Пространственная дифференциация и направление микроэволюционных процессов в формирующихся периферийных популяциях Cepaea nemoralis
ВВЕДЕНИЕ
Для некоторых достаточно хорошо изученных
фенетически видов можно выявить центры видового многообразия тех участков
ареала, где представлено большинство изученных фенов. В работе Н.И. Вавилова о
центрах происхождения культурных растений выделялись центры внутривидового
многообразия. Такие центры определяли район наиболее перспективных поисков
новых форм для селекции. Исходя из общих эволюционно - генетических
представлений, можно нарисовать гипотетическую схему разнообразия вида в
пределах ареала [1].
Как правило, центр разнообразия фенов
соответствует зоне оптимума вида. Эта зона характеризуется сравнительно
стабильными по численности и значительными по размеру популяциями. Здесь
поддерживается и проявляется разнообразие фенов, характерное для всего вида. По
мере удаления от центра набор оптимальных условий существования уменьшается.
Популяции на периферии видового ареала обычно более мелкие, подвержены более
значительным колебаниям численности, в большей степени изолированы друг от
друга; тут должны резко колебаться векторы отбора. В результате у таких
периферических популяций будет обедняться фенофонд. У каждой периферической
популяции фенофонд будет беднее, чем у любой из центральной популяции. В таких
краевых популяциях чаще будут выщепляться редкие фены (скрытые в гетерозиготном
состоянии в зоне оптимума). Поэтому сумма фенофондов периферийных популяций
может оказаться разнообразнее, чем у центральных популяций. Однако каждая из
краевых популяций в отдельности должна быть менее разнообразной фенетически,
чем центральная.
Если описанная схема верна, то можно сделать
такой вывод: когда исследователь встречает в природе сравнительно малочисленную
популяцию с резкими колебаниями численности, где фенетическое разнообразие
сравнительно мало, но встречаются редкие фены (то по отношению к другим
популяциям), можно предполагать, что перед ним периферийная популяция. Именно
такими (с пониженной гетерозиготностью но хорошо учитываемым фенам окраски
раковины) оказались изолированные колонии виноградной улитки на самой северной
границе ареала вида - в Шотландии. Некоторые из популяций были вообще
мономорфными по отдельным фенам [2].
Актуальность исследования. Изучаемые нами
группировки Cepaea
nemoralis относятся,
вероятно, к описанным выше периферийным малочисленным популяциям, поэтому
изучение фенетической структуры их представляет определенный интерес.
Цель работы: изучить пространственную
дифференциацию и направление микроэволюционных процессов в формирующихся
периферийных популяциях Cepaea
nemoralis.
Для решения данной цели поставлены следующие
задачи:
изучить изменчивость рисуночно-окрасочных форм Cepaea
nemoralis в периферийных
популяциях Бреста;
изучить изменчивость фенетической структуры
периферийных популяций в пространстве и во времени;
отработать методику культивирования Cepaea
nemoralis;
выявить реакции вида на действие солей тяжелых
металлов (азотистого свинца) в лабораторных условиях;
изучить особенности фенетической структуры вида
в различных экотопах;
определить направление адаптаций Cepaea
nemoralis к неблагоприятным
факторам и новым условиям существования.
Апробация результатов работы. Материалы работы
были представлены и доложены:
на заседаниях научно-исследовательской группы
«Фен» (2008, 2009, 2010 гг.);
на студенческих научных конференциях биологического
факультета «НИРС» (2008, 2009, 2010 гг.);
на межуниверситетской студенческой
научно-практической конференции «Природа, человек и экология» 30 апреля 2010
г., Брест;
на региональной научно-практической конференции
студентов 25 марта 2010 г., Брест.
Опубликованность результатов работы - 2 статьи:
. Синявская, А.С. Фенетическая структура Cepaea
nemoralis в формирующемся
ареале / А.С. Синявская // Сб. материалов студ. научн. конф. НИРС 2009, Брест.
гос. ун-т имени А.С. Пушкина : под общ. ред. Л.Н. Усачёвой. - Брест : БрГУ,
2009. - Ч. 2. - С. 24-25.
. Синявская, А.С. Изменчивость устья раковины Cepaea
nemoralis в формирующемся
ареале / А.С. Синявская // Состояние природной среды Полесья и сопредельных
территорий : материалы регион. научн.-практ. конф. студентов, Брест, 25 марта
2010 / Брест. гос. ун- т имени А.С. Пушкина; под общ. ред. Л.Н. Усачёвой. -
Брест : БрГУ, 2010 - С. 60-61.
1.
ОБЗОР ЛИТЕРАТУРНЫХ ИСТОЧНИКОВ
1.1
Современный ареал семейства Helicidae
Особи любого вида распределяются в пространстве
крайне неравномерно: они встречаются только в тех местообитаниях, где находят
подходящие для жизни условия. Часть поверхности суши, в пределах которой
распространены и проходят полный цикл своего развития особи данного вида (рода,
семейства или определенного типа сообщества), называется ареалом.
В пределах ареала особи того или иного вида
обычно распределяются по группам разной величины и занимают те его части, где
условия в наибольшей степени отвечают требованиям данного вида. Небольшие по
численности группы особей одного вида, как правило, существуют
непродолжительное время, после чего они обычно погибают в результате резких
изменений почвенно-климатических условий, нападения вредителей, возникновения
болезней и ряда других причин. Большие группы особей, которые приспособились к
условиям конкретного участка ареала, наоборот, сохраняются длительное время (на
протяжении бесчисленно многих поколений). Увеличиваясь в численности при
благоприятных условиях и уменьшаясь при неблагоприятных, они имеют перспективу
к длительному существованию на данной территории. Такие группировки особей
одного вида, в течение длительного времени населяющие определенную часть
ареала, свободно скрещивающиеся и относительно обособленные от других
совокупностей того же вида, называются популяцией [3].
Широко известные Helicidae
- яркая группа, имеющая компактный ареал, четко очерченная анатомически и
весьма разнообразна конхологически. При значительном разнообразии раковин Helicidae
отличаются большим постоянством черт внутреннего строения. Даже столь
неустойчивый в ряде других семейств признак, как число слизистых желез, здесь
жестко закреплен - слизистых желез всегда две, ветвящихся в разной степени - от
вообще нерасщепленных до многоветвистых, кустовидных.
Семейство разделяется на два подсемейства - Ariantinae
и Helicinae. Распространение
семейства в средиземноморье, причем ареал Ariantinae
укладывается целиком внутри ареала Helicinae,
занимая преимущественно Южную Европу и не заходя на Пиренейский полуостров, за
исключением рода Elona.
Но представители последнего встречаются только на склонах Пиренеев и не
спускаются ниже предгорий. Наибольшее разнообразие Helicinae
наблюдается в двух регионах: в-первых, как и Ariantinae,
в Южной и Юго-Восточной Европе и, во-вторых - на территории горной системы
Северо-Западной Африки: хребты Большой и Средний Атлас и Антиатлас, причем
наиболее своеобразные конхологические типы свойственны именно этим районам.
Распространение и местообитание Cepaea
nemoralis - это
преимущественно северные и средние районы Европы. Вид заходит также в Пиренеи и
занимает большую часть территории Великобритании. Населяет преимущественно
умеренно увлажняемые разреженные леса, опушки, встречается также на влажных
лугах, в полосе дюн и в кустарниковых зарослях. В ископаемом состоянии
обнаруживается, начиная с плиоцена [4].
Как выяснили A.
Cain, P.
Sheppard, что в масштабе
всего ареала Cepaea
nemoralis наблюдается
заметное увеличение доли желтых форм с севера на юг в районе исследования [5].
В распределении разных морф есть так называемый эффект ареала, несколько близко
расположенных группировок оказываются окрашенными одинаково и соседствуют с
резко отличной от них другой группой особей. На участке 5,4 ×
9
км 103 группировки улиток (5765 экз.) содержали только 11 экз. с 5 полосами. В
2-х км от этого участка все моллюски без исключения оказались пятиполосыми [6].
Эффект ареала был объяснен отбором климатических условий. Оказалось также, что
частоты полосатых раковин могут сохраняться, с одной стороны, крайне долго
(несколько тысяч лет, как показывают раскопки), а с другой стороны, могут
существенно меняться за десятки лет. Правда, в последнем случае изменения
частот морф совпало с изменением характера растительности. Обнаружились и
физиологические различия между морфами: желтые и коричневые в теплое и сухое
лето откладывают яиц достоверно меньше, чем розовые; оказалось также, что
улитки с более темными раковинами (в том числе многополосые) более
холодоустойчивы, разные морфы неодинаково выживают в разные сезоны, а также в
зависимости от плотности группы. Сейчас ясно, что нет никакого экологического
фактора, ответственного за распределение частот аллелей в пространстве ареала -
всегда действует, по меньшей мере, несколько разнонаправленных комбинаций
селективных сил в разных местообитаниях. В это же время, по-видимому, в
некоторых случаях (как, например, на насыпи вдоль дороги, вновь заселенной
улитками после сильного наводнения) частота морф может определяться немногими
выжившими, т.е. может проявиться эффект основателя.
На короткое время на небольших участках частоты
аллелей в природных группировках могут определяться случайными причинами
(эффектом основателя, дрейфом генов), но длительное поддержание определенной
частоты аллеля (равно как и поддержание определенной частоты на большом
пространстве) всегда связано с действием отбора [7].
1.2
Фенетическая изменчивость брюхоногих моллюсков
Наиболее изученными в фенетическом плане из
брюхоногих моллюсков являются: виноградная улитка, садовая улитка, лужанка
живородящая Bradybaena
fruticum, кустарниковая
улитка
Широко распространенный вид - лужанка
живородящая. Обитает практически во всех природных водоемах Бреста.
Цвет раковины зеленовато - коричневый с темными
продольными полосами на последнем обороте. Встречаются особи с отсутствием
полосы, а также с наличием 1 - 4 полос в водоемах Бреста [8].
При изучении данного вида использовался
морфологический подход. Учитывались четыре рисуночных варианта раковины:
бесполосные, одно-, двух-, и трехполосые. Для фенетического анализа сборы материала
производились в трех пунктах, расположенных на большом расстоянии друг от
друга. Пункты сбора: «Брестская крепость», «Суворовский мост», «Выстичи».
