Влияние внешних факторов на организм животных
ВЛИЯНИЕ ВНЕШНИХ ФАКТОРОВ НА ОРГАНИЗМ
ЖИВОТНЫХ
БЕЙРУТ 2012
Содержание
Введение
Факторы внешней среды и их влияние на естественную
резистентность и продуктивность свиней
Физиологическое значение минеральных веществ в организме свиней
Использование источников минерального питания в кормлении
свиней
Использование пикумина свиноматкам в течение первого периода
супоросности (до 84-го дня) и лактации
естественные защитные силы и показатели крови свиноматок
продуктивность и воспроизводительные способности свиноматок
энергия роста и гематологические показатели поросят-сосунов
Использование пикумина свиноматкам в течение второго периода
супоросности (с 84-го дня) и лактации
естественные защитные силы и показатели крови свиноматок
продуктивность и воспроизводительные способности свиноматок
энергия роста и гематологические показатели поросят-сосунов
Заключение
Выводы
Указатель литературы
Введение
В эволюционном развитии по мере интенсификации отрасли свиньи все более
отходят от естественных условий обитания. Создание регулируемых условий жизни
животных при промышленных методах производства, целенаправленное и усиленное
кормление для повышения продуктивности в соответствии с наследственными
возможностями, научно-обоснованная селекция являются лишь некоторыми из
основных условий современного ведения свиноводства.
Организм сельскохозяйственных животных находится под постоянным
воздействием самых разнообразных факторов внешней среды. К этим факторам
относится все то, что оказывает влияние на жизнеспособность, поведение и
продуктивность животных: воздушная среда, почва, количество, состав и качество
кормовых средств и воды, способы и распорядок кормления и поения животных,
технология их содержания, плотность размещения, размеры групп и т.д. Особенно
факторы внешней среды оказывают влияние на молодой организм. И это влияние
начинается уже с эмбрионального развития, когда идет закладка продуктивных
качеств животного, становление его защитных сил. Только оптимальные условия
содержания и кормления супоросных и лактирующих свиноматок, высокая
резистентность их организма могут способствовать получению большего количества
приплода с высокой жизнеспособностью и продуктивностью, развитыми естественными
защитными силами организма.
Важная роль в повышении естественной резистентности организма животных и
их продуктивности отводится биологически активным веществам, в том числе и
макро- и микроэлементам. Минеральные вещества, хотя и не представляют
энергетической ценности, однако имеют огромное значение для организма свиней.
Объясняется это той большой ролью, которую они играют во всех процессах обмена,
происходящих в организме.
Основным источником минеральных веществ для сельскохозяйственных животных
являются корма растительного происхождения. Но поскольку минеральный состав
кормов непостоянен, подвержен значительным колебаниям по сельскохозяйственным
регионам и находится в зависимости от вида растений, сорта, периода вегетации,
почвы и других условий, количество минеральных веществ в рационе не
обеспечивает физиологическую потребность животных. В связи с этим животноводы
вынуждены использовать другие источники минеральных веществ, содержащие те или
иные недостающие в рационе минеральные элементы.
Существуют различные способы компенсации недостатка минеральных веществ в
рационе. Это и приготовление химической промышленностью различных полисолей,
премиксов, минеральных брикетов, и использование природных источников
минеральных веществ, отходов промышленности, содержащих те или иные макро- и
микроэлементы. Зачастую разработка сырьевых минеральных источников,
приготовление минеральных добавок, их транспортировка требует больших
материальных затрат. Учитывая то, что в настоящее время Беларусь испытывает
недостаток в минеральных подкормках для животных, и их приходится закупать за
рубежом, большинство предлагаемых на рынке источников минерального питания
остаются слишком дорогими для многих хозяйств республики.
Альтернативой могут послужить отходы производства керамзита - пикумин.
Богатый минеральный состав пикумина, технологичность при приготовлении
кормосмесей служат предпосылкой к его использованию в качестве источника
недостающих макро- и микроэлементов в рационе свиней, в том числе и для
супоросных и подсосных свиноматок.
Факторы внешней среды и их влияние на естественную резистентность и
продуктивность свиней
Продуктивность клинически здоровых животных на 60…70% зависит от качества
и полноты кормления, на 20…30% от состояния микроклимата помещений и на 10…20%
от соответствующего ухода при эксплуатации. Доброкачественную продукцию можно
получить только от здоровых животных. Здоровье сельскохозяйственных животных
зависит от уровня естественной устойчивости организма к болезням, полученной по
наследству от родителей, сформированной в процессе роста и развития молодняка; от
условий содержания в помещениях, отвечающих физиологическим потребностям; от
качества и безвредности кормов, питьевой воды и состояния микроклимата
животноводческих помещений. Создание животным условий, обеспечивающих здоровье
и высокую продуктивность, является одной из важнейших задач в развитии
животноводства.
В настоящее время у ученых и практиков-свиноводов не ослабевает
стремление к изысканию такой системы содержания, которая обеспечивала бы
хорошее здоровье и высокую продуктивность животных, а также длительное
использование маток и хряков. Последние десятилетия в практике широко
используются следующие способы содержания свиней: индивидуальное и
станково-выгульное хряков и маток; свободно-выгульное ремонтных и откормочных
свиней; летнее лагерно-пастбищное свиней на племя; групповое безвыгульное;
фиксированное свиноматок. Перечисленные способы можно подразделить на две
системы - выгульную и безвыгульную.
Характеризуя выгульную и безвыгульную системы содержания свиней
большинство авторов отдают предпочтение выгульной системе. Свиньи, пользующиеся
при выращивании выгулом (особенно лагерным содержанием), отличаются более
высокой воспроизводительной способностью, лучшим физиологическим состоянием и
более выраженной резистентностью организма. Продуктивность этих животных, как
правило, выше, чем у содержащихся при безвыгульной системе.
Установлено, что предоставление ежедневных 1,5-часовых моционов
супоросным свиноматкам в течение 3 месяцев увеличивает многоплодие на 0,2
поросенка, количество поросят в гнезде при отъеме - на 0,4 головы и живую массу
в этом возрасте - на 12,2%.
В опытах Г. Походня и др. (1997) многоплодие и крупноплодность свинок при
выгульном содержании были на 7,0 и 7,3% соответственно выше, чем при
безвыгульном содержании.
В.А. Андросов и Н.В. Шабаев (1996) установили повышение уровня
естественной резистентности и иммунобиологической реактивности организма
поросят при содержании их в летних лагерях по сравнению с выращиванием в
помещениях.
Свиньи современных пород и типов отличаются генетически обусловленной
высокой продуктивностью, но в то же время это является причиной их
исключительно высокой чувствительности к воздействию неблагоприятных факторов
внешней среды. Промышленная технология, независимо от размеров фермы,
предполагает высокую скученность поголовья в ограниченном пространстве,
безвыгульное содержание и интенсивное использование животных. При этом их
организм испытывает большие функциональные нагрузки, изменяются его адаптивные
реакции на внешние раздражители. В результате нарушается физиологическое
состояние организма, снижается продуктивность, чаще проявляются заболевания,
обусловленные снижением естественной резистентности. Поэтому большое значение
приобретает учет факторов внешней среды, которые окружают животных и влияют на
организм в целом.
Влияние микроклимата на животный организм складывается из совокупного
действия физических, химических и биологических факторов: температуры,
влажности, движения и химического состава воздуха, наличия в нем пыли,
микроорганизмов, грибов, яиц гельминтов, а также тех или иных ядовитых газов.
Наиболее важным, определяющим фактором микроклимата животноводческих
помещений является температура воздуха.
Для каждой половозрастной группы свиней существует оптимум температурных
колебаний окружающего воздуха, при котором поддерживается на определенном
уровне температура тела и согласованно протекают все обменные процессы.
Взрослые свиньи менее требовательны к теплу, а молодые, особенно
поросята-сосуны, у которых механизм терморегуляции слабо развит, сильнее
реагируют на изменения температуры окружающей среды.
По данным И.П. Шейко и В.С. Смирнова (1997) потери поросят-сосунов от
низких температур при высокой влажности могут достигать 40-50 и даже 80%. Это
можно объяснить слабо развитой физической терморегуляцией молодняка,
отсутствием подкожного жира и волосяного покрова, вследствие чего поросята
практически не способны обеспечить надлежащую теплоизоляцию, сохранить
образующееся при обменных процессах тепло. Как указывает С.И. Плященко и В.Т.
Сидоров (1983), поддержание температуры тела у новорожденных поросят
осуществляется за счет энергии корма. Но такая терморегуляция наименее
экономична и не беспредельна. Если поросят после рождения помещают в условия с
температурой воздуха 18-200С, то температура их кожи снижается на 1,5-30С, а
при 120С - на 5-60С и восстанавливается до нормы лишь через 8-10 дней. Эти и
другие изменения, возникающие в организме, приводят к значительному ослаблению
защитных сил молодого организма, создаются благоприятные условия для возникновения
заболеваний.
Сохранность, рост и развитие поросят в значительной степени зависят от
температуры окружающей среды. По данным Е.А. Махаева (1998), оптимум
температуры для свиней должен быть следующим: при живой массе 25-45 кг - 220С,
46-85 кг - 200С, 86-120 кг - 170С. При отклонении от оптимума снижалась
переваримость питательных веществ рациона, повышалась теплопродукция и
возрастала потребность в энергии, снижалась продуктивность и увеличивались
затраты кормов на получение продукции.
Рядом авторов установлено, что понижение температуры воздуха свинарника
на 10 ниже оптимальной уменьшает среднесуточные приросты на 17,8 г, увеличивает
кормовую потребность на 0,33 г/кг массы животных при групповом содержании, и на
0,7-0,4 г/кг массы - у отдельно размещаемых животных. Кроме этого, у поросят в
условиях низкой температуры значительно ослабевает бактерицидная активность
сыворотки крови и фагоцитарная активность лейкоцитов.
Взрослые животные также реагируют на понижение температуры относительно
оптимума. С.И. Плященко и В.Т. Сидоров (1983) приводят данные, что наивысшая
оплата корма у холостых и легкосупоросных свиноматок достигается при
температуре не ниже 10-120, у подсосных свиноматок - 18-200. Снижение
температуры ниже указанных уровней повышает обмен веществ у животных, что на
10-20%, а в некоторых случаях на 50-100% увеличивает расход кормов.
Высокая температура для животных также не безразлична. По данным многих
ученых повышение температуры окружающей среды относительно оптимума
отрицательно сказывается на жизнедеятельности организма. У животных развивается
тепловое перенапряжение, сопровождающееся пониженным аппетитом, вялыми
процессами пищеварения и недостаточным использованием питательных веществ,
снижением уровня газообмена и теплопродукции. Отмечается снижение лактогенеза
(молочной продуктивности), угнетение воспроизводительных функций. По данным W.H. Close (1996), при
длительном воздействии высокой температуры на свиноматок перед опоросом могут
происходить выкидыши.
На теплорегуляцию организма, и прежде всего на теплоотдачу, оказывает
влияние влажность воздуха. Высокая влажность воздуха, особенно в сочетании с
низкой температурой, очень вредна для животных. При этом увеличивается
теплопроводность и теплоемкость воздуха, что приводит к большой потере тепла
животными. Холодный и влажный воздух вызывает затруднение дыхания, ухудшение
аппетита, ослабление пищеварения, снижение упитанности и продуктивности
животных, что ведет к лишним затратам кормов. Зимой, при содержании животных в
неблагоустроенных сырых помещениях появляются простудные заболевания.
Особенно неблагоприятно отражается высокая влажность воздуха на молодых,
ослабленных и больных животных. При относительной влажности 90-100% и
температуре 0-70С у поросят-сосунов задерживается формирование физиологических
функций.
В случае повышенной влажности в сочетании с высокой температурой
происходит задержка тепла в организме животного, тормозится обмен веществ,
появляется вялость, снижается продуктивность и устойчивость к различным
заболеваниям, увеличивается число случаев желудочно-кишечных заболеваний,
особенно у молодняка.
Установлено, что в помещениях, где содержатся подсосные свиноматки,
поросята-отъемыши и ремонтный молодняк, относительная влажность воздуха не
должна превышать 70%, а в помещениях для холостых и супоросных свиноматок,
откормочного поголовья - 75%.
Повышение влажности на каждый процент выше 85% снижает прирост живой
массы тела на 2,2-2,5%.
Неблагоприятно воздействует на организм и сухой воздух при повышенной
температуре. Он способствует высыханию кожи животных и слизистых оболочек, что
повышает их ранимость и увеличивает проницаемость для микроорганизмов.
Кроме этого, Y.C. Chung and others (1996) отмечают влияние пониженной влажности воздуха
в сочетании с повышенной температурой на увеличение образования в помещении
таких ядовитых газов как аммиак и сероводород.
При рассмотрении значимости температурно-влажностного режима необходимо
учитывать также влияние скорости движения воздуха. Скорость движения воздуха
относится к числу факторов, способных значительно повысить или снизить
воздействие температуры, влажности и других условий окружающей среды.
Большая подвижность воздуха, особенно при низких температурах, вызывает
резкое увеличение теплоотдачи, охлаждение поверхности тела, повышение обмена
веществ и, следовательно, неоправданную трату кормов на производство
дополнительного количества тепла.
I. Vrbanac, R. Yammine (1994)
установили высокую корреляцию между скоростью движения воздуха и падежом
поросят-отъемышей от желудочно-кишечных заболеваний.
Большое зоогигиеническое значение имеет газовый состав воздуха.
В закрытых животноводческих помещениях газовый состав воздуха может
значительно отличаться от атмосферного. В воздухе помещений обычно содержится
больше углекислого газа, водяных паров, нередко обнаруживают аммиак,
сероводород, а также газообразные продукты гниения и брожения органических
веществ. Количество кислорода может быть меньше, чем в атмосферном воздухе.
Большой вред животным наносят высокие концентрации в воздухе помещений
аммиака и сероводорода. Основным источником их образования являются
разлагающиеся органические вещества, содержащие азот и серу. Аммиак нарушает
барьерную функцию легких, замедляет активность и парализует деятельность
мерцательного эпителия, вызывает гемолитическую анемию, снижает гуморальный и
клеточный неспецифический иммунитет.
Нормами технологического проектирования допускается следующая
концентрация вредных газов: аммиака - 0,02 мг/л, сероводорода - 0,015 мг/л и
углекислого газа - 0,25%.
Воздух имеет в своем составе взвешенные механические частицы - пыль и
микроорганизмы. Воздушная пыль загрязняет кожу, раздражает слизистые оболочки
глаз, носа и верхних дыхательных путей, способствуя этим внедрению
микроорганизмов, в том числе возбудителей инфекционных заболеваний.
По данным С.И. Плященко (1980), уже при концентрации пыли во вдыхаемом
воздухе 0,6-6 мг/м3 у животных наблюдается ряд отклонений в физиологических
процессах.
От содержания в воздухе пыли и влаги находится в прямой зависимости
содержание в нем микроорганизмов. Воздух закрытых помещений имеет более
благоприятные условия для накопления и сохранения микроорганизмов, особенно,
если не соблюдаются санитарно-гигиенические нормы содержания животных.
По данным ряда авторов, в свинарниках содержится до 100 тыс. микробов в 1
м3 воздуха. Ф.Г. Торпаков (1980) утверждает, что в свиноводческих помещениях с
застойным нагретым воздухом в 1 м3 насчитывается до 1 млн. микроорганизмов, а в
помещениях с хорошим воздухообменом (35 м3/ч на 1 ц массы тела свиней) микробов
в 3-5 раз меньше.
Одним из условий возникновения неудовлетворительного микроклимата
является аэростаз, или застой воздуха. Аэростазы формируются при нарушениях
воздухообмена внутри животноводческих помещений, характеризуются ухудшением
практически всех микроклиматических показателей и являются одной из причин
массовых заболеваний молодняка.
Застойный (аэростазный) воздух помещений снижает естественную
резистентность организма и эффективность применения лекарственных препаратов.
Одним из важнейших факторов внешней среды, влияющих на организм животных,
в том числе и на его защитные механизмы, является кормление. При этом особое
значение приобретает тип и уровень кормления, соотношение отдельных кормов в
рационе, сбалансированность рационов по питательным и другим веществам.
Свиньи - многоплодные и интенсивно растущие животные. Это определяет
высокую напряженность физиологических процессов в их организме. Они хорошо
используют корма как растительного, так и животного происхождения. Однако из-за
недостаточного уровня синтеза микробного белка и витаминов группы В в
желудочно-кишечном тракте свиньи, по сравнению со жвачными, предъявляют более
высокие требования к аминокислотному составу рационов и обеспеченности их
витаминами и минеральными веществами.
В зависимости от соотношения в рационах концентрированных и сочных кормов
выделяют три основных типа кормления: концентратно-картофельный,
концентратно-корнеплодный и концентратный. Тип кормления определяется
конкретными природными и экономическими условиями хозяйствования. Независимо от
типа кормления зерновые корма являются преобладающими в рационах свиней,
особенно хряков и молодняка. Полностью удовлетворяя потребность свиней в
энергии и фосфоре, они в то же время дефицитны на 30-50% по протеину и основным
незаменимым аминокислотам, витаминам, особенно жирорастворимым, и кальцию.
Главным дополнением к зерну злаковых при балансировании рационов по
протеину являются зернобобовые. Они обогащают зерновой рацион белком и набором
аминокислот. Также источником полноценного протеина, а еще витаминов,
микроэлементов и клетчатки является травяная мука. Большое значение она имеет в
комбикормах для свиноматок: супоросным маткам травяная мука полезна как
средство профилактики ожирения, лактирующим - для предупреждения запоров, являющихся
основной причиной синдрома ММА (мастит-метрит-агалактия).
Не следует забывать, что как одностороннее концентратное, так и
одностороннее бесконцентратное или малоконцентратное кормление приводит к
отрицательным последствиям. В условиях крупного производства отсутствие в
рационах зеленых, сочных и грубых кормов вызывает нарушения минерального и
витаминного питания, снижает воспроизводительные способности свиноматок и
хряков, отрицательно влияет на развитие молодняка.
Большое значение для животного организма имеет уровень кормления. Так,
А.Ф. Кузнецов и Е.С. Сафронов (1977) установили, что обильное кормление
свиноматок не способствует повышению их иммунобиологической реактивности, в то
время как нормированное кормление активизирует адаптивно-защитную реакцию.
Кроме общего уровня питания, большое влияние на организм животного
оказывают компоненты рациона. Наиболее важным элементом питания являются
протеиновые соединения - белки и небелковые азотистые вещества. Они играют
главную роль в обмене веществ, являются регуляторами нормального состояния
организма. Также они несут структурные функции, входя в состав антител и
антитоксинов, ферментов, гормонов, участвуют в транспорте липидно-минеральных
соединений, витаминов и др.
Исследованиями С.И. Плященко и др. (1979) установлено, что белковый
недокорм способствует снижению уровня естественной резистентности.
Качество белка, определяемое количеством и соотношением в нем
аминокислот, и в первую очередь незаменимых, имеет важнейшее значение для
продуктивности животных. Дефицит одной или нескольких аминокислот вызывает
резкое снижение аппетита и, как следствие этого, депрессию роста.
Особая роль в кормлении животных отводится витаминам. Многочисленными
исследованиями установлено, что отсутствие или недостаток витаминов всегда
сопровождается нарушениями обмена веществ в организме, которые проявляются
задержкой роста и развития молодняка, снижением репродуктивных способностей
взрослого поголовья, ухудшением питательной ценности продуктов животноводства,
а также понижением естественной резистентности организма животных во все
возрастные периоды.
В условиях интенсивного промышленного производства свинины потребность
свиней в витаминах резко возрастает, и удовлетворить ее можно только за счет
применения витаминных препаратов и кормов.
О незаменимости витаминов в жизнедеятельности животных говорят В.А.
Алексеев и Л.В. Пыркина (1997), В.А. Медведский (1997, 1998), А.В. Соляник и
Т.В. Соляник (1998), В.Ф. Филенко с соавт. (1998), А.Г. Шипицин и Н.Ю. Басова
(1999), J. Prej. and others (1993), Schweigert Florian J. (1995) и многие другие исследователи.
Огромное значение для организма свиней имеют минеральные вещества, хотя
они и не представляют энергетической ценности. Объясняется это той большой
ролью, которую минеральные вещества играют во всех процессах обмена,
происходящих в организме. В опытах В.А. Кокорева и др. (1991, 1993, 1996, 1999)
оптимизация минерального питания свиней в соответствии с установленными нормами
повышала в организме азотистый, углеводный и жировой обмен, что обеспечивало
стабильный и интенсивный рост молодняка (578-608 г в сутки), высокую
скороспелость (191-203 дня), улучшало мясосальные качества и способствовало
рациональному использованию кормов. О положительном влиянии макро- и
микроэлементов на продуктивность и естественную резистентность указывается
также в работах Ф.Я. Беренштейна (1958), Я.М. Берзиня (1965), И.А. Буралги
(1974), В.Ф. Лемеша (1965), В.И. Сапего (1985) и многих других ученых.
Таким образом, приведенные данные свидетельствуют, что факторы внешней
среды, в частности содержание, кормление и микроклимат оказывают большое
воздействие на организм свиней. При интенсивном выращивании организм свиней
всецело зависит от факторов, обусловленных конструкцией свинарников,
микроклиматом в них, условий кормления и эксплуатации. Концентрация большого
количества свиней на одном предприятии, безвыгульное содержание, однообразный
концентратный тип кормления, ранний отъем поросят, частые перемещения с
перегруппировкой животных, интенсивное их использование, комплексная
механизация и автоматизация производственных процессов и другие факторы - все
это может привести и нередко приводит к ухудшению продуктивного здоровья
животных и, в конечном счете, к появлению различных болезней. Поэтому особое
внимание необходимо уделять комплексному анализу факторов внешней среды,
которые постоянно воздействуют на организм животных. Предупреждение
отрицательного влияния указанных факторов является важным моментом в увеличении
продуктивности сельскохозяйственных животных.
