразные органические молекулы, в том
числе из нефти и многих пластмасс, однако, вместо того чтобы использовать
результаты этих процессов, происходящих во внешней среде, некоторые морские ежи
перенесли их внутрь своего тела. Специальный участок их кишечника содержит
культуру анаэробных бактерий[1]. Заглатывая грубые ткани морских водорослей,
морские ежи обеспечивают бактерий пищей; те разлагают эти ткани, и
животные-хозяева переваривают как продукты метаболизма бактерий, так и их
самих. Следующая проблема, связанная с питанием морскими водорослями, - низкое
содержание в них азота на единицу веса, но в анаэробных условиях некоторые
бактерии способны фиксировать атмосферный азот, растворенный в морской воде,
т.е., живя в кишечнике иглокожих, могут повышать количество азота в их
преимущественно углеводной диете. Хотя морские ежи как бы преадаптированы к
наземной растительной пище, ни один из них не перешел к жизни на cуши, но
многие из наиболее успешных наземных фитофагов решили эту проблему сходным
образом[2].
Другая группа потребителей морских водорослей-это брюхоногие
моллюски, и неудивительно, что при их исходном образе жизни на мелководье,
включающем соскабливание пищи с субстрата, они оказались среди немногих групп
животных, обладающих ферментами, необходимыми для разложения большого числа
полимерных углеводов, в том числе и целлюлазами. В отличие от иглокожих
брюхоногие стали процветающей группой наземных фитофагов, что может
засвидетельствовать любой садовод.
Хотя у некоторых сухопутных членистоногих есть целлюлазы,
питание, основанное на деятельности кишечных симбиотических микроорганизмов,
характерно для многих успешных наземных фитофагов, включая тараканов, термитов,
некоторых жуков и млекопитающих. Все они для переваривания растительной пищи
используют сим-биотические бактерии и протистов, обеспечивающих распад
углеводных цепей и/или поступление дополнительных количеств органического
азота. Эти так называемые "травоядные" животные столь же зависимы от
бактерий и простейших, как и бескишечные плоские черви, сипункулиды и
питающиеся осадками аннелиды.
Однако не все части растения одинаково химически устойчивы;
например, молодые
фотосинтезирующие листья для животных доступнее. И все же
лишь немногие из фитофагов могут разрушать стенки растительных клеток, чтобы
добраться до их содержимого, используя свой собственный ферментный арсенал.
Наземные беспозвоночные, не способные разлагать целлюлозу с помощью симбионтов
или собственных ферментов, выработали два особых метода извлечения содержимого
растительных клеток или растительных соков. Гусеницы, кузнечики и другие
насекомые, питающиеся листьями, откусывают их мелкие кусочки или тонкие полоски
и потребляют содержимое клеток, разрушенных при разрывании и последующем пережевывании
тканей растения. Целые клетки при этом в пищу непригодны, поэтому лишь
относительно небольшая доля (в среднем одна треть) откусанных частей
усваивается и для возмещения низкой эффективности питания требуется
переработать большое количество корма[3]. Подобная система может существовать
только благодаря обильной биомассе растений. Второй способ основан на
проникновении в систему транспортировки жидкостей внутри растения с помощью
колюще-сосущего ротового аппарата[5]. Так питаются все равнокрылые, например
тли. Они избегают устойчивых химически структурных углеводов, однако иногда
нуждаются в симбиоти-ческих кишечных бактериях для увеличения с их помощью
количества азота в разбавленной и бедной этим элементом кормовой жидкости.
Растениям, используемым в пищу, очевидно, были выгодны
приспособления, препятствующие потреблению фотосинтезирующих тканей, и подобно
другим прикрепленным организмам (морским колониальным животным, например), мало
приспособленным к другим способам обороны, они развили сложную систему
механической и химической защиты[5]. Как обычно бывает в таких случаях, это
положило начало "гонке" наступательных и оборонительных
вооружений-образованию токсинов у кормовых объектов и механизмов детоксикации
или устойчивости у их потребителей.
Только одна категория фитофагов эффективно использует
вещества растительного происхождения без помощи симбиотических бактерий или
протистов. Это те виды, которые питаются веществами или структурами,
развившимися специально для привлечения животных (нектаром, плодами, семенами и
т.д.), необходимых растениям для опыления и/или распространения. Хотя плоды и
семена в основном предназначены для позвоночных, членистоногие и моллюски также
иногда "приобщаются" к этим ресурсам до того, как они съедены птицами
или млекопитающими.
Грибы с их хитиновыми клеточными стенками и отсутствием
прочных опорных тканей не столь трудны для переваривания животными и поэтому
широко используются ими. Некоторые насекомые в этом плане заходят столь далеко,
что собирают растительные ткани, пережевывают их до мягкой массы и затем
используют ее как субстрат для выращивания грибов, идущих в конце концов в
пищу. В какой-то степени грибы играют ту же роль, что и водоросли в питании
предков наземных животных.
Несмотря на широко распространенное представление о
преобладании на суше пищевых цепей типа растение->растительноядное
животное->плотоядное животное, сложившееся в связи с большим вниманием,
уделяемым в научных исследованиях травоядным (особенно пастбищным
млекопитающим), основу большинства наземных пищевых сетей составляют не живые
растительные ткани и не обгрызание их, а разлагающиеся органические остатки.
Основные потоки энергии идут через животных, использующих в пищу детрит и
лесную подстилку с участием бактерий, протистов и грибов. Таким образом,
наземные животные недалеко ушли от своих морских предков в смысле способности
"перерабатывать" различные типы корма, и даже сейчас фитофаги
потребляют менее 3% лесной продукции в виде живого растительного материала[15].
Типы
питания животных: способы захвата и переработки пищи
Влияние давних представлений о важнейшей экологической роли
цепей питания, включающих обгрызание растений, отражено даже в названиях
классических трофических уровней. Многие годы выделение типов питания животных
основывалось на таких пищевых цепях и на систематической принадлежности
поедаемых организмов; отсюда, например, "растительноядные",
"плотоядные" и (промежуточные) "всеядные" формы. Подобная
классификация порождает много неудобств. Во-первых, исключительно растительноядных
или плотоядных видов относительно мало, особенно если учитывать все стадии их
жизненного цикла, т. е. большинство животных попадают в неопределенную
категорию "всеядных"[2]. Во-вторых, будучи основана на старом
представлении о разделении живых организмов на два царства-растений и животных,
она не решает вопроса о том, куда отнести виды, питающиеся бактериями и
протистами и составляющие довольно обширную группу. В-третьих, филогенетические
связи видов добычи необязательно важны для экологии питания ее потребителей.
