Биохимические изменения в организме при выполнении соревновательных нагрузок
Введение
Обе фазы мышечной деятельности - сокращение и
расслабление - протекают при обязательном использовании энергии, которая
выделяется при гидролизе АТФ:
АТФ + Н20 - АДФ + Н3Р04 + энергия
Однако запасы АТФ в мышечных клетках
незначительны и их достаточно для мышечной работы в течение 1-2 с. Поэтому для
обеспечения более продолжительной мышечной деятельности в мышцах должно
происходить пополнение запасов АТФ. Образование АТФ в мышечных клетках
непосредственно во время физической работы называется ресинтезом АТФ и идет с
потреблением энергии. В зависимости от источника энергии выделяют несколько
путей ресинтеза АТФ.
Для количественной характеристики различных
путей ресинтеза АТФ обычно используются следующие критерии:
а) максимальная мощность, или максимальная
скорость, - это наибольшее количество АТФ, которое может образоваться в единицу
времени за счет данного пути ресинтеза. Измеряется максимальная мощность в
калориях или джоулях, исходя из того, что 1 ммоль АТФ соответствует в
физиологических условиях примерно 12 кал или 50 Дж. Поэтому данный критерий
имеет размерность кал/минкг мышечной ткани или соответственно Дж/мин-кг
мышечной ткани;
б) время развертывания - это минимальное время,
необходимое для выхода ресинтеза АТФ на свою наибольшую скорость, т.е. для
достижения максимальной мощности. Этот критерий измеряется в единицах времени;
в) время сохранения или поддержания максимальной
мощности - это наибольшее время функционирования данного пути ресинтеза АТФ с
максимальной мощностью. Единицы измерения - с, мин, ч;
г) метаболическая емкость - это общее количество
АТФ, которое может образоваться во время мышечной работы за счет данного пути
ресинтеза АТФ.
В зависимости от потребления кислорода пути
ресинтеза делятся на аэробные и анаэробные.
1. Анаэробные пути ресинтеза АТФ
1.1 Креатинфосфатный путь ресинтеза АТФ
(офеатинкиназный, алактатный)
В мышечных клетках всегда имеется креатинфосфат
- соединение, содержащее фосфатную группу, связанную с остатком креатина
макроэнергической связью. Содержание креатинфосфата в мышцах в покое - 15-20
ммоль/кг. Креатинфосфат обладает большим запасом энергии и высоким сродством к
АДФ. Поэтому он легко вступает во взаимодействие с молекулами АДФ,
появляющимися в мышечных клетках при физической работе в результате гидролиза
АТФ. В ходе этой реакции остаток фосфорной кислоты с запасом энергии
переносится с креатинфосфата на молекулу АДФ с образованием креатина и АТФ:
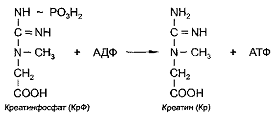
Эта реакция катализируется ферментом
креатинкиназой. В связи с этим данный путь ресинтеза АТФ еще называется
креатинкиназным.
Креатинфосфатная реакция обратима, но ее
равновесие смещено в сторону образования АТФ, и поэтому она начинает
осуществляться сразу же, как только в миоцитах появляются первые порции АДФ.
При мышечной работе активность креатинкиназы
значительно возрастает за счет активирующего воздействия на нее ионов кальция,
концентрация которых в саркоплазме под действием нервного импульса
увеличивается почти в 1000 раз. Другой механизм регуляции креатин-фосфатной
реакции связан с активирующим воздействием на креатин-киназу креатина,
образующегося в ходе данной реакции. За счет этих механизмов активность
креатинкиназы в начале мышечной работы резко увеличивается и креатинфосфатная
реакция очень быстро достигает максимальной скорости.
Креатинфосфат, обладая большим запасом
химической энергии, является веществом непрочным. От него легко может
отщепляться фосфорная кислота, в результате чего происходит циклизация остатка
креатина, приводящая к образованию креатинина:
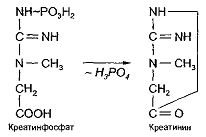
Образование креатинина происходит без участия
ферментов, спонтанно. Эта реакция необратима. Образовавшийся креатинин в
организме не используется и выводится с мочой. Поэтому по выделению креахинина
с мочой можно судить о содержании креатинфосфата в мышцах, так как в них
находятся основные запасы этого соединения.
Синтез креатинфосфата в мышечных клетках
происходит во время отдыха путем взаимодействия креатина с избытком АТФ:
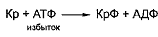
Частично запасы креатинфосфата могут
восстанавливаться и при мышечной работе умеренной мощности, при которой АТФ
синтезируется за счет тканевого дыхания в таком количестве, которого хватает и
на обеспечение сократительной функции миоцитов, и на восполнение запасов
креатинфосфата. Поэтому во время выполнения физической работы креатинфосфатная
реакция может включаться многократно.
Суммарные запасы АТФ и креатинфосфата часто
обозначают термином фосфагены.
Образование креатина происходит в печени с
использованием трех аминокислот: глицина, метионина и аргинина. В спортивной
практике для повышения в мышцах концентрации креатинфосфата используют в
качестве пищевых добавок препараты глицина и метионина.
Креатинфосфатный путь ресинтеза АТФ
характеризуется следующими величинами принятых количественных критериев:
Максимальная мощность составляет 900-1100
кал/мин-кг, что в три раза выше соответствующего показателя для аэробного
ресинтеза. Такая большая величина обусловлена высокой активностью фермента
креатинкиназы и, следовательно, очень высокой скоростью креатин-фосфатной
реакции.
Время развертывания всего 1-2 с. Как уже
указывалось, исходных запасов АТФ в мышечных клетках хватает на обеспечение
мышечной деятельности как раз в течение 1-2 с, и к моменту их исчерпания креатинфосфатный
путь образования АТФ уже функционирует со своей максимальной скоростью. Такое
малое время развертывания объясняется действием описанных выше механизмов
регуляции активности креатинкиназы, позволяющих резко повысить скорость этой
реакции.
Время работы с максимальной скоростью всего лишь
8-10 с, что связано с небольшими исходными запасами креатинфосфата в мышцах.
Главными преимуществами креатинфосфатного пути
образования АТф являются очень малое время развертывания и высокая мощность,
Что имеет крайне важное значение для скоростно-силовых видов спорта. Главным
недостатком этого способа синтеза АТФ, существенно ограничивающим его
возможности, является короткое время его функционирования. Время поддержания
максимальной скорости всего 8-10 с, к концу 30-й с его скорость снижается
вдвое. А к концу 3-й мин интенсивной работы креатинфосфатная реакция в мышцах
практически прекращается.
