Экологическая характеристика туводных рыб дельты Терека
Содержание
Введение
Глава I.
Общая характеристика исследования
.1 Состояние
изученности вопроса
1.2
Характеристика
места исследований и методических основ проведения исследований
Глава
II. Туводные рыбы
семейства карповых дельты Терека
.1
Проблемы
воспроизводства биоресурсов Каспийского бассейна
.2
Общая
характеристика туводных рыб дельты Терека
Глава III.
Годичные половые циклы и их особенности
Заключение
Список
использованной литературы
Введение
В данной работе рассматривается
экология размножения туводных рыб дельты р. Терек.
Актуальность темы определена
рядом факторов. В настоящее время, когда на многих внутренних водоемах
осуществляется комплексное использование водных ресурсов, необходимо сохранить
и приумножить запасы ценных видов рыб путем их эффективного воспроизводства.
Воспроизводство рыбных запасов
- единый процесс, который состоит из 2-х основных периодов: размножения рыб,
обеспечивающего восстановление численности в данном водоеме, и их нагула, в
результате которого образуется биомасса, составляющая собственно рыбные запасы.
Ухудшение условий размножения или нагула нарушает процесс воспроизводства
рыбных запасов. Дня его восстановления требуется проведение ряда мероприятий,
основными из которых являются:
размножение ценных промысловых
рыб путем улучшения естественных условий и при помощи искусственного
разведения;
улучшение видового состава
промысловых рыб в соответствии с особенностями водоемов;
улучшение режима рыбо-хозяйственных
водоемов как среды обитания.
Каждое из этих мероприятий по
воспроизводству рыбных запасов в естественных водоемах является самостоятельной
задачей, входящей в компетенцию рыбоводства.
Рыбоводство в естественных
водоемах предполагает проведение комплекса мероприятий, обеспечивающих процесс
воспроизводства рыбных запасов, их сохранение, увеличение и качественное
улучшение. Это достигается созданием благоприятных условий для размножения рыб,
путем искусственного их разведения, а также расширения ареала. Данные
мероприятия можно подразделить на 2 категории. Первая оказывает
непосредственное воздействие на рыбу - рыбоводные мероприятия, вторая - на ее
жизненную среду, т. е. мелиоративные мероприятия.
В качестве объекта исследования
выбраны рыбы семейства карповых - густера и красноперка. Большинство
классификаций относят их к туводным, хотя по ряду признаков они ближе к
полупроходным. Оба вида имеют второстепенное промысловое значение.
Предмет
исследования - разведение густеры и красноперки в водоёмах дельты Терека. В
данном случае используется практика ряда нерестово-выростных хозяйств (НВХ)
Кизлярского района.
Цель
данной работы состоит в определении экологии нереста красноперки и густеры, а задачи
исследования таковы:
1. охарактеризовать морфологию и
особенности нереста красноперки и густеры
. дать описание размножения и половых
циклик красноперки и густеры
Разведение этих рыб
осуществляется в нерестово-выростных хозяйствах, построенных в низовьях рек и
на водохранилищах.
Организация таких хозяйств
связана с ухудшением условий размножения рыб и падением их численности в результате
строительства гидроэлектростанций и зарегулирования стока рек.
Нерестово-выростные хозяйства позволяют направленно формировать ихтиофауну
водохранилищ, пополнять и увеличивать рыбные запасы рек благодаря созданию
благоприятных условий для нереста рыб.
НВХ представляют собой группу
водоемов, отделенных от реки или водохранилища возвышенными участками местности
или насыпными валами. Каждый из водоемов соединен с магистральным каналом, по
которому весной поступает вода из реки или водохранилища. Летом по каналу
производится сброс воды и скат выращенной молоди. Для полного сброса воды из
водоемов по их дну прокладывают сеть коллекторных канав. Подачу и сброс воды
регулируют при помощи шлюза, установленного на магистральном канале.
В зависимости от рельефа
местности водоемы заполняются водой самотеком или насосными станциями. В таких
хозяйствах нерест, инкубацию икры, выдерживание личинок до перехода на активное
питание и выращивание молоди рыб проводят в одном и том же водоеме. Условия
разведения рыб в этих хозяйствах близки к естественным, но более благоприятны:
постоянный уровень воды, отсутствие сорной и хищной рыбы. Кроме такой
упрощенной формы нерестово-выростных хозяйств существует и более совершенная,
предусматривающая проведение нереста, инкубацию икры, выдерживание личинок до
перехода на активное питание в одном водоеме, а выращивание молоди рыб в
другом.
При разведении туводных рыб
нерестово-выростные хозяйства располагают или на мелководье, или около
водохранилища (береговые) .
Береговые хозяйства бывают двух
типов. Первый тип представляет собой группу прудов, в которую входят маточные,
нерестовые и выростные. В маточных прудах осуществляют круглогодичное
содержание производителей. При наступлении нерестовых температур производителей
пересаживают в нерестовые пруды. Второй тип береговых хозяйств не имеет
нерестовых прудов. Они заменены инкубационным цехом и мальковыми прудами.
Работа состоит из трёх глав. Первая глава
посвящена обзору литературы и исследований по тематике работы. Во второй главе
даётся морфологическая характеристика густеры и красноперки, а в третьей
рассматриваются годичные половые циклы этих видов рыб.
Глава I.
Общая характеристика исследования
1.1
Состояние
изученности вопроса
Исследования по изучению
рыбоводства и рыбного населения водоемов Дагестана были начаты еще в конце
позапрошлого столетия (Россиков, 1895; Кузнецов, 1898; Соколов, 1878).
Некоторые данные о промысле и по биологии рыб водоемов Дагестана получили
освещение в последующих работах (Дмитриев, 1929; Глебов, 1932, 1941; Манкевич,
1932; Тамбовцев, 1932; Державин, 1932; Демин, 1932, 1946, 1947 и др.).
По утверждению этих авторов,
главное место в промысле из числа полупроходных видов рыб в водоемах Терской
системы в прошлом занимали сом, сазан, жерех, кутум и рыбец. В конце прошлого
века сазан был здесь в изобилии и фигурировал в уловах как «сор» (Кузнецов,
1898), а в 1928 г. он составлял около 19% от общего улова, занимая при этом
второе место после сома в промысловых уловах (Дмитриев, 1929). В 1931-1937 гг.
уловы сазана составляли от 25 до 32%, леща около 5-6%,
а воблы 1,2-4,2% от общего
улова (Демин, 1937). Вначале сом имел наибольшее промысловое значение среди
других видов рыб, составляя более 45% общего улова (Дмитриев, 1922), а в
последующие годы удельный вес его в общей добыче стал значительно снижаться и к
1937 г. дошел до 21,1% (Демин, 1946). Уловы жереха к 1935 г. определялись
порядка 7-7,5%, щуки и рыбца - 7% от общего улова по водоемам Дагестана (Демин,
1946). С 1901т., по сведениям А. Н. Державина (1949), уловы кутума в районе
Терека и Сулака достигали значительных величин. Так, в 1908- 191.1 гг. только в
Аграханском заливе и в бассейнах рек Терека, и Сулака кутума отлавливали от 6,0
до 10 тыс. ц в, год. Начиная с 50-х годов, уловы кутума в дагестанских водах
уже не исчислялись тысячами центнеров и в настоящее время (начиная с 1960- 1965
гг.) он по сути перестал в наших водах считаться промысловой рыбой и учет
ведется в общем улове частиковой рыбы. Уловы линя были незначительные (0,06%),
а красноперка и окунь вообще не имели промыслового значения (Демин, 1946).
По данным Д. 3. Демина (1946),
Т. X. Спасской (1972),
изучавших биологию размножения жереха, ход этого вида из моря через Аграханский
залив в реку и далее в озеро начинается в октябре и, усиливаясь по мере общего
похолодания, достигает своего осенне-зимнего максимума в конце ноября - начале
декабря.
Говоря о промысловом значении
линя, окуня и красноперки, Д. 3. Демин (1946), в частности, отмечает, что уловы
линя незначительные и обычно регистрируются совместно с другими мелкими видами
рыб, в качестве «мелочи» составляющими 4-5% общего улова. А красноперка и окунь
самостоятельного промыслового значения не имеют, поэтому их относят к прилову
или к «мелочи» (употребляется даже термин «сорные рыбы»; так, даже в тех
местах, где густера и красноперка имеют местное второстепенное промысловое
значение, они самостоятельно не учитываются и сдаются вместе с воблой и лещом).
Промысловая ихтиофауна
Аграханского залива впервые и более глубоко изучена А. К. Устарбековым.
Результаты его исследований опубликованы во многих литературных источниках.
Подробные сведения о промысле и
запасах многих видов рыб Дагестана приводятся в работах Ю. С. Саидова (1968,
1974). Им же изучены и паразиты рыб, встречающихся в водоемах Дагестана.
Эколого-морфологическая характеристика развития молоди некоторых видов рыб в
водоемах Дагестана дана в работе К. А. Аджимурадова (1972). Изучением кормовой
базы водоемов Дагестана занимался М. К. Гусейнов (1970) Растительноядные рыбы и
их эколого-физиологические особенности в водоемах Дагестана описаны М. О.
Омаровым (1970, 1972). Водная растительность водоемов Дагестана изучена Ф. М.
Магомаевым. Этим же автором проведены исследования по изучению возможностей
развития прудового рыбоводства, а также по акклиматизации северо-американских
рыб в водоемах Дагестана.
Работы по изучению питания
хищных и некоторых видов мирных рыб были выполнены 3. А. Юсуфовой, Н. М.
Мурзабековой и др. Гидробиологическими исследованиями в Северо-Западной части
Каспия занимался Г. А. Алигаджиев. Экономический анализ развития и размещения
рыбной промышленности Дагестана дан в многочисленных работах А. А. Надирадзе.
Паразиты и болезни рыб в водоемах Дагестана установлены многими исследователями
(Алигаджиев, Газимагомедов, Хайбуллаев, Атаев и др.). В последние годы
проводятся более глубокие исследования по изучению промысловых запасов рыб, их
биологии. Этому способствовало организованное в 60-х годах Дагестанское
отделение Каспийского научно-исследовательского института рыбного хозяйства
(КаспНИИРХ) на базе бывшей лаборатории промысловой ихтиологии Дагестанского
филиала АН СССР, которое в дальнейшем стало центром координации научных
исследований по ихтиологии и рыбоводству республики.
Таким образом, анализ
результатов ихтиологических исследований за период с 1890 г. и по сей день
показал, что хотя многие вопросы промысла и биологии отдельных видов рыб
Дагестана в определенной мере изучены, но биология размножения многих ценных в
промысловом и пищевом отношениях рыб в водоемах Дагестана все еще недостаточно
выяснена. В связи с зарегулированием стока рек, реконструкцией их придаточных
водоемов резко изменились и условия в них, необходимые для размножения рыб;
иным стал также количественный и качественный состав ихтиофауны придаточных водоемов
Дагестанского побережья Каспия, соответственно изменилась биология и, в
частности, биология размножения рыб, обитающих в этих условиях, а все это
требовало проведения комплексных и более глубоких ихтиологических исследований.
Такие вопросы, как эмбриогенез,
рост и развитие личинок и молоди(по этапам), скат и питание их в водоемах
Дагестана, изучались другими авторами (К. Аджимурадов, 3. Юсуфова, Т. Спасская,
Н. Мирзабекова и др.) и данные их работ опубликованы в различных сборниках и
журналах.
.2
Характеристика
места исследований и методических основ проведения исследований
Местом проведения исследований была выбрана
дельта реки Терек.
Дельта р. Терека играет важную
роль в естественном воспроизводстве рыбных запасов региона. Полноводность
Терека и развитость его речной сети, связывающей дельту с Северным Каспием,
обеспечивали на протяжении многих веков интенсивное воспроизводство ценных рыб.