Позже стало принято различать категории консументов по
способам питания. Можно говорить о хищниках, паразитах, обгрызающих и
ощипывающих животных, о потребителях суспензий и осадков, о питающихся с
помощью симбионтов. Эта классификация не учитывает систематического положения
организмов, используемых в пищу,- например, "обгрызатели",
потребители суспензий и осадков могут поедать бактерий, протистов, грибы,
растения и животных[15]. Если способы питания животных не делают различий между
царствами поедаемых организмов, не следует делать это и нам при анализе
биологии питания. Поэтому в следующем разделе мы от них абстрагируемся и более
детально рассмотрим особенности поглощения и переваривания пищи.
У большинства беспозвоночных "сквозной" кишечник с
ротовым отверстием, расположенным более или менее спереди и служащим для
захвата пищи, и анальным отверстием, сдвинутым более или менее назад и служащим
для вывода из кишечника во внешнюю среду непереваримых остатков и экскретов. У
некоторых животных кишечник заканчивается слепо, и единственное отверстие
служит как для поглощения пищи, так и для освобождения от "отходов".
Сюда относятся в основном книдарий и плоские черви, но также и ряд других
животных, у большинства из которых такое строение носит вторичный характер. У
других форм (некоторые паразиты, но также и свободноживущие виды)
пищеварительный канал вообще отсутствует, и пища поступает либо от внутренних
симбионтов, либо через наружную поверхность тела. Много и отклонений от этих
вариантов. Например, у форм, ведущих прикрепленный образ жизни, рот часто
расположен в центре верхней стороны тела, а не в его передней части; у
некоторых свободноживущих форм, предки которых были прикрепленными, например у
свободно передвигающихся иглокожих, рот находится в центре нижней поверхности
тела, а у пищеварительного канала хордовых и ряда других животных наружных
отверстий множество[27].
С точки зрения развития пищеварительный канал животных
включает три отдела: эктодермальную переднюю кишку (образуется при втягивании
части наружной поверхности тела), также эктодермальную заднюю кишку и
энтодермальную среднюю кишку, где происходят переваривание и всасывание, часто
снабженную слепыми дивертикулами. Каждый из этих основных отделов разделен на
функционально различные части [10]. Передняя кишка может состоять из а) ротовой
полости, в которую ведет ротовое отверстие и открываются протоки "слюнных
желез" (иногда специализированных на образовании клейких секретов,
антикоагулянтов или токсинов); б) мускулистой глотки, иногда участвующей в
поглощении пищи, либо действуя как насос, либо образуя выворачиваемый орган для
схватывания добычи, либо (как у хордовых) служа фильтром при питании
суспензией; в) короткого пищевода и/или г) накапливающего органа-зоба, особенно
хорошо развитого у животных, поглощающих через длительные интервалы времени
большие количества пищи, постепенно поступающей в среднюю кишку.
Средняя кишка обычно подразделяется на а) мускулистый
желудок, б) различные слепые секреторные и/или всасывающие отростки и в)
кишечник. В желудке происходит механическое измельчение и сортировка
проглоченной пищи; иногда в нем имеется особый отдел для растирания пищевых
частиц. Иногда здесь же начинается переваривание, но чаще переваривание и/или
всасывание протекают в различных дивертикулах- слепых выростах, отходящих от
кишки сразу за желудком. У моллюсков и ракообразных эти выросты превращаются в
крупные сложные органы - гепатопанкреасы.
У примитивных форм переваривание идет в основном
внутриклеточно, когда частицы пищи фагоцитируются клетками выстилки
абсорбирующего отдела пищеварительной системы и расщепляются в их вакуолях.
Такой способ питания преобладает у губок, кишечнополостных, плоских червей и
ряда других животных, у которых пища поступает в среднюю кишку в тонкодисперсном
состоянии. У более сложно организованных животных, особенно у поглощающих пищу
отдельными крупными массами, переваривание происходит в основном с помощью
ферментов, секретируемых в просвет кишки, а его продукты затем абсорбируются
клетками ее стенки. У таких видов дивертикулы имеют, как правило, чисто
секреторную функцию, и всасывание происходит в кишечнике. Внеклеточное
переваривание способствует ферментной специализации как отдельных клеток, так и
различных участков кишки, однако приводит к необходимости производить большее
общее количество ферментов, поскольку просвет кишки- крупная открытая система,
в которой нелегко поддерживать их оптимальную для пищеварительных процессов
концентрацию.
Наконец, задний отдел пищеварительного тракта (если он есть)
представлен прямой кишкой (ректумом), где может происходить всасывание воды (у
некоторых наземных животных[12] и образование фекальных масс перед их
выведением наружу через анальное отверстие.
Все животные нуждаются в получении из пищи энергосодержащих
веществ, используемых немедленно или несколько позже, аминокислот, необходимых
для синтеза структурных и метаболических белков, и ряда других химических
элементов и соединений, например витаминов, участвующих в катализе
биохимических реакций. Другими словами, животные должны поедать углеводы, жиры,
белки и витамины и, если они не могут всасываться сразу, переваривать их до
усвояемых форм. Виды, пища которых бедна белком и/или витаминами, обычно
нуждаются в симбиотических кишечных бактериях, синтезирующих эти вещества, а
потребители сложных полимерных углеводов - в симбиотических бактериях и
протистах, разлагающих эти вещества до более простых органических молекул.
Значительная часть фекалий состоит из кишечных бактерий.
Обгрызатели и соскабливатели
Обгрызатели и соскабливатели -это подвижные потребители
неподвижной добычи, использующие в пищу доступные для них ткани, но обычно не
убивающие при этом свои одиночные или колониальные жертвы. На суше источниками
питания для них являются растения и грибы, а в море- колонии животных (таких,
как книдарий, мшанки или оболочники), бактерий и многоклеточные водоросли.
