Возбудимость и ее измерение, лабильность
Волгоградская
государственная академия физической культуры
Реферат
На
тему: «Возбудимость и ее измерение, лабильность»
Выполнила Студентка 1го
курса
Группы 201-А Злобина
О.С.
Проверила: Суслова И.В.
Волгоград
2009г.
1. Возбудимость, раздражимость - это
способность живых клеток (от простейших одноклеточных организмов до нервных
клеток человека) воспринимать изменения внешней среды и отвечать на эти
изменения (раздражения) реакцией возбуждения. Возбудимость связана с
существованием в клеточной мембране особых молекулярных структур, обладающих
специфической чувствительностью к действию тех или иных раздражителей:
электрическому току, химическим, механическим, термическим и другим
воздействиям. В нервных и мышечных клетках (волокнах) главный компонент
возбуждения - потенциал действия (ПД), поэтому о Возбудимость указанных клеток
принято судить по пороговой силе тока или пороговому сдвигу потенциала,
достаточным для возникновения ПД. Пороговая сила тока находится в зависимости
от длительности действия раздражающего стимула; в отличие от этого, пороговый
сдвиг потенциала при варьировании длительности стимула не изменяется. В основе
генерации ПД лежит повышение проницаемости клеточной мембраны для ионов натрия.
Соответственно, все агенты, препятствующие повышению натриевой проницаемости
мембраны, ведут к снижению В. На этом механизме основано действие используемых
в медицинской практике местных обезболивающих средств и различных наркотических
веществ.
Лабильность - функциональная
подвижность, скорость протекания элементарных циклов возбуждения в нервной и
мышечной тканях. Понятие «лабильность» введено русским физиологом Н. Е.
Введенским (1886), который считал мерой лабильности наибольшую частоту
раздражения ткани, воспроизводимую ею без преобразования ритма. Лабильность
отражает время, в течение которого ткань восстанавливает работоспособность
после очередного цикла возбуждения. Наибольшей лабильностью отличаются отростки
нервных клеток - аксоны, способные воспроизводить до 500-1000 импульсов в 1 с.;
менее лабильны центральные и периферические места контакта - синапсы. Угнетение
жизнедеятельности тканей и клеток (например, холодом, наркотиками) уменьшает
лабильность, т. к. при этом замедляются процессы восстановления и удлиняется
рефрактерный период. Лабильность - величина непостоянная. Так, в сердце под
влиянием частых раздражений рефракторный период укорачивается, а,
следовательно, возрастает лабильность. Это явление лежит в основе т. н.
усвоения ритма. Учение о лабильности важно для понимания механизмов нервной
деятельности, работы нервных центров и анализаторов, как в норме, так и при
различных болезненных отклонениях.
. Все клеточные мембраны
представляют собой подвижные текучие структуры, поскольку молекулы липидов и
белков не связаны между собой ковалентными связями и способны достаточно быстро
перемещаться в плоскости мембраны. Благодаря этому мембраны могут изменять свою
конфигурацию, т. е. обладают текучестью.
Мембраны - структуры очень
динамичные. Они быстро восстанавливаются после повреждения, а также
растягиваются и сжимаются при клеточных движениях.
Мембраны разных типов клеток
существенно различаются как по химическому составу, так и по относительному
содержанию в них белков, гликопротеинов, липидов, а, следовательно, и по
характеру имеющихся в них рецепторов. Каждый тип клеток, поэтому,
характеризуется индивидуальностью, которая определяется в основном
гликопротеинами. Разветвленные цепи гликопротеинов, выступающие из клеточной
мембраны, участвуют в распознавании факторов внешней среды, а также во взаимном
узнавании родственных клеток. Например, яйцеклетка и сперматозоид узнают друг
друга по гликопротеинам клеточной поверхности, которые подходят друг к другу
как отдельные элементы цельной структуры. Такое взаимное узнавание - необходимый
этап, предшествующий оплодотворению.
Подобное явление наблюдается в
процессе дифференцировки тканей. В этом случае сходные по строению клетки с
помощью распознающих участков плазмалеммы правильно ориентируются относительно
друг друга, обеспечивая тем самым их сцепление и образование тканей. С
распознаванием связана и регуляция транспорта молекул и ионов через мембрану, а
также иммунологический ответ, в котором гликопротеины играют роль антигенов.