Площади затопления, которые образовывались после разливов реки, колебались в
отдельные годы от 30 до 50 тыс. га. Ежегодные уловы достигали 16-19 тыс. т,
доля ценных рыб составляла более 80 %.
Со строительством Каргалинского
гидроузла в 1956 г. и увеличением забора значительных объемов воды на орошение
изменился гидрологический режим нижнего течения р. Терека. Сток воды уменьшился
с 12 до 6 км3 в год, что вместе с падением уровня Каспийского моря
привело к резкому сокращению обводняемых площадей дельты реки и появлению
реальной угрозы их полной потери. В результате большинство озер высохло, а их
незначительные сохранившиеся площади потеряли связь с рекой и морем, обмелели,
отшнуровались и зарастали жесткой и мягкой водной растительностью. Стал
невозможен доступ полупроходных и проходных рыб на нерестилища, а также скат
молоди и производителей обратно в море.
Создавшиеся неблагоприятные
условия воспроизводства отразились на запасах промысловых рыб. Значительно
сократилась добыча полупроходных рыб: сазана, леща, воблы, судака. Не были
освоены промыслом кутум, усач, рыбец, шемая, утрачены стада терского лосося,
тогда как еще в 1930-1940-х годах уловы этой ценной рыбы составляли 350-400 т в
год.
Начиная с 1961 г. объединение
«Дагрыба» проводило работы по обводнению многочисленных пойменных водоемов в
дельте Терека. В начале 1965 г. были построены и сданы в эксплуатацию Аракумские
нерестово-выростные водоемы (НВВ) площадью 16 тыс. га. Весной 1966 г. вступили
в строй 2 водоема Нижнетерской системы (13 тыс. га), в 1970 г. было закончено
строительство Каракольского водоема (13 тыс. га). Общая площадь перестроенных и
введенных в эксплуатацию в дельте р. Терека НВВ вместе с южной частью
Аграханского залива (12 тыс. га) составила 54 тыс. га (рис. 1).
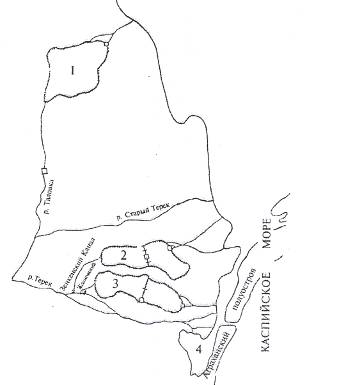
Рис. 1. Схематическая карта
яерестово-вырастных водоемов Дагестана
1
- Карагольский нвв, 2 - Аракумские нвв, 3 - Нижно-Терский нвн, 4 - Аграханский
залив
При проектировании НВВ были
поставлены следующие основные цели: создать водоемы с устойчивым регулируемым
гидрологическим режимом, что позволит установить оптимальные условия для
развития кормовой базы и создания возможности интенсивных нерестовых миграций и
нереста, улучшить условия нагула молоди рыб и обеспечить скат ее и
производителей в море. Проектом их предусматривалось использовать как
естественные нерестилища для воспроизводства ценных рыб в Каспийском море,
вылов предполагалось базировать в основном на местных, а также растительноядных
рыбах за счет массового вселения последних в водоемы, рыбопродуктивность
планировалась равной 1,2-1,7 ц/га.
Однако, как показывают
результаты исследований, указанные НВВ не дают проектного рыбопромыслового
эффекта. Это объясняется недостаточным водоснабжением и неудовлетворительной
эксплуатацией. Вследствие большего развития орошаемого земледелия в бассейне р.
Терека 80 % его стока потребляется для нужд сельского хозяйства и рыбоводство
испытывает нехватку водных ресурсов. В рыбохозяйственных целях в низовьях
Терека ниже Каргалинского гидроузла в весенне-летний период пропускается не
более 40 м3/с воды, что приводит к ухудшению гидрологического и
гидрохимического режимов, снижению кормовой базы, чрезмерному развитию высшей
водной растительности, заросли которой занимают до 80 % площади водоемов.
Предусмотренное проектом зарыбление водоемов растительноядными рыбами
проводилось только личинками, в редких случаях - небольшими партиями сеголеток,
что не дало промыслового и мелиоративного эффекта.
Основным источником
формирования ихтиофауны рассматриваемых водоемов явилось рыбное население
бывших пойменных озер, где в результате захода рыбы из Каспийского моря и р.
Терека сформировалась эвригалинная ихтиофауна: сочетание пресноводных видов
озерно-речного комплекса и некоторых полупроходных и проходных рыб. В водоемах,
связанных с Каспийским морем (Аракумские и Каракольский), больший удельный вес
имеют полупроходные рыбы, а в изолированной от моря южной части Аграханского
залива обитают в основном речные виды. Аналогичная с Южным Аграханом ихтиофауна
сложилась в конце 1980-х - начале 1990-х годов в Нижнетерских водоемах, имеющих
слабую связь с Каспийским морем.
Всего в составе ихтиофауны НВВ
в начале 1990-х годов насчитывался 21 вид, белоглазка, шемая, усач, чехонь
встречались лишь в рыбоходных каналах.
Основными промысловыми видами
во всех рассматриваемых водоемах в последние годы являются рыбы озерно-речного
комплекса (сом, щука, красноперка, линь, карась, окунь) и некоторые
полупроходные (лещ, сазан, судак), удельный вес первых в уловах составлял 85 %,
вторых - 15 %.
Исследования проводились в условиях
нерестово-выростных хозяйств дельты Терека (Аракумских Нижнетерских, Каракольского
нерестово-выростных водоёмов).
Исследование проводилось по следующим
направлениям:
. краткое описание и систематика
выбранных видов рыб - объектов изучения (густера и красноперка)
. анализ морфологических, в том числе
половозрастных и ихтиометрических характеристик (взаимозависимость длины, массы
тела, возраста и плодовитости). Для густеры было произведено изучение
упитанности и массы.
. изучение экологии нереста (время в
году, температура и др.)
Особое место в исследованиях занимало изучение
годичных половых циклов рыб и их особенностей для данных видов. Производилось
микроскопическое (гистологическое) изучение яичников и семенников
(макроскопическое изучение достаточно широко известно почти по всем описанным
выше источникам), это позволило выявить ряд особенностей в развитии половых
клеток в течение годичного полового цикла. Гистологическая картина всех шести
стадий зрелости у самок и самцов характеризуется наличием определенного
комплекса половых клеток, находящихся в разных фазах их роста и развития.
Наконец, при изучении использовалась
классификация рыб по различным признакам, составляющим экологию нереста.
Густера и красноперка попадали в группы, характерные для рыб с порционным
нерестом.
Глава II.
Туводные рыбы семейства карповых дельты Терека
.1 Проблемы воспроизводства
биоресурсов Каспийского бассейна
Россия, несмотря на распад СССР, занимает одно
из первых мест в мире по сырьевым запасам рыб. Двенадцать открытых морей,
омывающих границы России, крупнейшее в мире внутреннее море-озеро (Каспийское),
множество больших и малых рек и исключительное разнообразие климата обеспечили
благоприятные условия для обитания более 1000 видов рыб, из которых 250
являются рыбопромысловыми. Особенно богаты воды России наиболее ценными видами
рыб: осетровыми (более 90 % мирового улова) и карповыми (60 % мирового улова),
а также лососевыми и сельдевыми.
В настоящее время рыбо-хозяйственный комплекс
Каспийского моря и Прикаспия при незначительном удельном весе в общероссийских
уловах (около 3,5%) дает более 70% общероссийской добычи осетровых и более 60%
- крупночастиковых рыб. В последние годы наметилась тенденция роста вылова
рыбы.
Более 60% всех уловов в регионе приходится на
Каспийское море, пресноводные водоемы - 37,7%, пруды товарного рыбоводства -
2%. Основные промыслы - анчоусовая килька и крупный частик, которые являются
главными видами прироста уловов в последние годы. Уловы осетровых сохраняются
на уровне 0,5 тыс. тонн.
Общий фактический улов рыбы в Каспийском море и
внутренних водоемах Прикаспия составил в 2000 г. 174,2 тыс. тонн.
Основная часть российских уловов (88,2%)
приходится на Астраханскую область, на Республику Дагестан - 10,8% и Республику
Калмыкия - 1,0%.
Рыбохозяйственный потенциал Каспийского моря в
значительной степени зависит от поступления органического вещества с речным
стоком, режима последнего и от колебаний уровня моря. Наиболее продуктивной
частью моря является Северный Каспий, который обогащается волжской водой и
продуцирует около 20% органического вещества моря, занимая при этом лишь около
0,5% его объема. Регулирование речного стока каскадом водохранилищ на р. Волге,
результатом которого является увеличение зимних расходов в ее низовьях,
уменьшение объемов весеннего половодья и сокращение сроков затопления
нерестилищ осетровых и полупроходных рыб, отрицательно влияют на
биопродуктивность моря, на его рыбные запасы.
В настоящее время и в перспективе особую угрозу
для воспроизводства рыбных ресурсов моря и сохранения его экосистемы
представляет нарастающее загрязнение морской воды в результате хозяйственной
деятельности, в том числе проведение геологоразведочных работ, добыча нефти и
газа.
В случае сохранения уровня моря на современной
отметке, условий естественного и искусственного воспроизводства, аналогичных
последним годам, а также при низкой эффективности осуществляемых мероприятий по
охране рыбных запасов, объемы вылова осетровых в Волго-Каспийском бассейне в
ближайшие годы резко снизятся, а к 2010 г. их специализированный промысел может
быть прекращен.
Одной из основных причин катастрофической
ситуации с запасами осетровых является нелегальный промысел осетров в море.
Образование суверенных прикаспийских государств, новые рыночные отношения
разрушили сложившуюся в течение десятилетий систему рыболовства на Каспии.
Распалась также мощная единая система рыбоохраны. Повышенный спрос на осетровые
балыки и черную икру, как на внешнем, так и на внутреннем рынках породил
небывалый браконьерский промысел не только на реках, но и в море. По некоторым
оценкам, от браконьерского лова осетровых Россия ежегодно теряет сотни
миллионов долларов. Однако в данном случае речь идет не столько о потере экспортной
выручки, сколько о сохранении реликтового биологического ресурса Каспия.
Необходимо также увеличить производство в
искусственных условиях не менее чем 120 млн. штук рыбной молоди. Сегодня в
России на астраханских рыбоводных заводах производится около 60 млн. штук
молоди осетровых, в других государствах каспийского региона - еще 29, строится
осетровый завод на Дону, но этого явно недостаточно.
Увеличение уловов осетровых может произойти
только за счет осуществления комплекса достаточно дорогостоящих рыбоводных
мероприятий, а также улучшения состояния речного стока и моря. Большая часть
мероприятий, предусмотренных Федеральной программой развития рыбного хозяйства
России на период до 2000 года («Рыба») за счет бюджетных средств, не выполнена.
Поэтому часть этих мероприятий включена в настоящую Программу. При выполнении
намеченных мероприятий уловы осетровых могут возрасти до 3,0 тыс. т в 2010
году.
На фоне прогрессирующего снижения эффективности
естественного размножения осетровых в Волге и других реках Каспийского бассейна
сохранение их популяционного фонда и промыслового значения в сложившихся
условиях можно обеспечить только за счет интенсификации и повышения
эффективности заводского воспроизводства.
На действующем в настоящее время в Республике
Дагестан Сулакском осетровом рыбоводном заводе выпуск молоди осетра и севрюги в
последние годы снизился из-за дефицита зрелых производителей осетровых.
Построенные в 60-х годах осетровые рыбоводные
заводы Астраханской области в настоящее время приходят в упадок. Они нуждаются
в срочной реконструкции, смене оборудования, корректировке биотехнологии.