Добывание пищи требует наличия твердых грызущих или скоблящих ротовых частей
типа радулярной ленты моллюсков[15], аристотелева фонаря морских ежей[5], склеротизованных
челюстей насекомых и т.д., а поиск корма здесь не составляет главной проблемы
(сильное отличие их от большинства типов охотников), поскольку съедобный
материал, как правило, находится в изобилии. Трудности этой категории животных
в следующем: а) наличие систем химической защиты у неподвижных жертв; б)
небольшая доля пригодной для усвоения пищи на единицу веса всего поглощенного
материала из-за большого количества в нем устойчивых структурных или защитных
элементов и часто низкое содержание белка в усвоенной органике.
Проблема высокого содержания неперевариваемых углеводных
полимеров в растениях и водорослях (агар, альгин, ламинарии, целлюлоза и др.)
частично решается, как уже говорилось, благодаря специфическим ферментам и
одновременному удлинению пищеварительного тракта (особенно средней кишки [14]),
увеличивающему размер органов, переваривающих устойчивые материалы, а частично
- за счет симбиотических бактерий и протистов, обитающих в особых частях кишки
(часто в задней кишке, но иногда в зобе или желудке). Нередко присутствует
крупный зоб для накопления запасов[9].
Кишечные симбионты разлагают полисахариды анаэробным путем с
образованием жирных кислот и простых углеводов, пригодных для всасывания. Такой
способ достигает своего наивысшего развития у термитов, использующих в пищу
наиболее устойчивый из природных органических материалов древесину. У этих
насекомых задняя кишка длиннее всего остального пищеварительного тракта и
содержит плотную популяцию жгутиковых Нуpermastigina. Эти протисты фагоцитируют
частицы древесины и сами содержат симбиотические бактерии, которые,
по-видимому, в основном и отвечают за расщепление целлюлозы. Другой компонент
дерева, лигнин, вероятно, не переваривается.
Фактор размера также важен при потреблении растительного материала
с относительно низкой питательной ценностью. У крупных животных, например
позвоночных размером с кролика, низка удельная скорость метаболизма, что
снижает потребность в энергии на единицу веса тела. Благодаря большому объему
они способны накапливать в переваривающем отделе кишки много растительного
материала. Млекопитающие, будучи гомойотермными животными, поддерживают
температуру, необходимую для эффективной работы пищеварительных ферментов.
Поэтому они могут существовать за счет грубых трав и подобных им материалов,
поглощаемых в больших количествах. Мелкие животные, например растительноядные
беспозвоночные, вынуждены питаться кормом более высокого качества и
обеспечивать себе доступ к содержимому растительных клеток, протыкая их,
соскабливая целлюлозные стенки с помощью радулы или прокусывая их своими
ротовыми частями. Поглощение больших количеств пищи здесь, как правило,
невозможно просто из-за мелких размеров тела. Даже мелкие птицы и млекопитающие
не могут существовать за счет малопитательного корма: если это растительноядные
животные, их рацион ограничен богатыми энергией семенами или сходными
образованиями.
У растений много разных средств химической защиты, включая
алкалоиды (никотин, кокаин, хинин, морфин и кофеин), глюкозиноляты, цианогенные
гликозиды и танины[4]. Эти вещества, вероятно, специально образуются в их
тканях, чтобы воспрепятствовать потреблению последних, и иногда сильно
токсичны, являясь естественными инсектицидами - например, у дерриса и
пиретрума. Танины, высвобождаясь, связывают белки, делая их неусвояемыми и
таким образом уменьшая белковую ценность поглощенной пищи. Другие соединения
имитируют собственные гормоны и феромоны фитофага, отрицательно влияя на его
рост, развитие, размножение или вызывая неадекватные поведенческие реакции. У
некоторых растений механизм химической защиты включается только во время
выедания, исключая ненужную трату ресурсов, необходимых для роста и
размножения: недавно показано, что (по крайней мере в одном случае) растения,
даже не будучи объектом нападения, могут реагировать мобилизацией своей
химической защиты на поражение фитофагами соседних экземпляров того же вида.
Другие защитные механизмы растений бывают структурными,
например волосовидные выступы, выделяющие при прикосновении липкие секреты или
отпугивающие вещества, или анатомическое имитационное сходство здоровых листьев
с листьями, пораженными фитофагами[2], а также симбиотическими, особенно
заметными в случае ассоциаций разлчных покрытосеменных и других сосудистых
растений с муравьями. Муравьи изгоняют насекомых, обгрызающих растение, "в
обмен" на его нектар и место для устройства гнезд. Сходные структурные,
химические и другие системы защиты имеются и у морских животных, страдающих от
грызущих консументов, но они пока слабо изучены.
Тем не менее консументы коэволюционировали со своими
неподвижными жертвами в течение многих тысячелетий, и некоторые соскабливатели
и обгрызатели приобрели способность обезвреживать, избегать, связывать или
экскретировать специфические защитные метаболиты, обходить структурные барьеры
и ускользать от внимания охраняющих растения муравьев путем химической мимикрии
под этих насекомых. И все же в большинстве наземных местообитаний,
благоприятных для развития растений, накоплено огромное количество биомассы
живых растений так же, как в океане-водорослей. Это говорит о том, что большая
часть тканей макрофитов на самом деле остается недоступной для грызущих и
скоблящих животных и/или не используется ими и становится источником пищи
только после отмирания и разложения.
Однако даже при наличии целлюлаз и эквивалентных им ферментов
или симбиотических макроорганизмов в кишечнике (а тем более при отсутствии тех
и других) переваривание и ассимиляция тканей макрофитов, как правило, идут
очень неэффективно, а в случае питания жидкостями поступление углеводов намного
превышает потребность в них. Поэтому фекальные массы таких консументов обильны
и содержат много неассимилированного органического вещества[15]. В связи с этим
они являются важным экологическим путем доступа к углероду, фотосинтетически
фиксированному живыми макрофитами, для других категорий животных консументов,
особенно для потребителей осадков.