Исходя из такой характеристики креатинфосфатного
пути ресинтеза АТФ, следует ожидать, что эта реакция окажется главным источником
энергии для обеспечения кратковременных упражнений максимальной мощности: бег
на короткие дистанции, прыжки, метания, подъем штанги и т.п. Креатинфосфатная
реакция может неоднократно включаться во время выполнения физических нагрузок,
что делает возможным быстрое повышение мощности выполняемой работы, развития
ускорения на дистанции и финишный рывок.
Биохимическая оценка состояния креатинфосфатного
пути ресинтеза АТФ обычно проводится по двум показателям: креатининовому
коэффициенту и алактатному кислородному долгу.
Креатининовый коэффициент - это выделение
креатинина с мочой за сутки в расчете на 1 кг массы тела. У мужчин выделение
креатинина колеблется в пределах 18-32 мг/сутки-кг, а у женщин - 10-25
мг/сутки-кг. Креатининовый коэффициент характеризует запасы креатинфосфата в
мышцах, так как между содержанием креатинфосфата и образованием из него
креатинина существует линейная зависимость, поскольку это превращение протекает
неферментативным путем и является необратимым. Следовательно, с помощью креатининового
коэффициента можно оценить потенциальные возможности этого пути образования
АТФ, в том числе его метаболическую емкость.
Алактатный кислородный долг - это повышенное
потребление кислорода в ближайшие 4-5 мин после выполнения кратковременного упражнения
максимальной мощности. Этот избыток кислорода требуется для обеспечения высокой
скорости тканевого дыхания сразу же после окончания нагрузки для создания в
мышечных клетках повышенной концентрации АТФ. В этих условиях происходит
фосфорилирование креатина с образованием креатинфосфата:
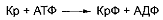
Таким образом, использование креатинфосфата во
время работы приводит к накоплению креатина, превращение которого снова в
креатинфосфат требует определенного количества кислорода. Отсюда следует, что
алактатный кислородный долг характеризует вклад креатинфосфатного пути
ресинтеза АТФ в энергообеспечение выполненной физической нагрузки и дает оценку
его метаболической емкости. Представление о мощности этого способа образования
АТФ дает показатель, полученный путем деления величины алактатного долга на
время выполнения нагрузки.
У квалифицированных спортсменов значение
алактатного кислородного долга после нагрузок максимальной мощности обычно
составляет 8-10 л.
В результате систематических тренировок,
направленных на развитие скоростно-силовых качеств, в мышцах увеличивается
концентрация креатинфосфата и повышается активность креатинкиназы, что находит
отражение в росте величины алактатного кислородного долга и суточного выделения
креатинина.
.2 Гликолитический путь ресинтеза АТФ
Этот путь ресинтеза, так же как и
креатинфосфатный, относится к анаэробным способам образования АТФ. Источником
энергии, необходимой для ресинтеза АТФ, в данном случае является мышечный
гликоген, концентрация которого в саркоплазме колеблется в пределах 0,2-3%. При
анаэробном распаде гликогена от его молекулы под воздействием фермента
фосфорилазы поочередно отщепляются концевые остатки глюкозы в форме
глюкозо-1-фосфата. Далее молекулы глюко-зо-1-фосфата через ряд последовательных
стадий превращаются в молочную кислоту, которая по своему химическому составу
является как бы половинкой молекулы глюкозы. В процессе анаэробного распада
гликогена до молочной кислоты, называемого гликолизом, образуются промежуточные
продукты, содержащие фосфатную группу с макроэргической связью, которая легко
переносится на АДФ с образованием АТФ.
Итоговое уравнение анаэробного расщепления
гликогена имеет следующий вид:
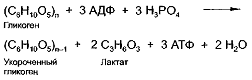
Все ферменты гликолиза находятся в саркоплазме
мышечных клеток.
Гликолизу может также подвергаться глюкоза,
поступающая в мышцы из кровяного русла. Анаэробный распад глюкозы протекает по
Уравнению:
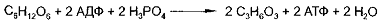
Регуляция скорости гликолиза осуществляется
путем изменения активности двух ферментов: фосфорилазы и фосфофруктокиназы.
Фосфорилаза катализирует первую реакцию распада гликогена - отщепление от него
глюкозо-1 - фосфата. Этот фермент активируется адреналином, АМФ и ионами
кальция, а ингибируется глюкозо-6-фосфатом и избытком АТФ. Второй регуляторный
фермент гликолиза - фосфофруктокиназа - активируется АДФ и особенно АМФ, а
тормозится избытком АТФ и лимонной кислотой. Наличие таких регуляторных
механизмов приводит к тому, что в покое гликолиз протекает очень медленно, при
интенсивной мышечной работе его скорость резко возрастает и может увеличиваться
по сравнению с уровнем покоя почти в 2000 раз, причем повышение скорости
гликолиза может наблюдаться уже в предстартовом состоянии за счет выделения
адреналина.
Количественные критерии гликолитического пути
ресинтеза АТФ:
Максимальная мощность - 750-850 кал/мин-кг, что
примерно вдвое выше соответствующего показателя тканевого дыхания. Высокое
значение максимальной мощности гликолиза объясняется содержанием в мышечных
клетках большого запаса гликогена, наличием механизмов активации ключевых
ферментов, приводящих к значительному росту скорости гликолиза, отсутствием
потребности в кислороде.
Время развертывания - 20-30 с. Это обусловлено
тем, что все участники гликолиза находятся в саркоплазме миоцитов, а также
возможностью активации ферментов гликолиза. Как уже отмечалось, фосфорилаза -
фермент, запускающий гликолиз, - активируется адреналином, который выделяется в
кровь непосредственно перед началом работы. Ионы кальция, концентрация которых
в саркоплазме повышается примерно в 1000 раз под воздействием двигательного
нервного импульса, также являются мощными активаторами фосфорилазы.
Время работы с максимальной мощностью - 2-3 мин.
Существуют две основные причины такой небольшой величины этого критерия.
Во-первых, гликолиз протекает с высокой скоростью, что быстро приводит к
уменьшению в мышцах концентрации гликогена и, следовательно, к последующему
снижению скорости его распада. Во-вторых, в процессе гликолиза образуется
молочная кислота, накопление которой приводит к повышению кислотности внутри
мышечных клеток. В условиях повышенной кислотности снижается каталитическая
активность ферментов, в том числе ферментов гликолиза, что также ведет к
уменьшению скорости этого пути ресинтеза АТФ.
Гликолитический способ образования АТФ имеет ряд
преимуществ перед аэробным путем. Он быстрее выходит на максимальную м0щность,
имеет более высокую величину максимальной мощности и не требует участия
митохондрий й кислорода.
