Сон и бодрствование у человека и животных
Реферат
тема: Сон и
бодрствование у человека и животных
Введение
С древнейших времен, вероятно, еще с тех пор, как человек обрел сознание
и только начал мыслить, его глубоко интересует вопрос о природе и механизмах
одного из самых таинственных, и даже мистических феноменов человеческой психики
- сон. Как только человек стал осознавать свои сновидения, он начал пытаться
найти им объяснение и задумываться об их происхождении. Поэтому за тысячелетия
человеческой цивилизации накопилось большое количество различных гепотиз и
теорий сна. В конце XIX в. Аршанский
разделил историю изучения сна на четыре периода: фетишистский, теологический,
метафизический и эмпирио-психолого-физиологический.
Фетишистский нашел свое выражение в народных воззрениях, поверьях и
сказках - удивительно похожих у древних и современных народов. Среди них мы
находим предания о странствующем духе, исчезающем во время сна и возвращающемся
утром; древние греки рассматривали сон как дар, посылаемый людям богом сна
крылатым Морфеем, одним из сыновей бога Гипноса.
Однако уже и тогда высказывались глубокие материалистические взгляды на
сон. Так еще Демокрит считал, что сущностью сна является продолжающаяся
автоматическая деятельность мозга при отсутствии восприятия.
Средние века мало, что добавили к уже имеющимся представлениям, но уже в XIX в. Начинается активная
исследовательская деятельность в области изучения механизмов сна. Поочередно,
ведущее положение захватывала та или иная теория.
Глава 1.
История развития представления о системе сна-бодрствования
Современные представления о механизмах сна имеют свою
историю. Многие ранние гипотезы о природе сна в большинстве своем являются
умозрительными и представляют в настоящее время лишь исторический интерес. С
ними можно ознакомиться в известных монографиях [Эпштейн, 1928; Рожанский,
1954; и др.]. Научный период в изучении сна в норме и патологии начался в конце
прошлого века с развитием невропатологии и психиатрии, способствовавших
изучению мозговой деятельности при различных патологических состояниях. Интерес
к мозговым структурам, регулирующим смену сна и бодрствования, в значительной
степени связан с клиническими наблюдениями за больными эпидемическим
энцефалитом, характерным для которого является нарушение цикла бодрствования и
сна, в результате поражения серого вещества вокруг сильвиева водопровода и
других подкорковых образований. Указанные поражения клинически проявляются
повышенной сонливостью, либо синдромом бессонницы или чередованием этих
состояний. Важными оказались наблюдения [Mauthner, 1890], при которых возникала
патологическая сонливость и глазодвигательные расстройства. Автор обратил
внимание на роль гипоталамической области в регуляции сна и бодрствования.
Наблюдения за больными летаргическим энцефалитом выявили зависимость сонливости
или бессонницы от локализации воспалительного процесса… Сон, по мнению автора,
наступает вследствие торможения таламуса и коры больших полушарий,
осуществляемого "центром" сна, расположенным в гипоталамусе и
представленным двумя функционально различными отделами, причем поражение
орального отдела приводит к бессоннице, нарушение же каудального центра влечет
за собой патологическую сонливость.
Следующим этапом в исследовании сна были физиологические
исследования Гесса с электрическим раздражением ограниченного участка
гипоталамуса, имеющего обширные связи со средним мозгом и корой больших
полушарий. В результате раздражения у кошек наступал сон, который продолжался и
после прекращения раздражения. На этом основании была сформулирована теория подкоркового
центра сна [Hess, 1931]. По мнению автора, центр сна состоит из двух
антагонистических частей, обладающих трофотропной и эрготропной функцией,
причем наступление сна связывалось с превалированием трофотропной функции.
Эти представления были подвергнуты критике И. П. Павловым,
отвергнувшим узко локализационистское представление о "пассивном" и
"активном" сне. Первый из них, по его мнению, наступает вследствие
отпадения массы афферентных раздражений. Активный сон возникает как результат
активного внутреннего торможения корковых клеток. Сон людей и животных в
физиологических условиях И. П. Павлов справедливо рассматривал как активный
процесс. Однако, в связи с несовершенством технического обеспечения
исследований того времени было невозможно дать точную идентификацию этого
активного процесса, и в этой связи предположение о разлитом корковом торможении
не получило в дальнейшем подтверждения. Необходимо отметить, что высказанная И.
П. Павловым корковая теория сна не исключала участия и подкорковых структур в
механизмах сна.
Развивая ряд положений павловской школы, П. К. Анохин [1945]
выдвинул корково-подкорковую теорию механизмов развития сна, согласно которой
активный сон наступает вследствие торможения корковой деятельности. Корковые
области, особенно ее лобные отделы, оказывают постоянное тормозящее влияние на
некоторые гипоталамические центры. Торможение корковой области способствует
высвобождению гипоталамических структур, остающихся активными во время сна.
Высвобожденные гипоталамические центры оказывают воздействие на зрительные
бугры, в которых блокируется прохождение импульсов, направляющихся к мозговой
коре от органов чувств, висцеральных органов, мышц.
Мощным стимулом в развитии современных представлений о
природе сна явились ставшие классическими опыты Бремера [Bremer, 1936] с
препаратами "encephale isole", "cerveau isole", а позднее и
исследования Moruzzi, Magoun [1949], Gellhorn [1953]. В результате этих
исследований возникло предположение о локализации механизмов поддержания
бодрствования в стволе головного мозга. В указанных экспериментах развитие
картины поведенческого и электроэнцефалографического сна объяснялось отсечением
от полушарий мозга активирующей восходящей ретикулярной формации. Перерезка
лемнисковых систем при условии интактности ретикулярной формации не вызывала
подобных изменений. Наступление сна наблюдалось только при разрушении
ретикулярной формации. Таким образом была установлена роль активирующей
ретикулярной формации в возникновении и поддержании бодрствования. На основании
опытов с разрушением различных отделов ствола мозга Клейтман [Kleitman, 1964]
также пришел к выводу, что "главным, если не единственным механизмом
чередования сна и бодрствования является ретикулярная формация ствола
мозга", которая при активации индуцирует и поддерживает бодрствование, а
при инактивации приводит ко сну.
Однако в регуляции уровня бодрствования, помимо ретикулярной
формации среднего мозга, важную роль играют диффузная и специфическая
таламические системы [Lindsley, 1952]. Автор выдвигает концепцию
"sleep-wakefulness continuum", причем этот цикл включает различные
промежуточные состояния. Различная степень бодрствования зависит от участия,
помимо ретикулярной активирующей системы, и таламических систем. Отмечая
значение активирующих структур заднего гипоталамуса, Tokizane [1965] считает,
что через специфические и неспецифические системы ядра таламуса осуществляется
гипоталамический контроль активности коры больших полушарий, причем гипоталамус
может выполнять более важную роль в регуляции сна и бодрствования, чем
ретикулярная формация среднего мозга.