Сравнение выборок показало, что в выборке
«Суворовский мост» статистически достоверно меньше особей, имеющих три темные
полосы, по сравнению с двумя другими выборками, что, вероятно указывает на
более благоприятные условия обитания вида в этом районе города.
Увеличение частоты трехполосых форм, а,
следовательно, и повышение степени меланизации раковины моллюсков в выборках
«Брестская крепость» и «Выстичи», на наш взгляд можно объяснить более
интенсивным потоком автомобильного транспорта, т.к. выборки были собраны в
водоемах, расположенных в непосредственной близости от автотрасс.
Другой широко распространенный в
Бугско-Полесском регионе вид - Виноградная улитка. Это один из самых крупных
наземных моллюсков фауны Республики Беларусь. Высота раковины до 45 мм, ширина
- до 45 мм. Цвет раковины: желтый, белый, коричневый и розовый.
Как отмечает Е.П. Климец в Бресте и его
окрестностях отмечены четыре варианта окрасочных форм. Выборки собраны в 3-х
пунктах Бреста и его окрестностей. Пункты сбора: «Газоаппарат», «Брестская
крепость», «Клейники». Выборки «Газоаппарат» и «Клейники» характеризуются
преобладанием коричневых форм. В выборке «Брестская крепость» максимальное
число особей с розовой раковиной и достаточно высокая частота встречаемости
моллюсков с белой раковиной. Статистическое сравнение выборок по частотам
встречаемости всех фенов показало, что выборка «Брестская крепость» резко
отличается по изучаемым параметрам от двух других и характеризуется наличием
большого количества особей со светлой окраской раковины. Вероятно, эти различия
связаны с относительно низким потоком автомобильного транспорта у места сбора
изучаемого вида в данном пункте [8].
Имеются данные Хохуткина, Лазаревой, Макеевой,
Зейферта указывающие на связь частот встречаемости отдельных морф в популяциях
с экологическими факторами среды - уровнем влажности и степенью затененности
местообитания, преобладанием низких или высоких температур в отдельные периоды
года [9]. А.Ф. Иваньковой изучалась связь отдельных морф кустарниковой улитки с
экологическими факторами среды. У кустарниковой улитки встречаются 4 морфы:
розовые с полосой, розовые без полосы, светлые с полосой и светлые без полосы.
Выборка составила 72 - 300 экземпляров с трех участков, пространственно
удаленных: д. Каменюки, Белое озеро, пригород Бреста. В исследовании условия
обитания популяций д. Каменюки и Белое озеро в значительной мере сходны по
характеру растительности покрова и уровню влажности, а условия популяции
пригород Бреста отличаются (более открытое пространство, меньше влажность).
Представляет интерес сравнение популяций по степени фенотипического
разнообразия, особенно в связи с тем, что установление степени дифференциации
пространственно изолированных популяций может способствовать выбору критериев
для выявления популяций и на сплошных участках ареала вида [9].
Bradybaena
fruticum изучалась И.М.
Хохуткиным и Л.И. Лазаревой. Авторы на выборке 15485 особей изучали
распределение полосатых и бесполосых морф в разных биотопах.
По соотношению частот фенотипов наблюдается
биотопическая изменчивость Bradybaena
fruticum обитающих на
болотах и в заболоченных канавах преобладает бесполосая морфа - от 83,3 % - 100
%.
В пойменных лесах и горных лесах Кавказа
бесполосая морфа - от 75,2 % до 82,1 %, а на высыхающих болотах и в некоторых
смешанных лесах - от 45,8 % до 50 %. По берегам водоемов, в более засушливых
лесных участках она составляет от 18 % - до 35 %. Моллюски разных видов,
обитающие на открытых склонах имеют больший процент раковин со слабовыраженным
спиральными полосами (47,9 %), чем моллюски обитающие под пологом деревьев
(13,8 %) [10].
1.3
Полиморфизм Cepaea nemoralis
Брюхоногие моллюски типичные полиморфные виды.
Определением понятия «полиморфизм», выяснением роли и механизмов, регулирующих
его, а также классификацией занимались Lewontin,
Оно, Майр, Грант, Салерно [11], [12], [13], [14], [15]. Разные авторы
вкладывают в понятие «полиморфизм» несколько разное содержание. Наиболее часто
определение полиморфизма дается по Форду, согласно которому, полиморфизм - «это
одновременное существование в одной и той же местности двух или нескольких
дискретных форм одного и того же вида в таких отношениях, что самая редкая из
них не может быть сохранена только под давлением мутационного процесса в донном
локусе, а является следствием гетерозиготности по уже имеющимся аллелям» [16].
Данное определение позволяет считать полиморфизм внутрипопуляционным
наследственным явлением, то есть считать полиморфизм генетическим. Генетический
полиморфизм, как считает А.В. Яблоков, представляет особый класс изменений, а
именно существование двух или более и фенотипически, и генотипически
различающихся форм в состоянии равновесия на протяжении ряда поколений в
популяции [7].
Классическое количественное изучение
сбалансированного полиморфизма провели А. Кэйн, Ж. Керри и Ф.М. Шеппард на
улитке Cepaea
nemoralis [17]. У этих
улиток раковина бывает желтая (если внутри находится живая улитка, раковина
выглядит зеленой), различных оттенков коричневого (в том числе светло-палевая),
розовая, оранжевая или красная. Устье раковины может быть темно- коричневым,
розовым или белым, а на самой раковине может быть до пяти темных полос, идущих
по ее контурам. Как цвет раковины, так и характер ее полос детерминированы
генетически. Окраска определяется множественными аллелями, причем коричневая
окраска доминирует над розовой, а обе они доминируют над желтой. Полосатость -
рецессивный признак.
Улиток поедают дрозды, которые переносят их на
ближайший камень и, используя его в качестве «наковальни», разбивают на нем
раковину, чтобы добраться до улитки. Изучая соотношение раковин разных типов
вокруг «наковален» и в самих местообитаниях улиток, А. Кэйн, Ж. Керри и Ф. М.
Шеппард показали, что в популяции действуют селективные факторы. В тех местах,
где фон был довольно однородным, например, на траве и на лесной подстилке,
селективным преимуществом обладали желтые и коричневые раковины без полос, так
как вблизи наковален таких раковин было меньше. Более темные полосатые раковины
имели селективное преимущество там, где фон был пестрым, как, например, на
пастбищах с грубыми травами и в живых изгородях. На любых участках дрозды
больше всего истребляют те формы, которые хорошо заметны. Обширная популяция
полиморфных улиток может занимать несколько участков, различающихся по фону.
Кроме того, характер и окраска фона могут изменяться со сменой времени года.
Хотя выедание хорошо заметных форм происходит непрерывно, ни одна форма не
обладает абсолютным селективным преимуществом; поэтому доля каждой формы в
популяции из года в год остается довольно постоянной [17].
А. Кэйном и Ж. Керри было также обнаружено, что
желтые формы (у которых сама улитка имеет зеленоватый оттенок) меньше поедаются
на зеленом фоне, а на коричневом фоне имеют преимущество розовые и коричневые
особи. Наблюдения показали, что ранней весной, когда земля в лесу коричневая,
дрозды поедают больше желтых форм (43 %), чем поздней весной, когда земля
зеленого цвета (14 %). Это свидетельствует о существовании естественного
отбора, где одним из факторов отбора были дрозды. Более того, существование
отбора хорошо объясняет высокую частоту желто - полосатых раковин в кустарнике
и большую приспособленность в лесу коричневых и розовых форм без полос [18].
Если бы отбор был обусловлен исключительно
хищниками, то не было бы в кустарнике никаких других раковин, кроме желтых и
полосатых, а в других местах коричневых и розовых без полос. Однако в действительности
это не так. Очевидно, что и другие факторы среды участвуют в поддержании
полиморфизма. Наиболее вероятное объяснение заключается в том, что гетерозиготы
имеют преимущества перед гомозиготами благодаря влиянию генов на плодовитость и
жизнеспособность. Такой отбор, если он достаточно эффективен, чтобы поддержать
полиморфизм, в состоянии изменить частоту генотипов в той же степени, в какой
это происходит под влиянием хищников. М. Шнеттер действительно обнаружил
различия в жизнеспособности и поведении между разными формами, но пока не
удалось показать, что гетерозиготы имеют преимущество именно по данным генам.
Профессор А. Кэйн и Ж. Керри после интенсивных
исследований колоний улитки Cepaea
nemoralis, обитающих на
открытой холмистой меловой местности в южной Англии, показали, что улитки не
только поедаются хищниками, но существуют и другие силы отбора. Они обнаружили,
что в этих суровых условиях обитания состав колоний иногда не определяется
фоном, на котором живут улитки. Согласно данным работ этих авторов, а также
ряда других работ, в крайних условиях обитания улитки с желтыми раковинами и
уменьшенным числом полос по некоторым неизвестным физиологическим причинам
имеют большее преимущество, чем в защищенных низменностях, где большое значение
приобретает отбор по внешнему виду. Действие такого отбора особенно заметно во
Франции, где сезонные температуры часто имеют крайнее значение, а влажность
сильно понижена [18].
Баланс численностей разных форм не обязательно
определяется только окраской и характером полосатости. Судя по некоторым
данным, сохранению равновесия при полиморфизме способствует также
физиологические факторы. В некоторых областях с сухой известковой почвой и
светлой поверхностью не всегда доминируют формы с наименее заметными окраской и
характером полос. Полагают, что полиморфизм у Cepaea
определяется генетическим сцеплением особого типа. Гены окраски и полосатости
сцеплены и образуют суперген, который действует и наследуется как одна
генетическая единица. Входящие в него гены определяю признаки, обладающие
такими селективными преимуществами, благодаря которым они сохраняются в
популяции. Именно разнообразие аллельных форм этих генов, охраняемое благодаря
широко распространенной гетерозиготности, составляет основу полиморфизма. К этому
добавляется сцепление генов, детерминирующих некоторые физиологические функции,
что, как полагают, тоже способствует поддержанию сбалансированного
полиморфизма. Наличие в одной и той же популяции нескольких особенных форм,
доля которых слишком велика, чтобы их можно было отнести за счет повторных
мутаций, называют генетическим полиморфизмом [17].