Несмотря на это, Астраханская область на сегодняшний день является практически
единственным регионом на Каспии, занимающимся воспроизводством осетровых на
площадях 6 заводов, здесь сосредоточены научные силы, занимающиеся проблемой
воспроизводства и сохранения биоресурсов Каспийского моря.
Гидрологические изменения, связанные с
повышенным уровнем Каспийского моря, оказали значительное влияние на
распределение килек: снизилась ее концентрация в районах традиционных
промысловых глубин, что осложняет промысел. До 2005 г. можно ожидать некоторого
увеличения запасов анчоусовидной кильки, стабилизации запасов большеглазой
кильки и рост запасов обыкновенной кильки. Данная тенденция сохранится в
условиях современного уровня Каспия и несколько усилится при его повышении.
Резервом увеличения объемов морского промысла, в том числе российского,
являются недоиспользуемые запасы обыкновенной и большеглазой кильки.
Состояние запасов воблы оценивается как
благополучное, промысловый запас предполагается на относительно высоком уровне,
в том числе в пределах вод России около 50 тыс. т. Высокий уровень запасов и
уловов леща в 1999-2000 гг., позволяет рассчитывать на их дальнейшее увеличение.
Наблюдается снижение уловов судака в северо-западной части Каспия в результате
сохраняющейся тенденции перехода судака в северо-восточную часть моря, а также
увеличивающегося пресса браконьерства.
Чрезвычайно важным для сохранения уникальной
экосистемы Каспийского моря, ее биоресурсов, особенно ценных осетровых,
является решение проблемы правового статуса Каспийского моря, которое бы
предусматривало совместное использование биоресурсов моря прикаспийскими
государствами, взаимные обязательства по их воспроизводству и охране и положило
конец браконьерству.
Рыбохозяйственный фонд внутренних водоемов
включает водоемы западно-подстепных ильменей дельты р. Волги общей площадью
около 500 тыс. га. Около половины из них потеряли рыбохозяйственное значение в
результате засоления. В то же время 60 тыс. га водоемов, прилегающих к р. Волге
и ее рукавам, имеют большое значение как нерестовый фонд дельты Волги и
нуждаются в особой охране. Повышение рыбохозяйственной значимости ильменей
связано с проведением их мелиорации, с дноуглубительными работами на малых
реках (Хордун, Бертюль, Бушма и т.д.), что может увеличить их
рыбопродуктивность в 1,5-2 раза.
Волго-Ахтубинская пойма до ограничения
рыболовства в 1958-1962 гг. имела важное промысловое значение. В настоящее
время пойма и дельта Волги рассматриваются как единая экосистема для
воспроизводства полупроходных и туводных рыб. Нерестилища Волго-Ахтубинской
поймы в пределах Астраханской области обеспечивают промысловый возврат в объеме
около 30,0 тыс. тонн. Для сохранения Волго-Ахтубинской поймы как места
воспроизводства ценных видов рыб необходимо жестко ограничить развитие в этом
регионе орошаемого земледелия.
В Республике Дагестан воспроизводство запасов
ценных проходных и полупроходных рыб сосредоточено преимущественно в нерестово-выростных
водоемах (НВВ) в бассейне р. Терек. Общая проектная мощность НВВ составляет 54
тыс. га. За последние годы промысловые запасы рыбы в этих водоемах снизились в
2,5-12 раз. Для повышения рыбопродуктивности водоемов необходимо осуществить их
реконструкцию и мелиоративные работы, в том числе зарыбление растительноядными
видами.
В республике имеется единственный полностью
спускаемый Самурский нерестово-выростной водоем, в котором осуществляется
естественное воспроизводство кутума и других частиковых рыб. Продуктивность
этого водоема составляет 50-70 кг/га, что в десятки раз выше, чем других
придаточных водоемов дельты Терека.
Общая водная площадь внутренних водоемов
Республики Калмыкия составляет 53,7 тыс. га, из них промыслом охвачено 36,7 тыс.
га. Рыбопродуктивность внутренних водоемов незначительна и имеет тенденцию к
снижению.
В целях сохранения и воспроизводства ценных
пород рыб намечено осуществить ряд мероприятий.
В целях воспроизводства рыбных ресурсов
Каспийского моря и развития сырьевой базы рыбохозяйственного комплекса
республики увеличиваются мощности по воспроизводству осетровых на Сулакском
осетровом рыбоводном заводах на 20 млн. шт. молоди; реконструируются
Нижне-Терские, Каракольские и Аракумские нерестово-выростные водоемы для восстановления
численности осетровых и частиковых рыб; расширяются нерестово-выростные
хозяйства в Тарумовском районе и увеличиваются мощности рыбоводных заводов в
Кизлярском и Магарамкентском районах, будет построен Нижне-Терский осетровый
рыбоводный завод.
Как показали многолетние
наблюдения, эффективность воспроизводсва рыб, особенно полупроходных, находится
в прямой зависимости от расхода воды в Тереке в каждом году. В годы с хорошим
водоснабжением водоемов урожайность молоди возрастает, в маловодные годы
снижается.
В последние годы фактические
расходы воды в апреле-июне для рыбного хозяйства низовий Терека не превышают 30
% от требуемого количества. Так, в апреле-июле в нижнем бьефе Каргалинского
гидроузла расход воды составляет 20-30 м3/с, что не обеспечивает
обводнения низовий и водоснабжения Аракумских, Нижнетерских, Каракольского НВВ
и других рыбохозяйственных объектов. При расходах менее 40 м3/с
водоподающие каналы, особенно Нижнетерских НВВ, не могут производить забор воды
из Терека, а малые глубины (10-15 см) на участках нижнего течения не
обеспечивают свободного прохода рыбы вверх по реке. Особенно недостаточны
рыбохо-зяйственные попуски воды в нерестовый период в марте-июне, когда только
на транзитные расходы воды для миграции рыб на нерест требуется подавать в
нижний бьеф Каргалинского гидроузла не менее 80-100 м3/с (при
фактическом расходе 20-30 м3/с).
В результате сложившегося
положения рыбное хозяйство несет большие потери. Если в 1950-е годы урожайность
пойменных водоемов дельты р. Терека составляла 1,5-2,5 ц/га и в некоторых
случаях - 4 ц/га, то в настоящее время рыбопродуктивность НВВ не превышает 2-3
кг/га, т. е. снизилась в 75-100 раз.
Кроме гидрометеорологических
параметров, на численность поколения каждого года влияют условия захода производителей
промысловых рыб из моря по рыбоходным каналам. В последнее время оказатась
заилены устья рыбоходов Аракумских и Нижнетерских водоемов, в результате чего
заход производителей из моря крайне затруднен. Нерестовые угодья фактически
непроточны, в связи с чем недостаточно нерестового субстрата для кладки икры.
Полупроходные рыбы вынуждены откладывать икру в тростниковых зарослях, т. к.
свежезалитая луговая растительность в водоемах отсутствует. В связи с чем
происходит совмещение нерестовых ниш у рыб различных систематических групп.
Вследствие резких суточных колебаний уровня воды на нерестилищах погибает
отложенная икра, что резко снижает эффективность естественного размножения рыб.
В результате скат молоди из НВВ с начала 1970-х до 1990-х годов снизился с
200-300 до 20-30 млн шт. в год (Магомаев, Столяров, 1993).
В последние годы в НВХ проведены работы по
повышению эффективности воспроизводства ценных промысловых рыб:
реконструированы Аракумские водоемы, мелиоративные работы в Каракольском
водоеме позволили решить проблему водообеспечения и проводить весной
необходимый сброс воды по рыбоходу для привлечения производителей. В результате
скат молоди в последние годы заметно увеличивается и достиг в 2002 году 60 млн
шт.
2.2.Общая характеристика туводных рыб
дельты Терека
Семейство карповых самое многочисленное,
характеризующееся наличием у рыб глоточных зубов, расположенных в 1-3 ряда,
трущихся о жерновик (роговидное образование). Усиков или нет, или не более двух
пар. Тело обычно покрыто чешуей, реже голое. Живут в быстротекущих потоках
(маринки, усачи, ельцы и др.), в озерах и прудах. Держатся как в толще воды,
так и у дна стоячих (линь, карась) и текучих вод (пескари, гольяны). Живут
также в солоноватой воде в Каспийском и Аральском морях.
Большинство видов карповых не совершают далеких
передвижений для нереста. Однако есть и типичные проходные рыбы, нагуливающиеся
в солоноватых водах, а для икрометания поднимающиеся вверх по рекам иногда
более чем на тысячу километров (вырезуб, кутум, аральский усач, каспийская и
азовская шемая и др.).
Молодь карповых питается зоопланктоном. Во
взрослом состоянии основная масса карповых питается зообентосом - животными,
сидящими на грунте (язь, плотва, черный амур и др.) и закапывающимися в грунт
(сазан, линь, карась и др.), а также зоопланктоном (уклея, амурская
востробрюшка и др.). Среди карповых есть хищники (жерех, желтощек, верхогляд,
голавль и др.). Высшую растительность поедают белый амур, красноперка.
Фитопланктоном питается толстолобик. Большинство карповых интенсивно питаются в
теплое время года и почти или полностью прекращают питание в холодное время.
Из карповых ряд видов являются объектами
искусственного рыборазведения в нерестово-выростных хозяйствах и рыбопитомниках
во многих регионах страны.
Густера (Blicca
bjoerkna) - единственный вид подсемейства густер семейства карповых
(Cyprinidae). Известна под различными обозначениями:
|
Cyprinus
Bjoerkna
|
Linnaeus,
1758: 326 (озера в Швеции)
|
|
Cyprinus
gibbosus
|
Pallas,
1814: 324 (Сура, Волга).
|
|
Cyprinus
laskyr
|
Pallas,
1814: 326 (Днестр, Дон, Сейм).
|
|
Blicca
bjoerkna
|
-
Heckel, 1843: 1032; - Кесслер, 1864: 98 (Невская губа, Нева, Ладожское озеро,
Волхов) ; - Берг, 1932: 494.
|
|
Blicca
argyroleuca
|
Heckel
et Kner, 1858: 120 (Нева, Австрия, Венгрия и др.)
|
|
Blicca
bjoerkna
|
-
Домрачев и Правдин, 1926: 180 (Ильмень, Волхов) ; - Берг, 1949: 763.
|
|
Abramis
bjoerkna
|
Kottelat,
1997: 42.
|
|
Blicca
bjoerkna
|
-
Решетников и др., 1997: 735; - Аннотированный каталог, 1998: 58 (структура
вида);
|
|
-
Атлас пресноводных рыб России, 2002: 219 (ареал).
|
Описание и систематика.
Тело высокое, с заметным горбом, сильно уплощенное с боков. Хвостовой плавник
сильно выемчатый, лопасти его приблизительно одинаковой длины. Голова
маленькая, глаз относительно большой. Рот косой, полунижний, маленький,
выдвигающийся в виде трубки, направленной вниз. Позади брюшных плавников есть
киль, не покрытый чешуей. На спине за головой чешуя с боков тела не смыкается,
и на гребне спины образуется не покрытая чешуей бороздка. Чешуя на затылке
крупнее, чем у леща. Чешуя толстая, плотно сидящая, от боковой линии вверх не
уменьшается в размерах. Окраска спины голубовато-серая, бока серебристые,
непарные плавники серые, грудные и брюшные в основании красноватые. Во время
нереста у самцов появляется брачная окраска (белые бугорки на голове и ярче
плавники). D III (7) 8-9, A III 19-23. В боковой линии 40-51 чешуй. Жаберных
тычинок 14-21. Глоточные зубы двурядные, 2.5-5.2 или 3.5-5.2. Позвонков 40-42
(Берг, 1949; Рыбы Подмосковья, 1988; Атлас пресноводных рыб России). Кариотип:
2n=50, NF = 90 (Васильев, 1985).