Физиологическое значение минеральных веществ в организме свиней
Для поддержания жизни, роста и проявления максимальной генетически
обусловленной продуктивности животные должны получать все незаменимые
питательные и другие биологически активные вещества в определенных количествах
и соотношениях.
Функция макро- и микроэлементов в организме свиней разнообразна и важна в
биохимии питания животных. Наряду со специфическими функциями большое значение
минеральные вещества имеют в поддержании осмотического давления, буферной
емкости жидкостей и тканей организма, нервного и мышечного возбуждения,
регуляции каталитических процессов, проявлении иммунобиологической реактивности
организма. Недостаток минеральных веществ в рационе отрицательно сказывается на
степени минерализации скелета у свиней, их здоровье, продуктивности,
продолжительности жизни, функциях воспроизводства.
В настоящее время при балансировании рационов учитывают следующие
жизненно необходимые для свиней макроэлементы: кальций, фосфор, калий, натрий,
хлор, магний, серу.
В организм животных кальций поступает с кормом и минеральными добавками в
виде солей, содержится он в основном в костях (97-99%) в виде фосфорнокислых и
углекислых солей, и только в ядрах клеток данный элемент связан с органическими
веществами. Богаты кальцием нервные клетки мозга, желез, особенно молочной.
Соли кальция в плазме крови способствуют образованию фибрина и фибриногена, от
чего зависит свертываемость крови. Кальций активизирует фермент
протромбокиназу, под действием которой протромбин превращается в активный
тромбин, а также ферменты липазу, лецитиназу, актомиозинаденозинтрифосфотазу,
фосфоглюкомутазу. Кальций участвует в регулировании мышечной и нервной
деятельности, стабилизирует трипсин и тормозит функции энолазы и др.
Ионы кальция укрепляют защитные функции организма, понижая клеточную
проницаемость и повышая фагоцитарную активность лейкоцитов. Bruckner J. and
others (1958), а также Fife E.H. and Muschel J.H. (1961) установили
положительное влияние кальция на активность комплемента.
Многочисленные исследователи отмечают положительное влияние кальция на
продуктивность животных. Использование минеральной подкормки животным,
обогащенной дикальцийфосфатом, способствует повышению переваримости питательных
веществ корма, использованию организмом азота, лучшему росту животных.
Нарушение фосфорно-кальциевого обмена отрицательно сказывается на росте
подсвинков, оплодотворяемости, плодовитости, молочности маток и качестве семени
производителей.
Воздействие кальция на организм зависит от его уровня в рационе. Большие
дозы отрицательно влияют на продуктивность животных. Так, установлено
отставание в росте 6-8-недельных поросят при повышенных дозах кальция в
рационе. Физиологическими опытами Концевенко В.В. (1998) выявлено, что при
повышении содержании кальция в рационе до 2% снижаются коэффициенты
переваримости сухого вещества корма на 3%, сырого протеина на 3,5%, что
безусловно вызывает снижение прироста живой массы. Надева Р. и Кънев Д. (1998)
отмечают, что в период откорма свиней увеличение уровня кальция и фосфора в 2
раза сказалось на снижении продуктивности (на 8%) и оплаты корма (на 5-8%).
Обмен кальция взаимосвязан с обменом других минеральных веществ: фосфора,
магния, цинка, калия и железа.
Фосфор входит в состав опорной ткани, сложных белков, жиров и углеводов.
Соединения, содержащие фосфор, активизируют ферментативные процессы, участвуют
в окислительном фосфорилировании, входят в состав ряда ферментов. С участием
фосфора протекает более десяти различных функций организма. Фосфор содержится в
каждой живой клетке. Он связан с межуточным обменом углеводов, сокращением мышц
и процессами выведения кальция из организма. Фосфор - активный катализатор и
стимулятор эффективного использования корма. Он участвует во всасывании,
транспортировке и в обмене органических питательных веществ в организме, а
также в делении клеток и в ростовых процессах, в значительной степени влияет на
качество мяса.
Многочисленные опыты подтверждают исключительно важную роль фосфора в
усвоении азотистых веществ. Добавка в дефицитные по фосфору рационы животных
минеральных фосфатов неизменно повышает использование азота, в зависимости от
условий опыта, от 5 до 23%.
Недостаток фосфора в рационах животных вызывает у них ухудшение общего
состояния, извращенный аппетит и костные заболевания (рахит, остеомаляция). При
хроническом недостатке фосфора наблюдается нарушение обмена веществ, снижение
продуктивности, плодовитости, увеличение прохолостов. Фосфорное голодание
обычно завершается снижением способности организма усваивать корм,
деминерализацией. При этом ухудшаются упитанность и здоровье животных. Обмен
энергии, лежащий в основе жизнедеятельности организма животного, невозможен без
участия фосфорной кислоты.
Важным свойством фосфора является влияние его на переваримость некоторых
веществ. Добавки минерального фосфора к рационам для животных на откорме
повышают коэффициент переваримости сырого жира, сырой клетчатки, кальция,
фосфора и магния.
О положительном влиянии фосфора на продуктивность свиней говорят
исследования Бажова Г. с соавт. (2000), Водолазского М.Г. с соавт. (1997).
Скармливание свиноматкам фосфатов в последние недели супоросности отражается на
их крупноплодности, росте и сохранности полученного приплода, на общем
состоянии свиноматок после отъема от них поросят.
В то же время большое увеличение содержания фосфора в рационе животных
может отрицательно сказаться на использовании других элементов. Так, при
увеличении содержания фосфора в рационе молодняка на 20% сверх нормы отмечена
тенденция к некоторому снижению степени использования кальция, а также к
снижению использования самого фосфора. Избыток его вызывает снижение содержания
кальция непосредственно в скелете.
Калий в основном сконцентрирован в клетках (97-98%), причем наиболшее его
количество находится в мышцах (особенно сердца), тканях мозга и эритроцитов
крови. Он принимает активное участие в регулировании осмотического давления в
жидкостях организма, в возбуждении нервной и мышечной тканей, нормализует
сердечную деятельность, улучшает переваримость и обмен питательных веществ.
Калий действует как активатор многих ферментов (пируваткиназы, фруктокиназы,
фосфофруктокиназы); является одним из главных катионов клеточной среды,
поддерживающих кислотно-щелочное равновесие в организме животных; снижает
проницаемость кровеносных сосудов.
Ряд ученых выявили участие калия в усилении неспецифической
резистентности и активации белкового обмена в организме животных, повышении их
продуктивности.
При недостатке калия в организме замедляется рост, ухудшается аппетит,
наблюдается атаксия, нарушение сердечной деятельности, общая слабость, судороги
и паралич.
Избыток калия тормозит процессы биохимического синтеза, а также уменьшает
число сердечных сокращений, вызывая так называемое “калиевое торможение”.
Длительное избыточное потребление калия нарушает также воспроизводительную
функцию и вызывает нарушение обмена магния, особенно при недостатке натрия.
Ведущая роль в поддержании осмотического давления в клеточных жидкостях и
кислотно-щелочного равновесия организма принадлежит натрию. Совместно с калием
он поддерживает в норме деятельность сердечной мышцы, участвует в процессах
нервно-мышечной возбудимости. Ферментативные процессы в митохондриях и ядре
могут происходить только при наличии натрия. Ионы натрия активизируют амилазу,
фруктокиназу, холинэстеразу и тормозят действие фосфорилазы. Участвуя в
энергетическом обмене, он стимулирует его, и оказывает адаптогенное влияние на
организм поросят при технологическом стрессе.
При содержании натрия в рационе супоросных маток в количестве 17,7 г и
соотношении его к калию, равном 0,62, наблюдается увеличение количества поросят
при рождении на 21,96 %.
Недостаток натрия приводит к снижению буферности крови и тормозит
окислительные процессы. При потреблении животными кормов, бедных натрием, у них
наблюдают извращение аппетита, торможение роста (у молодняка), снижение
жирности молока, нарушение воспроизводительной функции. Недостаток натрия может
быть вызван избытком калия в рационах.
Недостаток натрия в кормах может быть восполнен дополнительным введением
в рацион поваренной соли. При этом, однако, необходимо помнить, что избыток
натрия сказывается на организме также вредно, как и недостаток. В случае
острого отравления у свиней отмечают сильную жажду, частые мочеиспускания,
рвоту, цианоз слизистых оболочек, нарушение дыхания; возможен смертельный
исход.
Магний в значительном количестве (более 60%) находится в костях и зубах,
содержится также в мягких тканях и клеточной жидкости.
Он активизирует почти все ферменты, переносящие фосфатные группы в обменных
реакциях; способствует поддержанию нормального кислотно-щелочного равновесия и
осмотического давления в жидкостях и тканях; обеспечивает функциональную
способность нервно-мышечного аппарата; участвует в терморегуляции; активизирует
работу кишечника, поджелудочной железы и усиливает действие трипсина; участвует
в синтезе белков и других веществ.
Магний входит в комплекс миозина и АТФ. От него в значительной степени
зависит энергетический метаболизм в клетках.
О положительном влиянии магния на продуктивность животных сообщают многие
исследователи. Отмечено, что добавки соли магния стимулируют не только
поедаемость кормов, но и значительно снижают падеж поросят (0,34 % против 2,40
% в контроле).
Установлено, что гуморальная защита организма зависит от состояния
минерального обмена, она напрямую зависит от содержания в сыворотке ионов
магния. Ионы магния активизируют действие пропердина, увеличивают активность
комплемента, также участвующих в формировании естественной резистентности
организма животных. Соли магния, кроме того, обладают антагонистическим
действием по отношению к бактериальным токсинам.
Недостаток магния приводит к нарушению обмена веществ в организме и
довольно часто проявляется в виде гипомагниемии, вызывающей высокую смертность
среди животных. Дефицит магния у свиней напоминает низкокальциевую тетанию:
наблюдаются повышенная возбудимость нервной системы, шаткая походка, атаксия и
тетанические судороги.
Магний является антагонистом кальция, фосфора и марганца, поэтому избыток
магния сказывается отрицательно на усвоении этих элементов, увеличивя
потребность в них.
В организме животных (клетках и жидкостях) хлор находится в виде солей
натрия, калия, кальция и магния, а также в ионизированной форме. Вместе с
натрием и калием он принимает участие в регулировании кислотно-щелочного
равновесия и осмотического давления, играет важную роль в желудочной секреции,
являясь составной частью соляной кислоты, которая необходима для активации ряда
ферментов.
Недостаточность хлора в обычных условиях маловероятна, так как
потребность в нем животных значительно меньше, чем в натрии. Явления хлорной
недостаточности были получены лишь в опытах на цыплятах при скармливании им
синтетического, практически лишенного хлора рациона. Недостаточность
проявлялась в торможении роста, снижении электролитов в плазме, дегидратации
крови и параличах, приводящих к гибели.
В организме животных сера оказывает прямое действие на образование
серосодержащих аминокислот - метионина, цистина, цистеина и др. Она входит в
состав витаминов (битина и тиамина) и гормона инсулина. Атомы серы -
физиологически активные элементы в молекуле протеина, они стабилизируют
структуру белка.
Сера улучшает переваривание целлюлозы и поддерживает биосинтез витаминов
группы В.
При недостатке серы в рационах у животных отмечают потерю аппетита,
слезотечение, слюнотечение, общую слабость, нередко гибель.
Избыток в рационе неорганической серы у поросят приводит к задержке
роста, рахиту, гастроэнтеритам; у взрослых животных избыток серы легко
выводится через почки.
Введение в рацион подсвинков порошковой серы в количестве 0,1г на 1 кг
живой массы в течение 2-х месяцев способствовало увеличению приростов живой
массы на 3,79 - 7,89 кг в опытных группах по сравнению с контрольной. При этом
отмечалось улучшение состояния гематологических показателей и улучшение
качества мяса.
К микроэлементам относятся железо, кобальт, цинк, марганец, йод, медь,
фтор и др. Они содержатся в организме от сотых до миллионных долей процента.
Железо - составная часть белков, среди которых наиболее важными являются
гемоглобин крови (60-70%), миоглобин мышечной ткани (3-5%), трансферин плазмы
крови, ферритин и некоторые ферменты (каталаза, пероксидаза, цитохромоксидаза и
др.). В тканях легких атом железа в молекуле гемоглобина способен мобильно
связывать кислород и, таким образом, удовлетворять потребность в нем организма.
Железо играет большую роль в процессах тканевого дыхания и питания, способствуя
тем самым увеличению живой массы и сохранности молодняка.
Недостаток железа в кормах приводит к развитию алиментарной анемии,
которая часто отмечается у поросят. Симптомы анемии: исхудание, задержка роста
и пониженная иммунобиологическая реактивность, извращение аппетита, поносы,
бедный волосяной покров, снижение репродуктивных функций (у взрослых).
Многочисленные исследования указывают на успешное применение различных
железосодержащих добавок с целью предотвращения недостаточночти этого элемента
в организме животных. Введение в рацион свиноматок сульфата железа совместно с
витамином С приводит к повышению их уровня в крови и значительно улучшает
состояние поросят-сосунов. П.Я. Конопелько, Л.М. Морозова (1989) предлагают для
предупреждения анемии поросят применять препарат ферроглюкин.
Обогащение белково-витаминных добавок глицеро-фосфатом железа в сочетании
с витамином Е и ферментным препаратом протосубтилином Г3 положительно влияет на
продуктивность доращиваемых и откармливаемых свиней.
По данным И.А. Буралга (1974), ежедневное введение в рацион молодняка
свиней сернокислого железа привело к увеличению среднесуточного прироста на
9,4%. К концу опыта живая масса одной головы была выше, чем в контроле, на 5,5
кг. При этом отмечено увеличение на 8,4% общего белка сыворотки крови.
О.Г. Олейников и соавт. (1995) указывает на то, что внутримышечное
введение свиноматкам железосодержащего препарата глептоферрона за три недели до
предполагаемого опороса повышает продуктивность свиноматок. Полученные поросята
были более жизнеспособны, реже подвергались заболевания дыхательной и
пищеварительной систем, снижался отход молодняка, а прирост живой массы был
выше, чем у поросят контрольной группы.
Ряд авторов предлагает совместно с ферроглюкином вводить животным смесь
микроэлементов - медь, кобальт, цинк и селен. Введение этих микроэлементов
стимулировало гемо- и эритропоэз, способствовало увеличению в сыворотке крови
общего белка, бетта- и гаммаглобулинов, нуклеиновых кислот, повышало защитные
функции и резистентность организма.
Следует отметить, что содержание этого элемента в организме выше
физиологической нормы является негативным фактором. При избытке железа
ухудшается усвоение фосфора, меди, уменьшается отложение витамина А в печени
молодняка, замедляется рост. Высокие дозы железа токсичны. Так, содержание в
корме лишь 0,4% железа вызывает интоксикацию организма. Наиболее остро избыток
железа влияет на организм поросят.
Медь участвует в гемопоэзе и способствует образованию в крови гемоглобина
в присутствии железа, влияет на поступление железа в костный мозг. Микроэлемент
необходим для нормального развития костной ткани, поскольку стимулирует
образование оссеина и нормализует отложение кальция и фосфора. Медь в
значительной степени влияет на обмен в организме углеводов, липидов, белков и
минеральных веществ. Активизирует синтез йодированных соединений щитовидной железы,
влияет на активность половых гормонов, обмен витаминов и функциональное
состояние эндокринной и нервной системы.
Увеличение содержания меди в сыворотке крови сопровождается повышением
выработки антител.
М.Г. Коломийцева, Р.Г. Габавич (1970) приводят данные по состоянию
иммунобиологической реактивности организма в зависимости от содержания меди в
рационе. Авторы показали, что биотические дозы меди повышают фагоцитарную
активность лейкоцитов, бактерицидность и лизоцимную активность сыворотки крови.
Было также установлено, что иммунобиологическая реактивность находится в
определенной зависимости от концентрации и соотношения меди и марганца в
рационе.
Влияние меди как стимулятора роста зависит от уровня ее в корме, возраста
животных, а также содержания других минеральных веществ в рационе.
На усвоение меди оказывает влияние присутствие в рационе других макро- и
микроэлементов, причем большинство из них тормозит этот процесс. Молибден,
сера, цинк и железо являются антагонистами и снижают в процессе обмена веществ
адсорбцию и содержание меди в организме.
При недостатке меди у животных снижается аппетит, уменьшается
продолжительность жизни эритроцитов, постепенно замедляется рост, происходит
депигментация волосяного покрова, ослабляется костяк, снижается подвижность
суставов, наблюдаются поносы. В тяжелых случаях возникает анемия. Недостаток
меди может сопровождаться нарушением развития головного мозга, а также
прерыванием беременности у самок.
Дефицит меди ухудшает переваримость питательных веществ (сырого протеина,
жира), снижает использование азота и кальция. Оптимальный же уровень меди в
рационах позволяет повысить интенсивность роста свиней на 5,1-7,5%, коэффициент
мясности, калорийность мякотной части туш и снижает затраты кормов на
7,5-16,6%.
Имеются материалы, что использование несколько повышенных доз меди
свиньям (125-375 мг/кг) способствует улучшению использования корма на 10-15%,
повышает отложение кальция.
В.В. Николенко (1982) указывает на возможность увеличения количества
лейкоцитов и эритроцитов у молодняка при введении в рацион подкормки сульфата
меди совместно с хлоридом кобальта.
Высокие дозы меди в рационах оказывают токсическое действие. Так,
хроническая интоксикация приводит к некрозу клеток печени и гемолизу
эритроцитов. У животных наблюдают желтушность, апатию, отмечают повышенное
сердцебиение; животные испытывают жажду, подолгу лежат; смерть наступает в
результате печеночной комы. Малые дозы меди легко выводятся из организма.
Цинк обладает весьма широким спектром физиологического действия, участвует
в процессах дыхания, служит катализатором в окислительно-восстановительных
процессах, повышает активность витаминов и усиливает фагоцитоз. Входит в состав
многих ферментов, например карбоангидразы, которая обусловливает расщепление
угольной кислоты на двуокись углерода и воду; глутаминодегидрогеназы,
окисляющей глутаминовую кислоту; щелочной фосфотазы почек. Цинк влияет на
процессы оплодотворения и размножения, активизирует гормон инсулин, кишечную
фосфатазу, регулирует обмен кальция и меди, ослабляет гипертензивное действие
адреналина.
Положительное влияние цинка на переваримость питательных веществ,
нормализацию обменных процессов в организме и прирост живой массы отмечено В.А.
Кокоревым с соавт. (1991), Н.А. Давыдовым с соавт. (1995), A. Korniewicz (1992).
Недостаток цинка приводит к паракератозу. У животных отмечают дерматиты,
отсутствие аппетита, рвоту, поносы, замедленный рост, хромоту, дефекты
конечностей, нарушение воспроизводительной функции, особенно у самцов,
нарушение жирового и углеводного обмена. Приплод рождается слабым, с плохим
костяком, нередко гибнет.
Фактором, способствующим развитию цинковой недостаточности, является
содержание в рационе кальция при низком содержании цинка. При высоком
содержании кальция в кормах рекомендуется добавлять к рациону карбонат цинка.
Имеются материалы, свидетельствующие о влиянии недостатка цинка на
показатели естественной защиты организма. По этим данным в результате цинковой
недостаточности у молодняка свиней происходит снижение количества эритроцитов на
24,6%, бактерицидной активности - на 33,6%, активности лизоцима сыворотки крови
- на 40,6%. Дефицит цинка в рационе поросят способствует снижению содержания
лейкоцитов в крови.
Применение цинка, как в отдельности, так и в сочетании с другими
микроэлементами в кормлении животных, дает положительный результат. Добавление
тяжело супоросным свиноматкам цинка стимулирует внутриутробный рост поросят,
повышает многоплодие и крупноплодность свиноматок. Xu Zirong, Wang Minqi (1999)
установили положительное влияние различных форм цинка на среднесуточные
приросты и пищеварение поросят (увеличение переваримости сухих веществ и сырого
протеина, уменьшение диареи).
Различные препараты цинка в сочетании с другими микроэлементами
используются для стимуляции защитных сил организма. Установлено, что применение
0,4 мг/кг селенита натрия и 0,05 г сернокислого цинка на 100 кг живой массы
тела поросят активизирует некоторые клеточные и гуморальные факторы
неспецифической резистентности организма животных.
Добавки меди, железа и цинка способствуют увеличению активности щелочной
фосфотазы, концентрации гемоглобина в крови и стабильности мембраны
эритроцитов, увеличению прироста живой массы и превращения корма.
При избытке цинка наблюдают плохую поедаемость кормов, поносы, анемию, резкое
снижение приростов.
Хотя кобальт и мало распространен в природе, но является жизненно
необходимым элементом для животных. Он активизирует гидролитические ферменты,
увеличивает синтез нуклеиновых кислот и мышечных белков; в присутствии железа и
меди повышает активность кроветворной системы, входит в состав витамина В12.
Кобальт - важный возбудитель процессов образования эритроцитов, непосредственно
оказывает влияние на кроветворные функции костного мозга, ускоряет синтез
гемоглобина, повышает устойчивость организма к заболеваниям.
Кобальт и витамин В12 способствуют повышению среднесуточных приростов,
увеличивают продуктивность, положительно влияют на ассимиляцию азотистых
веществ, активируют использование аминокислот. Ионы кобальта принимают участие
в реакциях гликолиза и цикла трикарбоновых кислот, активируют ферменты
дипептидазу и фосфотазу, аргиназу, каталазу, но тормозят активность уреазы и
цитохромоксидазы.
Кобальт, являясь катализатором окислительно-восстановительных реакций,
повышает всасывание железа в кишечнике и использование его при образовании
гемоглобина. Так, А.В. Бушов с соавт. (1997) указывает на почти двухкратное
увеличение количества гемоглобина в крови поросят при введении им 2 мл/гол
ферродекса и 1 мл/гол тирозина кобальта.