Однако у этого пути есть и существенные
недостатки. Этот процесс малоэкономичен. Распад до лактата одного остатка
глюкозы, отщепленного от гликогена, дает только 3 молекулы АТФ, тогда как при
аэробном окислении гликогена до воды и углекислого газа образуется 39 молекул
АТФ в расчете на один остаток глюкозы. Такая неэкономичность в сочетании с
большой скоростью быстро приводит к исчерпанию запасов гликогена.
Другой серьезный недостаток гликолитического
пути ресинтеза АТФ - образование и накопление лактата, являющегося конечным
продуктом этого процесса. Повышение концентрации лактата в мышечных волокнах
вызывает сдвиг рН в кислую сторону, при этом происходят конформационные
изменения мышечных белков, приводящие к снижению их функциональной активности.
Таким образом, накопление молочной кислоты в мышечных клетках существенно
нарушает их нормальное функционирование и ведет к развитию утомления.
При снижении интенсивности физической работы, а
также в промежутках отдыха во время тренировки образовавшийся лактат может
частично выходить из мышечных клеток в лимфу или кровь, что делает возможным
повторное включение гликолиза.
Известные в настоящее время биохимические методы
оценки использования при физической работе гликолитического пути ресинтеза АТФ
основаны на оценке биохимических сдвигов в организме, обусловленных накоплением
молочной кислоты. Это прежде всего определение после физической нагрузки
концентрации лактата в крови. В покое, т.е. до начала работы, концентрация
лактата в крови обычно 1-2 ммоль/л. После интенсивных непродолжительных
нагрузок концентрация молочной кислоты в крови резко повышается и Может
достигать 18-20 ммоль/л, а у спортсменов высокой квалификации еще больших
значений. Другим показателем, отражающим накопление в кровяном русле молочной
кислоты, является водородный показатель крови. В покое этот показатель равен
7,36-7,40, после интенсивной работы он снижается до 7,2-7,0, причем в
литературе отмечено и еще более значительное снижение рН - до 6,8. Наибольшие
Изменения концентрации лактата и рН крови, наблюдаемые после на-Тузки "до
отказа" в зоне субмаксимальной мощности характеризуют Метаболическую
емкость гликолиза. Максимальную мощность гликолитического ресинтеза АТФ можно
оценить по скорости возрастания Концентрации лактата в крови или по скорости
снижения рН.
Еще один метод оценки скорости гликолиза,
фиксирующий последствия образования и накопления молочной кислоты, - это
определение щелочного резерва крови. Щелочной резерв крови - это щелочные
компоненты всех буферных систем крови. При поступлении во время мышечной работы
в кровь молочной кислоты она вначале нейтрализуется путем взаимодействия с
буферными системами крови, и поэтому происходит снижение щелочного резерва
крови.
Оценить вклад гликолиза в энергообеспечение
выполненной физической работы можно также путем определения лактата в моче. В
покое в моче лактат практически отсутствует. После тренировки, особенно с
использованием интенсивных упражнений, с мочой выделяются большие количества
молочной кислоты. При этом надо учесть, что в процессе тренировки гликолиз
включается многократно и поэтому анализ мочи дает информацию о суммарном вкладе
гликолитического пути ресинтеза в обеспечение энергией всех нагрузок,
выполненных за время тренировки.
Наряду с исследованием крови и мочи для оценки
гликолитического пути ресинтеза еще может быть использовано определение
лактатного кислородного долга. Лактатный кислородный долг - это повышенное
потребление кислорода в ближайшие 1-1,5 часа после окончания мышечной работы.
Этот избыток кислорода необходим для устранения молочной кислоты,
образовавшейся при работе. Наибольшие величины лактатного кислородного долга
определяются после физических нагрузок продолжительностью 2-3 мин, выполняемых
с предельной интенсивностью. У хорошо тренированных спортсменов величина лактатного
кислородного долга может достигать 20 л.
По величине лактатного кислородного долга можно
судить о возможностях гликолитического пути ресинтеза АТФ. Так, величина
лактатного долга свидетельствует о метаболической емкости гликолиза, а его
максимальная мощность может быть оценена по отношению величины лактатного долга
к времени выполнения предельной нагрузки субмаксимальной мощности.
В результате систематических тренировок с
использованием субмаксимальных нагрузок в мышечных клетках повышается концентрация
гликогена и увеличивается активность ферментов гликолиза. У высокотренированных
спортсменов наблюдается развитие резистентности тканей и крови к снижению рН, и
поэтому они сравнительно легко переносят сдвиг водородного показателя крови до
70 и ниже.
Аденилаткиназная реакция протекает в мышечных
клетках в условиях значительного накопления в них АДФ, что обычно наблюдается
при наступлении утомления. Аденилаткиназная реакция ускоряется ферментом
аденилаткиназой, который находится в саркоплазме миоцитов. В ходе этой реакции
одна молекула АДФ передает свою фосфатную группу на другую АДФ, в результате
образуется АТФ и АМФ:
2. Аэробный путь ресинтеза АТФ
Аэробный путь ресинтеза АТФ - это основной,
базовый способ образования АТФ, протекающий в митохондриях мышечных клеток. В
ходе тканевого дыхания от окисляемого вещества отнимаются два атома водорода и
по дыхательной цепи передаются на молекулярный кислород - 02, доставляемый
кровью в мышцы из воздуха, в результате чего возникает вода. За счет энергии,
выделяющейся при образовании воды, происходит синтез АТФ из АДФ и фосфорной
кислоты. Обычно на каждую образовавшуюся молекулу воды приходится синтез трех
молекул АТФ.
В упрощенном виде ресинтез АТФ аэробным путем
может быть представлен схемой:
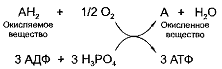
Чаще всего водород отнимается от промежуточных
продуктов цикла трикарбоновых кислот - цикла Кребса. Цикл Кребса - это
завершающий этап катаболизма, в ходе которого происходит окисление
ацетилкофермента А до С02 и Н20. В ходе этого процесса от перечисленных выше
кислот отнимается 4 пары атомов водорода и поэтому образуется 12 молекул АТФ
при окислении одной молекулы ацетилкофермента А.
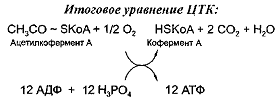
В свою очередь, ацетил-КоА может образовываться
из углеводов, жиров и аминокислот, т.е. через ацетил-КоА в цикл Кребса
вовлекаются углеводы, жиры и аминокислоты:
Скорость аэробного пути ресинтеза АТФ
контролируется содержанием в мышечных клетках АДФ, который является активатором
ферментов тканевого дыхания. В состоянии покоя, когда в клетках почти нет АДФ,
тканевое дыхание протекает с очень низкой скоростью. При мышечной работе за
счет интенсивного использования АТФ происходит образование и накопление АДФ.