Иногда этот вид включают в род Abramis (Шутов,
1969; Kottelat, 1997), но чаще считают валидным на основании анатомических
признаков (Богуцкая, 1986). Полагают, что в России обитает 3 подвида: кроме
номинативного подвида, занимающего основной ареал вида, выделяются еще B.
bjoerkna transcaucasica Berg, 1916 (Кура, Аракс, реки Ленкорани) и B. bjoerkna
derjavini Dadikjan, 1970 (Севджур в бассейне Аракса) (Аннотированный
каталог...,1998; Атлас пресноводных рыб России, 2002).

Рис 2. Густера
Густера отличается от родственных ей
видов лещей исключительно числом и расположением глоточных зубов, которых
находится с каждой стороны не по пяти, а по семи и притом в два ряда. Формой
тела она весьма похожа на молодого леща или, вернее, подлещика, но имеет
меньшее число лучей в спинном (3 простых и 8 ветвистых) и заднепроходном (3
простых и 20-24 ветвистых) плавниках; кроме того, чешуя у нее заметно крупнее,
а парные плавники имеют красноватый цвет. Тело густеры сильно сплющено, и
вышина его составляет не менее трети всей длины его; нос у нее тупой, глаза
большие, серебристые; спина голубовато-серая, бока туловища
голубовато-серебристые; непарные плавники серые, а парные при основании красные
или красноватые, к вершине темно-серые. Впрочем, эта рыба, смотря по возрасту,
времени года и местным условиям, представляет значительные видоизменения.
Распространение. Широко
распространена в Европе к востоку от Пиренеев и к северу от Альп и Балкан. Обитает
в реках и озерах бассейнов Северного, Балтийского, Черного, Азовского и
Каспийского морей. Из бассейна Белого моря густера отмечена в озерах бассейна
рек Онега и Северная Двина, редка в самой Северной Двине и ее притоках (Атлас
пресноводных рыб России, 2002).
Максимальные размеры. Живет не
более 15 лет, достигает длины 35 см и массы 1,2 кг, но обычно - до 25-30 см и
0,5 кг.
Образ жизни. Густера, как
и лещ, предпочитает водоемы со слабо проточной или непроточной водой, хорошо
развитой растительностью и заиленным или глинистым дном. Это теплолюбивый вид,
обитающий в водоемах с температурой в летнее время не ниже 16-17оС.
Малоподвижная рыба, долго держится на одном месте. Обычно предпочитает хорошо
прогреваемые мелководья с зарослями растительности. Весной и осенью густера
образует многочисленные стаи или густые скопления, откуда и произошло название.
Характер питания взрослых рыб зависит от типа водоема. Густера потребляет в
основном личинок хирономид, ручейников, моллюсков, а также часто водоросли и детрит.
Незначительную долю в откорме составляют воздушные насекомые и высшая
растительность. Растет густера медленно. Половой зрелости достигает в возрасте
3-4 лет при длине 11-15 см. Самцы созревают обычно на 1-2 года раньше самок и
при меньших размерах. Во время нереста у самцов голова и бока тела покрываются
эпителиальными беловатыми бугорками, а парные плавники становятся красноватыми.
Самки выметывают 2-3 порции икры с промежутком в 10-15 дней. Диаметр икринок
первой порции перед выметом 0,8-1,2 мм, второй 0,4-0,7 мм и третьей 0,2-0,3 мм.
В водохранилищах количество порций
меньше, и часть самок переходит к единовременному нересту. Нерест порционный,
происходит в конце мая - начале июня при температуре воды 15-17о С и
продолжается 1-1,5 мес. Нерест шумный, происходит ночью. Икра клейкая,
откладывается на затопленную растительность, подмытые корни растений на глубине
20-60 см. Плодовитость - 11-109 тыс. икринок. Развитие икры длится 4-6 сут.
Личинки до рассасывания желточного мешка прикрепляются к водной растительности.
Молодь сначала питается фито- и зоопланктоном, затем мелкими бентосными формами
(Промысловые рыбы СССР, 1949; Рыбы Подмосковья, 1988). Образует помеси с лещом,
плотвой, красноперкой, уклейкой и рыбцом.
Густера - рыба вялая, ленивая и,
подобно лещу, любит воду тихую, глубокую, довольно теплую, с иловатым или
глинистым дном, почему очень часто встречается с этим последним. Она долго
живет на одном месте и всего охотнее держится у самых берегов, особенно в
ветер, так как валы, размывая берега, а на мелких местах самое дно,
обнаруживают различных червяков и личинок. В небольшом количестве она,
по-видимому, держится в устьях рек и на самом взморье, как, например, в устьях
Волги и в Финском заливе между Петербургом и Кронштадтом. Весною и осенью густера
встречается чрезвычайно густыми стаями, откуда, конечно, и произошло ее
общеупотребительное название. Впрочем, она редко совершает очень дальние
странствия и почти не доходит, например, до среднего течения Волги, где живет
уже своя, местовая, густера. Вообще главная масса этих рыб скопляется в
низовьях рек, в море, и, подобно весьма многим другим, она совершает правильные
периодические движения: весною идут вверх для нереста, осенью - для зимовки.
Входя осенью на зимовку, они ложатся на ямы под перекаты такими большими
массами, что в низовьях Волги случается в одну тоню вытаскивать до 30 тысяч
штук. Пища густеры почти одинаковая с другими видами лещей: она кормится
исключительно тиною и заключенными в ней мелкими моллюсками, рачками и
червяками, чаще всего мотылем, но также истребляет и яйца других рыб, особенно
(по наблюдениям Блоха) икру красноперки.
Нерест густеры начинается очень
поздно, большей частью по окончании нереста леща - в конце мая или в начале
июня, на юге несколько ранее. В это время чешуя у нее изменяется в цвете, и
парные плавники получают более яркий красный цвет; у самцов, кроме того, на
жаберных крышках и по краям чешуи развиваются маленькие зерновидные бугорочки,
которые потом опять исчезают. Обыкновенно мелкая густера мечет икру раньше,
крупная позже. По наблюдениям же Блоха, в Германии сначала мечет самая крупная
густера, через неделю или девять дней - мелкая. Местом нереста густера выбирает
травянистые и мелкие заливы и выметывает икру чрезвычайно шумно, подобно лещу,
но несравненно смирнее его: в это время иногда даже случается ловить ее руками.
Нерестится она обыкновенно с захода солнца до десяти часов утра, и каждый
возраст кончает игру в 3 - 4 часа ночи, но если помешает холодная погода, то в
один день. В самке средней величины Блох насчитал более 100 тыс. икринок. По
Зибольду, густера становится способной к размножению очень рано, еще не
достигнув 5 дюймов (13 см) длины, так что надо полагать, что она нерестится
обычно по второму году.
Статус вида.
Малоценная промысловая рыба. Мясо костлявое и имеет низкие вкусовые качества.
Вылавливается в летние месяцы, в статистике отдельно не учитывается и сдается
вместе с лещом и воблой.
Красноперка (Scardinius
erithrophthalmus). Семейство Карповые Cyprinidae
|
Cyprinus
Erythrophthalmus
|
Linnaeus,
1758: 324 (Северная Европа).
|
|
Cyprinus
erythrops
|
Pallas,
1814: 317 (Россия и Сибирь).
|
|
Leuciscus
erythrophthalmus
|
-
Кесслер, 1856: 48 (юго-запад России).
|
|
Scardinius
erythrophthalmus
|
Heckel
et Kner, 1858: 153 (центр. Европа);
|
|
-
Берг, 1912: 270; - Берг, 1949: 593.
|
|
-
Аннотированный каталог, 1998: 71 (распространение);
|
Описание и систематика.
Тело умеренно удлиненное и уплощенное с боков. На брюхе есть киль, покрытый
чешуей. Перед спинным плавником спина сжата с боков. Рот конечный, но обращен
вверх. Окраска тела яркая, особенно во время нереста. Глаза оранжевые, с
красным пятном вверху. Плавники красные, кроме спинного. Спинной плавник серый
с красноватой вершиной. D III 7-10, A III 9-12. Чешуя крупная, в боковой линии
37-43 чешуи. Жаберных тычинок (8) 10-12. Глоточные зубы двурядные, пилообразно
зазубренные, 3.5-5.3. Позвонков 37-40. Кариотип: 2n = 50, NF = 88 (Васильев,
1985). Подвиды не выделяются (Берг, 1949; Аннотированный каталог, 1998).
По Л. Сабанееву: «С первого взгляда красноперка
представляет большое сходство с плотвой и, вероятно, часто принимается за
последнюю. Но, не говоря о том, что красноперка гораздо красивее плотицы, имеет
совсем другие глоточные зубы, она легко может быть отличена по своему косому
рту, направленному вверх, по острому выдающемуся ребру на брюхе, по желтой
каемке на губах, более мелкой и золотистой чешуе, а также по тому, что спинной
плавник у нее начинается не над брюшным, как у плотвы, но позади его. Кроме
того, красноперка не покрыта слизью, подобно плотве».

Рис.3. Красноперка (обычный вид)
У красноперки с каждой стороны
находится по 8 зубов, расположенных в 2 ряда, между тем как у плотвы только 6,
даже 5 зубов; брюхо, начиная от брюшных плавников до заднепроходного отверстия,
образует выдающееся ребро, покрытое чешуями. Красноперка притом всегда
несколько толще и шире плотвы и складом более походит на подлещика; кроме того,
она желтее, а плавники и глаза ее имеют гораздо более яркий цвет; спина у нее
коричневато-зеленая, бока туловища блестящего желтовато-золотистого цвета, а
края чешуи с золотисто-коричневой каймой; глаза оранжевые с красным пятном в
верхней половине; спинной плавник при основании черноватый, на вершине красный,
грудные - серые, на вершине тоже искрасна, а все остальные - кровяно-красные,
даже малиновые. Вообще, как видно из описания, красноперка одна из самых
красивых наших рыб; особенно ярки цвета у крупных во время нереста, молодые же
гораздо серебристое, светлее и плавники у них заметно бледнее. Очень крупные
красноперки (свыше 800 г) имеют более темную окраску.

Рис 4. Глоточные зубы красноперки.
Распространение. Повсеместно
в Европе, к востоку от Пиренеев. Из рек бассейна Северного Ледовитого океана
отмечена только в некоторых озерах Архангельской области (р. Вашка в бассейне
Северной Двины). Нет в Крыму и в северной части Скандинавского полуострова.
Есть в Малой Азии, Западном и Восточном Закавказье, в бассейне Аральского моря
и в реке Чу.
Главное местопребывание красноперки
- заливы и старицы рек, также проточные пруды и озера, где в изобилии растет
камыш, тростник и другие водяные растения. Здесь она довольно часто встречается
в сообществе с карасями, линями и лещами и ведет почти оседлую жизнь, редко
отдаляясь от раз избранного ею места. Поэтому она встречается далеко не всюду,
а местами, и где весьма многочисленна, где составляет большую редкость. Во
всяком случае, она, по-видимому, гораздо обыкновеннее в юго-западной и южной
России, чем в средних губерниях. Так, например, в низовьях Волги она держится
во множестве во всех ильменях и тихих заросших протоках, весьма обыкновенна
также во всех больших и средних озерах западной России. В подмосковных
губерниях она держится преимущественно в проточных прудах, хотя в весьма
немногих, но зато в большом количестве - в заливных озерах и старицах.
В Каспийском бассейне - Scardinius
erythrophthalmus Linne - пресноводная рыба, в море
встречается в предустьевой зоне Волги, Урала и Терека. После зарегулирования
стока Волги в низовьях дельты, култучной зоне и авандельте создались
благоприятные условия для обитания этого вида и численность красноперки
возросла [Тряпицина, 1965, 1975].