На основании экспериментальных исследований Rossi, Zancetti
[1960] предполагают наличие в стволе мозга десинхронизирующего (пробуждающего)
и синхронизирующего или вызывающего сон механизмов, находящихся в
антагонистических отношениях. Авторы считают, что сон не пассивное состояние,
вследствие отсутствия бодрствования, а результат активной деятельности мозга.
Экспериментальные исследования [Moruzzi, 1963] подтверждают наличие
синхронизирующих структур в каудальных отделах ствола мозга, действующих как
антагонисты восходящей ретикулярной активирующей системы. Опыты Ваtini с
соавторами [1958] …на среднемостовом претригеминальном препарате указывают на
наличие в верхних отделах моста (между перерезками cerveau isole и
претригеминальной) структур, играющих важную роль в поддержании бодрствования.
По предложению автора, состояние сна препарата cerveau isole обусловлено
отсутствием связи с этими структурами (перерезка произведена выше).
По мнению Моруцци, в стволе мозга имеются две тонически
активные системы - синхронизирующая и десинхронизирующая. Синхронизирующие
системы ствола мозга являются функциональными антагонистами активирующей
ретикулярной формации [Morussi 1962, Нарикашвили, 1965]… Преимущественная
локализация синхронизирующих структур - в каудальном отделе ствола мозга, в
зоне солитарного тракта тройничного нерва [Moruzzi, 1962], отсечение которого
от вышележащих отделов ствола сопровождается непрерывным бодрствованием, а
раздражение этой зоны током низкой частоты вызывает поведенческий сон с
синхронизацией на ЭЭГ. Подобные явления наблюдались при раздражении нижних
отделов ствола на людях во время хирургического вмешательства [Майорчик и др.,
1959]. Кпереди и латеральнее синхронизирующей системы Моруцци установлено
наличие еще одной синхронизирующей системы [Bonvallet, Allen, 1963], при
экспериментальном разрушении которой наблюдается учащение спонтанных
пробуждений и значительное удлинение периода активации после воздействия
афферентных раздражителей. Похоже, что эта структура осуществляет контроль за
активностью ретикулярной активирующей системы, смягчая эффекты ее возбуждения.
Наступление сна, по мнению автора, является следствием активного действия
синхронизирующих структур мозгового ствола, противоположно направленного
действия восходящей ретикулярной активирующей системы. При этом наряду с
несомненным влиянием химических медиаторов и гуморальных механизмов на
активность синхронизирующих аппаратов подчеркивается и возможная роль
кортикофугальной импульсации, способствующей реализации синхронизирующего
влияния соответствующих аппаратов продолговатого мозга.
Синхронизирующий эффект в ЭЭГ наблюдается при раздражении и
других структур мозга. При электрической стимуляции головки хвостатого ядра
наблюдается появление характерных веретен в коре и поведенческое торможение
[Buchwald et al., 1961]. По мнению Мэгуна [I960], при низкочастотном
раздражении хвостатого ядра наступление сна происходит в результате включения
таламокортикальной системы "рекруитирования".
Этот эффект, описанный впервые Dempsey, Morrison [1942],
возникает при низкочастотном раздражении ядер медиального таламуса.
Таламическая система, с одной стороны, оказывает активирующее влияние на кору
больших полушарий, являясь частью восходящей активирующей системы, а
раздражение ее ядер током низкой частоты приводит к развитию сна с
синхронизирующим эффектом в коре больших полушарий … К. структурам, при
низкочастотной стимуляции которых наблюдаются электроэнцефалографические
(синдронизация) и поведенческие признаки сна, относится передняя группа ядер
зрительных бугров и интраламинарная система [Hess, 1954], хвостатое ядро
[Buchwald et al., 1961], преоптическая зона и область диагональной связки
[Clemente, Sterman, 1967]. На основании опытов на кошках с раздражением
структур переднего мозга, и особенно латеральной преоптической области,
Hernandez-Peone [1963, 1964] предполагает наличие "гипногенного
цикла", простирающегося от медиального отдела верхней части преоптической
зоны в медиальный отдел покрышки мозга вдоль по медиальному пучку переднего
мозга, включая и ядра Бехтерева и Гуддена.
Таким образом, на всех уровнях мозговой оси, начиная с
каудальных отделов ствола, представлены синхронизирующие (сомногенные) и
десинхронизирующие (активирующие, пробуждающие) механизмы, нередко взаимно
перекрывающие друг друга с преимущественной концентрацией на определенном
уровне. Преимущественная концентрация элементов сомногенной системы находится в
каудальных отделах ствола (система Моруцци), синхронизирующей системе
медиального таламуса, хвостатом ядре, латеральной преоптической области и
базальных отделах переднего мозга. Образования сомногенной системы находятся в
конкурентных отношениях с активирующей пробуждающей системой,
структурно-функциональной основой которой является активирующая ретикулярная
формация ствола мозга, в том числе таламическая активирующая система. Обе
системы имеют кольцевые связи с лимбическими структурами и неокортексом [Hess,
1954; Анохин, 1957; Моруцци, 1962; Clemente, Sterman, 1963]. Различные отделы
синхронизирующей системы выполняют специфические функции. Системы каудального
ствола оказывают угнетающее влияние на активирующую ретикулярную формацию,
облегчая тем самым функционирование синхронизирующих образований таламуса.
Сомногенные образования латеральной преоптической зоны и базальных отделов
переднего мозга, по-видимому, осуществляют интеграцию цикла бодрствование - сон
с уровнем эндокринно-метаболических процессов в организме. Синхронизирующие
аппараты таламо-каудатно-неокортикального уровней, вероятно, обеспечивают
интеграцию цикла бодрствование - сон с внешнесредовыми условиями деятельности.
Глава 2. Фазы сна и их циклическая организация
Фаза медленного сна (синонимы: медленноволновой сон,
синхронизированный, ортодоксальный) у животных и человека характеризуется
замедленной ритмикой, в частности, появлением в ЭЭГ сонных веретен (сигма-ритм,
веретенообразные комплексы колебаний частотой 12-16 периодов в 1 сек.) и
высокоамплитудной дельта-активностью [Davis et al., 1938; Шпильберг, 1955].
У человека в фазе медленного сна выделяют ряд стадий,
характеризующихся определенной электроэнцефалографической картиной и глубиной
сна [Loomiset., 1937; Davis et al., 1938; Reehtschabben, Kales, 1968]. Стадия
"дремоты" (I стадия, стадии А и В) характеризуется исчезновением
альфа-ритма и уплощением на ЭЭГ с появлением низкоамплитудных медленных и
быстрых колебаний потенциала.