При недостатке кобальта возникает акобальтоз (сухотка), гиповитаминоз или
авитаминоз В12. У животных, больных акобальтозом, наблюдается угнетенное
состояние, потеря или извращение аппетита, лизуха, малокровие (снижение
гемоглобина и числа эритроцитов), прогрессирующее истощение, приводящее к
гибели.
Рядом иcследователей установлено, что
дополнительное введение в рацион ремонтных свинок и поросят-сосунов хлористого
кобальта и витаминов положительно влияет на естественные защитные силы
организма, рост и развитие животных. Это можно объяснить усилением биосинтеза
витамина В12 в организме животных и активирующим действием его на обмен веществ
организма.
Скармливание животным кобальтосодержащего витамина В12 вызывает резкое
увеличение концентрации лизоцима уже через день после его введения.
Введение в рацион хлористого кобальта положительно влияет на
продуктивность поросят, значительно увеличивает прирост и выход мясной
продукции, стимулирует накопление в организме свиней витаминов А, С, Е,
повышает количество эритроцитов и лейкоцитов в крови.
У всех животных резкий избыток кобальта вызывает полицитемию крови и
гиперплазию костного мозга, потерю аппетита, нарушение роста, животные
становятся вялыми, снижаются приросты.
Марганец усиливает окислительные процессы, потребление кислорода, синтез
гликогена, повышает выделение с мочой общего азота и мочевины, уменьшает
выделение хлоридов. Он необходим для кроветворения (особенно в сочетании с
железом, медью и кобальтом), принимает участие в тканевом дыхании, оказывает
влияние на обмен углеводов и жиров, пролонгирует действие витаминов (А, Е, В1,
В6, С и др.), активизирует целый ряд ферментов (тиаминазу, аргиназу,
дезоксирибонуклеазу и др.).
Марганец оказывает благоприятное влияние на рост и развитие молодняка,
воздействует на половое развитие и размножение животных. Недостаток его в
рационах может привести к атрофии семенников у самцов, а у самок - к снижению
деятельности яичников.
При дефиците данного элемента молодняк рождается слабый или мертвый, у
поросят проявляются структурные изменения в костях. Скармливание свиньям
кормов, дефицитных по марганцу, приводит к жировой инфильтрации печени и
повышенному отложению жира в туше. Недостаток марганца в тканях приводит к
образованию и накоплению в организме перекисных соединений.
Избыток марганца в кормах снижает использование йода, серы, меди,
фосфора, кальция и хлора в организме животных, оказывает отрицательное влияние
на рост, снижает уровень гемоглобина.
С профилактической целью рекомендуется вводить поросятам с кормом
сернокислый марганец в дозе 1,2-2,0 мг, а свиньям на откорме - 3-6 мг на голову
в сутки.
Йод входит в структуру гормона щитовидной железы - тероксина и
обусловливает его физиологическую активность в регуляции процессов белкового,
липидного, углеводного, водного и минерального обмена.
Йод способствует повышению продуктивности, улучшению состояния здоровья,
стимулирует рост и развитие молодняка. Дополнительное введение йода в рацион
способствует достоверному повышению переваримости сухого вещества, сырого
протеина, сырого жира у подсвинков 130-, 190-, 250-дневного возраста; оказывает
положительное действие на отложение азота и макроэлементов; ведет к увеличению
среднесуточных приростов живой массы свиней, снижению затрат корма на единицу
прироста и повышению убойного выхода.
Однократная имплантация йода в дозе 8 мг/гол в форме таблеток йодида
калия за 20 дней до опороса позволяет достоверно увеличить содержание в крови
эритроцитов и гемоглобина у супоросных и подсосных свиноматок, что указывает на
его положительное влияние на эритроцитарный гомеостаз.
Недостаток йода является основной причиной появления зоба, обусловливает
снижение окислительных процессов и азотистого обмена в организме, усиливает
отложение жира, снижает активность многих ферментов. У взрослых животных
нарушаются половые циклы, отмечаются перегулы, выкидыши. Новорожденные
недоразвиты, отечны, поросята без щетины. В тяжелых случаях рождается мертвый
приплод.
Повышение дозы йода в рационе вызывает снижение прироста массы, ухудшение
аппетита и признаки токсикоза у животных. Очень высокие дозы ведут к
прекращению деятельности яичников и перерывам лактации.
В организме животных фтор в основном депонируется в костной ткани и
зубах. Он участвует в фосфорно-кальциевом обмене при росте скелета, в
нормальных дозах предотвращает кариес зубов и способствует заживлению костных
переломов. Оказывает влияние на обмен йода в организме.
При недостатке фтора у животных наблюдают потерю аппетита, деформацию
скелета и зубов. Однако большой вред животноводству наносит не дефицит фтора, а
его избыток, который приводит к нарушению обменных процессов в костной ткани,
деформации и утолщению суставов и сужению костно-мозговых каналов. Также
избыточное поступление фтора в организм обусловливает отставание в росте и
снижение продуктивности маток.
Селен обладает высокой биохимической активностью, усиливает обмен
веществ. Установлено его влияние на белковый обмен (в частности, на обмен
серосодержащих аминокислот), на обмен жиров и углеводов, участие в регуляции
многих ферментативных реакций. Селен воздействует на процессы тканевого
дыхания, повышает иммунологическую реактивность организма, улучшает
плодовитость маток, регулирует усвоение и расход витаминов А, С, Е и К в
организме.
Добавление к рациону свиней солей селена способствует эффективному приросту
массы животных, улучшению роста и развития молодняка, более полному
использованию питательных веществ кормов; в значительной степени влияет на
воспроизводительные функции свиноматок; оказывает положительное влияние на
отложение у свиней азота, кальция и фосфора, увеличение в крови содержания
эритроцитов, гемоглобина и белка.
Скармливание поросятам селенопирана повышало активность оксидазных
ферментных систем нейтрофилов и, следовательно, резистентность организма
молодняка.
Недостаток селена в рационах вызывает беломышечную болезнь, дистрофию
печени, дегенерацию яичников, маститы, анемии, гемолиз эритроцитов, является
одной из главных причин появления мертворожденных и слабых поросят.
Избыток селена приводит к тяжелым эндемическим заболеваниям (алколоз,
щелочная болезнь). У животных наблюдают анемию, общее истощение, нарушение
сердечной деятельности, функции печени, частичную деформацию суставов,
нарушение функции нервной системы, параличи.
Молибден выполняет активную роль в окислительных процессах, так как
входит в состав флавиновых ферментов - ксантиноксидазы и альдегидоксидазы. При
этом он ускоряет азотистый обмен в организме. Элемент также влияет на
углеводный и липидный обмен, иммунобиологическую реактивность организма,
прирост массы, способствует повышению содержания общего белка в сыворотке крови
и λ-глобулинов в ней.
Увеличение количества нейтрофильных лейкоцитов при скармливании молибдена
вместе с сульфатом меди свидетельствует о его способности повышать защитные
функции организма.
По данным В.А. Кокорева, А.М. Гурьянова (1996), балансирование рационов
по молибдену способствует нормализации пищеварительных процессов в организме,
улучшению мясосальных качеств и более интенсивному росту молодняка.
Нарушений, вызванных недостатком молибдена у свиней, не обнаружено. У
жвачных симптомы недостаточности сходны с таковыми для меди.
При содержании в рационе более 1 мг молибдена на 1 кг корма отмечают
признаки отравления. Его избыток приводит к патологическим явлениям, в основном
у жвачных: истощению, поносам, ломкости костей, ослаблению сперматогенеза,
анемии, нарушению обмена серы, фосфора, меди, хлора, кобальта, марганца и
пуриновых оснований, гибели животных.
Таким образом, участвуя во всех обменных процессах организма, входя в
состав всех органов и тканей макро- и микроэлементы в значительной степени
стимулируют естественные защитные силы организма, продуктивность животных,
повышают качество животноводческой продукции.
Поэтому является важным обеспечение оптимального поступления минеральных
веществ в организм животных.
Использование источников минерального питания в кормлении свиней
Основным источником минеральных веществ для сельскохозяйственных животных
являются корма растительного происхождения.
На химический состав растительных кормов влияют климатические и погодные
условия, особенности химического состава растений, а также агротехнические
факторы, такие как количество, качество и сроки внесения в почву удобрений,
сроки уборки, технология заготовки и хранения кормов. Ценность кормов по
минеральным веществам также зависит от фазы развития растения. Так, к концу
вегетации количество фосфора, калия, хлора и меди в них, как правило,
уменьшается, а кальция увеличивается.
Дефицит в минеральных элементах устраняется путем добавления последних к
удобрениям. Например, при внесении в почву препаратов меди, повышается ее
содержание в растениях, за счет азотных удобрений снижается количество в почве
цинка, кобальта, но повышается - кальция, магния и натрия в травах, под
влиянием фосфатных удобрений в кормах возрастает содержание фосфора, но
уменьшается - микроэлементов. При внесении только калийных удобрений повышается
количество калия в растениях, но одновременно тормозится усвоение ими кальция и
магния.
Следовательно, в результате внесения удобрения дефицит в минеральных
веществах устраняется лишь частично.
При неполном минеральном составе кормов их недостаток восполняется за
счет минеральных добавок. В настоящее время известно довольно много подкормок,
содержащих различное количество макро- и микроэлементов с неодинаковой степенью
их усвояемости организмом животных.
Минеральные подкормки вырабатывает химическая промышленность, они
достаточно дорогие, поэтому очень важно изыскивать и использовать местные
естественные месторождения и источники минерального сырья.
Большинство кормов содержит слишком мало натрия, зачастую не хватает
фосфора. Недостаток в рационе натрия восполняется в основном поваренной солью.
Обычная кормовая поваренная соль в зависимости от сортности содержит 95-98%
хлористого натрия, в котором находится 39% натрия, 57 % хлора, 0,35-0,65%
кальция, 0,05-0,25% магния и в малом количестве другие минеральные и
органические примеси. Избыточное и недостаточное скармливание поваренной соли
отрицательно сказывается на состоянии животного. Свиньям поваренную соль дают в
молотом виде в смеси с концентратами, в составе комбикормов.
Сравнительно дешевым источником поваренной соли в условиях Беларуси могут
выступать галитовые отходы (галиты) 4-го РУ ПО “Беларуськалий”. О
целесообразности замены обычной поваренной соли в рационах животных солью
галитовых отходов говорят В.М. Голушко и др.(1982).
Хлористый калий (хлорид калия) - кристаллический порошок хорошо
растворимый в воде, не отличающийся по цвету и вкусу от поваренной соли, но
слегка с раздражающим действием кристаллов при прикосновении к языку. В этом
препарате содержится около 52% калия и 48% хлора, он используется для
балансирования рационов по калию и является обязательным компонентом заменителя
молока для поросят раннего отъема.
Среди фосфорно-кальциевых подкормок известен монокальцийфосфат кормовой,
кальций фосфорнокислый однозамещенный - мелкогранулированный серный порошок,
растворимый в воде, содержит 17,6% кальция, около 24% фосфора и не более 0,3%
фтора. Усвояемость фосфора организмом животных достигает 90,7%.
Трикальцийфосфат - трехзамещенный фосфат кальция - нерастворимый в воде
аморфный порошок, содержащий 14,5% фосфора и 32% кальция.
Обесфторенный фосфат кормовой- также нерастворимый в воде аморфный
порошок, получаемый из фосфоритов и апатитов. В его составе содержится 16%
фосфора, 36% кальция, не более 0,2% фтора и имеются примеси железа, магния,
кремния и других элементов.
Динатрийфосфат - двуокисный фосфорнокислый натрий - светлый
мелкокристаллический продукт без запаха, содержит 8,6% фосфора и 13,3% натрия.
Скармливается животным, в рационе которых имеется избыток кальция и недостаток
фосфора.
При избытке в рационах кальция и недостатке фосфора, скармливании больших
количеств корнеклубнеплодов для выравнивания оптимального соотношения между
калием и натрием в качестве натриевой кормовой добавки используют двуосновной
фосфорнокислый натрий.
Из многих кальциевых подкормок животным скармливают мел, известняк,
известковые туфы, гарныш, мергель, лимнокальций, травертины и другие природные
средства.
Кормовой мел представлен углекислым кальцием, используется как добавка в
тонкоизмельченном виде при условии, чтобы в нем содержалось не более 1%
примесей и фтора. В нем содержится в среднем 37% кальция, 0,18% фосфора, около
0,5% калия, 0,3% натрия и не более 5% кремния и других элементов. Мел вводят в
рационы поросят в количестве до 1%, а взрослых свиней - до 2% от сухого
вещества корма.
Широко применяются известняки, в состав которых входят: кальций (34-37%),
магний и кремний (по 1-3%), а также в незначительном количестве железо, сера и
фосфор. Используют их после предварительного измельчения в таком же, примерно,
количестве, как и мел. Известковый туф нередко состоит из почти чистого
углекислого кальция и не требует размола. В доломитовом известняке содержится
3-7% магния. Его используют в молотом виде, когда в рационе недостает магния.
Травертин по химическому составу близок к известковым туфам, но отличается
большим содержанием микроэлементов и применяется в качестве кальциевой
подкормки. Источниками кальция являются также ракушечная и мидиевая мука, мука
из створок мелких моллюсков.
В ряде случаев дефицит кальция в рационах животных восполняется за счет
использования гипса, который в чистом виде представляет сернокислый кальций. Он
является производным серной кислоты и может в определенной степени восполнять
недостаток в кормах серы.
В качестве серо-кальциевой добавки в рационах животных может выступать
фосфогипс (отходы производства фосфорных удобрений), получаемый при переработке
апатитов на Гомельском химическом заводе. Его химический состав следующий: вода
- 6%, кальций - 33, сера - 22,8, фосфор - 1,13%. В фосфогипсе присутствуют в
незначительных количествах калий, натрий, алюминий, железо, барий,
редкоземельные элементы. Отрицательной стороной фосфогипса является наличие в
нем до 0,3% фтора, который в большом количестве оказывает отрицательное влияние
на организм животных. Для устранения этого недостатка необходимо производить
обесфторивание его. По данным В.К. Пестиса и В.М. Голушко (1982, 1984) использование
фосфогипса при выращивании и откорме свиней повышает прирост живой массы на 4%,
снижает затраты кормов и себестоимость единицы прироста.
Чтобы восполнить недостаток серы и магния используется сульфат магния -
белый кристаллический порошок, содержащий 13% серы и 10% магния. Можно
скармливать глауберову соль, или сернокислый натрий - белый порошок, содержащий
10% серы. Этот препарат способствует лучшему усвоению азотистых веществ рациона
и повышает содержание жира в молоке.
В рационах с недостатком магния в качестве добавки используется жженая
магнезия, или окись магния - нерастворимый в воде аморфный порошок, содержит
60% магния, около 0,02% хлора, 0,15% кальция и 0,015% железа.
А.М. Венедиктов с соавт. (1992) в качестве источника магния предлагает
использовать карбонат магния основной (белая магнезия, углекислый магний) -
рыхло-аморфный порошок, малорастворимый в воде, содержащий около 23-25% магния.
Зачастую при выращивании поросят следует давать железистые подкормки, в
частности железный купорос - железо сернокислое закисное. По внешнему виду
препарат голубовато-зеленого цвета кристаллы или порошок растворимый в воде.
Его водный раствор используется как микродобавка в рационах молодняка с целью
профилактики анемии.
В качестве источника меди выступает медный купорос (сернокислая медь). Он
представляет собой прозрачные кристаллы или порошок синего цвета. Содержит
около 12% серы и 5% меди, используется как добавка в дефицитных по меди
рационах и при лечении алиментарной анемии.
Хлористый кобальт - по внешнему виду порошок вишневого цвета, растворим в
воде, и используется как подкормка при нехватке кобальта в кормах.
Цинк сернокислый, или сульфат цинка, цинковый купорос - бесцветные
кристаллы, хорошо растворимые в воде. Количество цинка около 22%, серы - 11%. Его
можно применять для всех видов сельскохозяйственных животных с целью
профилактики цинковой недостаточности, возможна замена окисью цинка или
хлористым цинком.
Сернокислый марганец - кубические кристаллы розово-сероватого цвета,
растворимые в воде; содержит около 23% марганца.
Йодистый калий - бесцветные кубические кристаллы, растворимые в воде,
имеющие солено-горький вкус с нейтральной реакцией. Используется как
микродобавка для всех видов животных, но его нельзя применять одновременно с
солями меди.
Селеновые подкормки применяются при беломышечных заболеваниях и как
профилактическое средство в виде селенита натрия.
Наряду с использованием макро- и микроэлементов промышленного
производства в последнее время привлекают внимание исследования и производственное
использование местных естественных месторождений минерального сырья.
Цеолиты - водные алюмосиликаты. Они характеризуются высокой ионообменной
способностью, обратимыми процессами дегидратации, высокой способностью
поглощать газы, главным образом двуокись серы, аммиак, газообразный хлор,
хлористый водород, высокой термостабильностью к агрессивным средам,
доступностью и дешевизной.
По химическому составу цеолитовые руды подразделяются на
натриево-кальциевые, кальциевые, калиевые, калиево-натриевые, калиево-кальциевые.
Химический состав цеолитов различных месторождений отличается. Например, 1 кг
цеолитсодержащего трепела Костюковичского месторождения Могилевской области
содержит: железа - 4518 мг, меди - 6,36, цинка - 25,5, марганца - 58,9, калия -
3,03 г, натрия - 0,51, кальция - 0,78, фосфора - 0,09, магния - 1,67 г.
Включение цеолитов в рационы свиней способствует росту естественных
защитных сил их организма, улучшению состояния многих гематологических
показателей, нормализует обменные процессы в организме животных; положительно
влияет на энергию роста молодняка, сохранность поголовья и оплату корма,
повышает воспроизводительные функции взрослого поголовья.
В зависимости от состава цеолитов и индивидуальных особенностей животных
в рацион вводят до 6% цеолитов на 1 кг сухого вещества корма.
В Беларуси имеются значительные запасы верхового сфагнового торфа,
степень разложения которого не превышает 20%. Кроме органических веществ,
преобладающих в его составе, в нем содержатся соли кальция, фосфора, железа, а
также микроэлементы: цинк, медь, молибден, марганец, кобальт. Это позволяет
рассматривать сфагновый торф как источник минерального питания животных. Торф в
чистом виде свиньи почти не едят, поэтому его необходимо смешивать с кормами.
Вводить в рацион свиней торфа рекомендуется до 10% от сухого вещества рациона.
В последнее время заметно возрос интерес к кормовым добавкам на основе
глины.
Микроминеральный состав бентонитовой глины, в частности, содержание в ней
кальция, марганца, цинка, меди и др., явился фактором, обеспечивающим
укрепление и лучшую минерализацию костяка свиней, способствующим более
интенсивному росту молодняка.
Л.А. Матюшевский и Н.Н. Коновалов (1983) указывают, что введение
свиноматкам 1,5% бентонита к весу корма на протяжении периода супоросности
способствует повышению показателей естественной резистентности у полученных
поросят, увеличению на 12,1% их продуктивности, снижению в 2,4 раза
заболеваемости и в 2,8 раза падежа.
Пикумин - минеральная добавка в корма для свиней. Это побочный продукт
(отходы) при производстве керамзита, представляющий собой обожженный при
высокой температуре порошок коричневого цвета, не слеживающийся при хранении,
технологичный при производстве кормосмесей и комбикормов. Он имеется на всех
заводах по термической обработке глины. Пикумин близок по химическому составу к
обычной глине, но не содержит органических веществ, а влажность составляет
всего 2-4%. В 1 кг добавки содержится кальция - 13,30 г, фосфора - 0,11 г,
магния - 13,85 г, натрия - 4,05 г, калия - 7,98 г, железа - 19,73 г, меди -
5,50 г, цинка - 72,7 мг, марганца - 215,05 мг и ряд других минеральных веществ.
При использовании пикумина в кормлении молодняка свиней В.А. Медведским
(1997) установлены оптимальные дозы его, которые оказывают стимулирующее
действие на отдельные показатели продуктивности и резистентности организма.
Так, у поросят-отъемышей, получавших добавку пикумина, среднесуточные приросты
живой массы были на 10,5% выше, чем у контрольных.
Исследованиями М.В. Рубиной (2001) установлено положительное сочетание
пикумина и цеолитсодержащего трепела в рационах откармливаемых свиней.
Вермикулит - это природный минерал, относящийся к алюмосиликатам, имеющий
слоистую структуру с размером частиц от 0,5 до 5 мм. После термической
обработки при температуре 1000-11000С приобретает свойства, позволяющие
использовать его в корм свиньям.
Введение в рацион супоросным и подсосным свиноматкам, поросятам-сосунам,
поросятам на доращивании и молодняку на откорме вермикулита в количестве 3% к
массе основного рациона оказывает положительное влияние на физиологическое
состояние свиней: улучшаются морфологические, биохимические и иммунологические
показатели крови. У маток, получавших вермикулит, на 1,5-10% выше
крупноплодность, на 2,6-6% - молочность, выше сохранность поросят на 5,5%, ниже
заболеваемость желудочно-кишечными заболеваниями на 15,0-20,3%. У поросят на
доращивании среднесуточный прирост живой массы выше на 4,3%, а сохранность - на
10%. У свиней на откорме вермикулит способствует приросту массы тела на 3,3%, улучшению
химического и минерального состава мяса.
В качестве минеральной добавки В.А. Медведский (1997) предлагает
использовать морскую соль. В ней содержится, мг/кг: кальций - 102,5, магний -
517,5, медь - 3,0, железо - 47,7, кобальт - 75,0, марганец - 67,5, цинк - 14,5,
калий 25,0 и другие элементы. Введение в рационы супоросных и подсосных
свиноматок 0,3-0,5% морской соли к массе сухого вещества корма способствовало
повышению уровня минерального обмена у животных, повышению их продуктивности,
гуморальных факторов защиты их организма. Поросята от свиноматок, получавших
соль, лучше развивались и росли, меньше подвергались желудочно-кишечным
заболеваниям.