Нерестится красноперка на
мелководных, малопроточных, заросших растительностью участках полоев, ильменей,
култуков, а также на мелководьях авандельты с мая по июль при температуре воды
19-23°С [Коблицкая, 1959]. В годы с ранней
весной личинки красноперки появляются на нерестилищах дельты во второй половине
мая. Наибольшие их концентрации наблюдаются в июне.
В период максимальной
концентрации численность личинок на нерестилищах средней зоны дельты колеблется
в разные годы в очень широких пределах - от 0,3 до 9,3 тыс. экз./га на западе и
от 2,0 до 35 тыс. экз./га на востоке. Наибольшая эффективность нереста в
средней зоне отмечается, как правило, в годы с большим объемом весеннего
половодья. В култучной зоне и авандельте, где расположены основные нерестовые
биотопы красноперки, ее нерест практически не зависит от характера половодья и
попусков воды.
Темп линейного роста
красноперки довольно высок. К концу первого года жизни она достигает средней
длины 4,2 см, в возрасте 2 лет - 9 см, в 3-летнем возрасте -- 18,5 см
[Тряпицина, 1975]. Молодь красноперки питается зоопланктоном и водорослями. В
питании взрослых рыб преобладает (90-100%) растительная пища.
Красноперка предпочитает озера. В реках выбирает
места со слабым течением, заросшие камышом и другими растениями. Питается
главным образом нитчатыми водорослями, но взрослые могут поедать и животную
пищу (ракообразные, икра моллюсков, личинки насекомых, черви, икра и мальки
рыб). Половой зрелости достигает на 3-5-м году жизни при длине более 12 см.
Нерест порционный, происходит в апреле-июне при температуре воды 18о С. Икру
откладывает на водные растения. Икринки клейкие, диаметром 1,5 мм. Плодовитость
4-232 тыс. икринок (Лебедев и др., 1969; Тряпицына, 1975). Инкубационный период
короткий, не более 3 сут. Личинки при выклеве имеют длину 5 мм. Молодь сначала
питается зоопланктоном, потом переходит на растительную пищу. Мальковый период
начинается при достижении длины 30 мм (Макеева, Павлов, 1998).

Рис.5. Краснопёрка - вид во время
нереста
У самцов во время нереста на голове
и спине выступают зернистые бородавочки, и все красноперки принимают более
яркие цвета; но (сравнительно) внешние покровы красноперки изменяются мало, и
чешуя у них не получает шероховатости, особенно резко выраженной у плотвы. По
Эренкрейцу, сначала к месту нереста идет стайка самцов, потом стайка самок и за
ними снова самцы; каждая стая идет густой массой и отдельно одна от другой.
Крупная красноперка, кажется, нерестится раньше - в камышах и тростнике,
средняя и мелкая - всегда в июне; икра выпускается в несколько приемов, а не
сразу. Самый процесс икрометания совершается весьма тихо, небольшими партиями и
его трудно заметить, тем более, что красноперки трутся (хотя днем) почти на дне
и не выпрыгивают на поверхность, а только всплескивают. Только качание камышовых
и тростниковых стеблей указывает на место нереста. Мелкая красноперка, кажется,
иногда трется и в водяном мху, но никогда под берегом. Вообще икру красноперки
разыскать почти так же трудно, как икру линя, и надо полагать, что она
прикрепляется б. ч. к корням водяных растений. Число икринок довольно
значительно (в 400-граммовой самке насчитывается до 100 тысяч), но, вероятно,
значительная часть икры пропадает, так как молоди красноперки всегда бывает
меньше, чем молодой плотвы, от которой первая отличается с первого взгляда по
своему общему красноватому цвету; кроме того, стайки молодой красноперки
никогда не бывают многочисленны. Держатся они, конечно, в траве, всегда на
солнечной стороне и почти у самой поверхности воды. Молодая красноперка растет
довольно быстро и к осени уже обгоняет плотичек-сеголетков, выведшихся месяцем
ранее. Через год, в июле, красноперка имеет в длину иногда более 4 см (в не
очень кормных водах), а двухгодовалая - до 9 см, и, по-видимому, уже мечет
икру. Впрочем, прирост, как всегда, неодинаков в различных местностях, и даже
годами в прудах рыба растет еще медленнее.
В промысловых уловах
красноперка встречается начиная со второго года жизни. Основу уловов (63-95%)
составляют 4-6-годовики длиной 18,8-27,0 см и массой 124-500 г (табл. 84).
Возрастная структура промысловой части популяции красноперки существенно
изменяется по годам и зависит от урожайности вступающих в промысел поколений.
Средний возраст рыб в промысловых уловах 4,6-5,5 лет. В последние годы (1981 -
1984) средний возраст красноперки был относительно высоким (5,1-5,5) лет) за
счет большого количества рыб старших возрастных групп - остатка от урожайных
поколений 1975, 1976, 1977 гг. Изменение возрастной структуры популяции
отразилось и на размерном составе. С 1981 по 1983 г. увеличилась доля крупных
рыб (24-29 см) с 17,1 до 44,3%.
Одновременно происходило
снижение длины и массы одновозрастных рыб (5-8-годовики). При стабильной
кормовой базе снижение линейно-весовых показателей одновозрастных групп
красноперки может быть результатом возросшей ее численности.
Средняя относительная
численность самок в нерестовой популяции 1976-1984 гг. составляла 70,0-82,7%. У
трехгодовиков соотношение полов приближается 1:1. В старших возрастных группах
преобладают самки.
В начале 50-х годов уловы
красноперки составляли 2,4 тыс. т в год. В настоящее время красноперка - одна
из наиболее распространенных и многочисленных рыб в волжском предустьевом
пространстве, где она добывается практически в течение всего года. Доля ее в
уловах мелкого частика колеблется от 29,2 до 65,4%. В 1966-1971 гг. уловы
составляли в среднем около 6,3 тыс. т, в 1972-1974 гг. они возросли до 10,2
тыс. т.
С 1976 по 1978 г. наблюдалось
уменьшение промыслового запаса красноперки, так как основу его составляли
малоурожайные поколения 1971, 1972, 1973 гг. (11,62-14,76 млн экз.). Уловы
красноперки в тот период колебались от 4,1 до 5,2 тыс. т при относительно
высокой интенсивности промысла. Количество секретов, используемых на промысле
туводных рыб, в те годы было самым высоким (90 тыс. шт.). В 1979-1982 гг.
добыча красноперки вновь возросла. При несколько меньшей интенсивности промысла
(в среднем 75,5 тыс. секретов) улов на усилие составил ПО кг/секрет, т.е. в 2.4
раза больше, чем в 1976- 1978 гг. В тот
период промыслом изымалось в среднем 36,9% от промыслового запаса, в 1976-1978
гг. эта величина составляла 22,9%. Несколько лет интенсивного промысла
отрицательно сказались на запасах красноперки. В 1983 г. наметилась тенденция к
снижению добычи. Значительную долю (61,5%) в уловах составляли рыбы старших
возрастов - остаток от поколений 1975, 1976, 1977 гг. рождения, которые по
убыли от лова показали себя как относительно урожайные (36,2-44,2 млн. экз.).
Названные годы были маловодными, с непродолжительным паводком, поэтому эффективность
нереста красноперки в полойной системе была невелика. Однако в авандельте,
наоборот, низкие уровни и малая проточность способствовали весьма эффективному
нересту и развитию красноперки. Благоприятные условия воспроизводства в
авандельте и обеспечили многочисленность этих поколений. Вступившее в промысел
поколение многоводного 1979 г. оказалось относительно малочисленным (17,3 млн
экз.). Несмотря на большой объем весеннего половодья, 1979 год был
неблагоприятным для размножения красноперки, так как в годы с большим, но
непродолжительным паводком часть самок не успевает повторно отнереститься,
ухудшаются условия выживания личинок, молодь не достигает покатных этапов и
погибает в личиночном периоде [Коблицкая, 1984]. Высокие уровни и большая
проточность, низкие температуры, слабое развитие кормовой базы создали и в
авандельте неблагоприятные условия для нереста и развития этого вида. Таким
образом, численность отдельных поколений определяется как величиной нерестового
стада, так и условиями воспроизводства (объемом и продолжительностью весеннего
половодья, температурой воды, субстратом и др.). В годы с низким и
непродолжительным паводком возрастает нерестовое значение авандельты. Однако
частая повторяемость маловодных лет может привести к общему ухудшению
экологической обстановки в авандельте, ее обмелению, заилению и заболачиванию,
что отрицательно скажется на формировании численности туводных рыб.
Красноперка является важным
компонентом питания хищников - сома и щуки. Значение в пищевом спектре этих рыб
колеблется от 40-50% (1970-1971 гг.) до 18-30% (1976-1977 гг.). Ежегодное
потребление красноперки сомом составляет в среднем 8,3-15,2 тыс. т, щукой -
3,7-5,3 тыс. т [Орлова, 1981].
По общепринятому мнению, красноперка довольно
вялая и ленивая рыба, по образу жизни занимающая как бы середину между плотвой
и карасем. Она постоянно держится на средних глубинах в траве, еще более
камышах, особенно в молодом возрасте, не любит сильного течения и в быстрых
реках даже вовсе не встречается. Пища ее, как и следует ожидать, состоит
преимущественно из растительных веществ, частью насекомых, личинок и червяков.
По свидетельству низовых рыбаков, красноперка весной кормится почти одним
шелкоником, нитчатой водорослью, которая плавает в затишьях и ильменях в виде
тонких ярко-зеленых нитей; от этого рыба будто бы принимает более зеленый цвет,
а мясо ее приобретает более горький вкус. Действительно, мясо красноперки летом
очень горькое, и весьма многие рыбаки объясняют эту горечь тоже ее
исключительно растительной пищей. Осенью красноперка обыкновенно уходит в
глубокие и тихие места и лежит там в продолжении всей зимы.
Как отмечал Л.Сабанеев: «По моим пятилетним
наблюдениям над красноперкой в подмосковных прудах - это, относительно говоря,
довольно сильная, бойкая и осторожная рыба; во всех этих отношениях она
превосходит плотву - свою постоянную спутницу, хотя местопребывание их
несколько различно. Именно красноперка встречается в наибольшем количестве в
самых густых зарослях трав, преимущественно в камышах, тростниках, между лопухами
(кувшинкой) и около водяного мха, так что в заросших прудах она всегда
многочисленнее плотвы. Подобно линю, она встречается, таким образом, в
наибольшем количестве в верховьях пруда и на открытые, незаросшие места почти
никогда не выходит; но она не любит, однако, тени и свежей воды и не
придерживается дна, как линь и плотва, а потому плавает все лето (с середины
мая до середины августа), б. ч. в полводы, а в солнечную погоду - в верхних ее
слоях, примерно на 15 см от поверхности; вообще она любит греться на солнце. В
жары даже плавает совсем поверху и ведет еще более дневной образ жизни, чем
щука и окунь: ее видно только с восходом и до заката солнца (кроме времени
нереста); вечером она уходит в нижние, более теплые слои воды и забивается в
траву, большей частью в т. н. водяной мох или в камыш и тростник. Самая крупная
красноперка держится в камышовых и тростниковых зарослях весь день и плавает
здесь на самом дне, выходя только на их окраины.