Вторая стадия (II стадия, С) характеризуется появлением
"сонных веретен" и К-комплексов (двух- или трехфазные медленные волны
длительностью 0,5 с, возникающие непосредственно перед сонными веретенами).
Стадии III и IV составляют дельта-сон, или глубокий сон,
характеризующийся наличием высокоамплитудных дельта-волн, причем в третьей
стадии наряду с дельта-волнами присутствуют сонные веретена. В IV стадии
преобладающими являются высокоамплитудные дельта-волны, которые по
международной классификации [Rechtschaffen, Kales, 1968] занимают в этой стадии
более 50% эпохи регистрации. В III стадии, согласно этой стандартизации,
дельта-волны высокой амплитуды должны составлять от 20 до 50%. У животных в
медленном сне обычно выделяют легкую, или поверхностную, стадию и глубокую, или
собственно медленно-волновой сон.
Электрическая активность диэнцефально-стволовых структур обычно
соответствует корковой активности, лишь иногда во время медленного сна в
специфических ядрах зрительного бугра и подушки наблюдается десинхронизация
[Hess et al, 1953; Jouvet, 1962]. В гиппокампе во время медленного сна
регистрируется высоковольтная тета-активность [Cadilhac et al., 1961; Jouvet,
1962] и дельта-волны [Ониани и др., 1970].
Механизмы, ответственные за регуляцию и поддержание различных
проявлений медленного сна, во многом еще не ясны, имеют сложную организацию и
функционируют в тесном контакте. Поскольку начало сна всегда, исключая
нарколепсию, идет с развертывания стадий медленного сна, целесообразным
является описание представлений о механизмах их включения. Как уже упоминалось,
с открытием роли ретикулярной формации в обеспечении состояния бодрствования
[Moruzzi, Mayoun 1949, Possi, Zancetti, 1960] наступление медленного сна
рассматривается как следствие временной блокады активирующей ретикулярной
системы и включения синхронизирующих таламо-кортикальных механизмов, описанных
еще в 1929 г. Гессом. Регистрация сонных веретен одновременно в таламусе и
коре, впервые осуществленная Dempsey, Morrison [1942], послужила поводом для
размышлений о месте первичной их генерации. Авторы предположили первичную их
генерацию в таламусе с дальнейшим распространением в кору больших полушарий.
Согласно представлениям ряда исследователей генерация сонных
веретен осуществляется неспецифической таламо-кортикальной проекционной
системой, пусковую роль в которой выполняют таламические ядра [Morrison,
Dempsey, 1942; С. П. Нарикашвили, 1969]. Вместе с тем экспериментальные
исследования указывают на важную роль коры, поскольку после полной экстирпации
последней наблюдается исчезновение веретен в таламусе [Jouvet, Michel, 1958].
Дальнейшими исследованиями показано, что для генерации сонных веретен
достаточна сохранность лишь небольшого участка коры, в частности орбитальной
коры [Velasco, Landsley, 1965]. При рассмотрении роли специфических и
неспецифических ядерных образований таламуса в генерации веретен предпочтение отдается
неспецифическим ядрам [Knott, Ingram, 1951].
Как уже указывалось, сомногенные, синхронизирующие механизмы
не ограничиваются таламо-кортикальной системой. Реакции синхронизации на ЭЭГ
наблюдаются при низкочастотном раздражении многих структур мозга, тогда как при
высокочастотной стимуляции тех же (синхронизирующих) структур наступает
десинхронизирующий эффект. Все это может свидетельствовать о широкой
представленности структур, ответственных за регуляцию цикла бодрствование -
сон, и лишь о преимущественной концентрации в той или иной области.
Фаза быстрого сна (синонимы:
парадоксальный сон, ромб энцефалический, активированный, Rein - сон, сон со
сновидениями и др.) характеризуется следующими основными
электрофизиологическими признаками: 1) реакцией десинхронизации на ЭЭГ; 2)
быстрыми движениями глаз (БДГ), регистрируемыми на электроокулограмме; 3)
падением тонуса субментальных и шейных мышц, регистрируемых на ЭМГ у человека
(у животных шейных мышц) [Aserinsky, Kleitman, 1953; Dement, Kleitman, 1957]. Динамика
изменения электроактивности соматической мускулатуры других областей не
претерпевает столь значительных изменений в быстром сне, сравнительно с фазой
медленного сна [Jacobson et al., 1964].
В картине этой фазы у человека и животных имеется некоторая
специфика. У людей картина ЭЭГ быстрого сна характеризуется вариабельностью
различных ритмов - от "плоской" ЭЭГ до ритмов всех частот, включая
фрагменты альфа-ритма. Считается характерным для этой фазы сна появление
"пилообразных" разрядов, нередко предшествующих вспышке БДГ. В фазе
быстрого сна животных наблюдается быстрая ритмика до 30 периодов в 1 сек
[Jouvet, 1962].
Появлению эпизода быстрого сна обычно предшествует
"переходная" стадия, в которой имеет место сочетание
электрографических признаков обеих фаз сна [Goldsteinas et al., 1966;
Шеповальников, 1971]. В переходной стадии, составляющей обычно 1-5% общей
длительности сна здоровых людей, наблюдается значительная представленность
вертекс-потенциалов (острые волны амплитудой около 200 мкВ, лучше выраженные в
теменном отведении), К-комплексов, замедленных колебаний, сонных веретен,
быстрой ритмики и участков "плоской" кривой, появление единичных
движений глаз, снижение активности на электродермограмме [Goldsteinas et al.,
1966; Lairy et al., 1966].
Переходный период констатирован также у кошек [Ониани и др.,
1970]. По мнению ряда авторов, он может являться результатом кратковременного
включения ретикулярной активизирующей системы. В гиппокампе электроактивность
при переходе из медленного сна в быстрый сон сменяется регулярным
высокоамплитудным тета-ритмом [Grastyan, 1961; Lissak et al., 1961], что
указывает на активацию лимбических структур [Ганжа, Богач, 1972].
В ряде исследований показана неоднородность электроактивности
гиппокампа в фазе быстрого сна [Ониани и др., 1970]: короткие эпизоды
тета-ритма, в среднем около 1 мин, периодически сменяющиеся дельта-активностью,
причем тета-активности соответствовали выраженные вегетативные изменения и
появление быстрых движений глаз. На уровне межуточного мозга в фазе быстрого
сна электрическая активность соответствовала корковой ритмике [Jouvet, 1962;
Tokizane, 1965].
Важными электрофизиологическими феноменами являются изменения
на электроокулограмме, отражающие движения глаз во время сна и являющиеся
следствием сдвига корнео-ретинального потенциала [Dement, 1966]. Во время сна
можно наблюдать движения глазных яблок двух видов. Медленные движения глаз
длительностью 1-2 с наиболее выражены при засыпании и дремотных стадиях. В фазе
быстрого сна появляются быстрые движения глаз частотой 1-2 с, одиночные или
группирующиеся в виде пачек.