Как минеральная добавка, а также источник витаминов, аминокислот,
углеводов, ферментов и других веществ выступает сапропель. Сапропель, или ил
(озерная грязь) - это донное отложение пресноводных озер. В сухом веществе
сапропеля, в зависимости от места залегания, содержится, %: органическое
вещество - до 26, зола (в основном карбонат кальция) - до 42, протеин - 1-6,
фосфор - до 0,2. В 1 кг высушенного сапропеля содержится (мг): марганца - до
90, цинка - до 60, молибдена - до 47, брома - до 58, бора - до 37, меди - до
26, кобальта - до 12,8, йода - до 6-7.
Включение в рацион сапропеля повышает его полноценность, способствует
увеличению скорости роста животных, улучшению их мясных качеств и значительной
экономии корма. В исследованиях И.Г. Елисеева и А.М. Карабанова (1982) при
скармливании добавки наблюдалось увеличение количества в крови эритроцитов,
гемоглобина и общего белка, повышение обмена кальция и фосфора. Е.Р. Сапалева
(1982) указывает на влияние сапропеля на воспроизводительные способности
свиноматок: многоплодие повышается на 11,0-14,1%, крупноплодность - на
3,2-10,1%, молочность - на 9,6 - 15,1%.
Применение добавок на основе сапропеля позволяет балансировать корма по
питательным веществам, витаминам, макро- и микроэлементам, аминокислотам.
Минеральной подкормкой служит также древесная зола. В ее состав входят
кальций, натрий, калий, магний, фосфор, а также микроэлементы. Химический
состав древесной золы зависит от вида растительности. Например, березовая зола
содержит (%): кальция - 22,4, марганца - 4,7, фосфора - 2,2, железа - 0,8,
цинка - 0,5, кобальта - 0,3, меди - 0,04, йода - 0,03.
Древесный уголь - также в основном кальциевая подкормка. Ценен тем, что
способствует улучшению пищеварения у животных, так как имеет свойство подавлять
нежелательные процессы брожения в кишечнике. Поросята древесный уголь охотно
поедают в чистом виде. В.Г. Мантикова (1995) указывает, что включение
древесного угля в качестве минеральной подкормки в рационы сосунов снижает
заболеваемость желудочно-кишечного тракта, увеличивает сохранность поросят на
7,8% и суточный прирост на голову на 6,5%.
Таким образом, минеральные вещества являются жизненно необходимыми
элементами, они оказывают существенное влияние на продуктивность, рост и
развитие животных, участвуют в формировании естественных защитных сил
организма. Однако мало изученной остается эффективность использования различных
источников макро- и микроэлементов. В животноводстве остро стоит необходимость
поиска и изучения новых дешевых источников минерального питания.
Особый интерес для животноводства Беларуси представляет местное сырье -
минеральная добавка пикумин. Однако влияние пикумина на организм свиней, на
естественные защитные силы и продуктивные качества как взрослого поголовья
свиней, так и получаемого от них приплода изучено недостаточно. Богатый
минеральный состав пикумина, его доступность и относительная дешевизна создают
предпосылку для более глубокого изучения этой добавки.
Использование пикумина свиноматкам в течение первого периода супоросности
(до 84-го дня) и лактации
Зоотехнический анализ рационов кормления свиноматок показал значительные
отклонения от нормативов по многим минеральным веществам. В рационах супоросных
и подсосных свиноматок при превышении крахмала, калия, переваримого протеина,
наблюдался дефицит кальция, магния, железа, меди, цинка, кобальта.
Для покрытия дефицита в отдельных макро- и микроэлементах к основному
рациону дополнительно вводили минеральную добавку пикумин, получаемый при
изготовлении керамзита. Это продукт обжига глины при температуре около 1,9
тыс.0С, легко смешиваемый с кормом порошок коричневого цвета. Минеральный
состав пикумина приведен в табл. 1.
Таблица 1
Состав
пикумина
|
Показатели
|
Ед. измерения
|
В 1 кг пикумина содержится
|
|
Макроэлементы:
|
|
Кальций
|
г
|
13,30
|
|
Фосфор
|
г
|
0,11
|
|
Магний
|
г
|
13,85
|
|
Натрий
|
г
|
4,05
|
|
Калий
|
г
|
7,98
|
|
Микроэлементы:
|
|
Железо
|
г
|
19,73
|
|
Медь
|
мг
|
54,97
|
|
Цинк
|
мг
|
72,70
|
|
Марганец
|
мг
|
215,05
|
Животные контрольных групп добавку не получали. Свиноматкам опытных групп
вводили в рацион 0,8, 1,2 и 1,5% пикумина в расчете на 1 кг сухого вещества
корма. При этом добавка применялась в течение первого периода супоросности (до
84-го дня) и лактации. Из четырех групп животных I-ая была контрольной, II-ой вводили в рацион 0,8% пикумина в расчете на 1 кг сухого
вещества корма, III-ей - 1,2%, IV-ой - 1,5%.
Продолжительность использования добавки составила: первый период
супоросности (84 дня) + подсосный период (45 дней)-всего 129 дней.
Естественные защитные силы и показатели крови свиноматок
Естественная резистентность организма свиноматок. Состояние
неспецифической резистентности организма животных обеспечивается гуморальными и
клеточными факторами. Из гуморальных факторов защиты организма наиболее
изученными остаются лизоцим, бактерицидная активность, естественные антитела,
белки крови и др. При определении гуморальных факторов защиты наиболее часто
исследуется бактерицидная активность сыворотки крови. Она дает возможность
судить о суммарных гуморальных защитных механизмах организма животных.
При изучении влияния минеральной добавки пикумин на свиноматок была
поставлена задача определить иммуностимулирующее действие ее на их организм и
полученный приплод.
До применения добавки бактерицидная активность сыворотки крови маток
находилась в пределах 64,14±2,294 - 70,26±2,421%
(рис. 1). К 4-му дню после опороса отмечено увеличение этого показателя, причем
в III группе, в которой свиноматки получали
1,2% пикумина, оно было достоверным по сравнению с контрольной. К концу
подсосного периода бактерицидная активность сыворотки крови снижалась у
свиноматок всех групп. Однако во II-й
группе этот показатель был выше, чем в контрольной на 2,9%, в III и IV - на 8,9%.

Рис.
1. Бактерицидная активность сыворотки крови, %
В
качестве гуморального фактора защиты организма также выступает лизоцим
(мурамидаза) - универсальный защитный фермент. Изучение лизоцимной активности
сыворотки крови не выявило достоверных различий между группами (рис. 2). Однако
необходимо отметить, что во все периоды исследований у свиноматок III и
IV групп она была выше на 3,2 - 8,3% по сравнению с
контролем.
Из
клеточных факторов защиты организма изучали способность лейкоцитов захватывать
проникающие в тело животных микроорганизмы, с последующим их перевариванием.
Фагоцитарная
активность лейкоцитов возрастала у всех подопытных животных в течение первого
периода супоросности и снижалась после опороса (рис. 3). На 4-й день после
опороса свиноматки II группы превосходили по этому показателю контрольных
на 3,9%, III - на 3,2%, IV - на 3,4%. К концу подсосного
периода отмечено достоверное увеличение фагоцитарной активности лейкоцитов у
свиноматок III и IV групп по сравнению с контрольными.
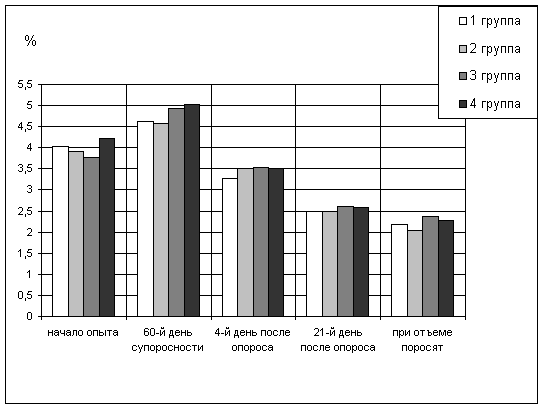
Рис.
2. Лизоцимная активность сыворотки крови, %
Результаты
исследования сиаловых кислот показали, что введение в рацион минеральной
добавки пикумин не вызвало аллергической реакции в организме свиноматок (рис.
4). Их концентрация в начале опыта составляла 24,13±0,76 - 27,35±0,74 ед.опт.пл. В течение первого периода
супоросности, а также подсосного периода, концентрация сиаловых кислот в крови
подопытных животных снижалась. При этом достоверных различий между группами по
этому показателю установлено не было.

Рис. 3. Фагоцитарная активность лейкоцитов, %

Рис.
4. Сиаловые кислоты, ед. опт. плотн.
В
связи с тем, что состояние естественной резистентности оценивалось по
нескольким иммунобиологическим показателям, выражающимся различными единицами
измерения, был определен комплексный показатель неспецифической резистентности.
Динамика комплексного показателя естественной резистентности свиноматок
показана на рис. 5.
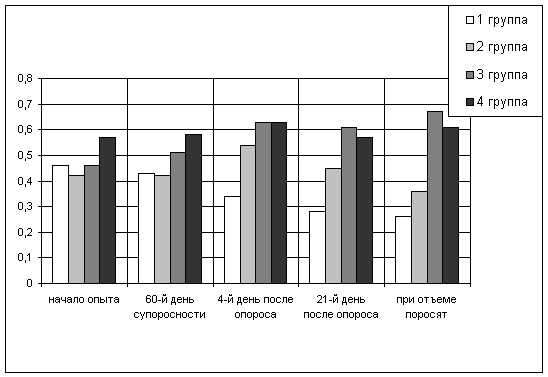
Рис.
5. Комплексный показатель естественной резистентности свиноматок
свиноматка пикумин продуктивность минеральный
Определение
комплексного показателя неспецифической резистентности организма свиноматок
позволило более объективно оценить влияние минеральной добавки пикумин на
показатели клеточной и гуморальной защиты. Рассчеты показали, что уже к 60-му
дню супоросности животные III и IV групп отличались повышенной
естественной резистентностью организма. Более существенные различия по
комплексному показателю отмечены к 4-му дню после опороса, когда величина этого
показателя во II группе была в 1,59 раза, в III и в IV - в 1,85 раза выше, чем в контрольной группе. На 21-й день после
опороса величина комплексного показателя в I группе составила 0,28, во II - 0,45, в III -
0,61, в IV - 0,57, а к отъему - соответственно
0,26, 0,36, 0,67 и 0,61.
Таким образом, использование минеральной добавки пикумин в кормлении
свиноматок способствует росту естественных защитных сил их организма.
Морфологические и биохимические показатели крови свиноматок. В крови
животных изучалось количество лейкоцитов и эритроцитов. Содержание этих
компонентов зависит от многих факторов, среди которых большое значение имеют
кормление, обеспеченность организма биологически-активными веществами. Добавка
подопытным животным к рациону пикумина в наших опытах определенным образом
отразилась на морфологических показателях крови (табл. 2).
При постановке на опыт количество лейкоцитов в крови свиноматок
находилось в пределах 9,95±0,622 - 11,16±0,702
Х109/л. В ходе опыта содержание лейкоцитов во всех группах возрастало. При этом
достоверных различий между группами по этому показателю не установлено. Вместе
с тем, на 60-й день супоросности свиноматки опытных групп по содержанию в крови
лейкоцитов превосходили контрольных на 3,8-6,7%, на 4-й день после опороса - на
5,1-9,3%, к отъему - на 5,5-7,0%.
Содержание эритроцитов на протяжении опыта было в пределах
физиологической нормы, без существенных колебаний.
Таблица 2
Морфологические показатели крови свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Лейкоциты, 109/л
|
При постановке на опыт
|
|
10,21±0,520
|
10,07±0,279
|
11,16±0,702
|
9,95±0,622
|
|
На 60-й день супоросности
|
|
11,38±0,472
|
12,12±0,379
|
12,14±0,526
|
11,81±0,716
|
|
На 4-й день после опороса
|
|
11,83±0,472
|
12,43±0,421
|
12,93±0,674
|
12,50±1,071
|
|
На 21-й день после опороса
|
|
12,62±0,429
|
12,55±0,435
|
13,09±0,590
|
12,90±0,772
|
|
При отъеме поросят
|
|
13,12±0,386
|
13,11±0,407
|
14,04±0,515
|
13,84±0,716
|
|
Эритроциты, 1012/л
|
При постановке на опыт
|
|
5,81±0,096
|
6,24±0,073
|
5,95±0,110
|
5,60±0,086
|
|
На 60-й день супоросности
|
|
5,62±0,062
|
5,97±0,126*
|
5,69±0,086
|
5,56±0,079
|
|
На 4-й день после опороса
|
|
5,70±0,060
|
5,84±0,096
|
5,71±0,096
|
5,63±0,122
|
|
На 21-й день после опороса
|
|
5,44±0,060
|
5,74±0,096*
|
5,57±0,060
|
5,61±0,096
|
|
При отъеме поросят
|
|
5,41±0,071
|
5,73±0,129
|
5,61±0,060
|
5,65±0,126
|
Примечание:
здесь и в последующих таблицах: * - Р<0,05;** - Р<0,01; *** - Р<0,001;
Результаты исследований белкового обмена представлены в табл. 3.
Таблица 3
Протеинограмма свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
При постановке на опыт
|
80,16±2,697
|
84,74±1,923
|
78,32±3,775
|
79,55±2,197
|
|
Альбумины, %
|
37,18±3,565
|
36,12±2,760
|
38,06±2,604
|
37,02±2,994
|
|
α-глобулины,%
|
16,22±1,264
|
17,84±1,837
|
14,27±0,455
|
18,29±1,657
|
|
β-глобулины,%
|
20,38±1,660
|
18,24±1,734
|
18,20±0,680
|
19,03±1,345
|
|
γ-глобулины,%
|
26,22±3,053
|
27,80±2,004
|
29,47±2,393
|
25,64±1,732
|
|
На 60-й день супоросности
|
|
Общий белок, г/л
|
76,42±2,953
|
78,40±2,337
|
77,95±1,923
|
79,12±3,236
|
|
Альбумины, %
|
42,07±2,118
|
41,86±3,874
|
44,16±1,654
|
43,59±3,129
|
|
α-глобулины,%
|
14,12±1,320
|
15,97±0,626
|
12,09±0,603
|
13,44±1,978
|
|
β-глобулины,%
|
21,72±1,811
|
18,10±1,003
|
23,16±1,225
|
23,93±1,694
|
|
γ-глобулины,%
|
22,08±1,942
|
24,07±2,247
|
20,58±1,159
|
19,05±1,657
|
|
На 4-й день после опороса
|
|
Общий белок, г/л
|
81,09±2,266
|
83,36±1,888
|
84,96±2,966
|
86,19±2,689
|
|
Альбумины, %
|
38,46±3,594
|
38,14±3,966
|
40,12±2,902
|
39,37±4,235
|
|
α-глобулины,%
|
16,86±1,159
|
17,13±0,483
|
15,24±1,517
|
16,29±2,017
|
|
β-глобулины,%
|
28,66±2,852
|
28,72±2,365
|
30,62±2,087
|
29,33±2,345
|
|
γ-глобулины,%
|
16,02±2,365
|
16,01±1,118
|
14,02±1,910
|
15,01±2,097
|
|
На 21-й день после опороса
|
|
Общий белок, г/л
|
74,37±1,785
|
74,64±3,146
|
77,57±1,579
|
81,60±1,978*
|
|
Альбумины, %
|
40,64±2,341
|
41,07±2,986
|
42,24±2,541
|
43,26±2,966
|
|
α-глобулины,%
|
17,86±1,588
|
17,23±0,924
|
16,10±1,172
|
15,32±1,500
|
|
β-глобулины,%
|
20,65±0,910
|
22,71±1,447
|
20,84±1,298
|
21,95±2,478
|
|
γ-глобулины,%
|
20,84±2,768
|
19,00±1,475
|
20,83±1,809
|
19,48±2,868
|
|
При отъеме поросят
|
|
Общий белок, г/л
|
55,42±1,571
|
56,24±2,876
|
59,78±1,236
|
63,44±1,528**
|
|
Альбумины, %
|
39,34±3,562
|
38,13±3,573
|
40,22±1,624
|
42,58±2,556
|
|
α-глобулины,%
|
19,78±0,998
|
20,80±2,399
|
22,41±1,212
|
21,27±2,079
|
|
β-глобулины,%
|
23,06±1,307
|
22,94±1,073
|
16,36±1,144
|
14,07±1,138
|
|
γ-глобулины,%
|
17,83±2,444
|
18,09±0,722
|
21,01±2,794
|
22,09±0,663
|
|
|
|
|
|
|
Количество общего белка в сыворотке крови в начале опыта находилось в
пределах 78,31±3,719 - 84,76±1,946 г/л. К 60-му дню супоросности
содержание общего белка снижалось во всех группах, но в опытных снижение этого
показателя было менее значительным. К опоросу количество общего белка в
сыворотке крови повышалось. В это время свиноматки опытных групп превосходили
контрольных на 2,7-6,4%. К отъему было отмечено достоверное увеличение
концентрации общего белка в сыворотке крови у свиноматок III и IV групп. Они превосходили контрольных соответственно на 8,0%
(Р<0,05) и 14,6% (Р<0,01).
При анализе распределения белка по фракциям установлено, что содержание
альбуминов в ходе опыта увеличивалось, при чем более существенно у животных,
получавших дополнительно пикумин. Количество α-глобулинов возрастало к концу
опыта,однако существенных различий по этому показателю установлено не было.
Также не было установлено разницы между группами по содержанию в сыворотке
крови β-глобулинов. В то же время в конце
опыта количество γ-глобулинов у свиноматок III и IV
групп было на 17,8 и 23,9% соответственно выше, чем у контрольных животных.
Свиноматки опытных групп, которым в рацион вводили минеральную добавку, к
концу опыта имели в крови больше гемоглобина. По этому показателю животные II группы превосходили контрольных на
4,8%, III - на 4,9%, а IV - на 7,6% (табл. 4).
Данные о состоянии углеводного обмена, полученные при проведении
исследований, показали, что содержание глюкозы в крови подопытных животных перед
опоросом несколько снижалось и находилось в пределах 3,42±0,112 - 3,52±0,160 ммоль/л. В конце опыта отмечено
увеличение концентрации глюкозы в крови свиноматок всех групп. Существенных
различий по этому показателю между группами выявлено не было.
Полученные данные по содержанию в крови холестерина показывают, что
введение в рацион пикумина оказало некоторое влияние на этот показатель.
Наиболее значительное повышение концентрации холестерина наблюдалось у животных
III группы. К 21-му дню после опороса оно
было достоверным.
Изменений концентрации мочевины в крови свиноматок при введении в рацион
минеральной добавки не наблюдалось. В течение всего периода исследований этот
показатель находился в пределах физиологической нормы.
Таблица 4
Биохимические показатели крови свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
1
|
2
|
3
|
4
|
5
|
|
Гемоглобин, г/л
|
При постановке на опыт
|
|
109,0±5,06
|
104,8±3,43
|
108,0±4,49
|
110,0±5,58
|
|
На 60-й день супоросности
|
|
112,0±4,29
|
118,5±6,18
|
120,4±4,29
|
117,5±7,30
|
|
На 4-й день после опороса
|
|
136,8±3,86
|
137,0±5,06
|
139,5±4,49
|
138,7±7,56
|
|
На 21-й день после опороса
|
|
112,2±4,94
|
113,5±7,30
|
115,2±4,72
|
117,5±6,74
|
|
При отъеме поросят
|
|
123,6±6,01
|
129,5±8,43
|
129,6±3,86
|
133,0±4,49
|
|
Глюкоза, ммоль/л
|
При постановке на опыт
|
|
4,05±0,104
|
3,90±0,099
|
3,84±0,129
|
3,96±0,139
|
|
На 60-й день супоросности
|
|
4,26±0,118
|
4,17±0,154
|
4,20±0,118
|
4,21±0,180
|
|
На 4-й день после опороса
|
|
3,42±0,118
|
3,54±0,154
|
3,70±0,157
|
3,49±0,118
|
|
На 21-й день после опороса
|
|
3,98±0,120
|
3,93±0,182
|
3,94±0,118
|
3,89±0,154
|
|
При отъеме поросят
|
|
4,04±0,079
|
3,91±0,129
|
4,03±0,139
|
3,93±0,104
|
|
Холестерин, г/л
|
При постановке на опыт
|
|
0,87±0,037
|
0,80±0,019
|
0,84±0,037
|
0,79±0,028
|
|
На 60-й день супоросности
|
|
0,78±0,039
|
0,77±0,037
|
0,80±0,039
|
0,81±0,051
|
|
На 4-й день после опороса
|
|
0,82±0,030
|
0,86±0,037
|
0,86±0,062
|
0,86±0,076
|
|
На 21-й день после опороса
|
|
0,72±0,019
|
0,75±0,062
|
0,78±0,028
|
0,82±0,039
|
|
При отъеме поросят
|
|
0,69±0,028
|
0,69±0,037
|
0,74±0,030
|
0,73±0,039
|
|
Мочевина, ммоль/л
|
При постановке на опыт
|
|
2,00±0,171
|
2,03±0,097
|
1,78±0,256
|
1,91±0,099
|
|
На 60-й день супоросности
|
|
1,88±0,131
|
1,90±0,084
|
1,69±0,163
|
1,89±0,129
|
|
На 4-й день после опороса
|
|
2,21±0,097
|
2,08±0,084
|
1,97±0,213
|
2,07±0,189
|
|
На 21-й день после опороса
|
|
2,06±0,097
|
1,93±0,084
|
1,91±0,195
|
1,85±0,129
|
|
При отъеме поросят
|
|
1,85±0,163
|
1,97±0,084
|
1,79±0,131
|
1,90±0,042
|
Минеральный состав крови свиноматок
Дополнительное введение в рацион свиноматок макро- и микроэлементов в
составе пикумина положительно сказалось на показателях минерального состава
крови свиноматок (табл. 5).