Красноперка никогда не уходит под плавучие
берега - любимое местопребывание линя в жары и холод, а также не прячется (в
жаркое время) под кусты и корни на берегу, подобно плотве, которая, по крайней
мере при совместном сожительстве с красноперкой, придерживается более открытых
мест и плавает большей частью близко от дна. Красноперка любит плавать около
купален, мостов, плотов, но только в том случае, если поблизости их растут
травы. Она положительно не отдаляется от зарослей и на несколько метров и не
любит даже и слабого течения, хотя охотно держится в более тихих местах
мельничного омута, привлекаемая сюда обилием пищи. В быстроте движений она
ничуть не уступает плотве, а кто видел, с какой силой она всплескивает или,
вернее, кувыркается, играя на поверхности, тот наверное скажет, что этот
всплеск производится более сильной рыбой, чем плотва.
Пища красноперки состоит, по моим наблюдениям,
из молодых побегов растений, нитчатых водорослей (Spirogyra, Cladophora -
шелковник, "зелень"), червей, насекомых, икры других рыб, а также
молоди. Летом красноперки весьма охотно, по-видимому, объедают икру улиток на
нижней стороне листьев кувшинок; нередко в зарослях кувшинок слышится
повсеместное чмокание - это красноперки счищают прилипшую к листьям слизистую
икру улиток. Подобный же звук часто издает в воздухе и пойманная красноперка. К
сентябрю, когда начнут пропадать травы, молодые красноперки переходят в камыши
и, вероятно, там же и зимуют. Взрослые рыбы в это время держатся более глубоких
мест, все реже и реже показываются у поверхности и, наконец, в октябре залегают
на зиму, как бы пропадают, и уже не берут на удочку. В прудах и озерах, при
зимнем сдыхании воды, красноперка всплывает на поверхность и ловится тогда в
большом количестве. Впрочем, это очень выносливая рыба, почти такая же
неприхотливая на качество воды, как линь, и много крепче и живучее плотвы».
Статус вида.
Широко распространенный, местами многочисленный, но малоценный промысловый вид
в большинстве районов ареала, особенно в дельтах крупных рек и в
водохранилищах. Является второстепенным объектом промысла в южных районах.
Глава III.
Годичные половые циклы и их особенности
экология нерест красноперка густера
Половое созревание у самок
густеры наступает в возрасте 3 лет, у самцов - 2 лет. В Заливе Кирова
(Азербайджан) можно встретить редкую карликовую форму, достигающую
половозрелости в возрасте года при длине тела самок 5,0 см, самцов - 4,5 см.
Плодовитость при размерах рыб
от 7,6 до 15,5 см в среднем составляет 10652 икринки с колебаниями от 1728 до
36850 икринок (табл. 1). В 1980 г. плодовитость достигала в среднем 21488
икринок (13750-34400). Размеры самок при этом составили 9,5-14,7 см, масса -
19-63 г.
Коэффициент зрелости самок
густеры в конце второй декады апреля в среднем составляет 15,35% с колебаниями
от 10,6 до 23,63%, самцов - 7,25% (4,66-10,0%).
Нерест происходит в конце
апреля при температуре воды 17-21°.
Средняя длина годовиков густеры
из Малого залива (май 1962) составляла 7,2 см с колебаниями
от 4,5 до I 10 см, масса
- 8,4г (3,3-30,5 г), упитанность (по Фультону) - 2,77 (2,71-3,5). Самки в возрасте
4 лет достигают I длины
в среднем 16,5 см, массы - 122 г, 5 лет - 18,6 см, 172 г, самцы -
соответственно 15,8 см и 112 г. 17,6 см и» 145 г.
Таблица 1 Плодовитость густеры
Длина тела рыб в неводном улове
колеблется от 13 до 20 см, в среднем 17,2 см, масса - от 57 до 366 г, в среднем
164 г. Самки по размеру и массе несколько превосходят самцов. Если средний
размер самок составляет 17,5 (14-20) см, а масса -- 182 (70-366) г, то самцов
-- 16 (13-20) см 1 135 (57-220) г. Длина тела исследованных в апреле 1980 г. 32
экз. густеры колебалась от 4,3 до 14,7 см, в среднем 8,1 см, масса - от 1,95 до
63, г. в среднем 15,5 г.
Индивидуальные колебания
коэффициента упитанности, по Фультону - от 2,20 до 3,04, в среднем 2,67; по Кларк-
от 1,98 до 2,74, в среднем 2,33. Модель по упитанности не уступает взрослым
рыбам, иногда даже превосходит их.
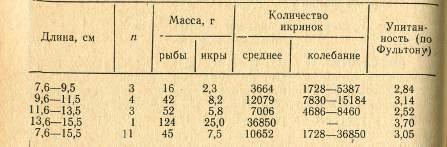
Таблица 2 Изменение упитанности
и массы густеры в Малом заливе в зависимости от размера рыб
Масса рыб обоих полов с
увеличением длины пропорционально увеличивается, при этом различия между полами
до определенного размера (19 см) не так заметны, затем прирост массы у самок
заметно увеличивается; по упитанности существенных различий между самками и
самцами одного размера не наблюдается (табл. 2).
В улове встречаются рыбы в
возрасте от 1 до 5 лет.
У красноперки в связи с
тем, что по меристическим и пластическим признакам различий между самцами и
самками почти нет, индексы их морфологических признаков определялись в
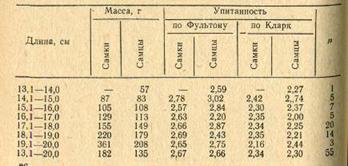
объединенных пробах. Результаты приведены в табл. 3.
Таблица 3 Морфологические
признаки красноперки
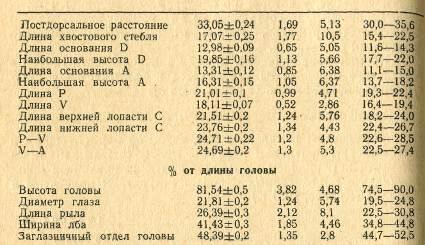
Половозрелость заливе наступает
в возрасте 2-3 лет, самцы с созревшими половыми продуктами часто встречаются в
возрасте одного года. Икрометание происходит в мае в зарослях травы при
температуре воды 16-19,5°.
В Каспии южнее Кизлярского
залива представлена некрупными экземплярами: индивидуальное колебание длины
тела - 8-18 см, массы - 14-1902 г. В возрасте одного года длина составляет 10,8
см, масса - 30,7 г (л=21), S лет
- 14,3 см и 82 г (л = 6), 3 лет - 16,3 см и 119 г Как видно, в первые 2 года
красноперка растет интенсивно, в дальнейшем темп роста, особенно линейного,
замедляется, что связано с наступлением половозрелости.
Биологические показатели
красноперки, исследованной в различные годы из несколько различаются (табл. 4).
В частности, в Азербайджане, где впервые были проведены исследования
биологических показателей, в придаточных водоемах рыбы оказываются более
крупными: исследованные в мае 1961 г. из оз. Нахалыхчала имели длину тела 15-25
см, массу 90-570 г.
Стадо красноперки представлено
3 возрастными группами - 2-4 лет, преобладают трехлетки.
Таблица 4 Биологические показатели
красноперки
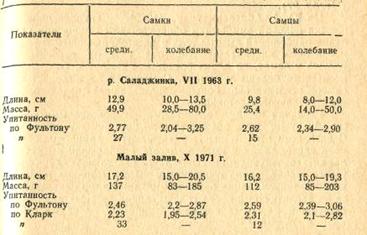
Размножение, как известно,
включает следующие процессы: развитие и созревание половых продуктов
(гаметогенез), наступление половой зрелости, нерест (икрометание),
оплодотворение икры и развитие эмбрионов (эмбриогенез), рост и развитие личинок
и молоди, скат производителей и молоди рыб, их пищевая миграция и др.
Экология нереста. После
проведения эколого-морфофизиологических исследований размножения рыб в водоемах
Дагестана все изученные виды рыб могут быть сгруппированы по многим
биологическим признакам. Так, по характеру нерестового субстрата все изученные
виды рыб, по классификации С. Г. Крыжановского, отнесли к двум экологическим
группам: фитофилы и литофилы. К первой группе отнесли рыб, откладывающих икру
на растительный субстрат, - сазан, лещ, вобла, красноперка, линь, кутум,
сом, щука, окунь и судак. Литофилы включают в себя рыб, размножающихся на
песчано-каменистых местах (на плотном грунте), обычно на течении или в местах с
высокой проточностью и с очень чистой водой. Следует заметить, что в водоемах
Дагестана после зарегулирования их стока, у некоторых видов как фитофильной,
так и литофильной группы (рыбец, жерех) стала наблюдаться индифферентность к
субстрату и глубине нереста, что, по-видимому, связано с высокими адаптивными
свойствами, характерными для этих видов к меняющимся в сторону ухудшения
экологическим условиям.
По периодам (сезонам)
размножения рыбы делятся на две группы: с весенне-летним и осенне-зимним
икрометанием. Исследованные нами виды относятся к первой группе.
Экологические наблюдения за
размножением исследованных рыб показали, что икрометание их происходит при
наличии определенных условий (температура, уровень воды, субстрат, проточность
и т. д.), которые носят видоспецифический характер. По требованиям к
экологическим условиям, необходимым для нереста, всех исследованных рыб мы
разделили на три группы: к первой отнесли рыб, нерест у которых ранний (в
марте), кратковременный (10--15 дней) и происходит при низкой температуре (5-10°С);
вторая группа - нерест поздний (май-июнь), растянутый (40-80 дней) и происходит
при более высоких температурах (20-25°С); третья группа занимает по всем этим
показателям-промежуточное положение между I
и
II группами
(апрель-май, 20-40 дней, 15-20°).
Нижний порог нерестовой
температуры (минимальная температура начала нереста) наиболее высокий у
красноперки (17-18°С). Соответственно и верхний порог нерестовой температуры
(максимальная температура нереста) у первой группы 20- 25°С, второй - 10-15°С, а
в третьей - 16-20°С.
По степени зависимости нереста
от паводкового режима (по классификации А. Ф. Коблицкой) всех исследованных (12
видов) рыб можно разделить на три группы: I-нерест
не зависит от хода паводка; II-успешность
нереста зависит от времени начала и продолжительности паводка; III
-
нерест происходит позднее, когда нерестилища уже залиты и меньше зависят от
хода паводка, - густера и красноперка входят в эту группу
По реакции к прочим факторам
водной среды (содержание кислорода, проточность воды, наличие субстрата и т.
д.) изученные виды рыб мы отнесли также к трем группам. Первая - те рыбы,
которые требовательны к содержанию кислорода, проточности вод и т. д. Для этих
видов рыб, вследствие интенсивной зарастаемости водоемов низовьев Терека, Сулака"
и Аграханского залива в весенний период, когда происходит икрометание, факторы,
обусловливающие их эффективный нерест, чаще оказываются значительно ниже
оптимальных;, поэтому численность их значительно сократилась и в промысловых
уловах их бывает очень мало. Вторая группа - линь, окунь, красноперка и
многие сорные виды рыб (густера, уклейка, карась), не требовательные к
содержанию кислорода, проточности воды и прочим факторам, водной среды,
прогрессируют в водоемах Дагестана и в ихтиофауне водоемов представляют самую
многочисленную группу. Третья группа занимает по требовательности к этим
условиям промежуточное положение. Численность этой группы в ихтиофауне водоемов
относительно ко второй сократилась.
Типы икрометания рыб, как
известно, определяются прежде всего условиями размножения. В литературе
существуют различные мнения по этому вопросу. В частности, указано, что у одних
видов рыб в процессе созревания половых продуктов в течение годичного полового
цикла развивается и выметывается одна порция икры, т. е. имеет место
единовременный тип икрометания. У других - асинхронное развитие и
неодновременное выметывание половых продуктов в течение нерестового периода, т.
е. порционный тип икрометания. А у некоторых видов - так называемые переходные
формы - в одних условиях наблюдается единовременный тип икрометания с
синхронным ростом овоцитов, в других же условиях (у этих самых видов) -
порционный тип икрометания с асинхронным ростом овоцитов. У рыб же с порционным
типом икрометания (в том числе густера и красноперка) процесс размножения более
сложный.