1.4 Реакция брюхоногих
моллюсков на изменение условий среды
Анализ литературных источников, показал, что
работ по изучению влияния химических веществ на фенетическую структуру и
физиологические процессы брюхоногих моллюсков относительно мало. Б.А. Флеров
[19] установил, что под действием хлорорганических соединений нарушается
синоптическая передача в возбудимых структурах организмов. Г.А. Виноградов и
П.А Гдовский [20] показали, что хлорофос в концентрациях 0,1 1 мг/л влияет на
водно-солевой обмен у прудовиков, а Меремiнський
А.Й., выявил подавление мышечной активности у ряда легочных моллюсков [21].
А.П. Стадниченко с соавторами изучили
токсичность хлорофоса в разных концентрациях на пресноводных моллюсков. Авторы
отмечают быстрые поведенческие, патоморфологические и патофизиологические
реакции на интоксикацию хлорофосом у двух видов пресноводных моллюсков -
катушки пурпурной и шаровки Бургинье, ранее в этом отношении не
исследовавшихся.
В опытах использовано 932 экз. животных,
собранных в 1982 г. в ряде водоемов Житомирской области (Украинская ССР):
катушки из небольшого заболоченного водоема (г. Житомир) и из пруда (с.
Смоковка) и шаровки из р. Каменки (Богуния - околица г. Житомира).
При постановке токсикологического эксперимента
животных помещали по 10 экз. в растворы метилового хлорофоса (диптерекс 91%-ной
активности) концентрацией 0,001, 0,01, 0,05, 0,1, 0,2, 0,3, 0,4, 0,5, 1, 10,
100, 200, 300, 400, 500, 600, 1000, 2000, 3000, 4000, 5000 и 10000 мг/л.
Растворы токсиканта заменяли через 24 часа. Экспозиция опытов - 48 часов.
Наблюдения снимались через 10, 30 мин. и через 1, 2, 4, 6, 24 и 48 часов.
Токсическое действие хлорофоса на моллюсков оценивали по изменению их
двигательной активности, наличию у них некоторых из них патоморфологических и
патофизиологических сдвигов и по общепринятым токсикологическим показателям - LC0,
LC50, LC100,
которые определялись по методике, описанной В.А. Алексеевым [22].
Экспериментальные условия создавали повышением
температуры до 30° и обсыханием (моллюсков в течение 7 суток выдержали в
кюветах с влажным песком).
В слабых растворах хлорофоса (0,001 мг/л)
исследованные виды хорошо противостоят воздействию токсиканта. Об этом свидетельствуют
обычная двигательная активность моллюсков, отсутствие у них явных
патоморфологических и патофизиологических нарушений, а также отдаленных
последствий воздействия этого вещества на организм подопытных животных. При
концентрации хлорофоса 0,01 мг/л и катушки и шаровки сохраняют двигательную
активность, не предпринимая при этом попыток покинуть отравленную среду в
течение первого часа с момента постановки эксперимента. Более концентрированные
растворы токсиканта (0,05 - 0,1 мг/л) на протяжении первого часа экспозиции
угнетающе воздействуют на двигательную активность катушек. Животные в них
обычно неподвижно лежат на дне сосудов, втянув ногу и голову в полость
раковины.
Часовое пребывание катушек в растворах хлорофоса
концентрацией 0,01 - 0,1 мг/л вызывает, как правило, возрастание двигательной
активности этих животных, проявляющееся отчетливо выраженной реакцией
избегания, которая является весьма обычной формой оборонительного поведения
животных, оказавшихся в неблагоприятных условиях среды. Она обусловлена
наличием нервной связи, соединяющей органы химического чувства моллюсков -
осфрадии с колумеллярной мышцей и мышцами ноги. Катушки устремляются вверх по
стенкам емкостей, к горловинам последних и, покинув затравленную среду,
располагаются кольцом над урезом воды. Чем выше концентрация токсиканта, тем
скорее обнаруживается реакция избегания. Например, у катушек при концентрации
хлорофоса 0,01 - 0,1 мг/л она отмечается по истечении часового пребывания
животных в растворах токсиканта, в то время как в более концентрированных
растворах (0,2 - 0,1 мг/л) выявляется уже впервые 10 мин.
Принудительное содержание катушек в растворах
токсиканта (0,001 - 2,0 мг/л), не позволяющее им покинуть затравленную среду,
приводит с течением времени к ослаблению двигательной активности животных. При
концентрации хлорофоса 0,001 мг/л способность к передвижению у катушек заметно
снижается через 24 часа, при 0,5 мг/л - через 4 часа, а при 2 мг/л - через 2
часа пребывания в растворах токсиканта.
Раствор хлорофоса концентрацией 10 мг/л
оказывает немедленное угнетающее воздействие на двигательную активность
моллюсков, которая не изменяется до окончания опыта.
У шаровок, подвергнутых воздействию хлорофоса,
часто проявляется другая форма оборонительного поведения - плотное смыкание створок
раковины и втягивание внутрь ее сифонов, чем существенно ограничивается
действие гидрокинетического аппарата и сокращается доступ токсиканта к телу
животных. Эта реакция обеспечивается наличием нервной связи, соединяющей
осфрадии с мышцами - замыкателями, притягивающими створки раковин к телу
моллюсков.
Отсутствие ярко выраженной реакции избегания у
шаровок в условиях эксперимента обусловлено, очевидно, тем, что в аквариумах не
было субстрата в виде мягких донных отложений, без которого передвижение двустворчатых
моллюсков невозможно.
Длительное пребывание моллюсков в растворах
хлорофоса сопровождается развитием разлитого отека ноги и головы. Скорость
появления этого симптома отравления прямо пропорциональна концентрации
токсиканта. Например, у катушек при концентрации хлорофоса 0,01 мг/л это
наблюдается через 48 часов, при 0,2 мг/л - через 24 часа, а при 0,4 мг/л -
через 6 часов. У шаровки разлитой отек ноги появляется при концентрации
хлорофоса 200 мг/л через 2 часа. Объем отекших частей тела возрастает при этом
в 2 - 3 раза. Развитие отека сопровождается обычно утратой способности моллюска
реагировать на механические раздражения.
При интоксикации у подопытных животных
наблюдаются расстройства периферического кровообращения. С ними связано
изменение окраски ноги моллюсков, которая у катушек в связи с усиленным
притоком гемолимфы, содержащей гемоглобин, становится темно - розовой. В слабых
растворах токсиканта (0,01 мг/л) нога катушек розовеет через 48 часов, а при
более высокой концентрации (0,3 мг/л) - гораздо быстрее через 24 часа.
Отмечаются случаи «выпотевания» гемолимфы,
свидетельствующие об изменении проницаемости поверхностных кровеносных сосудов
у моллюсков, подвергнутых воздействию хлорофоса. У катушек это наблюдалось при
концентрации токсиканта 2 мг/л после 24 часов экспозиции.
Под воздействием хлорофоса у моллюсков
стимулируется дефекация. Наблюдается быстрое (одномоментное) выделение обильных
экскрементов. Причем при концентрации токсиканта 0,01 мг/л у катушек это
происходит через 24 часа, при 0,3 мг/л - через 4 часа, а при 1 мг/л - через 2
часа от начала эксперимента. У шаровок интенсивное выделение экскрементов
наблюдается при всех концентрациях хлорофоса примерно через одинаковое время -
около 2 часов.
Интоксикация хлорофосом сопровождается у шаровок
обортированием эмбрионов из марсупиальных сумок. При концентрации токсиканта 1
мг/л это происходит через 24 часа, а при 200 мг/л - через 4 часа от начала
эксперимента.
К защитным физиологическим реакциям моллюсков
относится усиленное выделение слизи железистыми клетками покровов тела. Слизь
обволакивает выступающие из раковины части тела этих животных, ограничивая тем
самым в некоторой мере доступ к ним хлорофоса. У катушек эта реакция отмечается
при концентрации его 0,01 мг/л через 24 часа, при 0,05 мг/л - через 6 часов,
при 1 мг/л - через 4 часа и при 10 мг/л - через 30 мин. У шаровок обильное
выделение слизи происходит при содержании моллюсков в течение 24 часов в
растворе токсиканта концентрацией 10 мг/л. В более слабых растворах хлорофоса усиления
выделения слизи в них не наблюдается.
Наблюдения над ходом патологического процесса у
моллюсков, отравленных хлорофосом, свидетельствуют о том, что картина
отравления у шаровки и катушки - моллюсков различного систематического
положения в общем сходная. Это объясняется одинаковым механизмом,
обуславливающим развитие у них патологического процесса. При отравлении
хлорофосом наиболее ранимой функцией является синоптическая передача в
возбудимых структурах организма. Нарушением ее определяется все дальнейшее
течение патологического процесса в организме отравленных особей.
Сравниванием устойчивости шаровок и катушек по
отношению к растворам хлорофоса установлено, что первые из них отличаются более
высокой сопротивляемостью. Об этом судим по значениям основных
токсикологических показателей, которые у шаровок во много раз превышают таковые
у катушек: LC100 у
катушек составляет 10 - 10000, а у шаровок - не менее 10000 мг/л. Необходимо
отметить, что хотя полученные нами значения LC100
для шаровок и одной из исследованных групп катушек (пруд с. Смоковка) очень
велики, они тем не менее в 3 раза ниже этого показателя, приведенного А.И.
Мереминским для прудовика озерного, катушки роговой и катушки окаймленной из
водоемов Западного Полесья [21].
Постановкой токсикологических экспериментов в
экспериментальных условиях среды установлено, что неблагоприятные внешние
воздействия усугубляют вредное влияние хлорофоса на организм моллюсков. В норме
максимальная концентрация хлорофоса, при которой наблюдается 100 %-ная выживаемость
катушек, составляет 0,3 - 0,4 мг/л. Для животных, содержавшихся в условиях
повышенной температуры, этот показатель составляет 0,2 мг/л, а для особей,
подвергшихся обсыханию, - всего лишь 0,01 мг/л. У моллюсков, ослабленных
пребыванием в неблагоприятных условиях среды, при хлорофосной интоксикации
заметно ускоряются появление симптомов отравления. Так, если обычно разлитой
отек ноги и изменение ее окраски (порозовение) у катушек, находящихся в слабом
растворе хлорофоса (0,01 мг/л), наблюдается через 48 часов от начала
эксперимента, то у особей, подвергшихся как воздействию повышенной температуры,
так и условиям обсыхания, эти реакции отмечаются намного раньше - всего через
24 часа. У катушек, помещенных после условий обсыхания в растворы хлорофоса (0,01
- 0,3 мг/л), усиленное выделение экскрементов происходит через 10 мин, в то
время как обычно при указанных концентрациях токсиканта это наблюдается
примерно через 24 часа.