Таблица 5
Минеральный состав крови свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
1
|
2
|
3
|
4
|
5
|
|
Фосфор неорганический,
ммоль/л
|
При постановке на опыт
|
|
2,25±0,090
|
2,32±0,116
|
2,25±0,073
|
2,19±0,045
|
|
На 4-й день после опороса
|
|
2,81±0,082
|
2,82±0,152
|
2,99±0,093
|
3,00±0,084
|
|
При отъеме поросят
|
|
1,98±0,062
|
2,08±0,177
|
2,07±0,045
|
2,21±0,059*
|
|
Кальций, ммоль/л
|
При постановке на опыт
|
|
2,25±0,107
|
2,17±0,079
|
2,30±0,053
|
2,22±0,041
|
|
На 4-й день после опороса
|
|
2,37±0,069
|
2,38±0,107
|
2,47±0,042
|
2,48±0,105
|
|
При отъеме поросят
|
|
2,29±0,079
|
2,34±0,093
|
2,51±0,045*
|
2,51±0,070
|
|
Цинк, мкмоль/л
|
При постановке на опыт
|
|
32,93±2,441
|
30,30±1,236
|
34,91±1,947
|
32,02±1,339
|
|
На 4-й день после опороса
|
|
36,90±1,597
|
39,90±1,511
|
37,34±2,478
|
38,00±2,298
|
|
При отъеме поросят
|
|
34,40±1,601
|
36,54±2,118
|
36,72±1,573
|
36,38±1,486
|
|
Магний, ммоль/л
|
При постановке на опыт
|
|
0,96±0,101
|
0,99±0,079
|
0,92±0,062
|
1,09±0,047
|
|
На 4-й день после опороса
|
|
1,22±0,067
|
1,29±0,098
|
1,40±0,079
|
1,42±0,097
|
|
При отъеме поросят
|
|
1,10±0,090
|
1,23±0,098
|
1,36±0,067*
|
1,39±0,062*
|
|
Железо, мкмоль/л
|
При постановке на опыт
|
|
28,64±2,421
|
26,23±1,009
|
31,30±1,902
|
29,02±1,453
|
|
На 4-й день после опороса
|
|
34,76±1,676
|
36,94±1,784
|
41,05±1,893*
|
43,30±3,017*
|
|
При отъеме поросят
|
|
29,10±1,979
|
34,01±1,382
|
37,02±1,646*
|
38,33±2,174*
|
|
Натрий, ммоль/л
|
При постановке на опыт
|
|
142,2±4,33
|
139,6±2,51
|
139,8±5,06
|
142,4±3,00
|
|
148,2±3,76
|
149,4±3,26
|
154,0±5,08
|
155,8±5,92
|
|
При отъеме поросят
|
|
140,8±3,56
|
142,2±3,34
|
148,6±3,93
|
148,8±3,88
|
|
|
|
|
|
|
Содержание фосфора в крови свиноматок в начале опыта составляло 2,18±0,041 - 2,32±0,112 ммоль/л. К 4-му дню после
опороса уровень фосфора повышался у всех подопытных животных и в течение
подсосного периода наблюдался его спад. К отъему установлено увеличение его
содержания у свиноматок II и III групп на 5,1%, IV -на 11,6% (P<0,05).
Было отмечено заметное увеличение в крови опытных животных концентрации
кальция. К концу опыта количество кальция у свиноматок III и IV
группы превышало контроль на 9,6%, при этом в III группе это увеличение было достоверным.
Также изменения наблюдались по содержанию в крови цинка. Так, к 4-му дню
после опороса свиноматки опытных групп по этому показателю превосходили
контрольных на 1,2-3,0%, к отъему это превосходство составило 5,8-6,7%.
К концу опыта в крови животных III и IV группы наблюдалось также увеличение
концентрации магния. У маток этих групп содержание магния составило
соответственно 1,36±0,067
и 1,39±0,062 ммоль/л (Р<0,05), в то время
как у контрольных - 1,10±0,090
ммоль/л (на 25% меньше).
Высокое содержание в пикумине железа способствовало значительному росту
его количества и в крови свиноматок. Уже к 4-му дню после опороса у животных,
получавших минеральную добавку увеличение концентрации железа составило
6,3-24,6%, при этом в III и IV группах оно было достоверным. К
45-му дню после опороса разница с контрольной группой была 16,9-31,7%.
В течение всего периода исследований значительных колебаний концентрации
натрия в крови свиноматок отмечено не было. Его содержание было в пределах
139,6±2,51 - 155,8±5,92 ммоль/л с небольшим ростом в
опытных группах.
Таким образом, использование пикумина свиноматкам благоприятно сказалось
на клеточных и гуморальных факторах защиты их организма. Добавка не вызывала
аллергической реакции в их организме. При этом улучшались показатели
морфологического и биохимического состава крови животных, наблюдалось усиление
минерального обмена.
Продуктивность и воспроизводительные способности свиноматок
Продуктивность и воспроизводительные способности свиноматок оценивали по
следующим показателям: многоплодие, крупноплодность, молочность, масса гнезда
при рождении и отъеме, сохранность поросят, количество слабых и
мертворожденных.
Важнейшим хозяйственным признаком маток является их многоплодие. Этот
показатель выступает итогом эмбрионального развития плодов. На него влияют
условия содержания и кормления маток в период супоросности, состояние их
здоровья.
Повышение естественной резистентности организма животных, улучшение
некоторых показателей обмена скзалось на продуктивных качествах свиноматок
(табл. 6).
Не смотря на отсутствие достоверности по многоплодию свиноматок, в
опытных группах количество родившихся поросят было на 1,1-8,3% больше, по
сравнению с контрольной. При этом число слабых поросят в этих группах было
меньше на 15,7-30,6%, а мертворожденных - на 21,8-30,4%.
Результаты взвешивания приплода показали, что живая масса поросят,
полученных от свиноматок контрольной группы при рождении была 1,20±0,070 кг, а от животных опытных групп
- 1,25±0,051 - 1,31±0,043 кг, что выше контрольных на
4,2-9,2%. Масса гнезда при рождении во II группе была на 5,3%, в III - на 18,6% (Р<0,05), в IV - на 17,5% (Р<0,05) больше, чем в контроле.
Таблица 6
Показатели воспроизводительных функций свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Родилось живых поросят в
среднем на 1 матку, голов
|
9,08±0,606
|
9,18±0,599
|
9,92±0,647
|
9,83±0,462
|
|
В том числе слабых, голов
|
1,08±0,260
|
0,91±0,200
|
0,83±0,277
|
0,75±0,185
|
|
Родилось мертвых поросят в
среднем на 1 матку, голов
|
0,23±0,087
|
0,18±0,100
|
0,08±0,092
|
0,16±0,185
|
|
Масса гнезда при рождении,
кг
|
10,92±0,658
|
11,50±0,858
|
12,95±0,490*
|
12,83±0,563*
|
|
Молочность свиноматок, кг
|
47,2±3,06
|
54,7±3,20
|
58,9±3,00*
|
58,4±3,56*
|
|
Масса гнезда при отъеме, кг
|
81,1±4,79
|
87,8±7,61
|
100,3±6,12*
|
99,8±6,77*
|
|
Сохранность поросят, %
|
92,7±1,33
|
93,2±1,54
|
96,6±1,32
|
94,9±2,31
|
При определении молочности маток было установлено заметное отличие по
этому показателю у животных, получавших к рациону минеральную добавку пикумин.
Свиноматки опытных групп по молочности превосходили контрольных на 15,9-24,8%,
причем в III и IV группах это превосходство было достоверным.
К отъему в гнездах маток опытных групп сохранилось больше поросят (на
0,5-4,2%). Высокая сохранность полученного молодняка и повышенная энергия его
роста позволили получить к отъему гнезда с большей живой массой. У свиноматок II группы к 45-му дню подсосного
периода масса гнезда была на 8,3%, III группы - на 23,7%, IV -
на 23,1% (Р<0,05) больше, чем у животных контрольной группы.
Таким образом, при использовании изучаемой добавки повышались клеточные и
гуморальные факторы естественной защиты животных. Повышение иммунного статуса
подопытных свиноматок, активизация кроветворной функции их организма, усиление
некоторых обменных процессов способствовало росту продуктивных качеств животных,
улучшению их воспроизводительных способностей. При этом лучший эффект получен
при введении в рацион 1,2 и 1,5% пикумина в расчете на 1 кг сухого вещества
корма.
Энергия роста
и гематологические показатели поросят-сосунов
Применение
добавки пикумин свиноматкам в течение первого периода супоросности и лактации
определенным образом сказалось на полученном приплоде. Были изучены показатели
роста поросят, клеточной и гуморальной защиты организма, обмена веществ,
минерального состава их крови.
Энергия роста поросят-сосунов. Результаты взвешивания поросят показали
(табл. 7), что введение в рацион свиноматок минеральной добавки пикумин
повлияло на крупноплодность полученного молодняка. Живая масса поросят II группы при рождении была на 4,2%, III группы - на 9,2%, IV - на 8,3% больше, чем в контрольной
группе. Опытные поросята отличались повышенной энергией роста в течение всего
подсосного периода. Абсолютный прирост у молодняка II группы за 45 дней был больше на 4,6%, III группы - на 8,6%, IV - на 10,0%, чем в контроле.
Соответственно в опытных группах был выше и среднесуточный прирост живой массы.
Полученные данные свидетельствуют о том, что добавка к рациону маток
пикумина оказывает положительное действие на интенсивность роста
поросят-сосунов.
Таблица 7
Динамика приростов живой массы полученных поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Живая масса, кг: при
рождении
|
1,20±0,070
|
1,25±0,051
|
1,31±0,043
|
1,30±0,041
|
|
- в 21 день
|
5,63±0,190
|
6,40±0,128**
|
6,05±0,177
|
6,26±0,175*
|
|
-при отъеме (в 45 дней)
|
9,68±0,397
|
10,27±0,350
|
10,47±0,347
|
10,70±0,407
|
|
Абсолютный прирост, кг
|
8,47±0,311
|
8,86±0,364
|
9,20±0,319
|
9,32±0,321
|
|
Среднесуточный прирост, г
|
188±6,9
|
197±8,7
|
204±7,0
|
207±7,1
|
|
% к контролю
|
100,0
|
104,8
|
108,5
|
110,1
|
Естественные защитные силы организма поросят. Изучение гуморальных
факторов защиты поросят показало, что бактерицидная активность сыворотки крови
новорожденных поросят была выше у животных III и IV
групп (рис. 6). По этому показателю молодняк третьей группы превосходил
контроль на 1,5%, IV - на 1,9%. При
отъеме достоверное преимущество имели животные третьей группы.
По активности лизоцима в крови полученного приплода при рождении нами не
установлено заметных различий между группами (рис. 7). Однако на протяжении
всего подсосного периода отмечалась тенденция к повышению этого показателя у
опытных животных, а перед отъемом у поросят III и IV
групп лизоцимная активность сыворотки крови была достоверно выше, чем у
контрольных.
Исследование фагоцитарной активности лейкоцитов выявило некоторое
превосходство по этому показателю у молодняка опытных групп (рис. 8). При
рождении фагоцитоз был интенсивнее в этих группах в среднем на 1,9%. В ходе
опыта превосходство возрастало. К отъему у поросят второй группы фагоцитарная
активность лейкоцитов была выше на 1,2%, III - на 8,6%, IV -на 9,5%.

Рис.
6. Бактерицидная активность сыворотки крови, %

Рис.
7. Лизоцимная активность сыворотки крови, %

Рис. 8. Фагоцитарная активность лейкоцитов, %
Концентрация сиаловых кислот в крови поросят показала, что введение
минеральной добавки в рацион свиноматкам не вызывало существенных изменений
этого показателя в течение всего периода исследований (рис. 9).
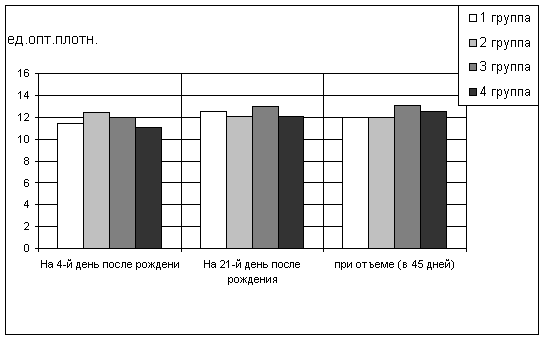
Рис.
9. Сиаловые кислоты, ед. опт. плотн.
При
определении комплексного показателя естественной резистентности организма
поросят стало очевидным, что наиболее развитыми естественные защитные силы были
у молодняка III и IV групп (рис.10).
Разница
по данному показателю между группами увеличивалась с ростом поросят. К 21-му
дню после рождения комплексный иммунологический показатель в I
группе составил 0,36, во II - 0,38, в III - 0,63, в IV -
0,63, а к 45-му дню - соответственно 0,20, 0,39, 0,68, 0,64.
Таким
образом, оптимизация минерального питания свиноматок способствует получению от
них более здорового приплода с повышенным уровнем естественных защитных сил их
организма.
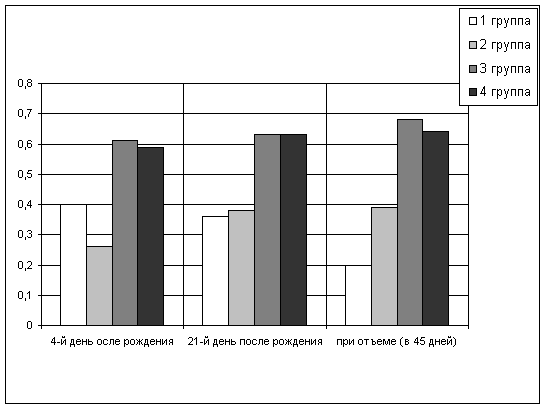
Рис.
10. Комплексный показатель естественной резистентности организма поросят
Морфологические
и биохимические показатели крови поросят. Изучение морфологических показателей
крови поросят показало, что дополнительное введение в рацион свиноматок
минеральной добавки пикумин не повлияло на концнтрацию лейкоцитов в крови
полученного от них приплода (табл. 8). Также не было установлено существенных
различий между группами по содержанию в крови эритроцитов.
Применение пикумина маткам III и IV групп вызвало некоторое увеличение
количества общего белка сыворотки крови у полученного приплода (табл. 9). В
течение опыта превосходство экспериментальных групп над контролем по этому
показателю возрастало, хотя достоверных различий не установлено. К концу опыта
у поросят II группы содержание общего белка в
сыворотке было на 5,8%, в III -
на 7,9%, IV - на 7,6% больше, чем в контрольной
группе.
Увеличение концентрации общего белка происходило главным образом за счет
роста альбуминовой и γ-глобулиновой фракций. К отъему молодняка установлено
достоверное увеличение содержания γ-глобулинов в сыворотке крови у
поросят III и IV групп.
Количество α-глобулинов в течение опыта находилось примерно на
одном уровне без видимых различий между группами. Процент β-глобулинов в течение подсосного
периода в опытных группах снижался.
Таблица 8
Морфологические показатели крови поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Лейкоциты, 109/л
|
На 4-й день после рождения
|
|
8,88±0,393
|
8,96±0,758
|
9,04±0,343
|
8,98±0,534
|
|
На 21-й день после рождения
|
|
9,00±0,300
|
8,85±0,801
|
9,13±0,463
|
9,08±0,548
|
|
При отъеме
|
|
9,28±0,322
|
9,33±0,885
|
9,30±0,354
|
9,24±0,604
|
|
Эритроциты, 1012/л
|
На 4-й день после рождения
|
|
6,02±0,143
|
6,02±0,112
|
6,15±0,109
|
6,14±0,160
|
|
На 21-й день после рождения
|
|
6,12±0,109
|
6,21±0,112
|
6,29±0,098
|
6,22±0,112
|
|
При отъеме
|
|
5,71±0,170
|
5,81±0,112
|
5,84±0,086
|
5,70±0,174
|
Содержание гемоглобина в крови молодняка к 4-му дню было в пределах 97,0±4,49 - 99,0±5,62 г/л с небольшим увеличением у
поросят III и IV групп (табл. 10). К 21-му дню наблюдался рост этого
показателя у животных всех групп без достоверных различий между ними. К отъему
количество гемоглобина в крови снижалось, однако, несмотря на отсутствие
достоверности, поросята опытных групп в это время по содержанию гемоглобина на
0,7-5,0% превосходили контрольных.
Изучение углеводного обмена выявило недостоверное увеличение содержания
глюкозы у поросят опытных групп. По этому показателю животные II группы при рождении превосходили
контрольных на 5,9%, III
группы - на 3,6%, IV - на 4,9%, а к
отъему - соответственно на 8,6, 5,1, и 7,8%.
Менее выраженная картина наблюдалась и по содержанию холестерина в крови
полученного приплода. Этот показатель во все периоды исследований у поросят III и IV групп был на 3,2-9,6% выше, чем в контроле.
По содержанию мочевины в крови полученных поросят достоверных различий не
установлено. Этот показатель во все периоды исследований находился в пределах
физиологической нормы.
Минеральный состав крови поросят-сосунов несколько отличался в опытных
группах. При рождении количество фосфора у поросят этих групп было на 1,1-5,0%
выше, чем в контроле. В течение подсосного периода концентрация фосфора в крови
животных возрастала, особенно у поросят III и IV
групп.
Таблица 9
Протеинограмма поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
На 4-й день после рождения
|
|
Общий белок, г/л
|
60,24±1,618
|
60,24±2,067
|
60,99±1,305
|
62,00±2,157
|
|
Альбумины, %
|
29,45±2,329
|
30,18±3,213
|
30,49±3,172
|
32,86±2,955
|
|
α-глобулины,%
|
21,08±1,517
|
20,87±2,197
|
19,33±0,850
|
23,33±1,143
|
|
β-глобулины,%
|
32,96±2,239
|
31,05±2,851
|
30,85±2,532
|
24,32±1,713
|
|
γ-глобулины,%
|
16,52±1,174
|
17,92±1,826
|
19,33±2,579
|
19,49±0,407
|
|
На 21-й день после рождения
|
|
Общий белок, г/л
|
60,35±1,167
|
62,32±1,708
|
62,72±1,888
|
65,12±1,798
|
|
Альбумины, %
|
32,90±2,142
|
33,41±3,792
|
35,01±3,978
|
38,77±3,480
|
|
α-глобулины,%
|
24,35±1,631
|
22,70±1,126
|
23,38±1,531
|
23,87±1,003
|
|
β-глобулины,%
|
26,17±1,685
|
25,68±1,756
|
21,53±2,278
|
17,03±1,275
|
|
γ-глобулины,%
|
16,59±1,174
|
18,22±1,104
|
20,09±1,222
|
20,33±1,360
|
|
При отъеме
|
|
Общий белок, г/л
|
52,16±1,648
|
55,20±1,348
|
56,19±1,305
|
56,16±1,169
|
|
Альбумины, %
|
40,61±2,478
|
41,56±4,225
|
42,40±2,957
|
43,33±3,972
|
|
α-глобулины,%
|
19,59±1,281
|
17,40±0,478
|
19,64±0,790
|
18,86±1,862
|
|
β-глобулины,%
|
19,39±1,492
|
20,75±1,447
|
11,07±0,551
|
9,64±0,433
|
|
γ-глобулины,%
|
20,42±0,831
|
20,30±3,253
|
26,90±1,824*
|
28,17±1,674**
|
Таблица 10
Биохимический и минеральный состав крови поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
1
|
2
|
3
|
4
|
5
|
|
Гемоглобин, г/л
|
На 4-й день после рождения
|
|
97,5±6,74
|
97,0±4,49
|
98,8±6,44
|
99,0±5,62
|
|
На 21-й день после рождения
|
|
126,4±6,01
|
125,5±4,49
|
127,0±8,99
|
129,0±4,49
|
|
При отъеме
|
|
94,8±4,72
|
95,5±7,30
|
98,4±4,72
|
99,5±8,99
|
|
Глюкоза, ммоль/л
|
На 4-й день после рождения
|
|
4,72±0,208
|
5,00±0,154
|
4,89±0,200
|
4,95±0,154
|
|
На 21-й день после рождения
|
|
5,20±0,238
|
5,32±0,180
|
5,29±0,137
|
5,32±0,154
|
|
При отъеме
|
|
4,74±0,238
|
5,14±0,258
|
4,98±0,118
|
5,14±0,208
|
|
Холестерин, г/л
|
На 4-й день после рождения
|
|
1,25±0,037
|
1,21±0,037
|
1,31±0,039
|
1,30±0,025
|
|
На 21-й день после рождения
|
|
1,17±0,028
|
1,14±0,062
|
1,25±0,037
|
1,29±0,076
|
|
При отъеме
|
|
1,15±0,039
|
1,15±0,050
|
1,25±0,039
|
1,25±0,037
|
|
Мочевина, ммоль/л
|
На 4-й день после рождения
|
|
0,57±0,045
|
0,61±0,000
|
0,58±0,034
|
0,54±0,045
|
|
На 21-й день после рождения
|
|
1,03±0,032
|
1,10±0,042
|
1,02±0,084
|
0,95±0,084
|
|
При отъеме
|
|
1,45±0,097
|
1,32±0,084
|
1,51±0,064
|
1,44±0,042
|
|
Фосфор неорганический,
ммоль/л
|
На 4-й день после рождения
|
|
2,80±0,166
|
2,84±0,048
|
2,92±0,099
|
2,93±0,070
|
|
На 21-й день после рождения
|
|
3,20±0,124
|
3,25±0,129
|
3,25±0,152
|
3,28±0,070
|
|
При отъеме
|
|
3,11±0,071
|
3,13±0,233
|
3,18±0,090
|
3,22±0,129
|
|
Кальций, ммоль/л
|
На 4-й день после рождения
|
1,60±0,081
|
1,68±0,041*
|
1,75±0,067*
|
|
На 21-й день после рождения
|
|
1,50±0,036
|
1,60±0,079
|
1,70±0,051*
|
1,69±0,081
|
|
При отъеме
|
|
1,4800,052
|
1,55±0,090
|
1,61±0,041
|
1,64±0,076
|
Достоверные различия между группами были выявлены по содержанию в крови
поросят кальция. Концентрация кальция при рождении поросят была выше во II группе на 6,0%, в III - на 11,3% (Р<0,05), в IV - на 15,9% (Р<0,05), чем в
контроле, а к 45-му дню жизни - соответственно на 4,7, 8,8, 10,8%.