Сахара, таким образом, могут функционировать как информационные молекулы
(подобно белкам и нуклеиновым кислотам). В мембранах содержатся также
специфические рецепторы, переносчики электронов, преобразователи энергии,
ферментные белки. Белки участвуют в обеспечении транспорта определенных молекул
внутрь клетки или из нее, осуществляют структурную связь цито - скелета с
клеточными мембранами или же служат в качестве рецепторов для получения и
преобразования химических сигналов из окружающей среды.
Важнейшим свойством
мембраны является также избирательная
проницаемость. Это значит, что молекулы и ионы проходят через нее с различной
скоростью, и чем больше размер молекул, тем меньше скорость прохождения их
через мембрану. Это свойство определяет плазматическую мембрану как
осмотический барьер. Максимальной проникающей способностью обладает вода и
растворенные в ней газы; значительно медленнее проходят сквозь мембрану ионы.
Диффузия воды через мембрану называется осмосом.
Существует несколько механизмов
транспорта веществ через мембрану.
Диффузия - проникновение веществ
через мембрану по градиенту концентрации, осуществляется при участии белков
мембраны, в которых имеются молекулярные поры, либо при участии липидной фазы.
При облегченной диффузии специальные
мембранные белки-переносчики избирательно связываются с тем или иным ионом или
молекулой и переносят их через мембрану по градиенту концентрации.
Активный транспорт сопряжен с
затратами энергии и служит для переноса веществ против их градиента
концентрации. Он осуществляется специальными белками-переносчиками, образующими,
так называемые ионные насосы.
В процессе активного транспорта
ионов в клетку через цитоплазматическую мембрану проникают различные сахара,
нуклеотиды, аминокислоты.
Функции биологических мембран
следующие:
Ø Обеспечивают
транспорт веществ в клетку и из нее, из цитоплазмы в органеллы и наоборот;
Ø Выполняют
роль рецепторов;
Ø Являются
катализаторами;
Ø Участвуют
в преобразовании энергии.
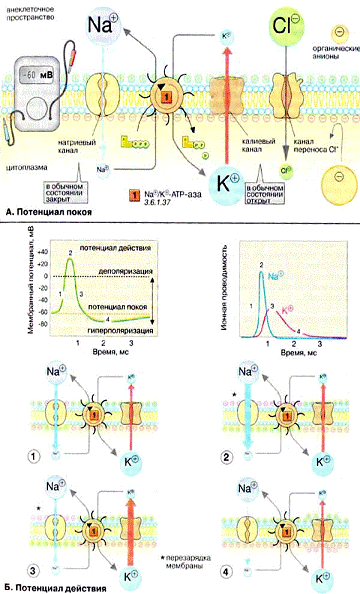
. Потенциал покоя. Мембраны, в том
чикле плазматические, в принципе непроницаемы для заряженных частиц. Правда, в
мембране имеется Na+, K+, АТФ фаза, осуществляющая активный перенос ионов Na из
клетки в обмен на ионы К+. Этот транспорт энергозависим и сопряжен с гидролизом
АТФ. За счет работы Nа+, К+ - насоса поддерживается неравновесное распределение
ионов Na+ и К+ между клеткой и окружающей средой. Поскольку расщепление одной
молекулы АТФ обеспечивает перенос трех ионов Na+ и двух ионов К+, этот
транспорт электрогенен, т. е. цитоплазма клетки заряжена отрицательно по
отношению к внеклеточному пространству.
Электрохимический потенциал.
Содержимое клетки заряжено отрицательно по отношению к внеклеточному
пространству. Основная причина возникновения на мембране электрического потенциала
- существование специфических ионных каналов. Транспорт ионов через каналы
происходит по градиенту концентрации или под действием мембранного потенциала.
В невозбужденной клетке часть К+ каналов находится в открытом состоянии и ионы
К+ постоянно диффундируют из нейрона в окружающую среду (по градиенту
концентрации). Покидая клетку, ионы К+ уносят положительный заряд, что создает
потенциал покоя равный примерно -60 мВ. Из коэффициентов проницаемости
различных ионов видно, что каналы, проницаемые для Na+ и Cl-, преимущественно
закрыты. Ионы фосфата и органические анионы, например белки, практически не
могут проходить через мембраны. С помощью уравнения Нернста можно показать, что
мембранный потенциал нервной клетки в первую очередь определяется ионами К+,
которые вносят основной вклад в проводимость мембраны.