Следует отметить, что устойчивость моллюсков по
отношению к хлорофосу подвержена популяционной изменчивости. Выживаемость
особей различных популяций, приуроченных в своем распространении к биотопам,
характеризующимся неодинаковыми условиями среды, варьирует в очень широких
пределах.
2. ОБЪЕКТ ИССЛЕДОВАНИЯ
Объектом нашего исследования является Cepaea
nemoralis, который в
исследуемом нами регионе в настоящее время формирует свою популяционную
структуру.
У Cepaea
nemoralis раковина со слегка
куполовидным завитком, высота которого превышает устья. Последний оборот
немного менее чем вдвое шире предпоследнего, к устью плавно, но энергично
опущен. Наиболее широкие темные ленты, если они присутствуют, - четвертая и
пятая. В проходящем свете ленты просвечивают немного сильнее фона. Скульптура в
виде тонких неравномерных радиальных морщинок и тончайшей струйчатости,
ориентированной в целом спирально; эта струйчатость видна лишь при значительном
увеличении. Устье широкое, очень косое, верхний край его прямой или образует
очень пологую дугу. Края устья, отворот коллумелярного края и париентальная стенка
имеют темную окраску. Если рассматривать раковину снизу, со стороны пупка, то
можно заметить, что базальный край устья иногда выгнут в виде лука и края
устья, таким образом, не лежат в одной плоскости. Гораздо чаще базальный край
устья плавно и равномерно выгнут или прямой. Прямой край связан с тем, что
базальная часть последнего оборота уплощена. Устья половозрелых особей имеет
обыкновенно в разной степени отогнутые края и снабжено концентрическим
утолщением - губой, расположенной несколько отступя от края устья. Губа
красновато - коричневая. Пупок полностью запаян [4].
Улитки считаются созревшими, когда в основании
раковины сформируется утолщение - «губа».
Сераеа nemoralis обитает в разнообразных
местообитаниях - в лесах, на лугах, в живых изгородях - в Англии и Западной
Европе. В популяциях этой улитки обычно наблюдается полиморфизм по окраске
раковины, которая может быть бурой, розовой или желтой. Окраска раковины
детерминируется рядом множественных аллелей, среди которых наибольшей
доминантностью обладает аллель бурой, а наибольшей рецессивностью - аллель
желтой окраски [18]. Цвет раковины играет важную роль в защитной окраске этих
улиток.
Одно из местообитаний Cepaea nemoralis - густые
буковые леса. Для этих лесов характерна подстилка из красно-бурых опавших
листьев, которая круглый год имеет примерно одинаковый вид. Здесь окраска
улиток с бурыми и розовыми раковинами оказывается покровительственной, и эти
формы здесь обычны. Улитки с желтой раковиной в буковых лесах встречаются
редко, но они обычны на зеленых лугах [18].nemoralis встречается ещё в одном
местообитании в смешанных лиственных лесах. В этих лесах характер фона
изменяется со сменой времен года. Ранней весной подстилка бывает бурой от
опавших листьев, но к лету она становится зеленой. Селективная ценность раковин
разного цвета здесь также изменяется со сменой времен года. Ранней весной бурая
и розовая окраска лучше защищают от уничтожения птицами, а летом это
преимущество переходит к желтым улиткам. В полиморфных популяциях улиток, обитающих
в лиственных лесах, в течение года происходят циклические изменения
относительных частот разных форм [18].
Окраска раковины несёт не только защитные
функции. Имеются данные, свидетельствующие о различиях в физиологической
выносливости между разными по цвету формами. Желтая форма, по-видимому, легче
переносит экстрёмальные температуры, как высокие, так и низкие, чем розовая.
Большая устойчивость желтой формы к колебаниям температуры, конечно, должна
давать ей преимущество в открытых луговых местообитаниях и в областях с жарким
летом. Эта форма, достигает высокой частоты на лугах; частота её повышается по
географической трансекте от северных к южным районам Европы.
К числу других признаков раковины относится
характер полосатости. Раковины бывают бесполосые и полосатые, а число полос
колеблется от 1 до 5. Наличие или отсутствие полос детерминируется геном.
В, тесно сцепленным с геном, детерминирующим
окраску раковины. В локусе В аллель бесполосости доминирует над аллелем
полосатости. Число полос контролируется особым локусом. В результате
рекомбинации образуются всевозможные сочетания окраски и полосатости раковины
[18].
Роль естественного отбора в отношении характера
полосатости менее ясна, чем в отношении окраски раковины. Известно, что у
других животных темные полосы, опоясывающие тело, участвуют в образовании
защитной окраски в полузатененных местообитаниях, где игра света и тени создаёт
разбитые контуры. Возможно, что такую же функцию несут полосы на раковинах у
Cepaea nemoralis. Во всяком случае, в некоторых местообитаниях полосатая
раковина лучше защищает улиток от уничтожения птицами. Возможно также, что
изменчивость по характеру полосатости контролируется дрейфом генов.
Полиморфные формы Cepaea
nemoralis по цвету раковины,
и количеству полос представлены на рисунке 2.1.

Рисунок 2.1 - Полиморфные формы Cepaea
nemoralis (фото автора)
3. МАТЕРИАЛ И МЕТОДИКА
ИССЛЕДОВАНИЯ
Материалом исследования послужили выборки Cepaea
nemoralis в четырех пунктах
г. Бреста и его окрестностях:
-ая выборка - микрорайон Восток. Сбор материала
проводился в зарослях придорожной растительности по ул. Молодогвардейской.
Данная местность характеризуется равнинным рельефом, растительность в виде
небольших кустарников и травянистая. Расселение Cepaea
nemoralis узкой полосой
вдоль автомобильной дороги длиной около 1 км. Было собрано 100 особей.
-ая выборка - Брестская крепость. Сбор материала
проводился в тупике железной дороги на углу проспекта П.М. Машерова и ул.
Ленина. Ценоз, аналогичен первой выборке. Расселение Cepaea
nemoralis наблюдалось по обе
стороны железнодорожного полотна. Было собрано 100 особей с площади 183 ×
20 м.
-я выборка - железнодорожный вокзал. Материал
собран в районе пересечения улиц Орджоникидзе и Комсомольской у пешеходного
моста. Участок близок по форме к треугольнику со сторонами 110; 90; 96 м. На
территории нет построек, в северной и восточной части бетонные стены, местами
покрытые известковым раствором. В западной части, участок ограничен торговыми
павильонами. В южной части имеется небольшой проход к ул. Орджоникидзе. По
территории участка проходит ветка железнодорожного полотна, поэтому пешеходное
движение ограниченно. Всего было собрано 100 экземпляров Cepaea
nemoralis.
-ая выборка - Ковалево. Материал собирался вдоль
полотна железной дороги, растительность травянистая, достаточно хорошо
освещенная. Расселение Cepaea
nemoralis по обе стороны
железной дороги на протяжении нескольких км. Было собрано 100 особей.
Общий объем исследуемого материала составил 400
особей.
Морфометрическое изучение велось по методике
описанной в определителе Е. М. Хейсин «Краткий определитель пресноводной
фауны».
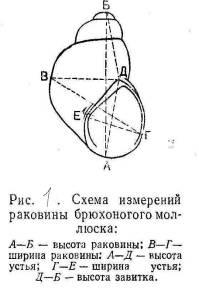
Рисунок 3.1 - Схема измерений раковины
брюхоногого моллюска (по Е.М. Хейсину)
Фенетическое описание (цветовые вариации
раковины, количество меланиновых полос на последнем обороте раковины велось по
методике Климец, Иванькова, Кароза) [25].
Пространственная и временная структура Cepaea
nemoralis изучалась по
методике Е.П. Климец разработанной для колорадского жука [26].
Обработка кормового растения солями азотистого
свинца велась следующим образом: пластинки кормового растения - моркови,
площадью 4,5×2,5 см и толщиной 1мм
выдерживались в концентрациях 1%, 2%, 4%, 8% растворов азотистого свинца в течение
5 мин. Затем эти пластинки служили кормовым растением в контрольных группах.
Статистические методы. При количественной
изменчивости как единого целого использовали среднее арифметическое (Х), лимиты
(lim), среднее
квадратическое отклонение ( ), коэффициент
вариации (V) и нормированное
отклонение (t). Нахождение
данных величин осуществляли по общей методике [27].
), коэффициент
вариации (V) и нормированное
отклонение (t). Нахождение
данных величин осуществляли по общей методике [27].
Для общей характеристики всего материала
необходимо найти такую величину, которая бы минимально отличалась от всех
вариаций. Такой величиной является средняя арифметическая (Х). Она и
представляет собой основной параметр и определяется:

Х - среднее арифметическое, число именное и
выражается в тех же единицах, в которых производилось измерение,
А - условное среднее, та, которая чаще
встречается,
а = Х - А, т.е. отклонение вариаций от условного
среднего,
n = Σf
- сумма всех частот, или объем выборки.
Для характеристики изменчивости
используют специальный параметр стандартное отклонение (или среднее
квадратическое отклонение);  - число именное и выражается в тех
же единицах, в которых производилось измерение. Стандартное отклонение
показывает, насколько в среднем отличается каждая из вариаций от среднего
арифметического. Для вычисления используют более простой метод (условного
среднего) и пользуются следующей формулой. Для того чтобы воспользоваться этой
формулой вычисляют «
- число именное и выражается в тех
же единицах, в которых производилось измерение. Стандартное отклонение
показывает, насколько в среднем отличается каждая из вариаций от среднего
арифметического. Для вычисления используют более простой метод (условного
среднего) и пользуются следующей формулой. Для того чтобы воспользоваться этой
формулой вычисляют «  ».
».
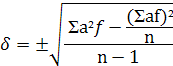
При равенстве средних
арифметических, чем больше величина , тем больше изменчивость. Однако
суждение о степени изменчивости по величине становится невозможным, если
средние не равны и тем более, если надо сравнивать изменчивость разных
признаков, а - величина именованная. Поэтому для
характеристики изменчивости вводят еще одну, относительную или безразмерную
величину - коэффициент вариации или изменчивость, который определяется по
формуле:
V =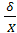
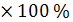
Если рассчитать величину, называемую
ошибкой средней арифметической, обозначаемую буквой m.