Эти данные позволяют судить о положительном влиянии пикумина на показатели
минерального обмена полученного приплода.
Таким образом, использование минеральной добавки пикумин в кормлении
свиноматок в течение первого периода супоросности и подсоса позволяет получить
большее количество здоровых и жизнеспособных поросят с повышенной их
сохранностью и энергией роста, более развитыми клеточными и гуморальными
показателями естественной резистентности организма. При этом заметно возрастали
показатели белкового и минерального обмена в организме молодняка, улучшалась
картина морфологического состава крови.
Использование пикумина свиноматкам в течение второго периода супоросности
(с 84-го дня) и лактации
По данным литературных источников второй период супоросности является
наиболее ответственным, так как у животного повышаются обменные процессы, и
только правильное кормление и содержание позволят получить здоровый приплод.
Целью второго опыта явилось изыскание возможности получения
жизнеспособного молодняка при введении пикумина с последнего месяца
супоросности.
Животные контрольных групп добавку не получали. Свиноматкам опытных групп
вводили в рацион 0,8, 1,2 и 1,5% пикумина в расчете на 1 кг сухого вещества
корма. При этом добавка применялась в течение второго периода супоросности (с
84-го дня) и лактации. Из четырех групп животных I-ая была контрольной, II-ой вводили в рацион 0,8% пикумина в расчете на 1 кг сухого
вещества корма, III-ей - 1,2%, IV-ой - 1,5%.
Продолжительность использования добавки составила: второй период
супоросности (30 дней) + подсосный период (45 дней)-всего 75 дней.
Естественные защитные силы и показатели крови свиноматок
Введение в рацион свиноматок минеральной добавки пикумин с 84-го дня
супоросности до отъема поросят определенным образом сказалось на показателях
естественной резистентности их организма.
Изучение иммунорегуляторного действия минеральной добавки на 4-й день
после опороса показало (рис. 11), что бактерицидная активность сыворотки крови
у животных II группы была выше, чем в контрольной
на 2,8%, III группы - на 4,9%, IV - на 3,1%. В течение подсосного периода
наблюдалось снижение этого показателя у животных всех групп. Однако в III и IV группах бактерицидная активность была значительно выше
контроля, причем, в этих группах к 21-му дню лактации рост был достоверным.
Схожая картина наблюдалась и к отъему поросят. В это время животные II группы превосходили контрольных на
4,4%, III - на 9,4%, IV - на 8,8%.
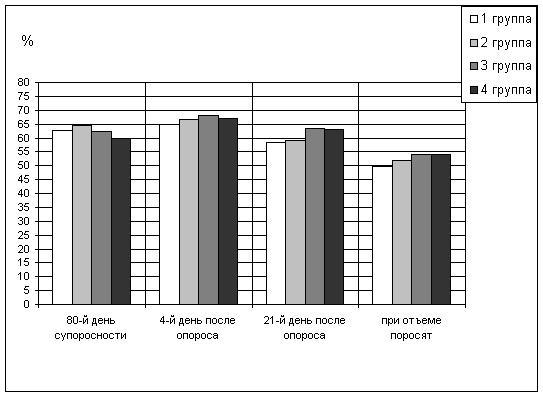
Рис.
11. Бактерицидная активность сыворотки крови, %
По активности лизоцима во все периоды исследований существенных различий
между группами установлено не было (рис. 12). При постановке на опыт лизоцимная
активность сыворотки крови находилась в пределах 4,10±0,193 - 4,50±0,253%. В ходе исследований
наблюдалась тенденция к снижению этого показателя до уровня 2,48±0,107 - 2,52±0,107%.
Было отмечено некоторое влияние сбалансированного кормления по макро- и
микроэлементам на фагоцитарную активность лейкоцитов в крови животных опытных
групп (рис. 13). На 4-й день после опороса опытные свиноматки отличались по
этому показателю от контрольных на 0,6-4,0%. Разница заметно увеличилась к
45-му дню подсосного периода, тогда превосходство составляло 1,5-11,9%.

Рис.
12. Лизоцимная активность сыворотки крови, %

Рис. 13. Фагоцитарная активность лейкоцитов, %
Применение минеральной добавки не вызвало аллергической реакции у
животных (рис. 14). Это подтверждают данные по концентрации сиаловых кислот в
крови. В начале опыта их содержание было в пределах 16,93-18,35 ед.опт.пл., к
4-му дню после опороса снижалось до 14,14-16,35 ед.опт.пл., а в конце
исследований достигало 12,71-13,99 ед.опт.пл. без достоверных различий между
группами.
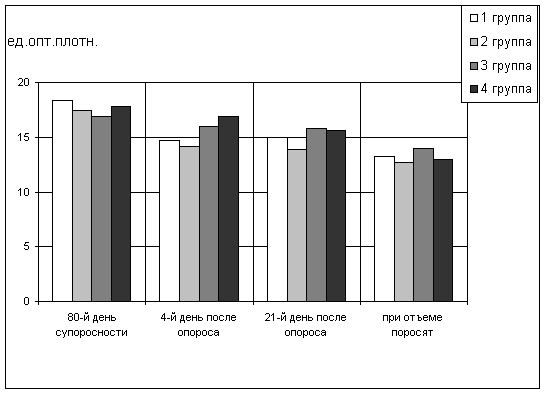
Рис.
14. Сиаловые кислоты, ед. опт. плотн.
Расчеты
комплексного иммунологического показателя позволили установить, что
дополнительное введение в рацион свиноматок минеральной добавки пикумин
положительно влияет на факторы защиты их организма (рис. 15). К 4-му дню
подсосного периода опытные животные отличались повышенной устойчивостью
организма к факторам внешней среды. В это время комплексный показатель
естественной резистентности составил у свиноматок I группы 0,35, II -
0,42, III - 0,61, IV группы - 0,52. К 45-му дню -
соответственно 0,36, 0,41, 0,62, 0,57. Таким образом, естественная
резистентность организма к концу опыта была выше у животных, получавших к
рациону 1,2 и 1,5% пикумина в расчете на 1 кг сухого вещества корма.
Морфологические
показатели крови свиноматок. Изучение содержания в крови свиноматок лейкоцитов
показало, что скармливание животным пикумина не повлияло на этот показатель
(табл. 11). В течение опыта количество лейкоцитов находилась примерно на одном
уровне у всех животных.

Рис.
15. Комплексный показатель естественной резистентности свиноматок
Таблица 11
Морфологические показатели крови свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Лейкоциты, 109/л
|
При постановке на опыт
|
|
12,20±0,449
|
11,94±0,622
|
13,05±0,618
|
11,49±0,343
|
|
На 4-й день после опороса
|
|
12,13±0,365
|
12,09±0,773
|
12,80±0,461
|
12,08±0,300
|
|
На 21-й день после опороса
|
|
12,88±0,322
|
12,76±0,536
|
13,47±0,365
|
12,91±0,365
|
|
При отъеме поросят
|
|
13,47±0,354
|
13,56±0,612
|
13,83±0,604
|
13,75±0,365
|
|
Эритроциты, 1012/л
|
При постановке на опыт
|
|
5,88±0,079
|
5,93±0,067
|
5,84±0,124
|
5,79±0,073
|
|
На 4-й день после опороса
|
|
5,69±0,062
|
5,89±0,062
|
5,53±0,049
|
5,70±0,054
|
|
На 21-й день после опороса
|
|
5,55±0,049
|
5,81±0,073*
|
5,48±0,109
|
5,70±0,098
|
|
При отъеме поросят
|
|
5,39±0,073
|
5,62±0,075
|
5,45±0,079
|
5,64±0,075*
|
Некоторое
увеличение числа эритроцитов к 21-му дню подсосного периода наблюдалось в крови
животных II и IV групп. Так, они превзошли по этому показателю контрольных во
II группе на 4,7% (Р<0,05), в IV - на 0,4%. К отъему рост количества
эритроцитов во II группе составил
4,3%, в IV - 4,6% (Р<0,05). Содержание
эритроцитов в крови животных контрольной группы и III опытной в течение опыта было примерно одинаковым.
Биохимические показатели крови свиноматок. При постановке на опыт
содержание общего белка в крови свиноматок находилось в пределах 67,77±2,056 - 69,99±1,704 г/л (табл. 12). До опороса
наблюдался рост этого показателя, причем, в III и IV
группах более заметный. На 4-й день подсосного периода концентрация общего
белка в этих группах была выше, чем в контроле соответственно на 2,8 и 1,9%.
Разница между животными опытных и контрольной группы возрастала к концу опыта.
Так, у свиноматок II группы
концентрация общего белка была на 3,8%, III - на 10,2%, IV - достоверно на 10,3% (Р<0,05) вьше, чем в контроле.
При изучении распределения белка по фракциям было установлено, что
процент α-глобулинов на протяжении опыта
оставался примерно на одном уровне у животных всех групп без достоверных
различий.
К концу опыта было отмечено незначительное увеличение содержания
альбуминов и β-глобулинов в крови животных опытных групп. В это время
увеличение концентрации альбуминов у свиноматок III группы составило 2,8%, IV группы - 7,7%, а увеличение количества β-глобулинов составило соответственно
10,2 и 14,3%.
К отъему поросят было отмечено снижение процента γ-глобулинов.
Свиноматки, получавшие с рационом минеральную добавку, имели в крови
больше гемоглобина (табл. 13). По этому показателю экспериментальные животные
превосходили контрольных к 4-му дню подсоса на 4,1-5,4%, к 21-му дню - на
3,9-10,7%. В конце опыта установлено достоверное увеличение концентрации
гемоглобина в крови свиноматок IV
группы.
Введение в рацион свиноматок пикумина не повлияло на содержание в крови
глюкозы. В течение опыта ее концентрация у животных всех групп находилась
примерно на одном уровне.
Более заметные изменения наблюдались по количеству в крови холестерина. К
21-му дню подсосного периода был отмечен достоверный рост этого показателя у
свиноматок IV группы.
Связи между введением добавки и изменением содержания у животных мочевины
не установлено.
Таблица 12
Протеинограмма свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
При постановке на опыт
|
|
Общий белок, г/л
|
68,48±3,416
|
70,02±1,648
|
67,76±2,067
|
68,42±1,648
|
|
Альбумины, %
|
35,32±2,593
|
36,71±3,466
|
36,46±3,157
|
34,25±2,921
|
|
α-глобулины,%
|
14,23±1,222
|
15,71±0,436
|
14,63±0,691
|
16,34±0,873
|
|
β-глобулины,%
|
19,74±1,346
|
20,73±0,830
|
18,22±0,708
|
21,78±1,094
|
|
γ-глобулины,%
|
30,71±1,919
|
26,85±2,202
|
30,69±2,107
|
27,63±0,593
|
|
На 4-й день после опороса
|
|
Общий белок, г/л
|
84,29±1,167
|
83,65±1,991
|
86,59±1,579
|
85,82±1,579
|
|
Альбумины, %
|
38,98±2,751
|
39,89±1,736
|
40,29±3,129
|
40,09±3,096
|
|
α-глобулины,%
|
15,84±1,242
|
16,64±0,938
|
14,57±0,723
|
15,97±0,865
|
|
β-глобулины,%
|
16,89±0,815
|
18,10±0,854
|
17,98±1,238
|
17,23±1,657
|
|
γ-глобулины,%
|
28,29±1,882
|
25,38±0,376
|
27,16±1,403
|
26,72±0,848
|
|
На 21-й день после опороса
|
|
Общий белок, г/л
|
73,28±1,991
|
73,02±1,236
|
75,33±1,236
|
76,08±1,618
|
|
Альбумины, %
|
40,86±1,794
|
42,04±2,148
|
42,87±2,350
|
42,96±2,843
|
|
α-глобулины,%
|
16,40±0,854
|
16,81±1,496
|
17,70±0,586
|
17,01±0,447
|
|
β-глобулины,%
|
17,75±0,513
|
17,49±1,135
|
18,81±1,273
|
17,60±1,177
|
|
γ-глобулины,%
|
24,99±0,858
|
23,66±0,541
|
20,63±0,852
|
22,43±1,427
|
|
При отъеме поросят
|
|
Общий белок, г/л
|
54,66±1,854
|
56,70±1,648
|
58,88±2,236
|
60,27±1,442*
|
|
Альбумины, %
|
38,22±3,058
|
37,32±2,942
|
39,37±2,584
|
41,22±2,170
|
|
α-глобулины,%
|
18,51±1,264
|
19,94±1,253
|
21,07±0,691
|
20,60±0,564
|
|
β-глобулины,%
|
15,96±0,401
|
17,58±1,440
|
17,47±0,480
|
18,18±1,350
|
|
γ-глобулины,%
|
27,30±1,738
|
25,16±1,667
|
22,09±1,632
|
20,00±0,440
|
Таблица 13
Биохимические показатели крови свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Гемоглобин, г/л
|
При постановке на опыт
|
|
113,0±5,62
|
120,84,72
|
115,5±6,18
|
119,2±5,58
|
|
На 4-й день после опороса
|
|
117,6±5,15
|
122,4±3,43
|
122,8±4,29
|
124,0±3,43
|
|
На 21-й день после опороса
|
|
112,0±3,86
|
116,4±3,43
|
119,6±3,00
|
124,0±4,49
|
|
При отъеме поросят
|
|
120,0±3,43
|
126,8±5,58
|
131,5±7,87
|
134,4±4,29*
|
|
Глюкоза, ммоль/л
|
При постановке на опыт
|
|
3,91±0,129
|
3,96±0,139
|
3,94±0,183
|
3,89±0,118
|
|
На 4-й день после опороса
|
|
3,57±0,139
|
3,61±0,120
|
3,61±0,139
|
3,65±0,120
|
|
На 21-й день после опороса
|
|
3,83±0,099
|
3,83±0,118
|
3,81±0,099
|
3,89±0,183
|
|
При отъеме поросят
|
|
4,09±0,099
|
4,13±0,099
|
4,14±0,154
|
4,04±0,079
|
|
Холестерин, г/л
|
При постановке на опыт
|
|
0,82±0,039
|
0,80±0,019
|
0,79±0,073
|
0,84±0,047
|
|
На 4-й день после опороса
|
|
0,86±0,019
|
0,80±0,019
|
0,89±0,019
|
0,86±0,028
|
|
На 21-й день после опороса
|
|
0,65±0,028
|
0,62±0,017
|
0,74±0,030
|
0,79±0,051*
|
|
При отъеме поросят
|
|
0,68±0,028
|
0,64±0,028
|
0,70±0,025
|
0,72±0,039
|
|
Мочевина, ммоль/л
|
На 60-й день супоросности
|
|
1,93±0,084
|
1,91±0,131
|
1,93±0,129
|
1,97±0,163
|
|
На 4-й день после опороса
|
|
2,18±0,097
|
2,12±0,129
|
2,12±0,129
|
2,21±0,161
|
|
На 21-й день после опороса
|
|
1,97±0,131
|
1,91±0,099
|
1,97±0,131
|
1,93±0,171
|
|
При отъеме поросят
|
|
1,75±0,099
|
1,79±0,131
|
1,82±0,087
|
1,78±0,099
|
Минеральный состав крови свиноматок. Изучение содержания в крови
подопытных животных макро- и микроэлементов показало, что введение в рацион
свиноматок минеральной добавки пикумин повлияло на концентрацию в крови
большинства изучаемых элементов (табл.14).
Несмотря на отсутствие достоверности, увеличение содержания фосфора в
крови опытных свиней к концу исследований составило 1,6-3,6% по сравнению с
контролем. Высокое содержание в минеральной добавке кальция сказалось и на
более заметном увеличении концентрации этого макроэлемента в крови свиноматок III и IV групп. Количество кальция у животных этих групп было больше
на 7,8-8,2%, чем у контрольных.
Таблица 14
Минеральный состав крови свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
1
|
2
|
3
|
4
|
5
|
|
Фосфор неорганический, ммоль/л
|
При постановке на опыт
|
|
2,56±0,045
|
2,52±0,124
|
2,44±0,059
|
2,50±0,107
|
|
На 4-й день после опороса
|
|
2,77±0,043
|
2,74±0,124
|
2,73±0,045
|
2,81±0,099
|
|
При отъеме поросят
|
|
1,93±0,054
|
1,97±0,116
|
2,01±0,056
|
2,00±0,107
|
|
Кальций, ммоль/л
|
При постановке на опыт
|
|
2,31±0,104
|
2,35±0,060
|
2,27±0,065
|
2,30±0,058
|
|
На 4-й день после опороса
|
|
2,40±0,092
|
2,44±0,071
|
2,49±0,056
|
2,48±0,073
|
|
При отъеме поросят
|
|
2,31±0,069
|
2,34±0,062
|
2,49±0,079
|
2,50±0,060
|
|
Цинк, мкмоль/л
|
При постановке на опыт
|
|
34,88±2,051
|
36,00±1,212
|
34,54±1,326
|
33,02±1,897
|
|
На 4-й день после опороса
|
38,64±1,457
|
38,71±1,854
|
38,03±1,721
|
|
При отъеме поросят
|
|
33,76±1,348
|
33,90±1,637
|
34,53±1,112
|
34,98±1,448
|
|
Магний, ммоль/л
|
При постановке на опыт
|
|
1,16±0,101
|
1,04±0,058
|
1,21±0,098
|
1,18±0,062
|
|
На 4-й день после опороса
|
|
1,16±0,101
|
1,22±0,054
|
1,34±0,077
|
1,37±0,069
|
|
При отъеме поросят
|
|
1,04±0,090
|
1,17±0,052
|
1,29±0,115
|
1,31±0,103
|
|
Железо, мкмоль/л
|
При постановке на опыт
|
|
30,41±1,907
|
32,63±1,946
|
29,51±2,581
|
31,00±1,412
|
|
На 4-й день после опороса
|
|
36,22±0,996
|
37,09±1,242
|
39,23±1,865
|
39,90±1,489
|
|
При отъеме поросят
|
|
26,74±1,418
|
30,70±2,118
|
35,46±2,472*
|
35,96±1,408**
|
|
Натрий, ммоль/л
|
При постановке на опыт
|
|
146,4±4,58
|
148,6±4,12
|
146,2±4,44
|
140,8±2,51
|
|
На 4-й день после опороса
|
|
150,4±3,37
|
152,6±4,25
|
158,0±3,45
|
160,0±3,84
|
|
При отъеме поросят
|
|
145,6±3,61
|
148,2±3,20
|
153,8±4,44
|
152,4±2,60
|
|
|
|
|
|
|
Исследования показали, что концентрация цинка в крови животных при
постановке на опыт находилась на уровне 33,02±1,897 - 36,00±1,212 ммоль/л. К концу подсосного периода матки II группы превосходили по этому
показателю контрольных на 0,4%, III -
на 2,3%, IV группы - на 3,6%.
При изучении содержания магния в крови свиноматок было выявлено, что на
4-й день подсосного периода у свиней опытных групп его концентрация была выше
на 5,2-18,1%, на 45-й день - на 12,5-26,0%.
Существенные изменения наблюдались по содержанию в крови железа. Так, к
4-му дню после опороса у свиноматок II группы содержание железа было выше на 2,4%, III - на 8,3%, IV - на 10,1%, чем в контроле, а к отъему - соответственно на 14,8, 32,6
(Р<0,05) и 34,5% (Р<0,01).
По концентрации натрия у животных достоверных изменений не установлено. К
45-му дню подсосного периода превосходство опытных групп над контрольными по
этому показателю составляло 1,7-4,6%.
Продуктивность и воспроизводительные способности свиноматок
Применение минеральной добавки со второго периода супоросности и в
течение всего подсосного периода оказало положительное влияние на
продуктивность свиноматок (табл. 15).
По многоплодию не отмечены достоверные различия между контрольной и
опытными группами. Однако слабых поросят во II и IV
группах было получено на 8,3%, в III -
на 25,5% меньше, чем от контрольных свиноматок, а мертвых во II и III меньше на 33,3%, в IV - на 44,4%. При этом масса гнезда при рождении была выше на
2,3-3,6% у животных, получавших к рациону комплекс макро- и микроэлементов в
составе пикумина.
В дальнейшем поросята от маток опытных групп лучше развивались и росли,
меньше подвергались заболеваниям. Этому способствовала и более высокая
молочность свиноматок.
Определение молочности по контрольному взвешиванию гнезда в 21-дневном
возрасте показало, что свиноматки, получавшие минеральную добавку пикумин,
имели более высокие показатели. По молочности животные II группы превосходили контрольных на 8,0%, III - на 9,1%, IV - на 9,7%.
Масса гнезда при отъеме также была выше в опытных группах. Сохранность
приплода в опытных группах превышала контроль на 2,1-3,0%.