Констатировано приблизительно равное соотношение быстрых
движений глаз одиночных и в пачках [Jeannerod et al., 1965]. У недоношенных
детей в большинстве случаев имела место тенденция к более редким движениям глаз
[Евсюкова, 1977]. Характерным для фазы быстрого сна у животных являются
понто-геникуло-окципитальные пики (ПГО-пики, спайки), возникающие в области
варолиева моста и распространяющиеся в коленчатые тела, верхние бугорки четверохолмия
и зрительную кору [Jouvet et al., 1959].
В спинном мозгу активность нейронов в фазе быстрого сна резко
заторможена, на что указывает почти полное отсутствие Н-рефлекса при сохранном
М-ответе [Giaquinto, Pompeiano, 1963]. Это угнетение синхронно с быстрыми
движениями глаз прерывается фазическим облегчением, что, очевидно, и
обусловливает наблюдаемые в эти периоды подергивания конечностей… Специальными
исследованиями показано, что во время быстрого сна имеет место
постсинаптическое тоническое торможение мотонейронов спинного мозга, на которые
в периоды проявления быстрых движений глаз наслаивается фазическое
вытормаживание пресинаптической природы [Baldissera et al., 1966; Kawamura,
Pompeiano, 1969]. Изменения активности спинальных нейронов имеют центральное
происхождение и устраняются после разрушения медиального и нисходящего
вестибулярных ядер и после перерезки вентрального и дорзального канатиков, по
которым направляются к спинному мозгу эти влияния.
В фазе быстрого сна различают фазические и тонические
компоненты (по электрофизиологическим и вегетативным показателям). Тоническими
проявлениями быстрого сна являются изменения на ЭЭГ, тета-активность
гиппокампа, падение мышечного тонуса, торможение спинальных рефлексов.
Фазическими феноменами этой фазы сна являются быстрые движения глаз, ПГО-пики у
животных, подергивания мышц, фазические изменения возбудимости спинальных
нейронов, вегетативные сдвиги - учащение и аритмия пульса и дыхания, перепады
уровня кровотока, КГР и др.
Структурами, ответственными за фазу быстрого сна, согласно
экспериментальным исследованиям, являются ретикулярные ядра варолиева моста
[Jouvet, 1964]. При отсечении мозгового ствола выше варолиева моста наблюдается
полное исчезновение признаков быстрого сна, в то время как у понтинных животных
продолжает регистрироваться эта фаза сна. Разрушение ретикулярных ядер моста
ведет к устранению быстрого сна, а половинные перерезки на этом уровне
сопровождаются асимметрией корковой активности в фазе быстрого сна [Jouvet,
1962]. Экспериментальные исследования на животных способствовали выявлению
структур, ответственных за отдельные электрофизиологические феномены быстрого
сна. Разрушение locus coeruleus ведет к прекращению падения мышечного тонуса в
фазе быстрого сна [Jouvet, Delorme, 1965]. Коагуляция структур покрышки моста
сопровождается исчезновением ПГО-пиков [Jouvet, 1962]. Десинхронизация корковой
активности в фазе быстрого сна, как показано в этих исследованиях, не зависит
от влияния активирующей ретикулярной формации, как и разрушение диэнцефальной
области, способное вызывать нарушение активации коры в бодрствовании, не
вызывает подобного действия в фазе быстрого сна. Показана роль вестибулярных
ядер в продукции фазических компонентов этой фазы сна [Pompeiano, Morrison,
1965]. Разрушение медиальных и нисходящих вестибулярных ядер с обеих сторон
вело к прекращению вспышек быстрых движений глаз, фазических изменений частоты
пульса и артериального давления, миоклонических подергиваний. Иного мнения
придерживается И. М. Гильман [1971], которая считает, что в происхождении
быстрого сна и его отдельных компонентов главное значение имеет торможение
активности вестибулярных ядер. Имеется и другая точка зрения…, согласно которой
вестибулярные ядра участвуют, но не являются обязательными в генезефазических
компонентов быстрого сна.
Благодаря методике микроэлектродного исследования на животных
в свободном поведении…, стало возможным проследить в динамике активность
отдельного нейрона в цикле сон-бодрствование. Исследования спонтанной и
вызванной нейронной активности не подтверждают гипотезы, со гласно которой сон
рассматривается как "разлитое корковое торможение", так как многие
данные указывают на некоторое повышение уровня возбудимости коры и ряда
подкорковых структур во время сна, особенно в фазе быстрого сна. Согласно этим
данным, во время сна не происходит общего торможения, а происходит
реорганизация активности нейронов, которая в значительной степени отличается в
различных функциональны; состояниях, в частности бодрствовании, фазе медленного
и бы строго сна. В зрительной коре кошки в медленном сне спайковая активность
многих нейронов возрастает [Hubel, 1960]. В фазе быстрого сна активность
большинства исследованных нейронов коры повышается по сравнению с медленным
сном [Фельдман, Владимирский, 1968], а в некоторых областях коры (зрительная,
слуховая) достигает и даже превосходит по этому показателю активное
бодрствование [Evarts, 1965]. По мнению Evarts [1967], во время сна
увеличивается активность тех нейронов, которые менее активны во время бодрствования.
При сензорном пробуждении у таких нейронов наблюдается длительное уменьшение
частоты разрядов [Creutzfeldt, Jung, 1961]. сон бодрствование нейрохимический физиологический
При исследовании клеточных популяций подкорковых структур у
человека во время хирургического лечения паркинсонизма было отмечено некоторое
снижение импульсной активности в медленном сне (в отдельных структурах может
даже повышаться) и значительное ее повышение в фазе быстрого сна… В нейронной
активности ядер зрительных бугров, гиппокампа и ретикулярной формации, как и в
коре, при наступлении медленного сна наблюдается усиление группирования
спайков, увеличение их частоты внутри групп и интервалов между ними
[Huttenlocher, 1961; Мухаметов, 1977]. В фазе быстрого сна это группирование
нарушается за счет резкого увеличения частоты разрядов. Повышение активности
нейронов в фазе быстрoго сна констатировано в таламусе [Filion et al., 1969] и
гиппокампе [Noda et al., 1969], тогда как в медленном сне частота разрядов
нейронов этих образований снижалась.
При исследовании вызванной активности в состоянии
бодрствования и в различных фазах сна получены неоднозначные результаты.
Вызванные потенциалы коркового ответа на зрительную стимуляцию обнаруживали
снижение в медленном сне, еще н большей степени в быстром сне и практически
исчезали в моменты появления быстрых движений глаз и ПГО-пиков [Giaquinto et
al., 1963; Mouret et al., 1963]. Снижение корковых вызванных потенциалов при
слуховой стимуляции [Bremer, Bennet, 1949; Jouvet, 1962] наблюдалось в
медленном сне и еще в большей степени в фазе быстрого сна, особенно синхронно с
быстрыми движениями глаз. Другие авторы сообщают об увеличении вызванного
ответа в коре во время медленного сна и быстрого сна [Guilbaud, 1970] с
одновременным увеличением вызванной нейронной активности [Mukhametov,
Rizzolatti, 1970] или без достоверных ее изменений [Lelord, Maho, 1969].