Определяется она по формуле:
m = 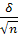
m - число
именованное, она показывает, на сколько ошибаются, когда считают, что среднее
арифметическое выборки соответствует среднее арифметическое выборки
соответствует среднему арифметическому генеральной совокупности. С ее помощью
на основании данных одной выборки можно определить пределы, в которых лежит
среднее арифметическое генеральной совокупности, или, иначе, определить
пределы, в которые будут укладываться среднее арифметические всех выборок,
сколько бы их ни было сделано из одной генеральной совокупности.
Если средние величины для
характеристики вида (генеральной совокупности) выражаются пределами, значит, и
сравнивать нужно через сравнение этих пределов. В математической статистике
предложен специальный метод определения коэффициента достоверности разности
средних:
t = 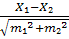
В числителе на первое место ставится большее
арифметическое, чтобы оценивать абсолютное значение разности. Сравнение выборок
по частотам признаков проводилось по формуле Н.А. Плохинского [28]:
χ² =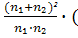 ∑
∑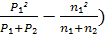
4. ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
4.1 Пространственно -
генетическая структура Cepaea nemoralis в формирующейся периферийной популяции
Изученные нами выборки относятся к формирующейся
периферийной популяции Cepaea
nemoralis. Наблюдения за
появлением Cepaea
nemoralis в Бресте и его
окрестностях показывают, что единичные экземпляры данного вида отмечаются с
60-х годов 20 столетия. Систематическое изучение становления фенетической
структуры вида в Бресте и его окрестностях ведется уже на протяжении 7 - 8 лет.
О. Морозова в своей дипломной работе «Становление популяционной структуры Cepaea
nemoralis в новом ареале»
показывает, что в Бресте формируются небольшие очаги Cepaea
nemoralis. По данным автора
на участке тупика железной дороги (район Брестской крепости) описана
группировка Cepaea
nemoralis насчитывающая 619
особей. Автором дается фенетическая характеристика всех особей, обитающих на
данной территории. В месте обитания по ул. Фортечной г. Бреста было описано 744
экземпляра [29].
Проведенные нами исследования по изучению
цветовых вариаций окраски раковины и изменчивости количества полос на последнем
обороте раковины Cepaea
nemoralis в четырех выборках
г. Бреста позволили выделить следующие вариации:
По цвету раковины: коричневые, розовые,
оранжевые, желтые, серые, бежевые. Естественно, что это чисто визуальная
дифференциация по окраске. Для более четкой дифференцировки цветовых вариантов
необходимо использовать цветовые шкалы, так как многие формы являются
переходными по цвету. Ф. М. Шеппардом установлено, что окраска наследуется
коричневая, которая доминирует над розовой, а коричневая и розовая - над желтой
[18].
По наличию полос на последнем обороте раковины
выявлены: полосатые и бесполосые, а по количеству полос различают: однополосые,
двуполосые, трехполосые, четырехполосые, пятиполосые.
По ширине полосы: тонкая, широкая, очень
широкая, а по степени прозрачности раковины моллюска выделены: прозрачные,
непрозрачные.
Для изучения пространственной структуры нами
учитывались такие параметры как: вариации цвета раковины, количество полос и
количественные показатели: ширина и высота устья.
Проведенные исследования по распределению
окрасочных форм и количества полос на раковине Cepaea
nemoralis в выборках г.
Бреста.
Анализируя частоты встречаемости различных
окрасочных форм в выборках можно отметить, что максимально количество
коричневых форм отмечено в выборке Ковалево (44,0 %), коричневые формы отмечены
и в выборке Железнодорожный вокзал (34,0 %). В выборках Брестская крепость и
Восток моллюски с коричневой окраской не отмечены. Для выборки Брестская
крепость максимальное количество особей отмечено с желтой окраской, а Восток -
оранжевой.
По количеству полос отмечены моллюски со всеми
вариантами - от 0 до 5. Максимальное разнообразие по данному параметру
характерно для выборки Ковалево. Выборка Брестская крепость характеризуется
наличием только пятиполосых и четерехполосых форм.
Судить о преимуществах особей в выборках с
определенной окраской и количеством полос довольно трудно, поэтому мы
попытались провести попарное статистическое сравнение фенофондов выборок
отдельно по окраске (таблица 4.1.1) и количеству полос (таблица 4.1.2).
Таблица 4.1.1 - Попарное сравнение выборок по
окраске раковины (значение χ²)
|
Сравнение
выборок
|
Железнодорожный
вокзал
|
Брестская
крепость
|
Ковалево
|
|
Брестская
крепость
|
145,92
|
|
|
|
Ковалево
|
38,72
|
124,40
|
|
|
Восток
|
63,68
|
126,32
|
66,04
|
Таблица 4.1.2 - Попарное сравнение выборок по
количеству полос (значение χ²)
|
Сравнение
выборок
|
Железнодорожный
вокзал
|
Брестская
крепость
|
Ковалево
|
|
Брестская
крепость
|
145,92
|
|
|
|
Ковалево
|
38,72
|
124,40
|
|
|
Восток
|
63,68
|
126,32
|
66,04
|
Анализ таблицы 4.1.1 показывает, что все выборки
между собой различаются по фенофондам окраски. Различия статистически
достоверны при вероятности 0,001. Однако максимальные различия наблюдаются
между выборками Брестская крепость и Железнодорожный вокзал, а значительно
меньшие - Ковалево и Железнодорожный вокзал.
Анализ таблицы 4.1.2 показывает, что и по
количеству полос все выборки также статистически различаются с вероятностью
0,001. Наиболее значимые различия между выборками Ковалево и Брестская
крепость, а наименьшие - Восток и Ковалево.
Полученные статистические данные показывают, что
каждая выборка Cepaea
nemoralis в г. Бресте
представляет самостоятельную внутривидовую группировку, ранг которой только по
морфологическим параметрам определить еще трудно.
4.2 Фенетическая изменчивость
Cepaea nemoralis во времени
Изучение процесса становления вида в новом
ареале является одной из важнейших задач популяционной биологии [7]. Так как Cepaea
nemoralis новый вид в фауне
РБ, поэтому интерес представляет изучение не только пространственной
дифференциации, но и изменение фенофонда вида во времени, что показывает,
выявление направления микроэволюционных процессов в формирующихся популяциях
этого вида. Нами изучалась изменчивость окраски раковины и количество
меланиновых полос на последнем завитке раковины Cepaea
nemoralis 2002-2009 гг.
Материал исследования составил более 826 особей,
собранных в г. Бресте (перекресток улиц Ленина и Машерова). Материалы за
2002-2005 гг. предоставлены выпускницей О. Мороз, а за 2008-2009 гг собраны
нами.
Анализ материалов 2002-2005гг. показал, что по
окраске раковины выявлены: желтые, розовые и зеленые; по наличию полос:
полосатые и бесполосые; по количеству полос однополосые, двухполосые,
трехполосые, четырехполосые и пятиполосые таблица 4.2.1. Обработка материалов
2008-2009 гг. показала большее фенетическое разнообразие у Cepaea
nemoralis. По окраске
выделено семь вариаций. Кроме уже отмеченных (желтые, розовые и зеленые),
появились новые окрасочные формы: оранжевые, серые, бежевые и коричневые. По
количеству полос в выборках 2008-2009 гг., также как в выборках 2002-2005 гг.,
отмечаются бесполосые и полосатые, а среди полосатых: однополосые, двухполосые,
трехполосые, четырехполосые и пятиполосые.
Таблица 4.2.1 - Частоты изменчивых элементов
окраски и количества меланиновых полос на последнем завитке раковины Cepaea
nemoralis
|
Год
|
Окраска
раковины
|
n
|
Бесполосые
|
Полосатые
|
|
|
|
|
1/п
|
2/п
|
3/п
|
4/п
|
5/п
|
|
|
|
n
|
%
|
n
|
%
|
n
|
%
|
n
|
%
|
n
|
%
|
n
|
%
|
|
2002
|
Желтая
|
34
|
-
|
-
|
16
|
47,1
|
-
|
-
|
17
|
50,0
|
-
|
-
|
1
|
2,9
|
|
Розовая
|
34
|
5
|
14,7
|
15
|
44,1
|
-
|
-
|
14
|
41,2
|
-
|
-
|
-
|
-
|
|
Зеленая
|
2
|
-
|
-
|
1
|
50,0
|
-
|
-
|
1
|
50,0
|
-
|
-
|
-
|
-
|
|
2003
|
Желтая
|
68
|
11
|
16,2
|
20
|
29,4
|
1
|
1,5
|
30
|
44,1
|
2
|
2,9
|
4
|
5,9
|
|
Розовая
|
59
|
6
|
10,2
|
13
|
22,0
|
1
|
1,7
|
29
|
49,2
|
4
|
6,8
|
6
|
10,2
|
|
Зеленая
|
18
|
-
|
-
|
6
|
33,3
|
-
|
-
|
4
|
22,2
|
2
|
11,1
|
6
|
33,3
|
|
2004
|
Желтая
|
98
|
15
|
15,3
|
37
|
37,8
|
1
|
1
|
39
|
39,8
|
3
|
3,1
|
3
|
3,1
|
|
Розовая
|
82
|
5
|
6,1
|
29
|
35,4
|
-
|
-
|
42
|
51,2
|
5
|
6,1
|
1
|
1,2
|
|
Зеленая
|
20
|
-
|
-
|
5
|
25,0
|
-
|
-
|
11
|
55,0
|
4
|
20,0
|
-
|
-
|
|
2005
|
Желтая
|
94
|
16
|
17
|
40
|
42,5
|
-
|
-
|
22
|
23,4
|
2
|
2,1
|
14
|
14,9
|
|
Розовая
|
87
|
17
|
19,5
|
29
|
33,3
|
-
|
-
|
32
|
36,8
|
2
|
2,3
|
7
|
8,1
|
|
Зеленая
|
23
|
2
|
8,7
|
10
|
-
|
-
|
7
|
30,4
|
-
|
-
|
4
|
17,4
|
|
2008
|
Коричневая
|
9
|
1
|
0,93
|
-
|
-
|
3
|
2,8
|
3
|
2,8
|
1
|
0,93
|
1
|
0,93
|
|
Оранжевая
|
68
|
14
|
13,08
|
1
|
0,93
|
22
|
20,6
|
27
|
25,2
|
2
|
1,87
|
2
|
1,87
|
|
Зеленая
|
4
|
-
|
-
|
-
|
-
|
1
|
0,93
|
3
|
2,8
|
-
|
-
|
-
|
-
|
|
Желтая
|
24
|
-
|
-
|
-
|
-
|
4
|
3,74
|
19
|
17,8
|
1
|
0,93
|
-
|
-
|
|
Розовая
|
2
|
-
|
-
|
-
|
-
|
1
|
0,93
|
1
|
0,93
|
1
|
0,93
|
-
|
-
|
|
2009
|
Оранжевая
|
18
|
-
|
-
|
16
|
16,0
|
-
|
-
|
-
|
-
|
-
|
-
|
2
|
2,0
|
|
Бежевая
|
25
|
1
|
1,0
|
24
|
24,0
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Коричневая
|
34
|
21
|
21,0
|
-
|
-
|
-
|
-
|
-
|
-
|
4
|
4,0
|
9
|
9,0
|
|
Серая
|
1
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
1
|
1,0
|
|
Розовая
|
6
|
-
|
-
|
-
|
-
|
-
|
-
|
1
|
1,0
|
1
|
1,0
|
4
|
4,0
|
|
Желтая
|
16
|
2
|
2,0
|
10
|
10,0
|
-
|
-
|
1
|
1,0
|
1
|
1,0
|
2
|
2,0
|
Анализ таблицы 4.2.1 показывает, что в выборках
2002-2005 гг. преобладают по окраске желтые формы, полосатые над бесполосыми
формами, среди полосатых наибольший процент однополосых и трехполосых форм. В
выборках 2008-2009 гг. оранжевые и бежевые, а по наличию полос также как в
выборках 2002-2005 гг. - однополосые и трехполосые.