Икрометание у красноперки
порционное и происходит в два или три приема (Чикова, 1966; Пенязь, 1973 и др.)
с промежутками в две недели.В то же время известно (Кузнецов и др., 1974), что
у красноперки в условиях водохранилища нерест единовременный, а в озерах -
порционный. Однако авторы не дают подробных объяснений причин этого явления и
неясно, чем оно вызвано у этого вида.
По типу икрометания и характеру
развития половых продуктов все изученные нами виды рыб мы отнесли к двум
группам: с единовременным типом икрометания и синхронным развитием половых
продуктов; с порционным икрометанием и асинхронным ростом овоцитов (красноперка
и сорные рыбы).
Порционность икрометания - одно
из важнейших свойств вида для обеспечения его высокой численности - имеет ряд
преимуществ: выметывание икринок порциями в течение длительного периода в
разных условиях среды создает большую вероятность выживания икры и личщкж. При
гибели помета (поколения) одной порции потомство особи имеет возможность
сохраниться за счет развития других порций. Кроме того, порционность удлиняет и
период нереста, что позволяет молоди равномерно и более полнее осваивать корма.
Изменение условий в водоемах Дагестана после зарегулирования их стока
благоприятствовало размножению рыб с порционным нерестом, т. е. с более
растянутым нерестовым периодом. В условиях искусственного рыборазведения
наиболее выгодно, на наш взгляд, использовать в качестве объектов разведения
порционно нерестующие виды рыб.
Изменения экологических условий
(температура, субстрат, уровень воды и др.) в связи с зарегулированием стока
отрицательно сказались не только на сроках нереста, продолжительности
нерестового периода, но также и на изменении мест (субстрат) и ареала
(сократились площади нерестелищ) нереста этих рыб.
Каждый из рассмотренных видов
рыб приспособлен к нересту в определенных условиях и имеет свой специфический
характер
икрометания, но при отсутствии
необходимых условий, как уже было отмечено выше, некоторые виды рыб (рыбец и
кутум) иногда нерестуют и на несвойственных им субстрате и глубинах.
Таким образом, у некоторых
видов рыб в измененных условиях выработались различные адаптивные свойства,
способствующие главным образом сохранению вида.
Знание плодовитости (количества
выметываемой икры) необходимо для решения ряда практических задач: установления
необходимого количества производителей с учетом обеспеченности хозяйства
рыбопосадочным материалом, определения эффективности естественного нереста,
расового изучения рыб и т. д.
Различают индивидуальную
(абсолютную), относительную, рабочую, популяционную и видовую плодовитость. При
анализе мы пользовались лишь двумя показателями: абсолютной или индивидуальной
плодовитостью (общее количество икринок, выметываемое самкой за один нерестовый
период) и относительной (количество икры, приходящееся на единицу веса самки).
Количество икры, продуцируемое разными видами рыб в водоемах Дагестана, сильно
варьирует.
Сравнительный анализ
плодовитости одних и тех же видов одноразмерных рыб, но обитающих в различных
водоемах, показал более высокую воспроизводительную способность у рыб в
водоемах южных широт. У тех видов рыб (окунь, линь, красноперка и др.), для
которых, изменившиеся экологические условия оказались более или менее
благоприятными (в отличие от некоторых других видов рыб) для роста,
плодовитость не снизилась. Доказательством этому в какой-то мере может служить
увеличение численности этих видов рыб в водоемах, особенно после
зарегулирования их стока.
Снижение плодовитости у
порционно нерестующих видов рыб, видимо, произошло за счет сокращения
количества мелких икринок, формирующих последующие порции икры, вследствие чего
показатель порционности также значительно уменьшился.
Исследования по изучению
половых циклов и гаметогенеза рыб в водоемах Северо-Западного Каспия (Дагестан)
до их зарегулирования не проводились и поэтому дать сравнительную
характеристику особенностей этих процессов в различных условиях (до и после
происшедших изменений) не представляется возможным.
В ходе работы приходилось
изучать внешнюю (макроскопическую) и внутреннюю (микроскопическую) картины
половых желез (самок и самцов) рыб в течение годичного полового цикла.
Половые органы рыб и самок
представлены яичниками, а у самцов - семенниками. Яичники и семенники у изученных
видов рыб парные и обычно имеют вытянутую форму (кроме окуня и сома, семенники
у которых имеют овальную форму), расположены в дорзальной полости тела. Вдоль
яичника проходит кровеносный сосуд с многочисленными ответвлениями, которые
наиболее ярко бывают выражены на II, III, IV и
V стадиях их
зрелости. В семенниках же такой кровеносный сосуд отсутствует, что служит
хорошим признаком для различия пола. Половые железы снаружи покрыты плотной
оболочкой, состоящей из нескольких слоев.
Микроскопическое (гистологическое)
изучение яичников и семенников позволило выявить ряд особенностей в развитии
половых клеток в течение годичного полового цикла. Гистологическая картина
каждой стадии зрелости у самок и самцов характеризуется наличием определенного
комплекса половых клеток, находящихся в разных фазах их роста и развития.
Переход половых желез от одной стадии к другой (от низшей к высшей)
определяется переходом половых клеток от одной фазы их развития к другой. Так,
для первой стадии зрелости яичников для всех видов рыб характерны образования
овоцитов ювенальной фазы развития и наличие первичных половых клеток -
овогоний. У самцов на этой стадии половые образования представлены первичными
половыми клетками - сперматогониями.
В яичниках второй стадии
зрелости основная масса овоцитов находится в фазе однослойного фолликула,
отличающегося от овогоний большим размером и характерной для премейотических
фаз зрелости- структурой ядра. Параллельно с процессом созревания овоцитов и
переходом в последующие фазы происходит заметное уменьшение количества
первичных половых клеток - овогоний. В семенниках второй стадии преобладают
сперматоциты 1 порядка и первичные сперматогонии.
Вторая стадия у рыб с
единовременным нерестом (вобла, окунь, щука и т. д.) наступает при завершении последующего
полового цикла, т. е. у самок - после каждого икрометания и окончания
резорбционных процессов, а у самцов - после выбоя и резорбции остаточных
спермиев. У некоторых видов порционно нерестующих рыб (сазан, карп) вторая
стадия зрелости яичников наблюдается только у неполовозрелых самок, а у
половозрелых она полностью выпадает.
Интересно заметить, что при
гистологических исследованиях гонад внутренние структуры яичника на второй
стадии у неполовозрелых самок и самок после икрометания, т. е. очередного
полового цикла резко различаются: у повторно созревающих самок на срезе яичника
хорошо заметны, кроме овоцитов в фазе протоплазматического роста, ещё и следы
или остатки прошедшего нереста в виде включений или скоплений пигментных
клеток, благодаря чему у них выше и коэффициент зрелости.
Для третьей стадии характерно
начало вителлогенеза, т. е. большинство овоцитов находятся в фазе
первоначального накопления желтка (фаза вакуолизации); кроме того, имеется и
комплекс овоцитов, характерных для предыдущих II
и
I стадий. У самцов на
III стадии половые
клетки в семенниках представлены главным образом сперматоцитами второго
порядка, но имеются сперматоциты первого порядка и единичные сперматогонии,
характерные для младших стадий зрелости.
При IV
стадии
зрелости яичников большая часть овоцитов находится в фазе наполненного желтком,
в остальной массе-овоциты всех фаз развития, обнаруживаемые в предыдущих
стадиях. Однако эта стадия имеет ряд особенностей, свойственных как для
единовременно, так и порционно нерестующих рыб. Так, у единовременно
нерестующих в яичниках на этой стадии имеются овоциты в фазе наполненного
желтком и комплекс овоцитов, присущих для II
и
I стадий зрелости, но
нет овоцитов, характерных для III стадии
зрелости (в фазе первоначального накопления желтка или вакуолизации), тогда как
у порционно нерестующих рыб в яичниках имеются овоциты всех фаз развития,
характерные для предыдущих III, II и
I стадий зрелости, т.
е. овоциты во всех фазах трофоплазматического и протоплазматического роста. У
последних в течение всего годичного полового цикла наблюдается асинхронность в
росте и развитии, которая стойко сохраняется в дальнейшем.
Гистологическая картина пятой
стадии по некоторым внешним признакам сходна с IV,
однако
появляются некоторые признаки, свойственные только для яичников пятой -
предовуляционной стадии, а именно: овоциты достигают дефинитивных размеров
(зрелое состояние), в них появляются «микропиле» с замыкающей клеткой,
являющейся местом проникновения сперматозоидов для оплодотворения. В семенниках
семенные ампулы приобретают вытянутую или округлую формы, заполненные
сперматозоидами, готовыми при наступлении оптимальных условий к размножению и
сразу же к выделению.
Гистологическая картина
яичников и семенников в посленерестовой VI
стадии
представляет интерес. У единовременно нерестующих рыб после икрометания
(овуляции зрелых овоцитов) в яичниках остаются лишь единичные, невыметанные по
разным причинам, икринки, многочисленные фолликулярные оболочки, которые
подвержены процессу резорбции, а также имеются и овоциты протоплазматического
роста. Вителлогенез начинается у них только по завершении резорбционных
процессов или даже при .наличии конечных фаз резорбции в виде включений
пигментных клеток.
У порционно нерестующих видов
рыб вителлогенез (образование желтка) в нерестовый и посленерестовый периоды не
прекращается. Поэтому в яичниках в течение всего года наблюдаются наряду с
овоцитами протоплазматического роста и овоциты на разных фазах
трофоплазматического роста. Однако у некоторых видов и с порционным нерестом
(линь, красноперка) в яичниках после выметывания всех порций икры овоциты
трофоплазматического роста не обнаруживаются или присутствуют лишь единичные,
причем в начальной фазе вакуолизации (с одним рядом вакуолей).
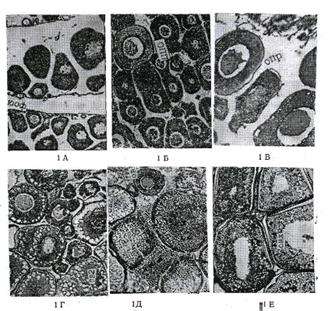
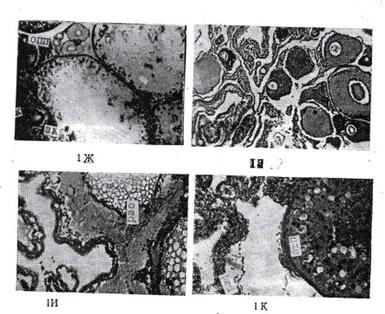
Рис. 6. Микрофотография
яичников на всех стадиях зрелости: А-I стадия,
Б-II стадия у
неполовозрелых самок. В - II стадия
после нереста. Г-III
стадия,
Д-IV стадия у порционно
нерестующих рыб, Е-IV стадия
у единовременно нерестующих рыб, Ж-овоциты дефинитивного размера (V-стадия),
видно микропиле с замыкающей клеткой 3-VI
стадия
у единовременно нерестующих рыб, И-резорбирующихся фолликул и остаточных
икринок, К-VI стадия у
порционно нерестующих рыб
У самцов исследованных рыб
после нереста семенники переходят в состояние полного выбоя (VI
стадия);
в некоторых семенных ампулах иногда остаются невыделенные сперматозоиды, но
они, очевидно, постепенно подвергаются процессу резорбции. Одновременно с
резорбцией остаточных спермиев на стенках семенных ампул (канальцев) появляются
первичные половые клетки - сперматогонии (крупного размера), которые обычно
расположены в них в один ряд, в виде цепочки.