При исследовании порогов коркового ответа на различные
афферентные раздражители в быстром сне обнаруживается значительное повышение их
по сравнению с медленным сном как на зрительную [Mouret et al., 1963], так и на
слуховую стимуляцию [Jouvet, 1962]. Отмеченная диссоциация между ЭЭГ-картиной,
близкой к таковой в состоянии активного бодрствования, повышением порогов
пробуждения собственно и характеризуется "парадоксальность" фазы
быстрого сна. Показано в эксперименте [Ониани, Мольнар, 1971] различие порогов
поведенческого ЭЭГ-пробуждения в этой фазе сна: порог пробуждения в быстром
сне, который устанавливается на основании депрессии гиппокампального тета-ритма
при раздражении мезенцефалической ретикулярной формации, мало отличается от
порога пробуждения в фазе медленного сна, в то время как порог поведенческого
пробуждения в быстром сне оказался гораздо выше, что авторы объясняют
результатом торможения специальных нейронов в быстром сне, препятствующего
поведенческому пробуждению.
При анализе электрополиграфических данных, регистрируемых
непрерывно на протяжении ночи, обращает на себя внимание циклическая
организация сна. В естественных условиях у здоровых людей сон начинается с фазы
медленного сна, с первой стадии постепенно углубляется до самых глубоких стадий
и затем закономерно сменяется фазой быстрого сна. Это и составляет цикл сна. В
течение ночи у здоровых лиц наблюдается 4-5 циклов, в среднем повторяющихся
через 90-120 мин. Первый цикл начинается с момента засыпания и заканчиваете
первым эпизодом быстрого сна, последующие циклы - от конца предыдущего периода
быстрого сна до окончания следующего эпизода быстрого сна. Длительность
эпизодов, минимальная в первых циклах, к утру постепенно возрастает, в то время
как представленность наиболее глубокого дельта-сна (III и IV стадии вместе),
максимальная в первых 2-3 циклах, прекращаете: в последних циклах сна (Dement,
Kleitman, 1957].
Сон рассматривается как одна из фаз циркадного ритма
[Sollberger, 1969; Berlucchi, 1970], в котором выявляется периодичность быстрой
фазы, являющейся филогенетически древ 90-минутным ритмом (Snyder, 1966].
Доказано, что смена фаз сна совершается через равные промежутки со средней
длительностью в 90 мин [Dement, Kleitman, 1957; Globus, 1966] Хотя зависимости
от режима освещения [Kleitman, 1939] происходят изменения сна, тем не менее он
сохраняет свойство периодичности и приурочен только к темному периоду суток.
Инверсия ритма бодрствование-сон при перелетах в новую временную, зону резко
изменяет картину сна: представленность быстрого сна и дельта-сна отмечалась в
первой части ночи. У лиц, засыпающих после трансмеридианального перелета в
утренние часы, по астрономическому времени места вылета, быстрый сон
максимально представлен в начале сна [Evans et al., 1972]. Отмеченный сдвиг по
фазе свидетельствует о том, что этот ритм является эндогенным. В зависимости от
расположения сна в течение суток различают [Hellpach, 1950] рано засыпающих
("жаворонки") и поздно засыпающих ("совы", "городской
тип").
Дневной сон характеризуется сокращением общей его длительности,
эпизодов медленного сна и быстрого сна, уменьшением фазы быстрого сна и
увеличением числа пробуждений [Weitzman et al., 1970]
Как показали эксперименты с депривацией, после полного или
частичного лишения сна у здоровых испытуемых во время восстановительной ночи в
первую очередь компенсаторно повышается представленность IV стадии - наиболее
глубокой стадии медленного сна [Dement, Greenberg, 1966]. Лишение сна
добровольцев в течение нескольких суток сопровождается в первую
восстановительную ночь значительным удлинением IV стадии медленного сна
[Berger, Oswald, 1962; Webb, Agnew, 1965], и лишь на вторую происходит
увеличение фазы быстрого сна. При частичном сокращении общего времени сна на
2,5-3 ч в течение 3-6 суток наблюдается увеличение длительности IV стадии
медленного сна на 40-75% [Dement, Greenberg, 1966]. Значительное увеличение IV
стадии наблюдается и после изолированной ее депривации [Agnew et al., 1964].
Необходимо отметить уменьшение IV стадии при ряде патологических состояний, в
частности при агрипническом и гиперсомническом синдроме.
У пожилых людей снижается длительность III и IV стадий
медленного сна, а относительная продолжительность фазы быстрого сна остается
неизменной [Kales, 1969]. По другим данным [Davies, 1976], с возрастом наблюдается
укорочение обеих фаз сна.
Избирательная депривация быстрого сна в течение нескольких
ночей [Dement, 1960; Sampson, 19651 сопровождается в последующем ее увеличением
(феномен "отдачи"). В период восстановления сна после его депривации
наблюдается повышение интенсивности фазических компонентов, в частности быстрых
движений глаз у людей [Pivik, Foulkes, 1966] и понто-геникуло-окципитальных
спайков у животных [Ferguson, Dement, 1968]. Степень восполнения тонических
явлений, которая оценивалась по общей продолжительности фазы быстрого сна,
составляла около 30%, в то время как процент восполнения фазических
компонентов, определяемый по частоте понто-геникуло-окципи-тальных спайков в
минуту, оказался равным 60% [Dusan-Peyerton et al., 1967].
Помимо обычного увеличения длительности фазы быстрого сна,
после избирательной депривации отмечается включение различных ее фрагментов в
электрополиграфическую картину медленного сна [Dusan-Peyerton et al., 1967].
Аналогичные изменения отмечены у больных, страдающих так называемой
"бессонницей".
Таким образом, как показывают далеко не полные данные опытов
с лишением сна, соотношение фаз медленного и быстрого сна в естественных
условиях является стабильным и при их недостаточности происходит компенсаторное
восполнение устраняемой фазы сна. Это свидетельствует о важной роли обеих фаз
сна в целостной мозговой деятельности.