В выборках г. Бресте наблюдается ежегодное
увеличение численности особей при стабильности границ ареала. Из трех форм по
окраске раковины преобладают желтые, а полосатые формы преобладают над
бесполосыми, среди которых наиболее частыми являются трехполосые. Сравнение
фенофонда Cepaea
nemoralis г. Бреста с
фенофондами популяций Западной Европы показывает, что в новом ареале
формируется популяция, характеризующаяся более низким процентным содержанием
меланистических форм, что вероятно обусловлено принципом основателя, а не
действием отбора, так как ареал популяции только начал формироваться.
Данные по хронологической изменчивости частот
встречаемости бесполосых и полосатых форм в период с 2002-2005 гг. и 2008-2009
гг. представлены в таблице 4.2.2.
брюхоногий моллюск культивирование helicidae
Таблица 4.2.2 - Частоты бесполосых и полосатых
моллюсков в выборках г. Бреста за 2002-2009 гг.
|
Год
|
n
|
Фенотип
|
|
|
Бесполосые
|
Полосатые
|
|
|
n
|
%
|
n
|
%
|
|
2002
|
70
|
5
|
7,14
|
65
|
92,86
|
|
2003
|
145
|
17
|
11,72
|
128
|
88,28
|
|
2004
|
200
|
20
|
10,00
|
180
|
90,00
|
|
2005
|
204
|
35
|
17,16
|
169
|
82,84
|
|
2008
|
107
|
15
|
14,02
|
92
|
85,98
|
|
2009
|
100
|
24
|
24,00
|
76
|
76,00
|
Анализ таблицы 4.2.2 показывает, что за
восьмилетний период изучения полосатые формы преобладают над бесполосыми.
Анализ динамики частот встречаемости данных фенов показывает, что идет
постепенное увеличение частот бесполосых форм от 7,14 % до 24,00 % и уменьшение
полосатых от 92,86 % до 76,00 %.
Изменчивость частот фенов по количеству полос в
период с 2002-2005 гг. и 2008-2009 гг. представлена в таблице 4.2.3.
Таблица 4.2.3 - Частоты моллюсков в выборках г.
Бреста по количеству полос
|
Год
|
n
|
Количество
полос
|
|
|
Одно-полосые
|
Дву-полосые
|
Трех-полосые
|
Четырех-полосы
|
Пяти-полосые
|
|
|
n
|
%
|
n
|
%
|
n
|
%
|
n
|
%
|
n
|
%
|
|
2002
|
65
|
32
|
49,23
|
-
|
-
|
32
|
49,23
|
-
|
-
|
1
|
1,54
|
|
2003
|
128
|
39
|
30,47
|
2
|
1,56
|
63
|
49,22
|
8
|
6,25
|
16
|
12,50
|
|
2004
|
180
|
71
|
39,44
|
1
|
0,56
|
92
|
51,11
|
12
|
6,67
|
4
|
2,22
|
|
2005
|
169
|
79
|
46,75
|
-
|
-
|
61
|
36,09
|
4
|
2,37
|
25
|
14,79
|
|
2008
|
92
|
1
|
1,09
|
31
|
33,70
|
53
|
57,61
|
4
|
4,35
|
3
|
3,29
|
|
2009
|
76
|
50
|
65,80
|
-
|
-
|
2
|
2,63
|
6
|
7,89
|
18
|
23,68
|
Анализ данных таблицы 4.2.3 показывает, что в
первый год наблюдений встречались преимущественно однополосые и трехполосые и
незначительное количество пятиполосых форм. В последующие годы увеличивается
фенетическое разнообразие и отмечается практически весь спектр изменчивости от
бесполосых до пятиполосых. Однако четкой закономерности в динамике изменчивости
не наблюдается.
4.3 Изменчивость
метрических параметров Cepaea nemoralis
Нами изучалось изменчивость ширины и высоты
раковины у Cepaea
nemoralis в четырех выборках
г. Бреста. Сборы проводились в следующих пунктах: Железнодорожный вокзал,
Брестская крепость, Ковалево, Восток. Параметры устья брюхоногих моллюсков
являются одним из таксономических признаков, учитываемых при характеристике
вида. Изучение вариабельности размеров устья у Cepaea
nemoralis представляет
определенный интерес, так как у данного вида интенсивно идут микроэволюционные
процессы в связи с расширением ареала с запада на восток. В таблицах 4.3.1,
4.3.2, 4.3.3, 4.3.4 представлены метрические характеристики выборок Cepaea
nemoralis.
Таблица 4.3.1 - Биометрическая характеристика
изменчивости ширины устья (мм.) Cepaea
nemoralis в выборках г.
Бреста за 2009 г.
|
Показатели
Выборка
|
X
|
lim
|
m
|

|
V
|
|
Железнодорожный
вокзал
|
8,24
|
4
- 13
|
0,19
|
1,91
|
23,18
|
|
Брестская
крепость
|
10,00
|
8
- 11
|
0,07
|
0,74
|
7,39
|
|
Ковалево
|
12,05
|
9 - 14
|
0,11
|
1,11
|
9,24
|
|
Восток
|
10,03
|
7 - 12
|
0,08
|
0,90
|
8,46
|
Анализ таблицы 4.3.1 показывает, что наибольшая
ширина устья характерна для выборок Ковалево (Х = 12,05 мм.), минимальная -
Железнодорожный вокзал (Х = 8,24 мм.). Выборка Железнодорожный вокзал
характеризуется максимальным варьированием ширины устья и максимальным
коэффициентом вариации, а наименьшие показатели характерны для выборки
Брестская крепость.
Попарное сравнение выборок по ширине устья по
критерию Стьюдента представлены в таблице 4.3.2. Данные таблицы 4.3.2 отражают,
что максимальные различия по изучаемому параметру характерны для выборок
Железнодорожный вокзал - Ковалево, а минимальные Железнодорожный вокзал -
Восток.
Таблица 4.3.2 - Попарное сравнение выборок по
ширине устья (по критерию Стьюдента)
|
Показатели
Выборка
|
Железнодорожный
вокзал
|
Брестская
крепость
|
Ковалево
|
|
Брестская
крепость
|
8,59
|
17,16
|
15,07
|
|
|
Восток
|
8,07
|
10,25
|
12,54
|
Таблица 4.3.3 - Биометрическая характеристика
изменчивости высоты устья у Cepaea
nemoralis в выборках г.
Бреста за 2009 г.
|
Показатели
Выборка
|
X
|
lim
|
m
|
|
V
|
|
Железнодорожный
вокзал
|
8,80
|
5
- 12
|
0,15
|
1,46
|
16,59
|
|
Брестская
крепость
|
10,17
|
8
- 11
|
0,07
|
0,70
|
6,85
|
|
Ковалево
|
11,30
|
10
- 13
|
0,09
|
0,93
|
8,20
|
|
Восток
|
8,70
|
3
- 14
|
0,10
|
1,50
|
7,10
|
Анализ таблицы 4.3.3 показывает, что наибольшая
высота устья характерна для выборок Ковалево (Х = 11,30 мм.), минимальная -
Восток (Х = 8,70 мм.). Выборка Восток характеризуется максимальным
варьированием высоты устья (от 3 до 14 мм.), а наименьшие показатели характерны
для выборки Брестская крепость.
Попарное сравнение выборок по высоте устья (по
критерию Стьюдента) представлены в таблице 4.3.4. Данные таблицы 4.3.4
отражают, что максимальные различия по изучаемому параметру характерны для
выборок Брестская крепость - Восток, а минимальные Железнодорожный вокзал -
Брестская крепость.
Таблица 4.3.4 - Попарное сравнение выборок по
высоте устья (по критерию Стьюдента)
|
Показатели
Выборка
|
Железнодорожный
вокзал
|
Брестская
крепость
|
Ковалево
|
|
Брестская
крепость
|
8,46
|
|
|
|
Ковалево
|
14,45
|
9,74
|
|
|
Восток
|
53,96
|
131,01
|
110,98
|
Анализ данных по изменчивости ширины и высоты
устья показывают, что максимальные средние параметры характерны для выборки
Ковалево. Остальные выборки по данному параметру отличаются незначительно.
Однако выборка Железнодорожный вокзал характеризуется большей изменчивостью
ширины устья, так как для нее характерны максимальные лимиты и коэффициент
вариации. Выборка Брестская крепость характеризуется максимальной стабильностью
ширины устья, так как для этой выборки характерны минимальные показатели lim
и V. Объяснить это явление можно тем, что выборка Брестская крепость, более
изолирована и, возможно, в ней уже наблюдается процесс стабилизации вида в
новом ареале. Выборка Железнодорожный вокзал, вероятно, еще не стабилизирована
до настоящего времени и в ней идет пополнение новыми особями с постоянным
завозом их на стенках вагонов с западных регионов.