Таблица 15
Показатели воспроизводительных функций свиноматок
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Родилось живых поросят в
среднем на 1 матку, голов
|
9,36±0,699
|
9,27±0,599
|
9,55±0,599
|
9,31±0,606
|
|
В том числе слабых, голов
|
1,09±0,299
|
1,00±0,299
|
0,91±0,200
|
1,00±0,260
|
|
Родилось мертвых поросят в
среднем на 1 матку, голов
|
0,27±0,100
|
0,18±0,100
|
0,18±0,100
|
0,15±0,866
|
|
Масса гнезда при рождении,
кг
|
11,69±0,778
|
11,96±0,808
|
11,88±0,858
|
12,11±0,762
|
|
Молочность свиноматок, кг
|
51,5±4,57
|
55,6±3,97
|
56,2±2,60
|
56,5±4,38
|
|
Масса гнезда при отъеме, кг
|
82,4±6,51
|
85,2±8,18
|
88,4±5,03
|
89,2±6,00
|
|
Сохранность поросят, %
|
92,2±1,43
|
94,1±2,50
|
94,3±1,25
|
95,0±1,24
|
Таким образом, полученные нами данные свидетельствуют, что улучшение
минерального питания глубокосупоросных и подсосных свиноматок способствовало
нормализации минерального обмена в их организме. Это в свою очередь
положительно сказалось на показателях белкового обмена животных и естественной
резистентности организма. При этом улучшались кроветворные и
воспроизводительные функции свиноматок, повышалась их продуктивность.
Полученный приплод обладал большей жизнеспособностью и более высокой
сохранностью к отъему.
Энергия роста
и гематологические показатели поросят-сосунов
Энергия роста
поросят-сосунов. Введение минеральной добавки пикумин в рацион
глубокосупоросных свиноматок позволило несколько повысить живую массу поросят II и IV групп при рождении (табл. 16). Так, живая масса родившегося
молодняка II группы была на 3,2%, IV группы - на 4,0% больше, чем в
контрольной группе.
Полученные от
опытных маток поросята в течение подсосного периода лучше развивались и росли.
К отъему живая масса поросят II
группы была на 2,4%, III -
на 3,0%, IV - на 5,7% выше, чем в контроле. При
этом в опытных группах был выше абсолютный прирост. По среднесуточному приросту
молодняк от свиноматок, получавших с рационом минеральную добавку, превосходил
контроль на 2,2-6,0%.
Однако
достоверных различий по живой массе между животными в течение всего периода
исследований установлено не было.
Таблица 16
Динамика приростов живой массы полученных поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Живая масса, кг: при
рождении
|
1,25±0,054
|
1,29±0,045
|
1,24±0,056
|
1,30±0,043
|
|
- в 21 день
|
5,95±0,218
|
6,37±0,228
|
6,25±0,214
|
6,38±0,210
|
|
-при отъеме (в 45 дней)
|
9,53±0,474
|
9,76±0,433
|
9,82±0,504
|
10,07±0,484
|
|
Абсолютный прирост, кг
|
8,28±0,333
|
8,47±0,396
|
8,58±0,394
|
8,77±0,294
|
|
Среднесуточный прирост, г
|
184±7,5
|
188±8,9
|
191±8,7
|
195±6,6
|
|
% к контролю
|
100,0
|
102,2
|
103,8
|
106,0
|
Показатели естественной защиты поросят-сосунов. Изучение гуморальных
факторов защиты полученного приплода показало, что на 4-й день после рождения
поросята опытных групп по уровню бактерицидной активности сыворотки крови на
1,3-3,4% превосходили контроль (рис. 16). В течение подсосного периода этот
показатель возрастал, причем, его рост был интенсивнее у животных опытных групп.
К 21-му дню подсосного периода поросята II группы отличались по этому показателю от контрольных на
6,0%, III - на 15,5%, IV - на 14,3%. К отъему молодняка разница составляла
соответственно 5,3, 10,1, 10,6%.

Рис.
16. Бактерицидная активность сыворотки крови, %
По
лизоцимной активности сыворотки крови при рождении поросят существенной разницы
не установлено (рис. 17). На 4-й день подсосного периода она находилась в
пределах от 5,22 до 5,45%. Несколько ниже у поросят всех групп активность
лизоцима стала к 21-му дню после рождения. Однако у молодняка от маток,
получавших с рационом минеральную добавку пикумин, она была значительно выше, а
у животных III группы установлено достоверное (Р<0,01) увеличение
активности лизоцима по сравнению с контрольными. К концу опыта у молодняка II
группы активность лизоцима была выше на 2,2%, III - на 18,3%
(Р<0,05), IV -на 17,2% (Р<0,05).
Исследование
клеточных факторов защиты организма поросят показало, что пикумин повлиял на
фагоцитарную активность лейкоцитов крови родившегося молодняка (рис. 18). Так,
поросята от опытных свиноматок при рождении на 2,3-6,0% превосходили по этому
показателю контрольных. В течение подсосного периода у животных опытных групп
наблядался рост фагоцитарной активности лейкоцитов. В конце опыта интенсивность
фагоцитоза в этих группах была на 3,8-9,5% выше, чем в контроле.

Рис.
17. Лизоцимная активность сыворотки крови, %

Рис. 18. Фагоцитарная активность лейкоцитов, %
Концентрация сиаловых кислот в крови подопытных поросят при рождении
колебалась от 11,20±1,180
до 12,45±0,951 ед.опт.пл. (рис. 19).
Практически на таком уровне она оставалась и в трехнедельном возрасте. В конце
опыта количество сиаловых кислот в крови подопытных поросят несколько
повысилось во всех группах без достоверных различий.

Рис. 19. Сиаловые кислоты, ед. опт. плотн.
Расчет комплексного показателя естественной резистентности показал, что
скармливание глубокосупоросным и подсосным свиноматкам пикумина значительно
повлияло на становление факторов естественной защиты родившегося молодняка
(рис. 20). Улучшение минерального питания свиноматок, более высокая их
молочность, позволили стабилизировать физиологические параметры резистентности
полученного приплода.

Рис.
20. Комплексный показатель естественной резистентности поросят
Морфологические и бихимические показатели крови поросят. Изучение
морфологических показателей крови полученного приплода свидетельствует о том,
что введение в рацион свиноматкам пикумина не повлияло на содержание лейкоцитов
в крови родившихся поросят (табл. 17). Концентрация лейкоцитов при рождении
молодняка находилась на уровне 8,98±0,236 - 9,10±0,408 Х109/л без видимых различий между группами. В течение подсосного
периода достоверных различий между группами по этому показателю не установлено.
Значительное увеличение числа эритроцитов в крови поросят III и IV групп также наблюдалось в конце опыта. Так, по этому
показателю они превзошли контрольных в III группе - на 2,8%, в IV - на 3,2%.
Введение пикумина свиноматкам вызвало некоторое увеличение количества
общего белка в сыворотке крови полученного молодняка к 21-му дню подсосного
периода (табл. 18). Так, в этот период исследований у поросят II группы этот показатель был выше, чем
в I на 6,2%, III - на 4,7%, IV - на 5,5%. К концу опыта наблюдался спад концентрации общего белка в крови
животных всех групп. Однако этот показатель оставался на более высоком уровне у
подопытных поросят.
Таблица 17
Морфологические показатели крови поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
Лейкоциты, 109/л
|
На 4-й день после рождения
|
|
9,07±0,418
|
8,98±0,236
|
8,90±0,337
|
9,10±0,408
|
|
На 21-й день после рождения
|
|
8,86±0,451
|
9,11±0,247
|
9,25±0,225
|
9,15±0,590
|
|
При отъеме
|
|
9,43±0,618
|
9,55±0,300
|
9,54±0,253
|
9,42±0,472
|
|
Эритроциты, 1012/л
|
На 4-й день после рождения
|
|
5,77±0,084
|
5,71±0,073
|
5,89±0,143
|
5,82±0,099
|
|
На 21-й день после рождения
|
|
6,10±0,060
|
6,10±0,073
|
6,17±0,097
|
6,22±0,112
|
|
При отъеме
|
|
5,64±0,160
|
5,59±0,086
|
5,81±0,096
|
5,62±0,098
|
Таблица 18
Протеинограмма поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
На 4-й день после рождения
|
|
Общий белок, г/л
|
58,37±1,442
|
57,47±1,236
|
57,92±1,798
|
58,82±1,854
|
|
Альбумины, %
|
30,19±2,929
|
31,35±1,637
|
31,48±3,826
|
31,11±2,391
|
|
α-глобулины,%
|
20,76±1,185
|
19,79±0,805
|
20,42±1,180
|
17,62±1,114
|
|
β-глобулины,%
|
29,97±2,296
|
28,43±1,798
|
27,33±2,702
|
28,47±2,200
|
|
γ-глобулины,%
|
19,08±1,215
|
20,44±1,509
|
20,77±0,789
|
22,79±1,627
|
|
На 21-й день после рождения
|
|
Общий белок, г/л
|
61,57±1,236
|
65,28±1,923
|
64,38±1,511
|
65,04±1,030
|
|
Альбумины, %
|
34,22±2,474
|
33,11±2,223
|
35,13±2,873
|
35,33±2,767
|
|
α-глобулины,%
|
21,44±0,697
|
23,62±1,644
|
19,85±0,886
|
18,08±1,183
|
|
β-глобулины,%
|
26,16±1,519
|
23,27±1,356
|
25,28±1,747
|
26,24±1,261
|
|
γ-глобулины,%
|
18,17±0,865
|
20,01±1,691
|
19,73±2,122
|
20,36±0,935
|
|
При отъеме
|
|
Общий белок, г/л
|
52,00±2,157
|
54,21±1,236
|
54,24±2,157
|
54,91±1,373
|
|
Альбумины, %
|
41,51±3,270
|
41,13±2,558
|
42,64±3,823
|
43,62±2,453
|
|
α-глобулины,%
|
19,83±0,806
|
20,16±0,936
|
13,25±0,621
|
12,14±0,893
|
|
β-глобулины,%
|
22,52±1,157
|
18,98±1,232
|
20,43±1,688
|
20,15±0,878
|
|
γ-глобулины,%
|
16,14±1,351
|
19,72±1,024
|
23,68±4,567
|
24,08±1,330**
|
|
|
|
|
|
|
Анализ
распределения белка по фракциям показал, что в ходе опыта в крови
экспериментальных животных повышался процент альбуминов и γ-глобулинов. Содержание β-глобулинов было примерно на одном
уровне, α-глобулинов - снижалось. При этом
следует отметить, что увеличение к концу опыта процента γ-глобулинов было достоверным.
При
определении содержания в крови гемоглобина было выявлено, что к окончанию опыта
у опытного молодняка наблюдалось значительное превосходство по этому показателю
над контрольными животными (во II
группе - на 8,6%, в III - на 12,6%, в IV - на 12,1%) (табл. 19).
При изучении
содержания в крови подопытных животных количества глюкозы и холестерина
выявлено, что введение в рацион свиноматок минеральной добавки пикумин не
вызывает у поросят их достоверных изменений. Не выявлено различий между группами
и по концентрации в крови мочевины.
Заметные изменения наблюдались по содержанию в крови неорганического
фосфора и кальция. При рождении концентрация фосфора в опытных группах была на
2,6-3,7% выше, чем в контрольной, а к отъему - на 2,3-4,8%. К 21-му дню жизни
установлено достоверное (Р<0,05) увеличение количества кальция в крови
поросят III группы. К 45-му дню подсосного
периода молодняк II группы
превосходил по этому показателю контроль на 1,3%, III - на 5,9%, IV группы - на 5,3%.
Таблица 19
Биохимический и минеральный состав крови поросят
|
Показатели
|
Группы
|
|
I
|
II
|
III
|
IV
|
|
1
|
2
|
3
|
4
|
5
|
|
Гемоглобин, г/л
|
На 4-й день после рождения
|
|
103,6±2,58
|
100,4±3,43
|
105,5±6,74
|
98,4±4,29
|
|
На 21-й день после рождения
|
|
131,6±3,86
|
130,0±3,00
|
134,4±6,01
|
132,0±5,06
|
|
При отъеме
|
|
103,5±6,18
|
112,4±5,58
|
116,5±7,87
|
116,0±5,15
|
|
Глюкоза, ммоль/л
|
На 4-й день после рождения
|
|
4,79±0,238
|
4,85±0,118
|
4,83±0,365
|
5,01±0,079
|
|
На 21-й день после рождения
|
|
5,18±0,200
|
5,18±0,139
|
5,24±0,200
|
5,28±0,154
|
|
При отъеме
|
|
4,68±0,258
|
4,98±0,137
|
4,97±0,208
|
5,05±0,178
|
|
Холестерин, г/л
|
На 4-й день после рождения
|
|
1,21±0,028
|
1,23±0,028
|
1,25±0,051
|
1,27±0,028
|
|
На 21-й день после рождения
|
|
1,21±0,019
|
1,26±0,047
|
1,27±0,019
|
1,26±0,076
|
|
При отъеме
|
|
1,19±0,037
|
1,20±0,036
|
1,24±0,022
|
1,23±0,056
|
|
Мочевина, ммоль/л
|
|
0,51±0,034
|
0,58±0,066
|
0,49±0,045
|
0,48±0,034
|
|
На 21-й день после рождения
|
|
0,97±0,032
|
0,97±0,032
|
0,99±0,032
|
0,95±0,042
|
|
При отъеме
|
|
1,36±0,084
|
1,42±0,097
|
1,47±0,126
|
1,36±0,064
|
|
Фосфор неорганический,
ммоль/л
|
На 4-й день после рождения
|
|
2,71±0,116
|
2,77±0,073
|
2,79±0,152
|
2,83±0,161
|
|
На 21-й день после рождения
|
|
3,18±0,107
|
3,23±0,082
|
3,26±0,116
|
3,23±0,199
|
|
При отъеме
|
|
3,12±0,093
|
3,17±0,071
|
3,20±0,140
|
3,26±0,133
|
|
Кальций, ммоль/л
|
На 4-й день после рождения
|
|
1,55±0,043
|
1,62±0,060
|
1,65±0,067
|
1,68±0,049
|
|
На 21-й день после рождения
|
|
1,49±0,043
|
1,58±0,060
|
1,67±0,052*
|
1,67±0,079
|
|
При отъеме
|
|
1,52±0,059
|
1,54±0,060
|
1,61±0,067
|
1,60±0,060
|
Следовательно, полученные данные свидетельствуют о том, что введение в
рацион глубокосупоросных и подсосных свиноматок минеральной добавки пикумин в
дозах 1,2-1,5% в расчете на 1 кг сухого вещества корма способствует получению
от них более крепкого и жизнеспособного приплода с повышенной энергией роста;
позволяет повысить уровень естественных защитных сил поросят-сосунов;
благотворно влияет на морфологические и биохимические показатели их крови,
показатели минерального обмена.
Заключение
Реализация генетического потенциала продуктивности животных возможна при
оптимизации факторов внешней среды: условий кормления и содержания животных,
микроклимата помещений, технологии производства, ухода при эксплуатации и т.д.
Благоприятное сочетание условий внешней среды позволяет повысить здоровье
животных, получить от них возможно большее количество здоровой и качественной
продукции, способствует получению и сохранению крепкого здорового молодняка с
развитой естественной устойчивостью организма к болезням.
Одним из важнейших факторов внешней среды, влияющих на организм животных,
в том числе и на его защитные механизмы, является кормление. При этом особое
значение приобретает тип и уровень кормления, соотношение отдельных кормов в
рационе, сбалансированность рационов по питательным и биологически активным
веществам.
Макро- и микроэлементы, не представляя энергетической ценности, играют в
организме большую роль, которая заключается в их участии во всех процессах
обмена. Наряду со специфическими функциями большое значение минеральные
вещества имеют в поддержании осмотического давления, буферной емкости жидкостей
и тканей организма, нервного и мышечного возбуждения, регуляции каталитических
процессов, проявлении иммунобиологической реактивности организма. Недостаток
минеральных веществ в рационе отрицательно сказывается на степени минерализации
скелета у свиней, их здоровье, продуктивности, продолжительности жизни, функции
воспроизводства.
Исследование кормов, применяемых в кормлении супоросных и подсосных
свиноматок, показало, что многих минеральных веществ, незаменимых в организме,
в них содержится в недостаточном количестве. Это сказывается на рождении
молодняка с низкой жизнеспособностью, несовершенной системой естественной
защиты организма, низкими продуктивными качествами. В связи с этим, была
предпринята попытка изучить возможность применения в качестве минеральной
добавки к рациону свиноматок отходов местного производства керамзита -
пикумина.
Установлено, что ежедневное применение свиноматкам пикумина в течение
первого периода супоросности (до 84-го дня) и лактации оказало положительное
влияние на гуморальные и клеточные факторы защиты их организма. Бактерицидная
активность сыворотки крови во все периоды исследований была выше у свиноматок,
получавших добавку пикумина. Особенно это повышение отмечалось в крови животных
третьей и четвертой группы. Изучение лизоцимной активности сыворотки крови не
выявило достоверных различий между группами. Тем не менее, во все периоды
исследований у свиноматок третьей и четвертой групп она была выше на 3,2-8,3%
по сравнению с контролем. Фагоцитоз протекал активнее в крови опытных животных.
С целью более объективной оценки влияния минеральной добавки пикумин на
показатели естественной защиты организма животных был определен комплексный
показатель неспецифической резистентности. Расчеты показали, что уже к 60-му дню
супоросности животные третьей и четвертой групп отличались повышенной
естественной резистентностью организма. В течение опыта комплексный
иммунобиологический показатель заметно возрастал у животных, получавших
минеральную добавку.
Результаты исследования сиаловых кислот показали, что введение в рацион
минеральной добавки пикумин не вызвало аллергической реакции в организме
свиноматок.
Высокое содержание железа в изучаемой добавке способствовало росту
концентрации гемоглобина в крови у свиноматок, получавших пикумин.
Известно, что минеральные вещества являются катализаторами обменных
процессов, происходящих в организме. Положительное влияние пикумина на белковый
обмен нашло подтверждение при изучении концентрации в крови животных общего
белка и анализе распределения его по фракциям. У свиноматок, получавших с
рационом 1,2 и 1,5% пикумина, на 8,0 и 14,6% соответственно возростало
содержание общего белка в крови по сравнению с контролем. При этом наблюдался
рост в крови процента альбуминовой и γ-глобулиновой фракций белка. В конце
опыта количество γ-глобулинов у свиноматок третьей и четвертой групп было
на 18,0 и 23,9% соответственно выше, чем у контрольных животных.
Полученные данные по содержанию в крови холестерина показывают, что
введение в рацион пикумина способствовало достоверному повышению этого
показателя у свиноматок третьей группы к 21-му дню подсосного периода.
Дополнительное введение в рацион свиноматок макро- и микроэлементов в
составе пикумина положительно сказалось на показателях минерального состава
крови свиноматок. Так, к концу подсосного периода установлено увеличение
содержания фосфора в крови опытных животных на 5,1- 11,6% по сравнению с
контролем, причем в третьей группе оно было достоверным. В это же время на 9,6%
у свиноматок третьей и четвертой групп возросла концентрация в крови кальция.
Существенные изменения наблюдались по содержанию в крови цинка, магния и
железа.
Введение в рацион свиноматок кормовой минеральной добавки пикумин оказало
положительное действие на гуморальные и клеточные факторы защиты организма
полученного приплода. Бактерицидная активность сыворотки крови новорожденных
поросят была выше у животных третьей и четвертой групп, а к отъему - достоверно
выше. В течение подсосного периода у поросят этих групп также заметно возрастала
и активность лизоцима. Исследование фагоцитарной активности лейкоцитов
показало, что фагоцитоз проходил на 1,9% интенсивнее в опытных группах, чем в
контрольной, а при отъеме - на 1,2-9,6%.
При определении комплексного показателя естественной резистентности
организма поросят стало очевидным, что наиболее развитыми естественные защитные
силы были у молодняка, чьи матки получали 1,2 и 1,5% пикумина в расчете на 1 кг
сухого вещества корма.
Поросята от свиноматок, получавших минеральную добавку пикумин,
отличались более высоким содержанием в крови гемоглобина.
Сбалансированное питание свиноматок по минеральным веществам сказалось на
белковом обмене у полученного приплода. Было выявлено недостоверное увеличение
количества общего белка сыворотки крови у молодняка опытных групп. К концу
исследований опытные животные отличались от контрольных по этому показателю на
5,8-7,9%. Увеличение концентрации общего белка происходило главным образом за
счет роста альбуминовой и γ-глобулиновой его фракций. К отъему
молодняка установлено достоверное увеличение содержания γ-глобулинов в крови поросят третьей и
четвертой группы. Это говорит об укреплении иммунного статуса организма
молодняка.
Изучение углеводного обмена выявило некоторое увеличение содержания
глюкозы у поросят опытных групп. По этому показателю животные опытных групп при
рождении отличались от контрольных на 3,6-5,9%, а к отъему - на 5,1-8,6%.
Минеральный состав крови поросят-сосунов несколько отличался в опытных
группах. Уже при рождении количество фосфора у поросят этих групп было выше,
чем в контроле на 1,1-5,0%. В течение подсосного периода концентрация фосфора в
крови животных возрастала, причем, более заметно у поросят третьей и четвертой
групп.
Достоверные различия между группами поросят были выявлены по содержанию в
крови кальция. Концентрация кальция при рождении молодняка была выше в опытных
группах на 6,0-15,9%, чем в контроле, а к 45-му дню жизни - на 4,7-10,8%.
Повышение иммунного статуса подопытных свиноматок и полученного от них
молодняка, активизация кроветворной функции их организма, усиление некоторых
обменных процессов способствовало росту продуктивных качеств животных.
Установлено, что ежедневное применение свиноматкам пикумина в течение первого
периода супоросности (до 84-го дня) и лактации в дозах 1,2 и 1,5% к массе
сухого вещества корма позволяло повысить многоплодие на 8,3-9,3%. При этом
количество слабых и мертвых поросят значительно снижалось. Живая масса опытных
поросят при рождении была на 4,2-9,2% больше, чем в контроле. При изучении
молочности маток были выявлены заметные отличия по этому показателю у животных,
получавших к рациону минеральную добавку пикумин. Свиноматки опытных групп по
молочности превосходили контрольных на 15,9-24,8%, причем это превосходство
было более заметным у животных третьей и четвертой групп, получавших 1,2 и 1,5%
пикумина. Полученный от опытных свиноматок молодняк отличался повышенной
энергией роста в течение всего подсосного периода. У этих животных абсолютный
прирост за 45 дней превзошел контроль на 4,6-10,0%. Соответственно у этих
поросят был выше и среднесуточный прирост живой массы. К отъему в гнездах
опытных маток сохранилось на 0,5-4,2% больше поросят.