Появившийся избыток АДФ ускоряет тканевое дыхание, и оно может достигнуть
максимальной интенсивности.
Другим активатором аэробного пути ресинтеза АТФ
является С02. Возникающий при физической работе в избытке углекислый газ
активирует дыхательный центр мозга, что в итоге приводит к повышению скорости
кровообращения и улучшению снабжения мышц кислородом.
Максимальная мощность составляет 350-450
кал/мин-кг. По сравнению с анаэробными путями ресинтеза АТФ тканевое дыхание
обладает самой низкой величиной максимальной мощности. Это обусловлено тем, что
возможности аэробного процесса ограничены доставкой кислорода в митохондрии и
их количеством в мышечных клетках. Поэтому за счет аэробного пути ресинтеза АТФ
возможно выполнение физических нагрузок только умеренной мощности.
Время развертывания - 3-4 мин. Такое большое
время развертывания объясняется тем, что для обеспечения максимальной скорости
тканевого дыхания необходима перестройка всех систем организма, участвующих в
доставке кислорода в митохондрии мышц.
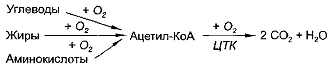
Время работы с максимальной мощностью составляет
десятки минут. Как уже указывалось, источниками энергии для аэробного ресинтеза
АТФ являются углеводы, жиры и аминокислоты, распад которых завершается циклом
Кребса. Причем для этой цели используются не только внутримышечные запасы
данных веществ, но и углеводы, жиры, кетоновые тела и аминокислоты,
доставляемые кровью в мышцы во время физической работы. В связи с этим данный
путь ресинтеза АТФ функционирует с максимальной мощностью в течение такого
продолжительного времени.
По сравнению с другими идущими в мышечных
клетках процессами ресинтеза АТФ аэробный ресинтез имеет ряд преимуществ. Он
отличается высокой экономичностью: в ходе этого процесса идет глубокий распад
окисляемых веществ до конечных продуктов - С02 и Н20 и поэтому выделяется
большое количество энергии. Так, например, при аэробном окислении мышечного
гликогена образуется 39 молекул АТФ в расчете на каждую отщепляемую от
гликогена молекулу глюкозы, в то время как при анаэробном распаде этого
углевода синтезируется только 3 молекулы АТФ в расчете на одну молекулу
глюкозы. Другим достоинством этого пути ресинтеза является универсальность в
использовании субстратов. В ходе аэробного ресинтеза АТФ окисляются все основные
органические вещества организма: аминокислоты, углеводы, жирные кислоты,
кетоновые тела и др. Еще одним преимуществом этого способа образования АТФ
является очень большая продолжительность его работы: практически он
функционирует постоянно в течение всей жизни. В покое скорость аэробного
ресинтеза АТФ низкая, при физических нагрузках его мощность может стать
максимальной.
Однако аэробный способ образования АТФ имеет и
ряд недостатков. Так, действие этого способа связано с обязательным
потреблением кислорода, доставка которого в мышцы обеспечивается дыхательной и
сердечнососудистой системами. Функциональное состояние кардиореспираторной
системы является лимитирующим фактором, ограничивающим продолжительность работы
аэробного пути ресинтеза АТф с максимальной мощностью и величину самой
максимальной мощности.
Возможности аэробного пути ограничены еще и тем,
что все ферменты тканевого дыхания встроены во внутреннюю мембрану митохондрий
в форме дыхательных ансамблей и функционируют только ffPH наличии неповрежденной
мембраны. Любые факторы, влияющие На состояние и свойства мембран, нарушают
образование АТФ аэробным способом. Например, нарушения окислительного
фосфорилирования наблюдаются при ацидозе, набухании митохондрий, при развитии в
мышечных клетках процессов свободно-радикального окисления липидов, входящих в
состав мембран митохондрий.
Еще одним недостатком аэробного образования АТФ
можно считать большое время развертывания и небольшую по абсолютной величине
максимальную мощность. Поэтому мышечная деятельность, свойственная большинству
видов спорта, не может быть полностью обеспечена этим путем ресинтеза АТФ и
мышцы вынуждены дополнительно включать анаэробные способы образования АТФ,
имеющие более короткое время развертывания и большую максимальную мощность.
В спортивной практике для оценки аэробного
фосфорилирования часто используют три показателя: максимальное потребление
кислорода, порог анаэробного обмена и кислородный приход.
МПК - это максимально возможная скорость
потребления кислорода организмом при выполнении физической работы. Этот
показатель характеризует максимальную мощность аэробного пути ресинтеза АТФ:
чем выше величина МПК, тем больше значение максимальной скорости тканевого
дыхания, это обусловлено тем, что практически весь поступающий в организм
кислород используется в этом процессе. МПК представляет собой интегральный
показатель, зависящий от многих факторов: от функционального состояния
кардиореспираторной системы, от содержания в крови гемоглобина, а в мышцах -
миоглобина, от количества и размера митохондрий. У нетренированных молодых
людей МПК обычно равно 3-4 л/мин, у спортсменов высокого класса, выполняющих
аэробные нагрузки, МПК - 6-7 л/мин. На практике, для исключения влияния на эту
величину массы тела МПК рассчитывают на кг массы тела. В этом случае у молодых
людей, не занимающихся спортом, МПК равно 40-50 мл/мин-кг, а у хорошо
тренированных спортсменов - 80-90 мл/мин-кг.
В спортивной практике МПК также используется для
характеристики относительной мощности аэробной работы, которая выражается
потреблением кислорода в процентах от МПК. Например, относительная мощность
работы, выполняемой с потреблением кислорода 3 л/мин спортсменом, имеющим МПК,
равное 6 л/мин, будет составлять 50% от уровня МПК. ПАНО - это минимальная
относительная мощность работы, измеренная по потреблению кислорода в процентах
по отношению к МПК, при которой начинает включаться гликолитический путь
ресинтеза АТФ. у нетренированных ПАНО составляет 40-50% от МПК, а у спортсменов
ПАНО может достигать 70% от МПК. Более высокие величины ПАНО у тренированных
объясняются тем, что аэробное фосфорилирование у них дает больше АТФ в единицу
времени, и поэтому анаэробный путь образования АТФ - гликолиз - включается при
больших нагрузках. Кислородный приход - это количество кислорода,
использованное во время выполнения данной нагрузки для обеспечения аэробного
ресинтеза АТФ. Кислородный приход характеризует вклад тканевого дыхания в
энергообеспечение проделанной работы.