4.4 Культивирование Cepaea
nemoralis в лабораторных условиях
В зимний период с 2008-2009 гг. нами
отрабатывается методика культивирования Cepaea
nemoralis в лабораторных
условиях. Нас заинтересовало, каким образом идет формирование меланинового
рисунка на последнем обороте раковины моллюска в онтогенезе и существует ли
зависимость между степенью меланизации и условиями существования.
Анализ литературных источников и сайтов
интернета показал, что уже опубликовано несколько рекомендаций по содержанию Cepaea
nemoralis.
Однако эти все методики предложены людьми
далекими от науки. Основная цель авторов это просто содержать что-то живое
дома, а не отработать методику культивирования вида.
Изучив особенности биологии и экологии Cepaea
nemoralis в природных
условиях, мы попытались, отработать наиболее простую методику культуры этого
вида и получить потомство в осеннее - зимний период.
Ниже приводится отработанная нами методика
культивирования Cepaea
nemoralis в лабораторных
условиях. Для содержания Cepaea
nemoralis мы использовали
аквариум прямоугольной формы объемом около 15 литров, предварительно тщательно
вымытый и обработанный дизраствором. На дно аквариума помещали грунт, из
прокаленных песка и глины, толщиной 5 см. (рисунок 4.4.1).

Рисунок 4.4.1 - Содержание Cepaea
nemoralis на
глиняно-песчаном грунте (фото автора)
Сверху аквариум закрывали стеклом, оставив
небольшую щель для поступления воздуха. Аквариумы подсвечивались
люминесцентными лампами на протяжении 14 часов. В качестве корма использовали
ветки крапивы, листья белокочанной капусты, корнеплоды моркови. Корм
предварительно ополаскивался отстоянной водопроводной водой и обновлялся
ежедневно. Корм чередовали. Сначала использовали в качестве корма крапиву до
тех пор, пока она вегетировала, потом в качестве корма использовали листья
капусты и, наконец, морковь в виде тонких пластинок.
Для содержания отбирались половозрелые особи Cepaea
nemoralis, собранные на
небольшом участке г. Бреста в тупике железнодорожного полотна недалеко от
Центрального железнодорожного вокзала сборы проводились в начале сентября. В
один аквариум помещали 50 особей.
Наблюдения за жизнедеятельностью особей в
аквариуме проводились ежедневно. Активность питания оценивалось по площади
съеденного корма.
Используя описанную выше методику, мы смогли
поддерживать культуру на протяжении 7 месяцев. Наблюдения за состоянием особи в
аквариуме показали, что активность питания особей на протяжении двух месяцев
была высокая. На третьем месяце активность питания снизилась. На грунте начали
появляться мухи горбатки. Грунт начал покрываться плесенью. Жизнедеятельность
особей снижалась, и наша культура прекратила свое существование к апрелю
месяцу.
Не смотря на неудачную попытку культивирования Cepaea
nemoralis по вышеописанной
методике, все-таки можно сделать некоторые выводы:
грунт, состоящий только из песка и глины, не
подходит для культуры;
грунт следует увлажнять и чистить или
обезвреживать;
предпочтительным кормом является капуста и
морковь, а не крапива.
Наблюдения за онтогенезом цепей показали, что
описанные выше условия не совсем благоприятные для культивирования, очень
быстро в аквариуме появились мухи горбатки. 16 особей, оставленных на зимовку -
погибло. Попытка провести спаривание также не увенчалось успехом т.к.
температура в лаборатории была относительно низкой, особенно в осенний период.
Нами изучен для культивирования и другой грунт, который создан на основе
природных компонентов, основу которого составляет торф (рисунок 4.4.2).
В грунт добавляли небольшое количество мела, а
для увлажнения среды использовали смоченные куртинки мха сфагнума. Изучался и
новый компонент рациона - яблоки. Как и в первом эксперименте, аквариум
освещался люминесцентными лампами на протяжении 8-10 часов.

Рисунок 4.4.2 - Содержание Cepaea
nemoralis на торфяном грунте
(фото автора)
В подготовленный аквариум в сентябре месяце были
посажены моллюски, взятые из природной среды, также в конце лета, как и в
первом эксперименте. Моллюски активно питались, успешно перенесли зимовку и
начали спариваться в мае месяце. После спаривания большинство моллюсков начало
откладывать яйца, которые находились в толще грунта. Улитка наполовину зарылась
в грунт и отложила 50 довольно крупных яиц (диаметром 5мм). Яйца белые, по
форме чем-то напоминают гусиные, но в миниатюре, и с очень плотной известковой
скорлупой. Яйца откладывают моллюски в почву. Сначала скорлупа яиц достаточно
плотная, а на просвет кажутся янтарными. Но со временем содержимое яиц мутнеет,
а скорлупа истончается, отдавая весь свой кальций на построение раковины. Через
65 дней начали вылупливаться моллюски, количество вылупившихся особей составило
- 50. Молодь начала питаться и расти, однако, через месяц начала наблюдаться
постепенная гибель молодых особей. Все полученное в лаборатории потомство
погибло к концу лета.
Данный эксперимент по культивированию показал,
что наличие в грунте торфа, мела и сфагнума снижает появление горбаток, а
предпочтительным кормом являются яблоки, которыми моллюски охотно питались.
Причиной гибели лабораторного потомства,
возможно, явилось все-таки недостаточное количество влаги и недостаток
кислорода, так как аквариум был закрыт стеклом и открывался нерегулярно.
Наш опыт по культивированию моллюсков показал,
что просто содержать моллюсков можно в условиях, описанных во втором
эксперименте, однако, эти условия не обеспечивают поддержания жизнеспособности
культуры. Методика культуры Cepaea
nemoralis должна
дорабатываться.
4.5 Влияние нитрата
свинца на Cepaea nemoralis
Свинец, относится к наиболее известным ядам.
Свинец находится в микроколичествах почти повсеместно. В почвах обычно
содержится от 2 до 200 мг/кг свинца. Свинец, как правило, сопутствует другим
металлам, чаще всего цинку, железу, кадмию и серебру. Свинец токсически
действует на 4 системы органов: кроветворную, нервную, желудочно-кишечную и
почечную.
Нитрат свинца
<http://ru.wikipedia.org/w/index.php?title=%D0%9D%D0%B8%D1%82%D1%80%D0%B0%D1%82_%D1%81%D0%B2%D0%B8%D0%BD%D1%86%D0%B0&action=edit&redlink=1>
Pb(NO3)2 - белое кристаллическое вещество, хорошо растворимое в воде. Вещество
разлагается при разогреве (290°C) с образованием токсичных паров оксидов азота,
а также оксидов свинца. Вещество является сильным окислителем и бурно реагирует
с горючими материалами и восстановителями. Вещество может всасываться в
организм при вдыхании аэрозоля и через рот. Нитрат свинца раздражает глаза,
кожу и дыхательные пути, может оказывать действие на кровь, желудочно-кишечный
тракт, почки, печень и нервную систему приводя к анемии, гипертонии, поражению
почек, поражению печени, судорогам, параличу.
Вещество токсично для гидробионтов,
может вызвать долговременные изменения в водной экосистеме. В пищевой цепи
имеет значение для человека, наблюдается биоаккумуляция, в особенности в
морских и наземных организмах.
В промышленности его используют в
спичечном производстве, крашении и набивке текстиля, окраске рогов и гравировке
[30].
Наши эксперименты проводились на Cepaea nemoralis.
Эксперимент проводился в марте 2010г. Контрольную и экспериментальную группу
составляли перезимовавшие в лабораторных условиях особи, поэтому
экспериментальные и контрольные выборки невелики. Группировка составила 60
особей. Моллюски зимовали в стеклянных банках с грунтом.
Для пробуждения моллюсков после
зимы, обрызгивали их отстоянной водопроводной водой из пульверизатора и
поставили под лампу накаливания. Через один час 40 особей начали активно
двигаться, 20 особей начали активно двигаться через 2 часа и 40 особей
зимовавших в лаборатории оказались мертвыми.
Наблюдения за пробуждением показали,
что первые 40 особей, начав активное движение, заработали радулами, отыскивая
корм. В этой группировке среди 40 особей по окраске раковины были: коричневые,
желтые и оранжевые в соотношении 10:10:20. По количеству полос однополосые и
пятиполосые. Однополосые: коричневые - 10, желтые - 10, оранжевые - 10 особей,
пятиполосые - желтые - 10.
Среди 20 особей, активность которых
наступила позже, были по окраске раковины бежевые, а по наличию полос -
однополосые.
Среди погибших 40 особей, по окраске
были оранжевые, желтые и коричневые; по наличию полос - бесполосые и
однополосые.
Из 60 перезимовавших особей
контрольную группу составили 12, а экспериментальную 48. Схема эксперимента
представлена в таблице 4.5.1. Контрольные и экспериментальные группы
выкармливались пластинками моркови площадью 4,5 × 2,5 см, толщина
0,1 мм.
Морковь является хорошим кормом для Cepaea nemoralis, что было
проверено нами при отработке методике культивирования данного вида. Контрольная
группа выкармливалась пластинками моркови смоченной отстоянной водопроводной
водой. Экспериментальные группировки выкармливались пластинками моркови такого
же размера, но выдержанные на протяжении 10 мин в растворе азотистого свинца в
концентрациях 1 %, 2 %, 4 %, 8 %.
Таблица 4.5.1 - Схема эксперимента
|
Фенотип
|
Контроль
|
Эксперимент
|
|
|
1 %
|
2 %
|
4 %
|
8 %
|
|
Коричневые
Одополосые
|
2
|
2
|
2
|
2
|
2
|
|
Оранжевые
Однополосые
|
2
|
2
|
2
|
2
|
2
|
|
Бежевые
Однополосые
|
4
|
4
|
4
|
4
|
4
|
|
Желтые
Однополосые
|
2
|
2
|
2
|
2
|
2
|
|
Желтые
Пятиполосые
|
2
|
2
|
2
|
2
|
2
|
|
Всего
|
12
|
12
|
12
|
12
|
12
|
Наблюдения за контрольной группировкой показали,
что активность питания довольно высокое, количество съеденного корма было
примерно таким же, как и у особей при культивировании в лабораторных условиях.