Глава 3. Нейрохимические механизмы сна
Характерной особенностью прежних представлений о механизмах
сна является противопоставление нейрофизиологических и гуморальных механизмов,
что безусловно противоречит установленным фактам. Дальнейшими исследованиями
показан важная роль обоих этих механизмов, взаимно дополняющих друг друга. До
последнего времени не отвергнуто окончательно мнения о существовании гуморального
фактора, который может быть медиатором наступления сна. Как известно, еще в
начале века была выдвинута гипотеза [Legendre, Piezon, 1910] о гипнотоксине,
способствующей наступлению сна. В исследованиях этих авторов экстракт ткани
головного мозга, сыворотка крови или ликвор собак, лишенных сна в течение 11
суток, вызывали немедленный сон при введении в большую цистерну бодрствующих
собак. К сожалению, в этих исследованиях, как и в других последующих
[Pappenheime'z, 1976], указанный фактор не был химически идентифицирован и не
проводились в последних работах электрополиграфические регистрации сна. Эта
гипотеза снова привлекла к себе внимание в связи со сравнительно недавними
исследованиями [Monnier, Hosli, 1965] на парах кроликов с пере крестным кровообращением.
При вызывании у одного кролика сна электрическим раздражением медиальных
интраламинарных ядер, введение диализата крови другому бодрствующему животному
вызывало сон с медленной дельта-активностью. Диализат крови от бодрствующего
донора, подвергнутого прямо электростимуляции среднего мозга, вызывал усиление
моторной активности, а в ЭЭГ - активацию у кролика-реципиента. К сожалению, до
сих пор не известен химический состав входящих в диализат веществ. По мнению
Monnier [1969], фактором, поступающим в кровь и вызывающим возбуждающий эффект,
является гистамин. К настоящему моменту получены данные, которые указывают на
то, что в развитии сна у реципиента играет существенную роль выделенный
дельта-фактор, представляющий собой вещество пептидной природы с молекулярным
весом] 355-1500 [Sehoenberger et al., 1972]
В то же время наблюдения за сиамскими близнецами показывают,
что наступление сна у близнецов развивается независимо друг от друга
[Алексеева, 1958], что указывает на малую вероятность зависимости сна
исключительно от гуморальных факторов. Исследования на нейронном уровне
обнаруживают характерные различия активности энзимов нервной клетки в
зависимости от состояния сна и бодрствования у кроликов. В частности,
активность сукциноксидазы оказалась высокой в нервных клетках низкой в
окружающей их глии во время сна и обратные отношения наблюдаются в состоянии
бодрствования [Hyden, Lange, 1965].
Накапливается все больше данных, подтверждающих участие
иогенных церебральных аминов в регуляции бодрствования иска [Jouvet, 1962;
Hartmann, 1973]. С помощью новых методов гистохимии, гистофлюоресценции и
нейрофармакологии была укрыта биохимическая специфичность некоторых структур,
участвующих в цикле сон - бодрствование. Благодаря гистохимическим
исследованиям и использованию гистофлюоресцентной техники, позволяющей
идентифицировать моноамины как медиаторы путем обнаружения их в нервных
окончаниях, представляется возможным довольно точно определять топографию
распределения моноаминов. В исследованиях на крысах большинство перикарий,
содержащих серотонин, находится в ядрах системы шва [Anden et al., 1966]. У
кошек отмечена в основном шалогичная топография, с небольшими отклонениями
[Morgane, 1969].
В отличие от серотонинсодержащих нейронов, располагающихся в
основном по средней линии, норадреналинсодержащие нейроны находятся в
латеральной части покрышки ствола [Anden et al., 1966]. Наиболее высокая
концентрация норадреналина обнаружена в системе locus coeruleus в варолиевом
мосту. Офамин сконцентрирован в черной субстанции среднего мозга,
нигростриарной системе.
Косвенным указанием участия биологически активных веществ в
регуляции сна явились колебания их накопления в зависимости от цикла
бодрствование-сон. Во время сна наблюдается увеличение ацетилхолина в головном
мозге [Richter, Cros-sland, 1949]. В опытах на кроликах показано значительное
увеличение выделения ГАМК из перфорированной пиальной поверхности коры во время
сна, по сравнению с бодрствованием [Jasper et al., 1965]. Содержание
норадреналина в мозгу увеличивается после пробуждения [Monnier, 1969].
В настоящее время наиболее разработанной является
моно-гаминергическая теория регуляции бодрствования и сна, предложенная Jouvet
[1969]. Основываясь на комплексных исследованиях в эксперименте на животных,
главным образом кошках и крысах, с использованием стереотаксических,
электрополиграфических, биохимических и гистохимических методов, автор
предложил следующую схему моноаминергических механизмов регуляции цикла
бодрствование - сон и, в частности, фаз сна…
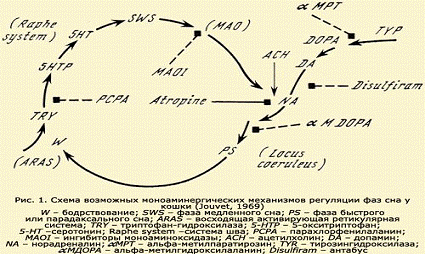
Согласно этой схеме постулируется, что в мозгу млекопитающих
в цикле бодрствование - сон происходят циклические биохимические колебания, от
бодрствования (W), зависящего от ретикулярной активирующей системы (ARAS), к
медленно-волновому сну (SWS), который зависит от нейронов системы шва (Raphe
system), содержащих серотонин (5-НТ), и затем к быстрому или парадоксальному
сну (PS), зависящем от норадреналинсодержащих нейронов (NA), локализованны в
передних отделах ретикулярной формации варолиева моста в neucleus locus
coeruleus, а также в группе нейронов среднее мозга. В схеме представлено также
действие некоторых фармакологических веществ, блокирующих отдельные ступени это
цикла. Известны исходные предшественники биогенных аминов норадреналина и
серотонина (аминокислоты тирозин и триптофан), а также промежуточные метаболиты
и ферменты, принимающие участие в этих превращениях. Установлены различные
ингибиторы и активаторы этих ферментов, а также различные эффекторы,
оказывающие влияние на действие серотонина, дофамина и норадреналина. На основе
этих данных проводились экспериментальные и клинические исследования с
использованием как самих медиаторов, так и его прекурсоров, антагонистов и
различных фармакологических веществ с различной направленностью их действия. В
схеме представлены ступени превращений моноаминов и места приложения некоторых
фармакологических средств. Так, парахлорфенилаланин (РСРА), подавляющий энзим
триптофангидроксилазу (TRY), ведет к снижению мозгового серотонина (5-НТ) и
вследствие этого к полной инсомнии, обратимой при повторном введении
5-гидрокситриптофана (5-НТР), непосредственного предшественника серотонина.