По данным литературных источников, второй период супоросности является
наиболее ответственным, так как у животного повышаются обменные процессы, и
только правильное кормление и содержание позволят получить здоровый приплод.
При введении пикумина в рацион свиноматок в течение второго периода
супоросности (с 84-го дня) и в период лактации установлено благоприятное
воздействие добавки на клеточные и гуморальные факторы неспецифической защиты
организма свиноматок. В опытных группах было отмечено достоверное увеличение в
крови бактерицидной активности сыворотки крови. Значительное, хотя и
недостоверное, наблюдалось повышение и фагоцитарной активности лейкоцитов.
Расчеты комплексного иммунологического показателя позволили установить,
что дополнительное введение в рацион свиноматок минеральной добавки пикумин в
изучаемые в этом опыте физиологические периоды положительно влияет на факторы
защиты их организма. Уже к опоросу опытные животные отличались повышенной
устойчивостью организма к факторам внешней среды. К концу опыта комплексный
показатель естественной резистентности у свиноматок контрольной группы составил
0,36, в то время как у опытных - 0,41-0,62.
Определение содержания сиаловых кислот позволяет утверждать, что их
количество в течение опыта находилось в пределах 12,71-18,35 ед.опт.пл., без
достоверных различий между группами.
Изучение морфологических показателей крови свиноматок показало, что у
опытных животных в крови была выше концентрация эритроцитов, возрастала
насыщенность эритроцитов гемоглобином.
Существенные изменения выявлены в белковом составе сыворотки крови.
Введение в рацион супоросных и подсосных свиноматок пикумина вызвало
достоверное увеличение количества общего белка у животных, получавших 1,5% этой
добавки. Этот показатель был на 5,63 г/л выше, чем в контроле. К концу опыта
было отмечено незначительное увеличение у животных опытных групп альбуминовой и
β-глобулиновой фракций. В это время
увеличение процента альбуминов у свиноматок третьей группы составило 2,8%,
четвертой группы - 7,7%; увеличение количества β-глобулинов в экспериментальных
группах составило 10,2-14,3%.
Заметные изменения наблюдались по количеству в сыворотке крови
холестерина. К 21-му дню подсосного периода был отмечен достоверный рост этого
показателя у свиноматок четвертой группы.
Изучение содержания в крови подопытных животных макро- и микроэлементов
показало, что введение в рацион свиноматок минеральной добавки пикумин повлияло
на концентрацию в крови большинства изучаемых элементов. Несмотря на отсутствие
достоверности, увеличение содержания фосфора у опытных свиней, по сравнению с
контролем к концу исследований составило 1,6-3,6%. Высокое содержание в
минеральной добавке кальция сказалось и на более заметном увеличении
концентрации этого макроэлемента в крови свиноматок третьей и четвертой групп.
Количество кальция у животных этих групп было больше на 7,8-8,2%, чем у контрольных.
Исследования выявили увеличение содержания цинка у свиноматок опытных групп.
При изучении содержания магния в крови свиноматок было установлено, что на 4-й
день подсосного периода у свиней опытных групп его концентрация была выше на
5,2-18,1%, на 45-й день - на 12,5-26,0%. Существенные изменения наблюдались по
содержанию в крови железа. Так, к 4-му дню после опороса у свиноматок второй
группы содержание железа было выше на 2,4%, третьей - на 8,3%, четвертой - на
10,1%, чем в контроле, а к отъему - соответственно на 14,8, 32,6 (Р<0,05) и
34,5% (Р<0,01).
Использование минеральной добавки в рационе подсосных свиноматок оказало
положительное влияние на показатели естественной защиты полученных от них
поросят. К 21-му дню подсосного периода поросята второй группы отличались по
бактерицидной активности сыворотки крови от контрольных на 6,0%, третьей - на
15,5%, четвертой - на 14,3%. К отъему молодняка разница составила
соответственно 5,3, 10,1, 10,6%. Аналогичное увеличение отмечено и по
активности лизоцима. В течение подсосного периода у животных опытных групп
наблюдался рост фагоцитарной активности лейкоцитов. В конце опыта интенсивность
фагоцитоза в этих группах была на 3,8-9,5% выше, чем в контроле.
Расчет комплексного показателя естественной резистентности показал, что
скармливание глубокосупоросным свиноматкам пикумина существенно не повлияло на
становление факторов естественной защиты полученного приплода. Тем не менее,
улучшение минерального питания свиноматок в указаный период и в период лактации,
более высокая их молочность, позволили стабилизировать физиологические
параметры резистентности поросят к концу подсосного периода.
Изучение морфологических показателей крови полученного приплода
свидетельствует о том, что скармливание свиноматкам пикумина в течение
подсосного периода способствовало увеличению у поросят содержания эритроцитов в
крови - на 2,8-3,2%, росту насыщенности эритроцитов гемоглобином на 8,6-12,6%.
У подопытных поросят наблюдалось недостоверное увеличение количества
общего белка в сыворотке крови. Анализ распределения белка по фракциям показал,
что в ходе опыта в крови экспериментальных животных повышался процент
альбуминов и γ-глобулинов. При этом следует отметить, что увеличение к концу
опыта содержания γ-глобулинов было достоверным.
Заметные изменения наблюдались по содержанию в крови неорганического
фосфора и кальция. При рождении концентрация фосфора в опытных группах была на
2,6-3,7% выше, чем в контрольной, а к отъему - на 2,3-4,8%. В трехнедельном
возрасте установлено достоверное увеличение количества кальция в крови поросят
третьей группы. К отъему молодняк второй группы превосходил по этому показателю
контроль на 1,3%, третьей группы - на 5,9%, четвертой- на 5,3%. Нарушений в
фосфорно-кальциевом соотношении не установлено.
Применение глубокосупоросным и подсосным свиноматкам пикумина сказалось
на продуктивности животных. Несмотря на то, что многоплодие маток находилось
без видимых различий между группами, количество слабых поросят было на
8,3-25,5%, а мертвых на 33,3-44,4% меньше у свиноматок, получавших минеральную
добавку пикумин. Живая масса поросенка при рождении в контрольной группе
составляла 1,25 кг, а в опытных доходила до 1,30 кг. Приплод от свиноматок
опытных групп лучше рос. Среднесуточный прирост живой массы у них был на
2,2-6,0% выше, чем у контрольных животных. Молочность свиноматок, получавших
добавку, была на 8,0-9,7% выше, а сохранность поросят за время подсосного
периода составляла 94,1-95,0% против 92,2% в контроле.
Таким образом, добавка к рациону супоросным свиноматкам пикумина
оказывает на организм самих животных и полученного от них приплода
разнообразное воздействие. Дополнительное введение в рацион комплекса макро- и
микроэлементов в составе изучаемой добавки оказало положительное действие на
морфологические, биохимические и иммунобиологические показатели крови.
Повышение естественной резистентности организма животных, улучшение обменных
процессов в их организме способствовало росту продуктивных и
воспроизводительных качеств свиноматок, повышению сохранности полученного
молодняка, увеличению энергии его роста.
Выводы
. В условиях промышленной технологии в организме свиноматок наблюдается
нарушение минерального обмена, что выражается пониженным содержанием в крови
жизненно необходимых элементов (кальция, магния, железа, цинка и др.),
снижением уровня естественной резистентности организма свиноматок и получаемого
от них приплода, их продуктивности. Для оптимизяции минерального обмена
свиноматок можно использовать минеральную добавку пикумин, получаемую из местного
дешевого сырья. Наиболее целесообразным является введение в рацион свиноматок
1,2% пикумина в расчете на 1 кг сухого вещества корма.
. Использование пикумина свиноматкам в течение первого периода
супоросности (до 84-го дня) и лактации повышает бактерицидную активность
сыворотки крови свиноматок - на 8,9%, лизоцимную активность сыворотки крови -
на 8,3%, фагоцитарную активность лейкоцитов - на 12,8%; в крови возрастает
содержание гемоглобина на 4,9%, общего белка - на 7,9% (главным образом за счет
альбуминовой и γ-глобулиновой его фракций), кальция - на 9,6%, цинка -
на 6,7%, магния - на 23,6% и железа - на 27,2%. При этом наблюдается улучшение
воспроизводительных функций животных: многоплодие увеличивается на 9,3%,
крупноплодность - на 9,2%, молочность - на 24,8%, на 23,2% снижается количество
слабых и на 65,3% мертвых поросят при рождении.
. Применение пикумина свиноматкам в течение второго периода супоросности
(с 84-го дня) и лактации позволяет повысить бактерицидную активность сыворотки
крови свиноматок - на 9,4%, фагоцитарную активность лейкоцитов - на 11,0%; в
крови возрастает содержание - гемоглобина - на 9,6%, общего белка - на 7,7%,
кальция - на 7,8%, магния - на 24,0% и железа - на 32,6%. При этом наблюдается
увеличение молочности свиноматок - на 9,1%, на 16,5% снижается количество
слабых и на 33,3% мертвых поросят при рождении.
. Установлена зависимость становления неспецифической защиты организма
приплода от уровня естественной резистентности организма матери. У поросят
возрастает бактерицидная активность сыворотки крови на 13,8%, лизоцимная
активность сыворотки крови - на 16,4%, фагоцитарная активность лейкоцитов - на
8,6%; при этом повышается энергия роста поросят на 8,5% и их сохранность - на
4,2%.
Указатель литературы
1. Алексеев В.А., Пыркина Л.В.
Эффективность использования аскорбиновой кислоты в кормлении свиней // Изв.
Нац. акад. наук и искусств Чуваш. Респ.- 1997.- №5.- С. 60-64.
2. Андросов В.А., Шабаев Н.В.
Естественная резистентность и иммунологическая реактивность организма свиней
при разных технологиях содержания // Сб. науч. тр // ВНИИ вет. сан., гигиены и
экол.- 1996.- 100.- С. 108-117.
3.
Бажов Г.,
Бахирева Л., Пищулин В. Фосфатиды в кормлении тяжелосупоросных свиноматок //
Свиноводство.- 2000.- №6.- С. 16-18.
4. Безмен В.А. Метод комплексной оценки
естественной резистентности у сельскохозяйственных животных // Исследования
молодых ученых в решении проблем животноводства: Материалы междунар.
научно-практической конференции молодых ученых и преподавателей, Витебск, 22-23
мая 2001 г. - Витебск, 2001.- С.10-11.
5. Беренштейн Ф.Я. Микроэлементы, их
биологическая роль и значение для животноводства.- Мн.: Госиздат БССР, 1958.-
С. 120-129.
6. Берзинь Я.М. Микроэлементы в
животноводстве.- Рига., Латвийское государственное издательство, 1961.- С.
43-82.
7. Буралга И.А. Продуктивность молодняка
свиней, его физиолого-биохимические и морфологические показатели при добавлении
в корм солей микроэлементов / / Комплексное использование биологически активных
веществ в кормлении сельскохозяйственных животных: БСХА.- 1974.- С. 344-347.
8. Бушов А.В., Сергатенко А.С., Тен Э.В.
Использование противоанемических средств в свиноводстве / / Биохим. аспекты
использ. хелат. структур происход. мет. в животновод. / Ульян. гос. с.-х. акад.
- Ульяновск, 1997. - С. 20-22.
9. Венедиктов А.М., Дуборезова Т.А.,
Симонов С.А. Кормовые добавки: Справочник.- 2-е изд., перераб. и доп.- М.:
Агропромиздат, 1992.- 192 с.
10. Водолазский М.Г., Касьянов М.С.,
Лемешко Т.Ф. Микроэлементы - фактор нормализации воспроизводства свиней //
Вестн. вет.- 1997.- №2.- С. 21-25.
11. Голушко В.М. и др. Кормовая добавка
// Сел. хоз-во Белоруссии.- 1982.- №10.
12. Голушко В.М. и др. Фосфогипс -
источник серы и кальция в рационах животных // Химия в сел. хоз-ве, Агрихем,
84: Тез. докл. 6-го конгр. Братислава, 25-27 июн.1984 г.
13. Давыдов Н.А., Андреев А.И., Лапшин
С.А. Роль цинка в питании ремонтных телок / / Огарев. чтения: Тез. докл. науч.
конфер., Саранск, 4-9 дек., 1995. Ч.2. - Саранск, 1995.- С. 156-157.
14. Елисеев И.Г., Карабанов А.М. Биохимические
показатели крови и развития внутренних органов поросят при скармливании
сапропеля // Белорусская сельскохозяйственная академия. Сб. науч. трудов.-
Горки, 1982. Вып.97: Минеральные подкормки в рационах животных., 1982.- С.
23-26.
15. Использование сапропелей в кормлении
животных.: Рекомендации.- Мн., БелНИИНТи, 1990.- 12 с.
16. Использование сфагнового торфа в
кормлении животных: (временные рекомендации).- Мн., 1980.- 23 с.- (БелНИИЖ.
Ин-т торфа АН БССР).
17. Кальницкий Б.Д. Минеральные вещества
в кормлении животных. - Л.: Агропроиздат, 1985. - 207 с.
18. Конопелько П.Я., Морозова Л.М.
Профилактика анемии поросят на промышленных комплексах / / Ветеринария. - 1989.
- № 10. - С. 45-46.
19. Концевенко В.В. Некоторые проблемы
нарушения минерального питания свиней в условиях промышленной технологии //
Профилактика нарушений обмена веществ и незаразных болезней молодняка
сельскохозяйственных животных: Материалы науч.-метод. конференции по
диагностике и терапии болезней с.-х. животных, 2-4 октября 1997 года /
Казанская ГАВМ.- Казань, 1998.- С. 32-33.
20. Концевенко В.В., Коган Э.С.
Резистентность поросят при нарушении минерального питания // Ветеринария.-
1985.- №5.- С. 59-60.
21. Кузнецов А.Ф., Сафронов Е.С.
Резистентность организма свиней при различном кормлении // Ветеринария. -
1977.- №6. - С. 35-38.
22. Лемеш В.Ф. Применение микроэлементов
в кормлении сельскохозяйственных животных / / Химизация животноводства. - Мн.,
1965. - 34-53.
23. Лумбунов С., Игнатьев Р. Природные
минералы для животных // Молоч. и мяс. скотовод. (Москва).- 1998.- №4.- С. 6-8.
24. Мантикова В.Г. Использование
древесного угля в кормлении поросят-сосунов // Пробл. и пути повыш. продуктив.
животновод. Сибири // РАСХН.СО. Сиб. Н.-и. и проект.-технол. ин-т
животновод.-Новосибирск, 1995.- С. 121-123.
25. Матюшевский Л.А., Коновалов Н.Н.
Влияние бентонита на резистентность поросят-сосунов // Проблемы повышения
резистентности животных: Сб. науч. тр // ВНИИ незараз. болезней животных.-
Воронеж, 1983.- 188 с.
26. Махаев Е.А. Переваримость, обмен
веществ и потребность в энергии растущих и откармливаемых свиней в зависимости
от температуры окружающей среды // Докл. Рос. акад. с.-х. наук.- 1998.- №2.- С.
33-36.
27. Медведский В.А. Использование
минеральной добавки пикумин в рационах отставших в росте поросят //
Свиноводство.- 1997.- №5.- С.21-22.
28. Медведский В.А. Продуктивность и
резистентность организма свиноматок при введении в рацион витаминов С и U // Зоотехния.- 1997.- №9.- С. 16-17.
29. Медведский В.А. Эффективная
минеральная добавка в рационы свиней // Весцi Акад. аграр. навук Беларусi.- 1997.- №3.- С. 53-55.
30. Медведский В.А., Свистун М.В., Вакар
А.Н. Использование биологических стимуляторов в свиноводстве: Аналит. Обзор.-
Мн.: Белорусский научный институт внедрения новых форм хозяйствования в АПК,
2002.- 48 с.
31. Надева Радка, Кънев Данаил. Влияние
на различните нива на калций и фосфор в смеските върху продуктивността на
подрастващи и уроявани прасета. Животновъд. Науки.- 1998.- 35, №5.- С. 30-35.
32. Николенко В.В. Динамика количественного
содержания лейкоцитов и эритроцитов у телят в связи с возрастом, сезоном и
применением хлорида кобальта и сульфата меди // Профилактика нарушения обмена
веществ у сельскохозяйственных животных.: Сб. науч. тр.- Одесса: ОСХИ, 1982.-
С. 41-44.
33. Олейников О.Г., Комарова Л.Н., Летов
И.И. Влияние инъекции препарата железа на продуктивность свиноматок и
жизнеспособность плода (помета) // Диагностика, лечение и профилактика
сельскохозяйственных животных // Ставроп. гос. с.-х. акад.- Ставрополь, 1995.-
С. 50-51.
34. Пестис В.К., Галушко В.М.
Использование фосфогипса в качестве минеральной подкормки для свиней. //
Минеральные подкормки в рационах сельскохозяйственных животных // Сб. науч.
тр., Вып. 97.- Горки, 1982.- С. 50-51.
35. Пилюк Н.В. Использование фосфогипса в
рационе молодняка крупного рогатого скота // Науч. основы развития ж-ва в
Республике Беларусь: Межведомств. сб.,- 1994.- Вып. 25.
36. Плященко С.И., Сидоров В.Т.
Естественная резистентность организма животных.- Л.: Колос, 1979.- 184 с.
37. Плященко С.И., Сидоров В.Т.
Предупреждение стрессов у сельскохозяйственных животных.- Мн.: Ураджай, 1983.- 136
с.
38. Плященко С.И., Сидоров В.Т., Сапего
В.И. Влияние факторов внешней среды на резистентность животных в условиях
современной технологии: Обзор. Информ.- Мн.: БелНИИТИ, 1980.- 39 с.
39. Плященко С.И., Хохлова И.И.
Микроклимат и продуктивность животных.- Л.: Колос.- (Ленингр. отд-ние), 1976.-
208 с.
40. Походня Г., Москалева Н., Абдурачимов
М. Влияние моциона на рост, развитие и воспроизводительные функции ремонтных
свинок // Свиноводство.- 1997.- №2.- С. 19-20.
41. Рубина М.В. Продуктивность свиней на
откорме при введении в рацион трепела и пикумина: Автореф. дис… канд. с.-х.
наук: 16.02.04.- Жодино, 2001.- 20 с.
42. Сапалева Е.Р. Влияние сапропелевой
подкомки на продуктивность свиноматок // Белорусская сельскохозяйственная
академия. Сб. науч. трудов.- Горки, 1982. Вып.97: Минеральные подкормки в
рационах животных., 1982.- С. 52-54.
43. Сапего В.И. Эффективность применения
микроэлементов в животноводстве.- Мн., 1985.- 46 с.
44. Слесарев И.К., Пилюк Н.В. Минеральные
источники Беларуси для животноводства.- Жодино-Минск, 1995.- 176 с.
45. Соколов Г.А. Ветеринарная гигиена. -
Мн.: Дизайн ПРО, 1998.- 160 с.
46. Соляник А.В., Соляник Т.В. Влияние
витамина С и фолиевой кислоты на продуктивность и неспецифическую реактивность
организма свиноматок // Фундаментальные и прикладные проблемы повышения
продуктивности сельскохозяйственных животных.: Материалы международной научной
конференции, посвященной 70-летию С.А. Лапшина, Саранск, 1998.- С. 91-92.
47. Торпаков Ф.Г. Зоогигиена в
промышленном свиноводстве.- Л.: Колос, 1980.- 229 с.
48. Филенко В.Ф., Орлянский Ю.А., Мамышев
С.А. Некоторые аспекты технологии племенного свиноводства // Вестн. вет.-
1998.- №2.- С. 21-24.
49. Шейко И.П., Смирнов В.С.
Свиноводство: Учеб. пособие для с.-х. вузов.- Мн.: Ураджай, 1997.- 352 с.
50. Bruckner J., Purice S.
Studii Si Cecretari fiziol. Acad PPR3, 1958, 15-19.
51. Chung Y. C., Huang
Chihpin, Tseng Ching-ping. Reduction of H2S/NH3 production from pig feces by
cont rolling environmental conditions // J. Environ. Sci. and Helth. A.- 1996.-
31, №1.- C. 139-155.
52. Close W.H. Housing systems
for optimum environment // Inf. Pig. Top.- 1996.- 11, №6.- C. 15-17.
53. Fife E.H., Mushel J.H. J.
Immunol. 87, №6, 1961.- P. 688-695.
54. Kornievicz A. Dodatki
paszowe w zywieniu trzody chlewnij / / Biul. inform. / / Inst. Zootechn. Zakl.
Inform. - Krakow, 1992. - № 190 / 193. - Р. 66-90.
55. Pestis Witold, Dobruk
Evgenij. Sapropeli w zywieniu svin // Ann. Warsaw. Agr. Vniv.- SCCW. Anim.
Sci.- 1999.- №36.- c. 117-123.
56. Prej J., Fuchs B.,
Schleicher A. The effect of carotene and vitamins A and E supplementation on
reproduction of sows // Arch. vet. pol.- 1993.- 33, №1-2.- C. 55-64.
57. Schweigert Florian J.
Neues zum Stoffwechsel und zur Wirkung vom
β-carotin und vitamin A // Kraftfutter.- 1995.- №6.- C. 271-272.
58. Vrbanac Ivan, Yammine
Rayanne, Krsnik Boris. Utjecai okolisa na trbnje patologije u intenzivnoj
svinjogojskoj proizodnji // Zb. sazet priopcen. 5 kongr. biol. Hrv., Pula, 3-7
okt., 1994.- Zagreb, 1994.- c. 233-234.
59. Xu Zirong, Wang Minqi //
Jiangxi nongue daxue xuebao= J. Zhejiang Agr. Univ.- 1999.- 25, №1.- c.
103-106.