Питание
взвесями
Основные механизмы, с помощью которых животные, питающиеся
суспензиями, отфильтровывают пищевые частицы от воды, были рассмотрены выше. У
групп, использующих для этого наружные секреты неанатомического характера,
по-видимому, отсутствуют другие специальные приспособления для улавливания
взвесей. Эти животные, по крайней мере по строению своего кишечника, вероятно,
наиболее близки к охотникам-засадчикам, улавливающим свои небольшого размера
жертвы сетями. В самом деле, эти два способа питания не имеют четкой границы:
паук, плетущий тенета, может рассматриваться и как потребитель "воздушной
суспензии", и как охотник-засадчик. Такое же двойственное положение у
морских книдарий; вокруг их центрального ротового отверстия расположены
радиально симметричные кольца щупалец, а в фазе полипов эти животные ведут
прикрепленный образ жизни; оба этих признака характерны для потребителей
суспензий, и, действительно, жертвами здесь являются в основном зоопланктонные
виды, взвешенные в воде. Однако их особи улавливаются не совсем пассивно, а
индивидуально поражаются нематоцистами, пусть даже сама жертва способствует их
выстреливанию, случайно коснувшись щупальца, и книдария не нападает "по
собственной инициативе".
В отличие от этого у групп со слизисто-ресничной системой
фильтрационного питания четко прослеживается специализация пищеварительной
системы. У большинства таких видов с фильтрационным аппаратом, либо свободно
омываемым водой, либо защищенным снаружи раковиной (например, у плеченогих и
двустворчатых моллюсков) [5], передняя кишка представляет собой короткую
трубку, соединяющую наружный фильтр с желудком. Однако у хордовых передняя
кишка сама является местом, где происходит фильтрация, и поэтому сильно
специализирована. Глоточная стенка у этих животных пронизана по бокам
многочисленными мелкими отверстиями ("стигмами"), которые, прободая
стенку тела, открываются на его поверхности. Вода поступает в рот, оттуда в
глотку, затем проходит через стигмы и выводится обратно в окружающую среду,
т.е. ее поток однонаправленный. Хотя сам по себе процесс фильтрации глоткой и
необычен, аналогичная система известна и в других группах, что, возможно,
указывает на ее эволюционное происхождение. Животные, питающиеся мелкими
частицами, часто используют для втягивания их в рот поток воды, которая затем
должна как-то выводиться из организма. У лофофоровых вода просто периодически
"выплевывается", и поглощение частиц в эти моменты прекращается.
Глотка питающихся суспензиями хордовых очень велика и
занимает большую часть объема тела. Пронизанная тысячами отверстий стенка тела
в глоточной области у многих видов почти исчезает и поэтому заменяется вторичной
стенкой, образованной у головохордовых складками ткани, а у туникат -
секретируемой целлюлозной оболочкой. В связи с этим собственно тело целиком или
частично окружено морфологически внешней полостью, или атриумом, между истинной
и ложной стенками тела. Сюда поступает вода, проходящая через стигмы, и отсюда
она выводится в окружающую среду через единственное отверстие, т.е. атриопор
или атриальный сифон[15]. В сущности такая же система наблюдается у
позвоночных. Однако у рыб жаберных щелей (= стигм), ведущих из кишечника к
поверхности тела, значительно меньше, а сама стенка тела толще, поэтому нет
необходимости в дополнительной ложной стенке, а поток воды используется прежде
всего для газообмена, а не для поглощения пищевых частиц.
Специальная железа, эндостиль, расположенная вдоль
вентральной средней линии глотки, вырабатывает слизь, которая тонкими слоями
гонится вверх по перфорированным боковым стенкам. Когда вода вытекает наружу
через стигмы, эти слои перехватывают потенциальные пищевые частицы в некоторых
случаях размером до 0,5 мкм. В конце концов потоки слизи с пищевыми частицами
сходятся дорсально, где образуют продольный тяж в гипербранхиальном желобе.
Движение в этой системе как воды, так и слизи обеспечивается полосками ресничек
внутри глотки (исключение - пелагические сальпы)[2].
У пелагических оболочников поток воды с пищевой взвесью
обычно обеспечивает и движение животного вперед, поскольку ротовое и атриальное
отверстия расположены на противоположных концах тела. В связи с этим реактивным
типом движения число стигм уменьшается до двух (у сальп), слизь образует одну
внутреннюю сеть конической формы, подвешенную в полости глотки, а проталкивание
воды обеспечивается мышечными кольцами, охватывающими тело.
Не только у этих беспозвоночных хордовых, но и у лофофоровых
и моллюсков-фильтраторов пища поступает из фильтрующего органа (независимо от
его типа) в желудок в виде слизистого жгута, несущего съедобные частицы. У всех
животных со слизисто-ресничной системой питания в желудке кислая реакция. Это
снижает вязкость слизистого жгута, обеспечивая выход из него пищевых частиц.
Переваривание и абсорбция питательных веществ происходят в желудке и/или в
дивертикулах средней кишки, отходящих поблизости от него. Непереваренные
остатки попадают в обычно короткий кишечник, где щелочная реакция вновь
увеличивает вязкость слизи и способствует образованию фекальных комочков или
нитей.
Движущая сила, обеспечивающая доставку слизистого жгута в
желудок, также обеспечивается ресничками, у хордовых действующих на этот жгут
непосредственно, а у лофофоровых и моллюсков - косвенно. У двух последних групп
в желудке находится вращающийся стержень, наматывающий на себя слизистый жгут.
У лофофоровых он состоит из слизи и фекальных масс, расположен в пилорической
части желудка и вращается расположенными там ресничками. Время от времени
использованный стержень поступает в кишечник, превращается в фекальный комок,
выбрасывается наружу и заменяется новым.
Питание осадками
Вероятно, большинство современных животных питаются детритом
(органическим материалом, состоящим из мелких частиц) или подстилкой (из частиц
большего размера), но, несмотря на это, четкого представления о природе этого
рациона пока нет. Лист, съедаемый, скажем, земляным червем, - это не просто
мертвый лист, а целая экосистема. На его поверхности и в его тканях обитают
бактерии и грибы, обеспечивающие разложение органики, различные протисты,
потребляющие деструкторов (например, амебы, ресничные, гетеротрофные
жгутиковые), а также лишь немногим более крупные нематоды и клещи, питающиеся
этими мелкими организмами. На поверхности листа иногда оказываются
фотосинтезирующие водоросли и цианобактерии, мертвые и разлагающиеся остатки
других организмов (не в последнюю очередь - из фекалий животных). Все эти живые
и мертвые компоненты могут быть поглощены животным вместе с листом, и задача
состоит в том, чтобы определить, какие из них перевариваются и ассимилируются и
какие в наибольшей степени удовлетворяют метаболические запросы потребителя.