Под влиянием систематических тренировок,
направленных на развитие аэробной работоспособности, в миоцитах возрастает
количество митохондрий, увеличивается их размер, в них становится больше
ферментов тканевого дыхания. Одновременно происходит совершенствование
кислородтранспортной функции: повышается содержание миоглобина в мышечных
клетках и гемоглобина в крови, возрастает работоспособность дыхательной и
сердечнососудистой систем организма.
Длительное время этот путь образования АТФ
рассматривался как аварийный механизм, обеспечивающий синтез АТФ в условиях,
когда другие способы получения АТФ становятся неэффективными. Кроме того,
считалось, что аденилаткиназная реакция ведет к уменьшению в миоцитах общего
количества адениловых нуклеотидов, так как образующийся в этой реакции АМФ может
дезаминироваться и превращаться в инозиновую кислоту:

Однако в настоящее время этой реакции отводят не
энергетическую, а регуляторную роль. Это связано с тем, что АМФ является мощным
активатором ферментов распада углеводов - фосфорилазы и фосфофруктокиназы,
участвующих как в анаэробном расщеплении гликогена и глюкозы до молочной
кислоты, так и в их аэробном окислении до воды и углекислого газа. Оказалось
также, что превращение АМФ в инозиновую кислоту имеет положительное значение
для мышечной деятельности. Образующийся в результате дезаминирования аммиак
может нейтрализовать молочную кислоту и тем самым предупреждать наступление
изменений в миоцитах, связанных с ее накоплением. При этом общее содержание
адениловых нуклеотидов в клетках не изменяется, так как инозиновая кислота при
взаимодействии с одной из аминокислот - аспарагиновой кислотой снова
превращается в АМФ.
В табл. приведены величины критериев описанных
выше путей Ресинтеза АТФ. Количественные критерии основных путей ресинтеза АТФ:
|
Критерии
|
Пути
ресинтеза
|
|
Креатин-фосфатный
|
Гликолити-ческий
|
Аэробный
|
|
Максимальная
мощность, кал/мин-кг
|
900-1100
|
750-850
|
35О-450
|
|
Время
развертывания
|
1-2
с
|
20-30
с
|
3-4
мин
|
|
Время
сохранения максимальной мощности
|
8-10
с
|
2-3
мин
|
десятки
минут
|
Соотношение между различными путями ресинтеза
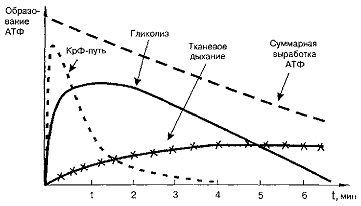
АТФ при мышечной работе
При любой мышечной работе функционируют все три
пути ресинтеза АТФ, но включаются они последовательно. В первые секунды работы
ресинтез АТФ идет за счет креатинфосфатной реакции, затем включается гликолиз
и, наконец, по мере продолжения работы на смену гликолизу приходит тканевое
дыхание.
Включение путей ресинтеза АТФ при выполнении
физической работы
Из рисунка видно, что переход энергообеспечения
мышечной деятельности с анаэробных путей на аэробный ведет к уменьшению
суммарной выработки АТФ за единицу времени, что находит отражение в снижении
мощности выполняемой работы.
Конкретный вклад каждого из механизмов
образования АТФ в энергообеспечение мышечных движений зависит от интенсивности
и продолжительности физических нагрузок.
При кратковременной, но очень интенсивной работе
главным источником АТФ является креатинкиназная реакция, при более
продолжительной интенсивной работе большая часть АТФ образуется гликолитическим
путем. При выполнении упражнений большой продолжительности, но умеренной
мощности энергообеспечение мышц осуществляется в основном за счет аэробного
окисления.
3. Зоны относительной мощности мышечной работы
В настоящее время приняты различные
классификации мощности мышечной деятельности. Одна из них - классификация по
B.C. Фарфелю, базирующаяся на положении о том, что мощность выполняемой
физической нагрузки обусловлена соотношением между тремя основными путями ресинтеза
АТФ, функционирующими в мышцах во время работы. Согласно этой классификации
выделяют четыре зоны относительной мощности мышечной работы: максимальной,
субмаксимальной, большой и умеренной мощности.
Работа в зоне максимальной мощности может
продолжаться в течение 15-20 с. Основной источник АТФ в этих условиях -
креатин-фосфат. Только в конце работы креатинфосфатная реакция замещается
гликолизом. Примером физических упражнений, выполняемых в зоне максимальной
мощности, является бег на короткие дистанции, прыжки в длину и высоту,
некоторые гимнастические упражнения, подъем штанги и др.
Работа в зоне субмаксимальной мощности имеет
продолжительность до 5 мин. Ведущий механизм ресинтеза АТФ - гликолитический. В
начале работы, пока гликолиз не достиг максимальной скорости, образование АТФ
идет за счет креатинфосфата, а в конце работы гликолиз начинает заменяться
тканевым дыханием. Работа в зоне субмаксимальной мощности характеризуется самым
высоким кислородным долгом - До 20 л. Примером физических нагрузок в этой зоне
мощности является бег на средние дистанции, плавание на короткие дистанции,
велосипедные гонки на треке, бег на коньках на спринтерские дистанции и др.
Работа в зоне большой мощности имеет предельную
продолжительность до 30 мин. Для работы в этой зоне характерен примерно
одинаковый вклад гликолиза и тканевого дыхания. Креатинфосфатный Путь ресинтеза
АТФ функционирует только в самом начале работы, и поэтому его доля в общем
энергообеспечении данной работы мала. Примером упражнений в этой зоне мощности
является бег на 5000 ц бег на коньках на стайерские дистанции, лыжные гонки по
пересеченной местности, плавание на средние и длинные дистанции и др.
Работа в зоне умеренной мощности продолжается
свыше 30 мин. Энергообеспечение мышечной деятельности происходит
преимущественно аэробным путем. Примером работы такой мощности является
марафонский бег, легкоатлетический кросс, спортивная ходьба, шоссейные
велогонки, лыжные гонки на длинные дистанции, турпохода и др.
В ациклических и ситуационных видах спорта мощность
выполняемой работы многократно изменяется. Так, у футболиста бег с умеренной
скоростью чередуется с бегом на короткие дистанции со спринтерской скоростью;
можно найти и такие отрезки игры, когда мощность работы значительно снижается.
Такие примеры можно привести в отношении многих других видов спорта.
Однако в ряде спортивных дисциплин все же
преобладают физические нагрузки, относящиеся к какой-то определенной зоне
мощности. Так, физическая работа лыжников обычно выполняется с большой или
умеренной мощностью, а в тяжелой атлетике используются максимальные и
субмаксимальные нагрузки.