Установлены также для каждого
вида и пола сроки и продолжительность прохождения отдельных стадий зрелости
гонад (яичников и семенников) в течение годичного полового цикла в конкретных
условиях водоемов. Все обнаруженные особенности в прохождении годичного
полового цикла у изученных видов рыб (самок и самцов) отражены в разработанной
нами схеме (шкале) применительно для водоемов южных широт. Из схемы видно, что
время прохождения и длительность отдельных стадий зрелости гонад самок и самцов
в течение годичного полового цикла у разных видов рыб, независимо от их
близости в систематическом и биологическом отношениях, заметно различаются. По
данной схеме можно легко определить сроки начала и продолжительность каждой
стадии зрелости как яичников, так и семенников указанных в ней различных видов
рыб, поэтому она имеет не только теоретическое значение, но может быть
использована и в практических целях, в частности, при искусственном
рыборазведения.
По отличительным свойствам ово-
и сперматогенеза, срокам прохождения половых циклов и длительности отдельных
стадий зрелости все изученные виды рыб мы отнесли к двум различным группам.
Так, самки основных промысловых рыб входят в группу с ранневесенним кратким
нерестом. Более разнообразно протекает сезонная циклика у порционно нерестующих
видов. У густеры, линя и красноперки после выметывания последней порции
(выметывают при наличии оптимальных условий для нереста не менее двух порций
икры) яичники их переходят в посленерестовую VI-II
стадию.
По завершении резорбционных процессов яичники переходят во II-III
стадии,
точнее даже во II стадию
зрелости, которая длится недолго - менее одного месяца. В то же время у этих
двух видов рыб более продолжительное время (6-7 месяцев) половые железы
находятся в переходной II -
III и III
стадиях
(сентябрь-март) зрелости, зимуют они с яичниками в III
стадии
зрелости, т. е. с незавершенным вителлогенезом.
Заключение
Результаты исследований по
экологии размножения изученных видов рыб позволяют не только составить шкалу их
нереста применительно для водоемов южных широт, но и выявить некоторую
несогласованность в сроках запрета лова рыб в водоемах Дагестана, установленных
с 15 апреля по 15 июня. Во-первых, наиболее интенсивный период нереста у
высокоплодовитых особей (модальные группы) многих ценных видов рыб (кутум,
жерех, вобла, лещ, судак и т. д.) приходится именно на апрель. Следовательно,
при существующих сроках запрета эти особи изымаются из водоемов до получения от
них наиболее высокопродуктивного потомства. Во-вторых, интенсивный нерест
многих сорных и малоценных видов рыб (потенциальные пищевые конкуренты ценных
видов рыб и их молоди) начинается с первых чисел июня. Таким образом, за 15
дней июня они успевают отнереститься. Исходя из этого, мы предлагаем внести
корректив в сроки запрета лова рыб в водоемах данного региона, установив их с 1
апреля по 1 июня. При этом продолжительность запрета остается прежней.
Из анализа обширного
литературного материала видно, что сроки начала и продолжительности
икрометания, а также температурные условия нереста у одних и тех же видов рыб,
но обитающих в разных водоемах, различны. Эти показатели у рыб, обитающих в
водоемах южных широт, резко отличаются от обитающих в средних и тем более
северных широтах, что также необходимо учитывать при установлении сроков лова
рыб.
Анализ возрастного и размерного состава
нерестовых стад позволил выявить ряд особенностей, специфичных для каждого из
всех исследованных видов рыб. Сопоставление литературных и наших данных
показывает, что половая зрелость у одних и тех же видов рыб в водоемах южных
широт наступает значительно раньше (в раннем возрасте) и при относительно
меньших размерах производителей, чем в водоемах средних и северных широт,
по-видимому, это связано с экологическими и прежде всего температурными
условиями. По-видимому, изменение плодовитости у одних видов рыб в сторону увеличения,
а у других-уменьшения соответственно их численности в указанных водоемах
вызвано ухудшением условий для размножения. Условия размножения для большинства
изученных видов рыб в водоемах Дагестана после зарегулирования их стока резко
ухудшились в связи с недостатком нерестилищ (сужение их ареала), резкими
колебаниями уровня воды, особенно в период нереста, когда он совпадает с
периодом максимального водозабора для орошения, неравномерным прогревом масс
воды и т. д. В результате этого большая часть рыб с единовременным нерестом
остается яловой, с порционным нерестом (линь, красноперка, сазан - карп) не
выметывает третью и большую часть - вторую, а некоторые особи даже и первую
порцию икры.
Таким образом, по особенностям
ово- и сперматогенеза, срокам прохождения и продолжительности отдельных стадий
зрелости можно утверждать, что самки густеры, линя, рыбца и красноперки, у
которых вителлогенез интенсивно протекает весной незадолго до начала или
непосредственно перед нерестом, а зимуют они с яичниками в III
стадии
зрелости, т. е. с незаконченным вителлогенезом. По отличительным свойствам
сперматогенеза самцов названных рыб сперматогенез усиленно протекает в весенний
период (перед началом нереста), а зимуют они с семенниками в переходной II-III
или
III стадиях зрелости.
В результате исследований
выявлены некоторые специфические особенности в биологии размножения отдельных
видов рыб в водоемах данного региона. При этом наблюдаются как общие
биологические закономерности, свойственные для рыб, обитающих в разных экологических
условиях, так и отклонения от нормы некоторых биологических показателей,
проявляемые в измененных условиях водоемов в связи с зарегулированием их стока.
Список использованной литературы
1. Водный
кодекс Российской Федерации от 16 ноября 1995 г. N 167-ФЗ (с изм. и доп. от 30
декабря 2001 г., 24 декабря 2002 г., 30 июня, 23 декабря 2003 г.). Федеральный
закон от 31 июля 1998 г. N 155-ФЗ "О внутренних морских водах,
территориальном море и прилежащей зоне Российской Федерации" (с изм. и
доп. от 22 апреля, 30 июня, 11 ноября 2003 г.).
. Федеральный
закон от 8 декабря 1995 г. N 193-ФЗ "О сельскохозяйственной
кооперации" (с изм. и доп. от 7 марта 1997 г., 18 февраля 1999 г., 21
марта 2002 г., 10 января, 11 июня 2003 г.).
. Федеральный
закон от 30 ноября 1995 г. N 187-ФЗ "О континентальном шельфе Российской
Федерации" (с изм. и доп. от 10 февраля 1999 г., 8 августа 2001 г., 22
апреля, 30 июня, 11 ноября 2003 г.)
. Федеральный
закон от 24 апреля 1995 г. N 52-ФЗ "О животном мире" (с изм. и доп.
от 11 ноября 2003 г.).
. Указ
Президента РФ от 29 августа 1997 г. N 950 "О мерах по обеспечению охраны
морских биологических ресурсов и государственного контроля в этой сфере"
. Указ
Президента РФ от 8 декабря 1992 г. N 1551 "О мерах по стабилизации
финансового положения предприятий и организаций рыбного хозяйства"
. Постановление
Правительства РФ от 9 декабря 1998 г. N 1457 "О выплате в 1998 году
выходного пособия и компенсаций работникам органов рыбоохраны и воспроизводства
рыбных запасов Министерства сельского хозяйства и продовольствия Российской
Федерации"
. Абдурахманов
Ю.Ф. Рыбы залива Кирова. Баку: 1988 - 328 с.
. Аннотированный
каталог круглоротых и рыб континентальных вод России // Под ред.
Ю.С.Решетникова. М.: Наука, 1998. 218 с.
. Атлас
пресноводных рыб России: В 2 т. Т.1. // Под ред. Ю.С.Решетникова. М.: Наука,
2004 - 379 с.
. Багров
AM., Виноградов В.К.,
Магомаев Ф.М. Проекты ученых должны осуществиться // Рыбоводство и
рыболовство.-2001. - № 1. - С. 60-61.
. Бекин
АГ., Виноградов В.К., Магомаев Ф.М., Орлов Ю.И. Технология непрерывного
выращивания: Обзор, информ. - М.: ЦНИИТЕЭРХ, 1999. - Вып. 3. - 72 с.
. Берг
Л.С. Рыбы пресных вод СССР и сопредельных стран. М.; Л.: Изд-во АН СССР. 1949
Т. 2. С. 469-929.
. Бердичевский
Л.С. Биологическое обоснование рационального использования запасов промысловых
рыб Каспийского бассейна. Проблемы Каспийского моря. - Баку: Изд. АН Азерб.
ССР, 1963. - С. 27-35.
. Богерук
А.К., Волчков Ю.А., Илясов Ю.И., Касатонов В.Я. Концепция селекционных
достижений в аквакультуре: информ. пакет. - М.: ВНИЭРХ, 1997. - С. 1-43.
. Васильев
В.П. Эволюционная кариология рыб. М.: Наука, 1985. 300 с.
. Вилер
А. Определитель рыб морских и пресных вод М.: Инфра-М, 2003 - 457 с.
. Виноградов
В.К. Концепция развития пресноводной аквакультуры России // Рыбное хозяйство. -
2003. - № 5. - С. 17-18.
. Виноградов
В.К., Ерохина Л.В. Повышение эффективности товарного рыбоводства // Рыбоводство
и рыболовство - 1999.- № 3. - С. 6.
. Иванов
А.П. Рыбоводство в естественных водоемах. - М.: Агропромиздат, 1988. - 368 с.
. Казначеев
Е.Н. Рыбы Каспийского моря. М.:Пищепром, 1981 - 167 с.
. Кулиев
З.М. Рыбы Каспийского моря. Баку: 1988 - 224 с.
23. Магомаев Ф.М., Бер С.Б., Шайхулисламов
АО., Гимбатов И.М. Эффективное рыбоводство. - Махачкала: Изд. ДГУ, 2001. - 70
с.
. Магомаев Ф.М., Гимбатов Г.М.,
Шайхулисламов А.О., Гаджимусаев Н.М. Осетроводство в Дагестане. - Махачкала:
Изд. Юпитер, 2001. - 40 с.
. Магомаев Ф.М., Кривцов В.К., Богерук
А.К. Рекомендации по разведению и выращиванию растительноядных рыб в рыбхозах
Дат. АССР. - М.: ВНИИПРХ, 1975. - 38 с.
. Магомаев Ф.М., Магомедов ГА, Бекин АГ.,
Бер С.Б., Ахмаев Э.А Рекомендации по подращиванию личинок карпа и
растительноядных рыб в условиях Дагестана. - М.: ВНИИПРХ, 2003. - 22 с.
. Магомаев Ф.М., Магомедов М.А Перспективы
использования горных водохранилищ Дагестана в рыбохозяйственных целях //
Материалы междун. конференции - М.: Изд-во ВНИРО, 2002. - С. 98-102.
. Магомаев Ф.М., Столяров И.А
Рыбохозяйственное освоение внутренних водоемов Дагестана: Обзорная информация.
- М:. ВНИЭРХ, 2003. - 62 с.
29. Решетников
Ю.С., Богуцкая Н.Г., Васильева Е.Д. и др. Список рыбообразных и рыб пресных вод
России //Вопросы ихтиологии. 1997.Т. 37, вып. 6. С. 723-771.
. Рыбоводство
/ авт.-сост. А.И. Исаев М.: Агропром, 1996 - 432 с.
. Сорвачев
К. Ф. Основы биохимии питания рыб М.: Лёгкая и пищевая промышленность, 1988 -
247 с.
. Справочник
по рыбоохране М.: Агропромиздат, 1986 - 288 с.
. Цепкин
Е.А. Изменения промысловой фауны рыб континентальных водоемов Восточной Европы
и Северной Азии в четвертичном периоде //Вопр. ихтиологии. 1995. Т. 35, вып. 1.
С. 3-17.
. Шихшабеков
М. М. Рыбы Каспийского моря М.: Дагучпедгиз, 1980 - 96 с.