Это не обязательно означает, что потребитель просто
заглатывает основной материал и переваривает все, что может. Теперь известно,
что некоторые потребители осадков способны к значительно более избирательному
поглощению, чем представлялось ранее (впрочем, лишь немногие животные были настолько
детально изучены с этой точки зрения[1]).
Избирательное поглощение, безусловно, должно давать
преимущества, поскольку пищевая ценность различных компонентов детрита весьма
неодинакова. Сравнение органики в осадке, заглатываемом некоторыми морскими
животными, не способными к такому отбору, и их фекалий, а также обработка
известными ферментами из их кишечника проб этого осадка показали, что основная
часть того небольшого количества органического вещества, что содержится в
донных отложениях, остается таким видам недоступной. Они усваивают менее 5-10%
его детритного "пула". (На чисто песчаных пляжах вся органика иногда
составляет менее 1% заглатываемого материала. Слой лесной подстилки ею богаче,
но все равно часто лишь небольшая ее доля пригодна в пищу[15].)
К тому времени, когда любой кусочек органических остатков
становится частью поверхностного слоя подстилки или детрита, он уже теряет
большую часть своей питательной ценности. Если это лист, то перед сбрасыванием
все растворимые вещества переходят из него в остающиеся на родительском
растении ткани, а выщелачивание приводит к потере других органических веществ
уже в первые часы после падения на землю. Органические частицы попадают на
субстрат только пройдя через кишечник консумента, где из них обычно извлекается
большинство питательных соединений, а фекальные массы - один из основных
источников органического вещества в осадках и почве. Поэтому органика в них-это
чаще всего устойчивые структурные, скелетные или защитные соединения,
проходящие без изменений через пищеварительные тракты большинства животных (у
детритофагов, как правило, отсутствуют целлюлазы).
Однако в случае свежих остатков других организмов потребитель
осадка имеет шанс усвоить некоторые содержащиеся в них органические вещества,
хотя и тогда белка скорее всего будет очень мало или он окажется недоступен
из-за присутствия танинов . Вообще наиболее вероятно, что потребители мертвой
органики (за исключением питающихся падалью) существуют за счет живых
микроорганизмов, связанных с этими остатками, служащими лишь удобным
транспортным средством для доставки пиши в кишечник. На суше и в пресных водах
грибы, разлагающие подстилку, возможно, являются основной пищей для
потребителей осадков, а на прибрежном морском мелководье наибольшую питательную
ценность, вероятно, имеют обрастающие частицы грунта фотосинтези-рующие
одноклеточные протесты. Показано, что у животных, способных выборочно питаться
только диатомовыми водорослями, эффективность ассимиляции достигает 70%, в то
время как у неизбирательных потребителей органического материала она менее 4%.
Так или иначе, на основной площади морского дна, где фотосинтез невозможен и
частицы, оседающие из верхних слоев воды, редки, к тому времени, когда они
достигают субстрата, в них остаются только устойчивые к разложению вещества, и
для консументов источником как энергии, так и белка служат бактерии.
Пищеварительная система потребителей осадков, как правило, не
специализирована, но кишечник часто удлинен [4],что обусловлено теми же
причинами, что и в случае обгрызающих и соскабливающих животных. Как и у
последних, обычно образуется очень много фекальных масс, питательная ценность
которых для других потребителей намного ниже, чем у проглоченного материала.
Питание с участием симбионтов
Хотя питание относительно немногих видов животных полностью
зависит от фотосинтеза их эндосимбионтов, некоторые представители таких
филогенетически различных групп, как губки, моллюски, книдарий, хордовые и т.
д., в той или иной степени используют в пищу продукты жизнедеятельности своих
фотосинтезирующих партнеров. Ими могут быть цианобактерии (например, у
некоторых губок и эхиурид), прохлорофиты (у оболочников), а чаще
ди-нофлагеллаты (в виде зооксантелл во многих морских беспозвоночных) или
зеленые водоросли, представленные зоохлореллами (обычно у пресноводных видов).
Некоторые мадрепоровые кораллы удовлетворяют 2/3 своих метаболических
потребностей за счет внутриклеточных зооксантелл, и только 1/3 - за счет
внешних источников питания, включая типичное для кишечнополостных улавливание
зоопланктона. У некоторых актиний зооксантеллы и приспособления для ловли
добычи расположены в различных частях тела: "органы" с симбионтами
расправляются в дневное время, а щупальца с нематоцистами - ночью. Симбиоз
может дополнять растительнояд-ность. Мешкоязычные моллюски высасывают клетки
макроскопических зеленых (и других) водорослей; некоторые их виды могут
сохранять в дивертикулах средней кишки целые функционирующие хлоропласта своей
добычи, которые захватываются пищеварительными клетками-фагоцитами, но
продолжают фотосинтезировать в некоторых случаях еще более двух месяцев. До
половины фиксируемого углерода передается моллюску, чего вполне достаточно для
удовлетворения его энергетических потребностей.
Животные, улавливающие пищу путем процеживания потока воды,
очевидно, тратят энергию не на преследование или поиски добычи, а на фильтрацию
и отбраковку ненужных частиц.
Размеры отверстий в их фильтрах обычно могут меняться лишь в
масштабах эволюционного времени, поэтому трудно предполагать возможность отбора
из воды только частиц с наибольшей питательной ценностью: фильтр
специализирован на размер, а не на съедобность пищи. Прикрепленные фильтраторы
могут максимизировать чистую скорость потребления энергии, изменяя только
следующие параметры:
) скорость фильтрации в зависимости от относительного обилия
в воде частиц разного типа;
) скорость отбраковки в зависимости от относительной
питательной ценности частиц и затрат на отбраковку.