Поэтому при подготовке спортсменов необходимо
применять тренировочные нагрузки, развивающие путь ресинтеза АТФ, являющийся
ведущим в энергообеспечении работы в зоне относительной мощности, характерной
для данного вида спорта.
4. Биохимические изменения в мышцах, головном
мозге, печени, крови, моче
.1 Биохимические изменения в скелетных мышцах
При выполнении физической работы в мышцах
происходят глубокие изменения, обусловленные прежде всего интенсификацией
процессов ресинтеза АТФ.
Использование креатинфосфата в качестве
источника энергии приводит к снижению его концентрации в мышечных клетках и
накоплению в нихкреатина.
Практически при любой работе для получения АТФ
используется мышечный гликоген. Поэтому его концентрация в мышцах снижается
независимо от характера работы. При выполнении интенсивных нагрузок в мышцах
наблюдается быстрое уменьшение запасов гликогена и одновременное образование и
накопление молочной кислоты. За счет накопления молочной кислоты повышается
кислотность внутри мышечных клеток. Увеличение содержания лактата в мышечных
клетках вызывает также повышение в них осмотического давления, вследствие чего
в миоциты из капилляров и межклеточных пространств поступает вода и развивается
набухание мышц.
Продолжительная мышечная работа небольшой
мощности вызывает плавное снижение концентрации гликогена в мышцах. В данном
случае распад гликогена протекает аэробно, с потреблением кислорода. Конечные
продукты такого распада - углекислый газ и вода - удаляются из мышечных клеток
в кровь. Поэтому после выполнения работы умеренной мощности в мышцах
обнаруживается уменьшение содержания гликогена без накопления лактата.
Еще одно важное изменение, возникающее в
работающих мышцах, -повышение скорости распада белков. Особенно ускоряется
распад белков при выполнении силовых упражнений, причем это затрагивает в
первую очередь сократительные белки, входящие в состав миофиб-рилл. Вследствие
распада белков в мышечных клетках повышается содержание свободных аминокислот и
продуктов их последующего расщепления - кетокислот и аммиака.
Другим характерным изменением, вызываемым
мышечной деятельностью, является снижение активности ферментов мышечных клеток.
Одной из причин уменьшения ферментативной активности может быть повышенная
кислотность, вызванная накоплением в мышцах лактата.
И наконец, мышечная деятельность может привести
к повреждениям внутриклеточных структур - миофибрилл, митохондрий,
разнообразныхбиомембран. Так, повреждение мембран саркоплазматического
ретикулума ведет к нарушению проведения нервного импульса к цистернам,
содержащим ионы кальция. Нарушение целостности сарколеммы сопровождается
потерей мышцами многих важных веществ, в том числе ферментов, которые через
поврежденную сарколемму уходят из мышечных клеток в лимфу и кровь.
Повреждение мембран также негативно влияет на
активность иммобилизованных ферментов, т. е. ферментов, встроенных в мембраны.
Эти ферменты могут полноценно функционировать только при Наличии неповрежденной,
целостной мембраны. Например, при мышечной работе может снижаться активность
кальциевого насоса -фермента, встроенного в мембрану цистерн и обеспечивающего
транспорт ионов кальция из саркоплазмы внутрь цистерн. Другой Пример: при
продолжительной физической работе уменьшается активность ферментов тканевого
дыхания, локализованных во внутренней мембране митохондрий.
4.2 Биохимические сдвиги в головном мозге
Во время мышечной деятельности в мотонейронах
коры головного мозга происходит формирование и последующая передача
двигательного нервного импульса. Оба эти процесса - формирование и передача
нервного импульса - осуществляются с потреблением энергии молекул АТФ.
Образование АТФ в нервных клетках происходит аэробно, путем окислительного
фосфорилирования. Поэтому при мышечной работе увеличивается потребление мозгом
кислорода из протекающей крови. Другой особенностью энергетического обмена в
нейронах является то, что основным субстратом окисления является глюкоза,
поступающая с током крови.
В связи с такой спецификой энергоснабжения
нервных клеток любое нарушение снабжения мозга кислородом или глюкозой
неминуемо ведет к снижению его функциональной активности, что у спортсменов
может проявляться в форме головокружения или обморочного состояния.
.3 Биохимические сдвиги в миокарде
Во время мышечной деятельности происходит
усиление и учащение сердечных сокращений, что требует большего количества
энергии по сравнению с состоянием покоя. Однако энергообеспечение сердечной
мышцы осуществляется главным образом за счет аэробного ресинтеза АТФ.
Анаэробные пути ресинтеза АТФ включаются лишь при очень интенсивной работе.
Большие возможности аэробного энергообеспечения
в миокарде обусловлены особенностью строения этой мышцы. В отличие от скелетных
мышц в сердечной имеется более развитая, густая сеть капилляров, что позволяет
извлекать из протекающей крови больше кислорода и субстратов окисления. Кроме
того, в клетках миокарда имеется больше митохондрий, содержащих ферменты
тканевого дыхания. В качестве источников энергии миокард использует различные
вещества, доставляемые кровью: глюкозу, жирные кислоты, кетоновые тела,
глицерин. Собственные запасы гликогена практически не используются; они
необходимы для энергообеспечения миокарда при истощающих нагрузках.
Во время интенсивной работы, сопровождающейся
увеличением концентрации лактата в крови, миокард извлекает из крови лактат и
окисляет его до углекислого газа и воды. При окислении одной молекулы молочной
кислоты синтезируется до 18 молекул АТФ. Способность миокарда окислять лактат
имеет большое биологическое значение. Использование лактата в качестве
источника энергии позволяет дольше поддерживать в крови необходимую
концентрацию глюкозы, что очень существенно для биоэнергетики нервных клеток,
для которых глюкоза является почти единственным субстратом окисления. Окисление
лактата в сердечной мышце также способствует нормализации кислотно-щелочного
баланса, так как при этом в крови снижается концентрация этой кислоты.
.4 Биохимические сдвиги в печени
При мышечной деятельности активируются функции
печени, направленные преимущественно на улучшение обеспечения работающих мышц
внемышечными источниками энергии, переносимыми кровью. Ниже описаны наиболее
важные биохимические процессы, протекающие в печени во время работы.
. Во время выполнения физических нагрузок клетки
печени активно извлекают из крови жир и жирные кислоты, содержание которых в
крови возрастает вследствие мобилизации жира из жировых депо. Поступающий в
печеночные клетки жир сразу же подвергается гидролизу и превращается в глицерин
и жирные кислоты. Далее жирные кислоты путем Р-окисления расщепляются до
ацетил-КоА, из которого затем образуются кетоновые тела - ацетоуксусная и
Р-оксимасляная кислоты. Синтез кетоновых тел обычно называется кетогенезом.