Никаких заметных морфофизиологических изменений не наблюдалось, поведение этой
группировки аналогично поведению природной группировки.
Экспериментальная группировка выкармливается
морковью обработанной 1 % раствором нитрата - свинца, активно съела корм, особи
активно ползали по банке и возвращались назад к корму. Было видно активное
движение радулой. Выделение слизи не наблюдалось.
Экспериментальная группировка корм, которой был
обработан, 2 % раствором нитрата-свинца активно уползала от корма, количество
потребляемого корма мало, особи выделяли небольшое количество слизи, которое
концентрировалось в устье. Моллюски втягивали тело в раковину и не выползали.
Экспериментальная группировка, которая питались
морковью обработанной 4 % раствором нитрата-свинца, характеризовалась тем, что
особи активно уползали от корма, количество потребляемого корма также мало,
активно прятали тело в раковину и выделали большое количество слизи.
Экспериментальная группировка, корм которой был
обработан, 8 % раствором нитрата-свинца, проявляла наибольшую реакцию с момента
помещения корма в банку. Моллюски начинали активно двигаться к краю банки,
пытаясь выползти из нее, количество потребляемого корма очень мало. Выделяли
огромное количество слизи, которое заполняло все устье. На второй день вся
экспериментальная группировка погибла.
Подводя итог нашему эксперименту можно сделать
следующий вывод:
нитрат - свинца приводит к изменению
поведенческих реакций Cepaea
nemoralis, прежде всего
усилению двигательной активности от источника содержания свинца;
снижением активности питания, выделением слизи
устьем и снижением жизнеспособности, при концентрации 8 % приводит к летальному
исходу.
ЗАКЛЮЧЕНИЕ
Изучение изменчивости окраски раковины и
количества полос на последнем обороте раковины в природных условиях среды
показало, что за период с 2002 по 2009 гг. в выборках г. Бреста выделено семь
вариантов окраски: серые, бежевые, желтые, розовые, оранжевые, зеленые,
коричневые. По наличию полос - бесполосые и полосатые, а по количеству полос -
от одной до пяти.
Сравнение четырех выборок, собранных в разных
районах города по частоте окрасочных форм и количеству полос, а также по ширине
и высоте устья, показало, что каждая выборка Cepaea
nemoralis, характеризуется
специфическим фенофондом.
Изменение фенофонда во времени позволяет
констатировать, что с момента начала наблюдения за становлением фенетической
структуры вида в г. Бресте, происходит увеличение окрасочных форм и изменение
доминирующих феноформ (от желтой до оранжевой и коричневой). Отмеченные в
выборках Бреста зеленые и розовые формы исследователями в других регионах не
отмечаются. За восьмилетний период наблюдений в выборках г. Бреста произошло
увеличение бесполосых форм с 7,14 % до 24,00 % и одновременно уменьшение
полосатых от 92,86 % до 76,00 %. Четкой закономерности в динамике форм с разным
количеством полос не наблюдается.
Морфометрическое сравнение выборок в
пространстве показывает, что каждая из четырех выборок представляет самостоятельную
внутривидовую группировку, ранг которой на данный момент определить еще трудно.
Динамика фенофондов во времени отражает направление микроэволюционных процессов
и показывает, что в формирующемся новом ареале происходит увеличение бесполосых
форм, но с более темной окраской раковины. Увеличение количества коричневых
форм, возможно, является отражением наличия неблагоприятных факторов в
окружающей среде, а увеличение бесполосых форм подтверждает то, что в
формирующемся ареале для Cepaea
nemoralis более благоприятны
для существования открытые пространства, чем затененные.
Проведенные нами лабораторные исследования
показали, что Cepaea
nemoralis может быть
объектом культивирования, но для продолжительной культуры необходима доработка
методики ее содержания.
Выявление реакции Cepaea
nemoralis на присутствие
солей тяжелых металлов подтверждает, что наземные брюхоногие моллюски могут
использоваться, как виды индикаторы качества среды.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Вавилов,
Н.И. Центры происхождения культурных растений Н.И. Вавилов Тр. по приклад.
ботан., генет. и селекции. - 1926. - Т. 16. - Вып 2. - С. 3-138.
2. Яблоков,
А.В. Фенетика / А.В. Яблоков. - М. : Наука, 1980. - 132 с.
. Парамонов,
А.А. Дарвинизм : учеб. пособие по биол. спец. для студентов пед. ин-тов / А.А.
Парамонов. - М. : «Просвещение», 1978. - 335с.
. Шилейко,
А.А. Наземные моллюски надсемейства Helicoidea
/ А.А. Шилейко // Серия : Фауна СССР. Моллюски.
- Л.
: Наука,
1978. - Т.
3.- Вып.
6. - 384 с.
. Cain,
A.J. Genetics / A.J. Cain, P.M. Sheppard, 1954. - Р.
89-116.
. Cain,
A.J. Area effects in Cepaea on the Larkhill Artillery ranges, Salisbury plain /
A.J. Cain, J. D. Currey // Soc. Zool. London, 45, № 303, 1963. - Р.
1-15.
7. Яблоков,
А.В. Популяционная биология : учебное пособие / А.В. Яблоков. - М. : Высшая
школа, 1987. - 303 с.
. Биотест
: сб. научных трудов ; под ред. Е.П. Климец. - Брест : БрГУ им. А.С. Пушкина,
2003. - 169 с.
. Фенетика
популяций : материалы III
Всесоюзного совещания, Саратов, 7-8 февраля 1985 г. / Саратов. гос. ун-т ; под
ред. Н.И. Ларина [и др.]. - саратов, 1985.- 239 с.
. Физиологическая
и популяционная экология животных / Межвузовский научный сборник, Сарат. ун-т,
1978. - 200 с.
11. Lewontin,
R.C. Interdeme selection cont rolling a polymorphism ih the house mouse / R.C
Lewontin. // Amer. Nat, 1962 - Vol. 96. - P. 65-78.
12. Оно,
С. Генетические механизмы прогрессивной эволюции / С. Оно. - М. : Мир, 1973. -
227 с.
. Майр,
Э. Популяции, виды и эволюция / Э. Майр. - М. : Мир, 1980. - 407 с.
. Грант,
В. Эволюция организмов / В. Грант. - М. : Мир, 1980. - 407с.
. Салерно,
А. Биохимический полиморфизм / А. Салерно // Молекулярные механизмы
генетических процессов. Молекулярная генетика Эволюция и генетическая
инженерия. - М., 1982. - С. 139-147.
16. Ford,
E.B. Polymorphism / Е.В.
Ford Biol. Rev. - London, 1945. - P. 73-88.
17. Грин,
Н. Биология : в 3 т. / Н. Грин, У. Стаут, Д. Тейлор. - М. : Мир, 1993. - 3 т.
. Шеппард,
Ф.М. Естественный отбор и наследственность / Ф.М. Шеппард ; под ред. Д.К.
Беляева. - М. : Просвещение, 1970. - 216 с.
. Флеров,
Б.А. Физиологические механизмы действия токсических веществ и приспособления к
ним водных животных / Б.А Флеров // Гидробиол. журн. - 1977. - Т. 14, № 1. - C.
80-86.
. Виноградов,
Г.А. Исследование действия хлорофоса на водно-солевой обмен пресноводных
брюхоногих моллюсков / Г.А. Виноградов, П.А. Гдовский // Информ. бюлл. ин- та
биол. внутрен. водоем. АН СССР, 1976, № 32. - С. 35-41
. Меремiнський,
А.Й. Вивчення деяких хiмiчних
спролук на молюскiв Planorbis
planorbis
L. в лабораторних та
польових умовах / А.Й. Меремiнський
Ветеринарiя. - 1968. - Вип.
17. - C. 54-59.
. Алексеев,
В.А. Основные принципы сравнительно - токсикологического эксперимента / В. А.
Алексеев // Гидробиол. журн. - 1981. - Т. 17, № 3. - C.
92-100.
. Антропогенные
воздействия на популяции животных : сб. науч. трудов. Волгоград : ВГПИ им. А.С.
Серафимовича, 1986. - 144 с.
. Хейсин,
Е.М. Краткий определитель пресноводной фауны / Е.М. Хейсин. - М. : Гос. уч.-
пед, 1962. - 148 с.
. Климец,
Е.П. Фенетика некоторых видов беспозвоночных юго-западной части Беларуси / Е.П.
Климец, А.Ф. Иванькова, С.Э. Кароза // Веснiк
Брэсцкага унiверсiтэта,
Навукова-тэарэтычны часопic.
- 2009. - С. 71-80.
. Популяционная
фенетика. - М. : Наука, 1997. - 254 с.
. Рокицкий,
П.Ф. Введение в статистическую генетику. - Мн. : Вышэйшая школа, 1974. - 447 с.
. Плохинский,
Н.А. Биометрия / Н.А. Плохинский. - М. : МГУ, 1970. - 270 с.
. Морозова,
О.А. Становление популяционной структуры Cepaea
nemoralis в новом ареале :
дип. Морозова О.А. студ. биол. фак-та : 10.02.05 / О.А. Морозова. - Брест,
2005.
. Институт
промышленной безопасности, охраны труда и социального партнерства -
Санкт-Петербург, 1999. - Режим доступа: http: // www.safework.ru.
- Дата доступа : 25.03.2010.
. Ватти,
К.В. Руководство к практическим занятиям по генетике : учеб. пособие / К.В.
Ватти, М.М. Тихомирова - 2-е изд. - М. : Просвещение, 1979. 189 с.
. Синявская,
А.С. Фенетическая структура Cepaea
nemoralis в формирующемся
ареале / А.С. Синявская // Сб. материалов студ. научн. конф. НИРС 2009, Брест.
гос. ун-т имени А.С. Пушкина : под общ. ред. Л.Н. Усачёвой. - Брест : БрГУ,
2009. - Ч. 2. - С. 24-25.
. Синявская,
А.С. Изменчивость устья раковины Cepaea
nemoralis в формирующемся
ареале А.С. Синявская Состояние природной среды Полесья и сопредельных
территорий : материалы регион. научн. - практ. конф. студентов, Брест, 25 марта
2010 / Брест. гос. ун-т имени А.С. Пушкина; под общ. ред. Л.Н. Усачёвой. -
Брест : БрГУ, 2010. - С. 60-61.