Скорость фильтрации должна увеличиваться при повышении
концентрации частиц с высокой питательной ценностью (рис. 9.35), однако здесь
может быть верхний предел, определяемый возможностями пищеварительного тракта
хищника перерабатывать поступающий материал. При сли-зисто-ресничном механизме
это зависит от движения слизистого тяжа внутри кишки, обусловленного
деятельностью ресничек. У членистоногих, просеивающих пищу с помощью щетинок,
может произойти полное заполнение полости кишки заглоченными частицами. Как
только это происходит, скорость поглощения энергии ограничивается временем
прохождения частиц через пищеварительный тракт и интенсивностью их
переваривания. Следовательно, скорость фильтрации падает до величины,
способствующей поддержанию кишечника в заполненном состоянии. Как и в ситуации
с хищниками, поедающими только наиболее питательные части
добычи, чтобы после этого перейти к другой жертве[5], потребителям суспензий,
зависящим от качества и обилия пищевых частиц, бывает выгоднее быстро
пропускать пищу через кишечник, усваивая лишь часть ее, чем медленно
переваривать большую долю заглоченного.
Затраты на отбраковку бывают высоки, поэтому по мере роста
затрат на отбраковку частиц определенного типа можно ожидать увеличения
скорости их заглатывания. Когда эти затраты очень велики, бывает дешевле
избавляться от даже полностью несъедобных частиц через кишечник, а не с помощью
сортирующих органов. В этих условиях потребитель суспензий может столкнуться с
дефицитом энергии, и ему иногда выгоднее полностью прекратить питание до
изменения состава взвешенного в воде вещества.
При подобных системах сбора пищи возможность стать
"специалистом" очень мала (если вообще существует), разве только в
эволюционном масштабе времени и в отношении определенного размерного диапазона
частиц. Вода просто проталкивается вперед, почти не проходя через ячеи. Такая
избирательность у потребителей суспензий, естественно, не позволяет
регулировать питательную ценность частиц, а проявляется в основном у
свободноживущих личинок, имеющихся у многих видов, когда они расселяются в
места, способные обеспечить прикрепленные взрослые стадии необходимыми
взвешенными пищевыми частицами.
В целом живые существа в рационе потребителей суспензий и
осадков очень малы по сравнению с размером самого потребителя, и, хотя
некоторые из поглощаемых видов и располагают химическими средствами защиты,
большинство, по-видимому, полностью лишены препятствующих выеданию систем.
Вместо этого они наделены способностью быстро размножаться половым и/или
бесполым путем, которой более чем достаточно для компенсации потерь от
хищничества. В этом они отчасти сходны с добычей обгрызателей и
соскабливателей, обычно достаточно интенсивно растущей или бесполо делящейся,
чтобы возмещать утраченные структуры типа листьев, полипов или зооидов, так что
только при исключительно высокой скорости выедания наблюдается гибель отдельных
особей или колоний.
Хотя и ограниченные в разной степени своим эволюционным
прошлым, все животные тем не менее оказываются в масштабах экологического
времени перед выбором: какую из особей-жертв предпочесть в данных
пространственно-временных условиях; продолжать питаться на этом участке или
перейти на другой и т. д. По-видимому, потребители не принимают случайных
"решений", и теория оптимизации дает модель, довольно успешно
предсказывающую поведение животных в ходе фуражирования. В рамках ограничений,
создаваемых факторами, не относящимися к качеству пищи и обеспеченности ею
(например, хищничеством, биологией разможения), некоторые животные,
действительно, питаются таким образом, чтобы максимизировать получаемую прибыль
(выгода минус затраты). С другой стороны, отбор среди видов-жертв ведет к
максимизации относительных расходов, связанных с использованием их
в пищу, за счет особенностей поведения, морфологии или
биохимии. Как и следовало ожидать, различные типы питания подразумевают
неодинаковые потенциальные решения проблемы максимизации прибыли, дающие
селективное преимущество либо генералистам. либо специалистам.
Пищевое поведение, очевидно, ежедневно находится под
давлением не менее интенсивного отбора, чем другие, более изученные аспекты
биологии животных.
Сравнительный обзор питания разных групп животных
Губки. Принципиально неподвижные организмы, осуществляющие
один тип питания - фильтрацию микропланктона. Имеют только внутриклеточное
пищеварение, что требует определенной ловкости от самих клеток, выхватывающих
пищевые частицы из тонких межклеточных каналов, по которым просачивается
гонимая ресничками вода.
Книдарии. Как сидячие полипы, так и плавающие медузы главным
образом ориентированы на питание зоопланктоном. Рачки улавливаются разными
клейкими и стрекательными клетками на щупальцах и отправляются в гастральную
полость. Кроме того, для большинства коралловых полипов установлено
симбиотическое взаимодействие с фотосинтезирующими водорослями, живущими прямо
в тканях полипа.
Турбеллярии. Хищники с замкнутой (мешковидной)
пищеварительной системой. Обладая в общем небольшой подвижностью, охотятся как
поисковики, нападая на еще менее подвижных животных.
Нематоды. При очень однородном внешнем облике «идеального
червяка» довольно разнообразны по типу питания: среди них известны
бактериофаги, альгофаги, детритофаги, хищники и паразиты. В зависимости от
этого они имеют разный ротовой аппарат: без ротовой полости и вооружения
(бактериофаги), с ротовой полостью без вооружения (детритофаги), с небольшими
скребуще-колющими зубами (альгофаги, соскребающие и прокалывающие клетки
водорослей), с крупными зубами (хищники, хватающие протистов и других нематод).
Глотка нематод обычно совершает ритмичные пульсирующие движения, в ходе которых
содержимое ротовой полости поступает в кишечник - выпивается (у бактериофагов,
альгофагов и паразитов) или проглатывается (у детритофагов и хищников).