Кетоновые тела являются важными источниками энергии. С током крови они
переносятся из печени в работающие органы - миокард и скелетные мышцы. В этих
органах кетоновые тела вновь превращаются в ацетил-КоА, который сразу же
аэробно окисляется в цикле Кребса до Углекислого газа и воды с выделением
большого количества энергии.
. Еще один биохимический процесс, протекающий в
печени во время работы, - глюконеогенез. Уже отмечалось, что этот процесс
Инициируется глюкокортикоидами. За счет глюконеогенеза в клетках Печени из
глицерина, аминокислот и лактата осуществляется синтез глюкозы. Этот процесс
идет с затратой энергии АТФ. Обычно глюконеогенез протекает при длительной
работе, ведущей к снижению концентрации глюкозы в кровяном русле. Благодаря
глюконеогенезу организму удается поддерживать в крови необходимый уровень
глюкозы.
. При физической работе усиливается распад
мышечных белков, приводящий к образованию свободных аминокислот, которые далее
де-заминируются, выделяя NH3. Аммиак является клеточным ядом, его
обезвреживание происходит в печени, где он превращается в мочевину. Синтез
мочевины требует значительного количества энергии. При истощающих нагрузках,
несоответствующих функциональному состоянию организма, печень может не
справляться с обезвреживанием аммиака, в этом случае возникает интоксикация
организма этим ядом, ведущая к снижению работоспособности.
.5 Биохимические сдвиги в крови
Изменения химического состава крови является
отражением тех биохимических сдвигов, которые возникают при мышечной
деятельности в различных внутренних органах, скелетных мышцах и миокарде.
Поэтому на основании анализа химического состава крови можно оценить
биохимические процессы, протекающие во время работы. Это имеет большое
практическое значение, так как из всех тканей организма кровь наиболее доступна
для исследования.
Биохимические сдвиги, наблюдаемые в крови, в
значительной мере зависят от характера работы, и поэтому их анализ следует
проводить с учетом мощности и продолжительности выполненных нагрузок.
При выполнении мышечной работы в крови чаще
всего обнаруживаются следующие изменения:
. Повышение концентрации белков в плазме крови.
Это происходит по двум причинам. Во-первых, усиленное потоотделение приводит к
уменьшению содержания воды в плазме крови и, следовательно, к ее сгущению, в
результате чего возрастают концентрации всех компонентов плазмы, в том числе
белков. Во-вторых, вследствие повреждения клеточных мембран наблюдается выход
внутриклеточных белков в плазму крови. Однако при очень продолжительной работе
возможно снижение концентрации белков плазмы. В этом случае часть белков из
кровяного русла переходит в мочу, а другая часть используется в качестве
источников энергии.
. Изменение концентрации глюкозы в крови во
время работы характеризуется фазностью. В начале работы обычно уровень глюкозы
в крови возрастает. Это объясняется тем, что в начале работы в печени имеются
большие запасы гликогена и глюкогенез протекает с высокой скоростью. С другой
стороны, в начале работы мышцы тоже обладают значительными запасами гликогена,
которые они используют для своего энергообеспечения, и поэтому не извлекают
глюкозу из кровяного русла. По мере выполнения работы снижается содержание
гликогена как в печени, так и в мышцах. В связи с этим печень направляет все
меньше и меньше глюкозы в кровь, а мышцы, наоборот, начинают в большей мере
использовать глюкозу крови для получения энергии. При длительной работе часто
наблюдается снижение концентрации глюкозы в крови, что обусловлено истощением
запасов гликогена и в печени, и в мышцах.
. Повышение концентрации лактата в крови
наблюдается практически при любой спортивной деятельности, однако степень
возрастания концентрации лактата в значительной мере зависит от характера
выполненной работы и тренированности спортсмена. Наибольший подъем уровня
лактата в крови отмечается при выполнении физических нагрузок в зоне
субмаксимальной мощности, так как в этом случае главным источником энергии для
работающих мышц является анаэробный гликолиз, приводящий к образованию и
накоплению молочной кислоты.
В покое, до работы содержание лактата в крови
равняется 1-2 ммоль/л. После работы «до отказа» в зоне субмаксимальной мощности
у спортсменов средней квалификации концентрация лактата в крови увеличивается
до 8-10 ммоль/л, у высокотренированных этот рост может достигать 18-20 ммоль/л
и выше. В литературе описаны случаи повышения лактата в крови у очень хорошо
подготовленных спортсменов до 30-32 ммоль/л.
При проведении анализа крови на содержание
лактата необходимо учитывать, что увеличение его концентрации в крови
происходит не сразу, а через несколько минут после окончания работы. Поэтому
забор крови следует делать примерно через 5 мин после завершения нагрузки. При
взятии крови в более поздние сроки концентрация лактата окажется заниженной,
так как часть его будет извлечена из кровяного русла клетками миокарда и
печени.
. Водородный показатель. Образующийся при
интенсивной работе лактат является сильной кислотой и его поступление в
кровяное русло должно вести к повышению кислотности крови. Однако первые порции
лактата, диффундирующие из мышц в кровяное русло, нейтрализуются буферными
системами крови. В дальнейшем, по мере исчерпания емкости буферных систем,
наблюдается повышение кислотности Крови, возникает так называемый
некомпенсированный ацидоз. В покое значение рН венозной крови равно 7,35-7,36.
При мышечной работе вследствие накопления в крови лактата, величина рН
уменьшается. При выполнении физических упражнений субмаксимальной мощности РН
снижается у спортсменов средней квалификации до 7,1-7,2, а у спортсменов
мирового класса снижение водородного показателя может быть до 6,8.
. Повышение концентрации свободных жирных кислот
и кетоновых тел наблюдается при длительной мышечной работе вследствие
мобилизации жира из жировых депо и последующим кетогенезом в печени. Увеличение
концентрации кетоновых тел также вызывает повышение кислотности и снижение рН
крови.
. Мочевина. При кратковременной работе
концентрация мочевины в крови увеличивается незначительно, а при длительной
физической работе уровень мочевины в крови может возрасти в 4-5 раз. Причиной
увеличения содержания мочевины в крови является усиление катаболизма белков под
воздействием физических нагрузок, особенно силового характера. Распад белков, в
свою очередь, ведет к накоплению свободных аминокислот, при распаде которых
образуется в большом количестве аммиак. В печени большая часть образовавшегося
аммиака превращается в мочевину.
.5 Биохимические сдвиги в моче
Выполнение физических нагрузок приводит также к
значительным сдвигам в химическом составе мочи и существенно влияет на ее
физико-химические свойства.