Как показано в эксперименте [Jouvet et al., 1966], локальное
разрушение ядер шва ведет к резкому подавлению медленного сна и отсутствию сна
вообще, при установленном снижении концентрации серотонина в мозгу. Введение
серотонина в ядра шва G0 провождается увеличением представленности
медленного сна [Dahlstrom, Fuxe, 1964] Ингибиторы моноаминоксидазы (МА01), предотвращающие
разрушение серотонина, вызывают увеличение медленного сна и подавление быстрого
сна. [1972] и Bremer [1974] относят к гипногенным струкурам преоптическую
область и бульбарную систему в области ядра солитарного тракта, ставя под
сомнение ключевую роль системы шва. По предположению Koella [1977], существует
двусторонняя связь ядра солитарного тракта и ретикулярной форлации среднего
мозга. Серотонинергическая система шва, по мнению автора, может иметь двойную
функцию - гипногенного передатчика и антипробуждающего фактора. Ряд
исследователей [Jouvet, 1965; Pujol et al., 1968] придают большое значение
норадреналину в регуляции быстрого сна. Эта точка зрения основывается на
экспериментах с последовательным исследованием влияния резерпина, 5-окситриптофана
и ЦОФА на две фазы сна в хроническом эксперименте на кошках [Matsumoto, Jouvet,
1964]. При введении больших доз резерпина наступала редукция быстрого сна,
точнее не наблюдались тонические феномены этой фазы, в частности
соответствующие изменения ЭЭГ и падение тонуса шейных мышц. Однако М. Жуве
придает значение другому признаку быстрого сна у кошек -
понто-геникуло-окципитальным электрическим потенциалам, усиление которых
констатируется при введении находящимся под действием резерпина кошкам ДОФА в
дозе 30-50 мг/кг веса. При этом наблюдается наступление медленного сна,
прерываемого полной картиной быстрого сна в течение 5-6 ч, вслед за этим вновь
развивается резерпиновый синдром. Основываясь на том факте, что при введении
ДОФА наблюдается появление медленного сна и быстрого сна, в то время как
5-гидрокситриптофан в дозе 30-50 мг/кг вызывает лишь короткое возобновление
прерванного медленного сна, М. Жуве склоняется к мысли о возможности развития
быстрого сна под влиянием норадреналина.
По мнению ряда исследователей [Oswald, Priest, 1965; Jouvet,
1972], в организации сна участвуют и холинергические механизмы. При лишении сна
крыс наблюдалось снижение содержания ацетилхолина в переднем мозге, но не в
стволовой его части [Bowerus et al., 1966]. Вместе с тем по другим данным
[Tsuchiya et al., 1969], у того же вида животных только селективное лишение
быстрого сна вызывало падение количества ацетилхолина в теленцефалоне, в то
время как при полном лишении сна там же наблюдалось его увеличение.
Согласно представлениям Jouvet [1969], атропин действует на
холинергический механизм, который, по-видимому, отвечает за запуск
норадренергических механизмов быстрого сна. Показано, что атропин тормозит
тонические процессы быстро сна (но не ПГО-активность), не оказывая в то же
время влияния на развитие медленного сна [Jouvet, 1962; Yamamoto, Domino,
1967]. В то же время, по данным Weiss с соавторам [1964], полученным на крысах,
атропин не оказывает статистически значимого действия ни на длительность
быстрого сна, ни на характер возникновения его периода как при больших, та и
малых дозах препарата. По мнению Itil [1969], дремотное стояние у людей после
приема антихолинергических веществ картине по ЭЭГ отличается от естественного
сна.
При применении ацетилхолина или карбахолина [Hernandez-Peone,
1964], эзерина [Jouvet, 1962] и холиномиметиков [Yamamoto, Domino, 1967]
наступил сон с увеличением фазы быстрого сна и сокращением фазы медленного сна.
По мнению Magherini с соавторами быстрые движения глаз в фазе быстрого сна,
по-видимому, связаны с возбуждением холинергических ретикулярных нейрона (через
вестибулярные ядра). По предположению Koella [19701 "ацетилхолин,
возможно, является переносчиком в нервных сетях, контролирующих процессы сна".
Однако конкретные механизмы участия ацетилхолина в организации сна остаются не
ясными. Предполагается влияние ацетилхолина на медиатор сна посредством
воздействия на ГАМК, снижая ее распад и повышая синтез [Нилова, 1973].
Внимание исследователей в последнее время привлечено
исследованию возможной роли гамма-аминомасляной кислоты (ГАМК) в организации
сна, особенно после выяснения ее значения как нейромедиатора, участвующего в
регуляции процесса торможения. Уже указывалось о тенденции к повышению во время
сна содержания ГАМК в центральной нервной системе [Godin, Mandell, 1965].
Депривация быстрого сна у кошек сопровождалась значительным снижением ее
содержания в четверо холмии и хвостатом ядре, в то время как в коре лобных
долей таламусе и особенно ретикулярной формации среднего мозга содержание ГАМК
возрастает [Micic et al., 1967; 1969; Rakic, 1972]. Вместе с тем торможение
гидроксиламином фермента ГАМК аминотрансферазы, разрушающего ГАМК, вызывало
накопление последней и сопровождалось сокращением быстрого сна. Депривация
быстрого сна приводила к торможению ГАМК-ами нотрансферазы [Mendoza et al.,
1970], а подавление этого фермента динорпропилацетатом сопровождалось
увеличением медленного сна и снижением ПГО-активности.
Близкими по химической структуре естественными компонентами
центральной нервной системы являются гамма-оксибутират и гамма-бутиролактон.
Указанные производные, по данным ряда исследователей [Laborit et al., 1960;
Benda, Perles, I960], стимулировали сон и особенно быстрый сон у кошек (но не у
крыс), причем у кошек они вызывали эту фазу сна и на фоне ее угнетения
атропином, но не после угнетения ингибиторами моноаминоксидазы [Delorme et al.,
1966]. Предполагается, что эти эффекты могут быть получены посредством влияния
на пентозофосфатный шунт [Laborit et al., 1960] либо через воздействие на
метаболизм ГАМК [Fishbein, Bessman, 1964]. Однако имеющиеся в настоящее время
данные не дают ясного представления о конкретных механизмах участия ГАМК и ее
производных в регуляции сна и его различных проявлений.
Нельзя не отметить, что большинство данных
"моноаминной" схемы регуляции и поддержания различных проявлений сна
получены в опытах на животных, кошках и отчасти на крысах. Экстраполяция этих
данных на человека оказалась в ряде пунктов несостоятельной, в связи с видовой
спецификой сна у различных животных. Помимо уже затрагивающихся в процессе
изложения отличий реакций у людей, по сравнению с животными, можно
дополнительно указать, что ингибиторы моноаминоксидаз, вызывая повышение у
людей содержания норадреналина (сравнительно с серотонином), в то же время
вызывали и угнетение быстрого сна [Wyatt et al., 1969], а введение
парахлор-фенилаланина приводило к подавлению быстрого сна, а в некоторых
случаях вызывало удлинение медленного сна [Wyatt et al., 1970]. Авторы полагают,
что лишь серотонинергическая система индуцирует быстрый сон у людей, в то время
как адренергическая система скорее подавляет быстрый сон и участвует в
механизмах поддержания бодрствования, что подтверждается опытами и других
исследователей [Bowers, Kupfer, 1971].