возбудимость раздражимость
лабильность
Потенциал действия. Возбуждение
нервной клетки под действием химического сигнала приводит к возникновению
потенциала действия. Это означает, что потенциал покоя -60 мВ скачком изменяется
на +30 мВ и спустя 1 мс принимает исходное значение. Процесс начинается с
открывания Nа+ канала (1). Ионы Na+ устремляются в клетку (по градиенту
концентрации), что вызывает локальное обращение знака мембранного потенциала
(2). При этом Na+ каналы тотчас закрываются, т. е. поток ионов Na+ в клетку
длится очень короткое время (3). В связи с изменением мембранного потенциала
открываются потенциал - управляемые К+ каналы (2) и ионы К+ устремляются в
обратном направлении, из клетки. В результате мембранный потенциал принимает
первоначальное значение (3), и даже превышает на короткое время потенциал покоя
(4). После этого нервная клетка вновь становится возбудимой.
За один импульс через мембрану
проходит небольшая часть ионов Na+ и К+, и концентрационные градиенты обоих
ионов сохраняются. Поэтому по мере получения клеткой новых импульсов процесс
локального обращения знака мембранного потенциала может повторяться
многократно. Распространение потенциала действия по поверхности нервной клетки
основано на том, что локальное обращение мембранного потенциала стимулирует
открывание соседних потенциал - управляемых ионных каналов, в результате чего
возбуждение распространяется в виде деполяризационной волны на всю клетку.
. Воздействуя раздражителями разной
силы в различные фазы потенциала действия, можно проследить, как изменяется
возбудимость в ходе возбуждения. На рисунке видно, что период локального ответа
характеризуется повышенной возбудимостью; во время фазы деполяризации мембрана
утрачивает возбудимость, которая постепенно восстанавливается в ходе
реполяризации.
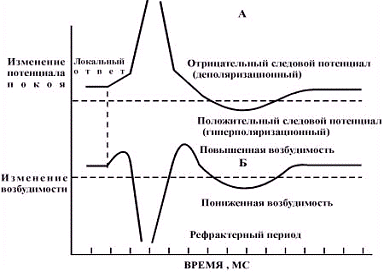
Выделяют период абсолютной рефрактерности,
который в нервных клетках продолжается около 1 мс. и, характеризуется их полной
невозбудимостью. Период абсолютной рефрактерности возникает в результате
практически полной инактивации натриевых каналов и повышения калиевой
проводимости мембраны. По мере реполяризации мембраны происходит реактивация
натриевых каналов и снижается калиевая проводимость. Это период относительной
рефрактерности: потенциал действия может возникнуть только при действии
более сильных раздражителей.
В период отрицательного следового
потенциала фаза относительной рефрактерности сменяется фазой повышенной
возбудимости. В этот период порог раздражения снижен по сравнению с исходным
значением, поскольку мембранный потенциал ближе к критической величине, чем в
состоянии покоя.
Фаза следовой гиперполяризации,
обусловленная остаточным выходом калия из клетки. Напротив, характеризуется
снижением возбудимости, поскольку мембранный потенциал больше, чем в состоянии
покоя, и требуется приложить более сильный раздражитель для его
"смещения" до уровня критической деполяризации.
Таким образом, в динамике
возбудительного процесса изменяется способность клетки реагировать на
раздражители, т.е. возбудимость. Это имеет большое значение, поскольку в момент
наибольшего возбуждения (пика ПД), клетка становится абсолютно невозбудимой,
что защищает ее от гибели и повреждений.
Список литературы
1. ФИЗИОЛОГИЯ ЧЕЛОВЕКА: Учеб. Для ин-тов физ. культ.\ Под
ред. Н. В. Зимкина.-М.:Физкультура и спорт, 1975.
2. ФИЗИОЛОГИЯ ЧЕЛОВЕКА: Учебник для медицинских
институтов.-М.:Медицина, 1984.
. ФИЗИОЛОГИЯ ЧЕЛОВЕКА: В 4-х томах \ Под ред. Р.Шмидта и Г.
Тевса.-М.:Мир, т. 1, 2, 1985; т. 3,4, 1986.