Коловратки. Очень мелкие и довольно подвижные животные,
распространенные главным образом в планктоне
<#"587942.files/image001.gif">
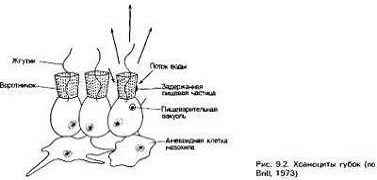
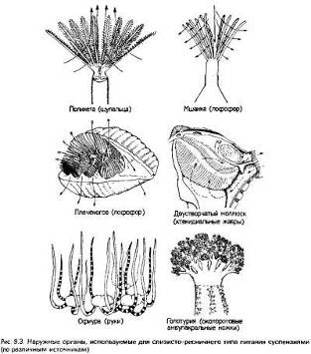
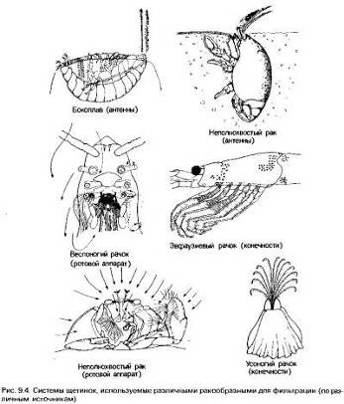

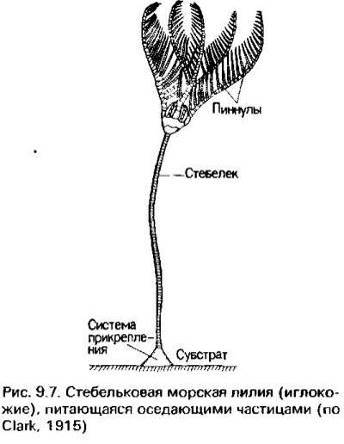



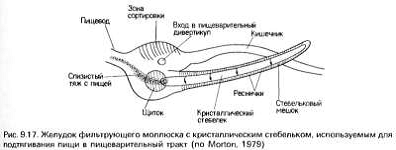
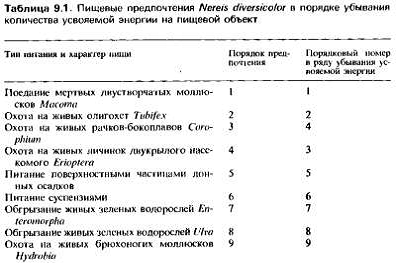
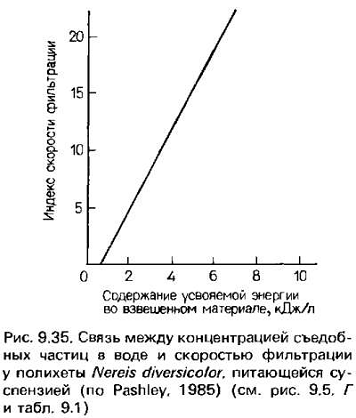
Альтруисты гибнут, но дело их живет
В журнале Science опубликованы результаты примечательной
работы американских микробиологов, изучавших конкуренцию между искусственно
созданными линиями кишечных палочек - "лабораторных крыс"
бактериального мира. Бактерии выращивали на среде с антибиотиком; с помощью
генной инженерии в них "вшили" способность его обезвреживать, для
чего требовалось наличие в среде определенного сигнального вещества. Всего было
создано две линии бактерий: одни синтезировали требуемое сигнальное вещество
сами (их можно условно назвать "альтруистами"), другие
("эгоисты") были лишены такой способности. В смешанных колониях
"альтруистов" и "эгоистов" сигнальное вещество передавалось
от бактерии к бактерии, позволяя каждой из них противостоять действию
антибиотика. Воочию наблюдать за конкуренцией двух линий можно было благодаря
тому, что "альтруистов" заставили вдобавок синтезировать зеленый
флуоресцирующий белок. Как и следовало ожидать, в смешанных колониях
"эгоисты" размножались быстрее, и доля "альтруистов"
снижалась. Причина в том, что "альтруистам" приходилось тратить часть
энергии на синтез сигнального вещества и флуоресцирующего белка, проигрывая в
скорости размножения.
Все это вполне закономерно. Но чем должен закончиться такой
процесс? То ли стабилизацией численности "альтруистов" на низком
уровне, то ли их полным вымиранием (выбор между этими возможностями зависит от
конкретных значений интенсивности действия антибиотика, способности сигнального
вещества к распространению, энергетических потерь "альтруистов" и
других факторов). Однако в описываемом эксперименте был предусмотрен еще один
важный процесс. Периодически колонии смешивали, а смесь бактерий разбавляли и
высевали на новую среду. Там возникали новые колонии с разным соотношением
"альтруистов" и "эгоистов". Каждая из колоний росла, и в
каждой из них доля "альтруистов" снижалась. Однако (вот что важно!)
те колонии, где доля "альтруистов" была выше, росли быстрее. Потом
опять конец цикла, и снова образование новых колоний. Итог: в каждой колонии
доля "альтруистов" снижалась, а во всей экспериментальной популяции -
росла!
Авторы эксперимента трактуют это как иллюстрацию парадокса
Симпсона (парадокса объединений).
Ограничивается ли описанное сравнение "альтруистов"
и "эгоистов" подтверждением этого парадокса? Конечно, нет! Одна из
тем, вызывающих ожесточенные споры в эволюционной биологии, -
противонаправленность индивидуального и группового отбора. Понятно, что если
действие некоего наследственного задатка приводит к увеличению доли его
носителей в среде, дальнейшая эволюция пойдет в его сторону. А могут ли
закрепиться признаки, которые невыгодны особи, но выгодны группе? Иногда да.
Социобиологи подробно разработали модели, показывающие, что
если некий ген заставляет особь жертвовать собой во имя других носителей этого
же гена, доля таких носителей может возрастать.
А если акт "альтруизма" совершается во имя всей
группы, без оценки родства? В каждой из колоний были как
"альтруисты", так и "эгоисты", и несмотря на то, что доля
"альтруистов" неизменно снижалась, их образ жизни восторжествовал и
стал нормой!
Как показал описанный эксперимент, динамичная среда, где
происходит образование новых групп и имеет место конкуренция между ними, может
приводить к распространению качеств, носители которых проигрывают внутри каждой
из групп, способствуя, тем не менее, общему успеху. В этом исследовании
показано не только действие одного из статистических парадоксов, но и доказана
эффективность группового отбора.
И весьма вероятно, что социальные структуры, где обучают
собратьев, заботятся о больных, сочиняют музыку или читают
"Компьютерру", тоже возникли с участием подобных процессов.