После завершения мышечной работы наиболее
характерным является появление в моче химических веществ, которые в покое
практически отсутствуют. Эти соединения часто называют патологическими
компонентами, так как они появляются в моче не только после физических
нагрузок, но и при ряде заболеваний. У спортсменов после выполнения
тренировочных или соревновательных нагрузок в моче обнаруживаются следующие
патологические компоненты:
. Белок. У здорового человека, не занимающегося
спортом, в сутки выделяется не более 100 мг белка. Поэтому в порциях мочи,
взятых для анализа до тренировки, обычными методами белок не обнаруживается.
После выполнения мышечной работы отмечается значительное выделение с мочой
белка. Это явление носит название протеинурия. Особенно выраженная протеинурия
наблюдается после чрезмерных нагрузок, не соответствующих функциональному
состоянию спортсмена. Вероятными причинами протеинурии являются повреждение
почечных мембран, возникающее под влиянием мышечных нагрузок, а также появление
в крови во время физической работы продуктов деградации тканевых белков - различных
полипептидов, легко проходящих через почечный фильтр из кровяного русла в
состав мочи.
. Глюкоза. В порциях мочи, полученных до
выполнения физической нагрузки, глюкоза практически отсутствует. После
завершения тренировки в моче спортсменов общепринятыми методиками нередко
обнаруживается значительное содержание глюкозы, что может быть обусловлено
двумя основными причинами. Во-первых, как уже отмечалось, при выполнении
физических упражнений в крови повышается уровень глюкозы и он может превысить
почечный порог, вследствие чего часть глюкозы не будет подвергаться обратному
всасыванию в извитых канальцах нефрона и останется в составе мочи. Во-вторых,
из-за повреждения почечных мембран нарушается непосредственно процесс обратного
всасывания глюкозы в почках, что также ведет к развитию глюкозурии.
. Кетоновые тела. До работы кетоновые тела в
моче не обнаруживаются. После соревновательных или тренировочных нагрузок с
мочой могут выделяться в больших количествах кетоновые тела - ацето-уксусная и
Р-оксимасляная кислоты, а также продукт их распада - ацетон. Это явление
называется кетонурией, или ацетонурией. Причины кетонурии аналогичны причинам,
вызывающим глюкозурию. Это повышение в крови концентрации кетоновых тел и
снижение реабсорбционной функции почек при мышечной работе.
. Лактат. Появление молочной кислоты в моче
обычно наблюдается после тренировок, включающих упражнения субмаксимальной
мощности. Каждое такое упражнение приводит к резкому возрастанию концентрации
лактата в крови и последующему его переходу из кровяного русла в мочу. Таким
образом, происходит аккумулирование молочной кислоты в моче. В связи с этим по
выделению лактата с мочой можно судить об общем вкладе гликолитического пути
ресинтеза АТФ в энергообеспечение всей работы, выполненной спортсменом за
тренировку.
Наряду с влиянием на химический состав
физические нагрузки приводят к изменению физико-химических свойств мочи.
Наиболее существенные изменения следующие:
Плотность. Вследствие повышения роли внепочечных
путей выделения воды из организма объем мочи после тренировки или соревнования,
как правило, уменьшается. Это, в свою очередь, сказывается на плотности. Данный
показатель после работы чаще всего повышайся. В среднем плотность мочи до
нагрузок колеблется в пределах 1.010-1,025 г/мл. После тренировки этот
показатель может быть равен 1,030-1,035 г/мл и даже еще выше. Одной из причин
увеличения плотности мочи является, как отмечалось выше, увеличение внепочечных
потерь воды, что приводит к возрастанию концентрации растворенных в моче
веществ. Другой причиной повышения плотности мочи после физической работы может
оказаться появление в моче веществ, отсутствующих в ней в состоянии покоя.
По плотности можно рассчитать содержание
растворенных химических соединений в отдельных порциях мочи. Для этого используется
следующая формула:
Сухой остаток = а • 2,6,
где а - величина, численно равная тысячным долям
плотности.
Это делает возможным оценивать экскрецию
отдельных компонентов мочи в расчете на 1 г сухого остатка, что позволяет
исключить влияние плотности мочи на эти показатели.
Кислотность. Вследствие выделения после
тренировки с мочой молочной кислоты, а также кетоновых тел, которые тоже
являются кислотами, рН мочи снижается. До работы при обычном питании рН мочи
равен 5-6. После работы, особенно с использованием интенсивных нагрузок, рН
мочи может быть в пределах 4-5, что соответствует примерно десятикратному
увеличению концентрации в моче ионов водорода.
В заключение необходимо подчеркнуть, что глубина
биохимических сдвигов, возникающих в мышцах, во внутренних органах, в крови и в
моче, зависит от мощности и продолжительности физической работы. Чем выше
интенсивность работы и чем дольше она длится, тем более глубоки и значительны
биохимические изменения в организме спортсмена. Достигнув определенной глубины,
биохимические сдвиги оказывают отрицательное влияние на возможность выполнения
данной работы и приводят к снижению работоспособности спортсмена, т. е.
вызывают утомление.
Библиографический список
ресинтез аденозинтрифосфат нагрузка
мышца
Медведева
Г.Е. Биоэнергетика мышечной деятельности: учебное пособие. - Челябинск, 2006.
Биохимия
физической культуры и спорта: учебно-методическое пособие (составители Г.Е.
Медведева, Т.В. Соломина) - Челябинск, 2006.
Соломина
Т.В. Особенности процессов энергообеспечения физических нагрузок в циклических
видах спорта. Учебное пособие - Омск, Челябинск, 1987.
Биохимия.
Учебник для институтов физической культуры/ под ред. В.В. Меньшикова и Н.И.
Волкова - М.: ФиС, 1986.
Биохимия
физической культуры и спорта. Учебно-методическое пособие (сост. Г.Е.
Медведева, Т.В. Соломина) - Челябинск, 2003.
Биоэнергетика
мышечной деятельности. Учебное пособие. Г.Е. Медведева - Челябинск, 2003.
Особенности
процессов энергообеспечения физических нагрузок в циклических видах спорта. Учебное
пособие. Т.В. Соломина - Омск, Челябинск, 1987.
Биохимия
обменных процессов. Учебное пособие для студентов институтов и факультетов
физической культуры. - Челябинск, 1999.
Биохимические
изменения в организме при мышечной деятельности. Методические рекомендации по
выполнению курсовой работы. Сост. Т.В. Соломина, Н.В. Князев - Челябинск,
2005.://topref.ru/referat/11842.html
Биохимия
физической культуры и спорта: учебно-методическое пособие (составители Г.Е.
Медведева, Т.В. Соломина) - Челябинск, 2006. - С. 69.
1.