Бодрствование, по мнению Jouvet [1968], поддерживается
благодаря активации метаболизма норадреналина в нейронах передних отделов
ретикулярной формации варолиева моста, а также в группе нейронов среднего
мозга. С этим связано тоническое облегчающее влияние на восходящую активирующую
ретикулярную систему и поддержание корковой активности. Не исключается при этом
включение и другой катехоламинергической системы, связанной с активностью
дофамина - нигроотриарной системы, связывающей Substantia nigra с полосатыми
телами и, по-видимому, выполняющей динамогенную роль в экстрапирамидных
механизмах постурального регулирования в состоянии бодрствования [Демин, 1975].
Важная роль моноаминов мозга в регуляции сна подтверждается
фармакологическими исследованиями с использованием "специфических"
средств. Большинство фармакологических агентов, способных оказывать влияние на
церебральные моноамины, неизбежно будет вмешиваться и в сон [Jouvet, 1972].
Используемые в настоящее время фармакологические препараты снотворного
действия оказывают влияние на фазу быстрого сна, чаще всего вызывая ее
подавление. Этот эффект особенно выражен под влиянием препаратов барбитуровой
кислоты. В связи со сложностью переплетений нейромедиаторных влияний при
возникновении быстрого сна Жуве [1972] подчеркивает деликатность триггерной
организации этой фазы сна, в развитии которой необходимы три ключа: серотонин,
ацетилхолин и норадреналин. Допускается, что в индукции быстрого сна могут
участвовать продукты катаболизма серотонина, в частности 5-оксииндолуксусная
кислота, триптофол. Развитие фаз цикла бодрствование-сон связывается с
относительными концентрациями серотонина и норадреналина в структурах головного
мозга при освобождении в соответствующих механизмах (Torda, 1969]. Высказывается
мнение, что для возникновения быстрого сна необходим тонкий баланс и
определенный уровень между аминами [Tabushi, Himwich, 1969]. Отсюда и
проистекает особая чувствительность быстрого сна к различным фармакологическими
и другим факторам. Наблюдающийся феномен "отдачи" после отмены ряда
подавляющих фазу быстрого сна фармакологических препаратом и после тотальной
или избирательной депривации сна, по мнению Oswald [1969], может отражать
повышенный синтез протеиина в мозгу. Синтез рассматривается как создание нового
нейронного механизма или для замещения того, который был до этого создан для
противодействия или для выработки к препарату толерантности, развитию которой
препятствуют ингибиторы синтеза протеина [Сох et al., 1968]. Ряд авторов
[Jouvet, 1964; Dement, 1965] считают, что во время бодрствования и медленного
сна происходит накопление в организме химических веществ, нейтрализация которых
совершается во время быстрого сна. Подводя итоги имеющимся в литературе данным,
следует подчеркнуть отсутствие единого мнения о нейрофизиологических и
нейрохимических механизмах регуляции и поддержания различных проявлений
бодрствования и сна, в частности его фаз и стадий и их циклической организации.
Достигнутые успехи в результате экспериментальных исследований на животных,
свидетельствуют о сложной иерархии взаимосвязей нейрофизиологических и
нейрохимических систем мозга в регуляции цикла бодрствование-сон и многообразии
его проявлений. Опубликованные данные показывают большую сложность
нейрохимической регуляции сна и бодрствования и связанные с этим значительные
расхождения в результатах исследования различных фармакологических веществ на
сон животных и в особенности человека. Изучение механизмов сна человека
является весьма актуальным и представляет несомненный теоретический и
практический интерес, так как дальнейшее их углубление может способствовать
целенаправленной регуляции столь распространенных агрипнических расстройств. В
заключение необходимо отметить, что этим не исчерпывается обзор имеющихся в
литературе данных по вопросам химии и фармакологии сна.
Заключение
Поскольку проблемы сна находятся в той области научных исследований,
каждое новое открытие в которой является не только ответом на поставленную
исследователем задачу, но и ставит все новые вопросы и открывает новые
направления для дальнейших исследований, дать более или менее подробный отчет
на эту тему в короткой работе не представляется возможным. Именно поэтому в
настоящей работе освещены лишь те представления о механизмах сна, которые,
выдержав проверку временем и многократное экспериментальное подтверждение,
стали уже классическими. Но, несмотря на это, необходимо отметить, что за
последние десятилетия в изучении природы и механизмов сна был сделан прорыв,
причем не только физиологами и психологами, но и представителями других,
относительно новых смежных наук: кибернетики, квантовой психологии,
хронобиологии и др.
Так, например, в настоящее время, не подлежит ни какому сомнению важная
роль в механизме регуляции сна и бодрствования, открытых хронобиологией
биоритмов - суточных ритмов. В ходе исследований было установлено, что человек
не рождается с суточным ритмом, а постепенно его заучивает. Так, например, у
новорожденных отмечаются часовые ритмы: половину часового цикла ребенок
проводит неподвижно, а половину в движении. Постепенно ритм перестраивается, и
период относительной неподвижности составляет восемь часов и приходится на
ночь. Этот цикл был назван циркадным (от лат. Cirka dien - вокруг дня). Он
характеризуется закономерной сменой интенсивности физиологических обменных
процессов, совершаемых в организме.
Дальнейшие исследования показали, что циркадный ритм является строго
индивидуальным и отличается большой устойчивостью. У ряда людей вообще не
проявляется приспособление к меняющимся условиям жизни, у других же он Может
быть сдвинут путем систематической тренировки.
Множество интереснейших открытий, раскрывающих “тайны” сна, было сделано
учеными при изучении гипнотических состояний человека, патологических форм сна,
состояния пассивного бодрствования. Не меньший интерес представляют, сделанные
за последние годы открытия при изучении природы и физиологии сновидений. А в
самое последнее время ученые обратили наконец-то свое внимание на издавна
интересующий человека, но относимый наукой в сферу проблем, если не
эзотерических, то, по крайней мере, мета психологических, феномен так
называемого “осознанного сновидения”. И хотя не все из последних открытий в
области сна можно считать фундаментальными, многие из них противоречивы, а
некоторые даже и сомнительны, но, тем не менее, современные направления научных
исследований и темпы научно-технического прогресса дают возможность
предположить, что дальнейшее изучение механизмов сна, в течение довольно
продолжительного времени, будет весьма плодотворным, и приведет еще не к одному
крупному открытию в области физиологии и психологии.
Литература
1)
Вейн А.
Бодрствование и сон. М., “Наука”, 1970.
2)
Каструбин Ю. Ключ
к тайнам мозга. М., “”, 1995.
3)
Физиология
человека. Под ред. Г. И. Косицкого, - М.,: Медицина, 1985