Воскрешение РНК-мира
К цепочке экспериментов, моделирующих возникновение жизни,
добавилось новое звено. Но прежде чем приступить к рассказу о нем, необходимо
вступление.
«КТ» уже обсуждала представления о том, что жизнь появилась
от случайного взаимодействия примитивных органических молекул. Эту точку зрения
обычно приписывают ученым те, кто зарабатывает очки на ее разоблачении. Науке
уже давно ясно, что сложность не является результатом однократной случайности,
а возникает в результате запоминания развивающейся системой целой серии
случайных выборов, когда изменения в определенном направлении фильтруются неким
направляющим механизмом. Подобный механизм фильтрации неплохо описан. Для
организмов это естественный отбор, для молекул - отбор автокатализаторов и иные
виды отбора относительно более конкурентоспособных структур и процессов.
Современная жизнь построена на взаимодействии двух классов
биополимеров - белков и ДНК. Но удивительным образом при их взаимодействии
«между» ними почти всегда оказывается другой (помимо ДНК) класс нуклеиновых
кислот - РНК. Может, наблюдаемое нами состояние - результат наложения новых
молекулярных механизмов на старую РНК-основу? РНК пригодна для хранения и
передачи наследственной информации (пусть и хуже, чем более устойчивая ДНК) и
может обладать каталитической активностью (конечно, уступая в этом отношении
ферментам - белкам-катализаторам). В 1986 году нобелевский лауреат Уолтер
Гилберт подвел итог вызревавшим в течение двух десятилетий идеям и описал
«РНК-мир» - форму преджизни, где и каталитическую, и информационную функции
выполняли молекулы рибонуклеиновой кислоты. РНК-мир, считает Гилберт, возник в
результате отбора на молекулярном уровне и привел к многократному повышению
эффективности дальнейшего отбора.
Можно ли экспериментально доказать, что происходило на Земле
почти четыре миллиарда лет назад? Увы, нет. Но попытки моделирования этих
процессов способны дать весьма ценное теоретическое знание.
В конце 1990-х А.Б. Четверин и другие сотрудники Института
белка РАН научились выращивать колонии размножающихся и конкурирующих друг с
другом клонов РНК на чашках Петри с гелем, содержащим РНК-мономеры и
необходимые ферменты. Достаточно выставить такую чашку на открытый воздух, как
в ней начнут размножаться молекулы РНК. Но это, конечно, еще не РНК-мир, ведь
сборку новых полимеров осуществляют белки.
В начале 2008 года были опубликованы результаты работ
сотрудников Исследовательского института Скриппса (SRI) в Калифорнии, создавших
установку для размножения и отбора рибозимов (РНК-катализаторов). Авторам
исследования удалось создать условия, при которых лучшие катализаторы
размножаются на среде с белками и мономерами нуклеиновых кислот быстрее худших.
Молекулы РНК сами «решают», как изменить свою структуру в соответствии с
требованиями экспериментаторов.
И, наконец, начало 2009 года. В том же SRI запущен процесс
эволюции молекул РНК, которые обходятся без белков. Ученым удалось подобрать
пары молекул РНК, каждая из которых синтезирует свою напарницу. Поселенные на
соответствующую среду, разные пары начинают конкурировать друг с другом и
совершенствовать свою структуру.
Что дальше? Как скоро будет создан реактор, где оживет
преджизнь? Если все рассуждения верны, ДНК-РНК-белковый мир вытеснил когда-то
своего предшественника - РНК-мир. Вытеснил, чтобы воскресить нашими руками
заново?
Голиаф-киборг
Начиная с каменноугольного периода за господство на суше
борются представители двух «вершин» эволюции животного мира - тип членистоногих
и подтип позвоночных (третья «вершина» - класс головоногие - не смогла освоить
ни пресные воды, ни сушу). Трудно представить, насколько мы отличаемся от
членистоногих! В начале XIX века Этьен Жоффруа Сент-Илер, один из предтеч
эволюционной теории, попытался вывести «план построения» позвоночных из
членистоногих. Чтобы сделать это, потребовалось вывернуть тело наизнанку (у
членистоногих скелет наружный, а у позвоночных - внутренний) и перевернуть его
вверх тормашками (брюшной стороне тела членистоногих соответствует спинная
сторона позвоночных; как ни удивительно, последний вывод подтверждается и
современной наукой).
Каждая из групп приспособилась и заняла подходящие для нее
экологические ниши. Признавая наличие массы исключений, можно резюмировать, что
членистоногие многочисленны, приспособлены к строго определенным условиям жизни
и в своем поведении отрабатывают жестко заданные программы. Каждая особь
позвоночных ценнее, и она чаще учитывает индивидуальный опыт. Эволюционная
гибкость членистоногих связана с обилием их видов, а позвоночных - с
изменчивостью поведения, а иногда даже и с индивидуальными отличиями между
особями одного вида.
А какую из групп лучше использовать для создания киборгов -
управляемых устройств на биологической основе? Перед специалистами из
Университета Беркли в Калифорнии этот вопрос не стоял. Недавно они представили
ролик, на котором запечатлен полет радиоуправляемого жука-голиафа. Больше того,
ученые заявили о готовности наладить выпуск подобных киборгов в самое ближайшее
время. На жуке было укреплено устройство весом 1,3 г. Оно управляется по радио
и подает сигналы на шесть электродов, внедренных в мускулатуру и нервную
систему жука. Только представьте себе радиоуправляемого жука, направление
полета которого изменяется по воле экспериментатора!
Жук-голиаф получил такое название не случайно, он один из
самых крупных и сильных среди себе подобных. Это насекомое способно поднять в
воздух примерно три грамма груза - а значит, имеется запас, который можно
использовать, например, для установки крохотной видеокамеры. Решать в первую
очередь военным, ведь именно они финансируют исследования. Но сами создатели
голиафа-киборга убеждены, что для него найдутся и мирные применения.
А если не вешать на жука дополнительную видеотехнику, а задействовать
его собственные глаза? И облегчить искусственный модуль управления за счет
максимального использования нервной системы биологического носителя? Вот только
размножаться, производя таких же киборгов, как они сами, техножуки не смогут.
Или этому стоит порадоваться, ведь, как-никак, речь идет о представителе
конкурирующей с нами группы животных?