Особенности показателей вегетативной регуляции и функционального состояния у лиц с разным уровнем двигательной активности
Содержание
Ведение
Глава
1. Вегетативная регуляция функционального состояния организма
.1
Соотношения между парасимпатической и симпатической регуляциями сердца
.2
Парасимпатическая регуляция
.3
Симпатическая регуляция
.4
Гуморальная регуляция
Глава
2. Особенности показателей вегетативной регуляции и функционального состояния у
лиц с разным уровнем двигательной активности
.1
Основные физиологические свойства сердечной мышцы
.2
Ритм сердца. Показатели сердечной деятельности
.3
Оценка функционального состояния организма с учетом уровня их двигательной
активности. Нарушение сердечного ритма
.4
Экспериментальное исследование
Заключение
Список
литературы
Введение
Актуальность темы исследования обусловлена
интенсификацией процесса обучения в институте и ростом требований,
предъявляемых студенту при поступлении в ВУЗ, современной системой образования.
Известно, что начальный период обучения в институте является очень
ответственным как в социальном, так и в физическом отношении моментом в жизни
студента. Новые условия обучения, высокая суммарная учебная нагрузка, большой
объём, новизна и сложность материала, которым должен овладеть студент,
предъявляют к организму повышенные требования.
Существенная система обучения изначально
подавляет физиологические возможности организма, воздействуя на него в
прессинговом режиме однонаправленного характера, не учитывающем
морфофункциональных особенностей и индивидуальных темпов биологического
созревания. Это приводит к напряжению и срыву механизмов адаптации, существенно
снижает уровень здоровья интеллектуальный и физический потенциал студентов.
Ведущую роль в обеспечении адаптационной
деятельности организма играет система кровообращения. Эта роль определяется,
прежде всего, её функций транспорта питательных веществ и кислорода - основных
источников энергии для клеток и тканей. Таким образом, изменение параметров
сердечнососудистой системы может расширять или же наоборот, лимитировать
адаптационные возможности организма, так как энергетический механизм занимает
главное место в процессах адаптации.
Потенциальным подходом к анализу
приспособительных возможностей организма является изучение его функционального
состояния. Согласно концепции В.В.Парина и Р.М.Баевского (1967) особенности
адаптивных реакций можно оценить с помощью анализа механизмов регуляции
кардиоритма, потому что его специфика дает возможность получения
прогностической информации о функциональном состоянии всего организма.
В основе любого адаптивного цикла лежит
эволюционно обусловленная программа приспособления, реализующаяся в том или
иной форме двигательной активности, которая является одним из ведущих
гомеостатических факторов (Лобзин В.С., 1979). Вместе с тем все параметры
двигательной активности, обеспечивающие наилучший эффект строго индивидуальны
(Пирогова В.А., 1989). Нахождение оптимума возможно с помощью оценки
интегративных характеристик функционального состояния организма.
Согласно Р.М.Баевскому (1979), иерархическая
структура управления ритмом сердца включает последовательные уровни гуморальной
гормональной, вегетативной и центральной (корковой) регуляции. При оптимальном
управлении участие высших уровней минимально.
Цель:
использование показателей сердечного ритма для оценки функционального
состояния студентов.
Объект исследования: функциональное
состояние студентов.
Предмет исследования: показатели
сердечного ритма, влияющие на функциональное состояние студентов.
Выдвинутые цель, объект и предмет исследования
обусловили решение следующих задач:
выявить особенности показателей сердечного
ритма, влияющие на функциональное состояние студентов;
изучить зависимость показателей сердечного ритма
от функционального состояния учащихся;
Гипотеза: мы
предполагаем, что вегетативная регуляция в симпатикотоническим типом приводит к
напряжению механизмов адаптации сердечнососудистой системы к учебным нагрузкам.
Методы исследования: Методом
вариационной пульсометрии по Р.М.Баевскому обследованы 25 студентов факультета
музыкально-педагогического 1 курса (девушки) и 25 студентов 1 курса девушки
факультета физической культуры. В работе использован автоматизированный способ
регистрации кардиоинтервалов (бралось не менее 100 кардиоинтервалов) с помощью
электронного кардиографа посредством связи блютус (Поли-Спектр-8/EХ
фирмы Нейрософт) и кардиотренажер (KETTLER).
Алгоритм математической обработки массива
включает в себя вычисление моды (Мо), амплитуды моды (АМо), индекс
вегетативного равновесия (ИВР), вегетативный показатель ритма (ВПР), показатель
адекватности процессов регуляции (ПАПР), индекс напряжения регуляторных систем
(ИНрс). Методами математической статистики определяем среднее арифметическое
(М), среднее квадратическое отклонение (r),
ошибку выборки (m). Проверку
гипотезы достоверности различий параметров ритма сердца осуществляем по t
- критерии Стьюдента.
Глава 1. Вегетативная регуляция
функционального состояния организма
1.1 Соотношения
между парасимпатической и симпатической регуляциями сердца
Взаимодействие
парасимпатических и симпатических регуляторных аппаратов сердца не подвергалось
систематическому углубленному экспериментальному изучению. Существующее
представление рисует это взаимодействие в виде динамического балансирования
антагонистических влияний на основе реципрокных отношений между
соответствующими центральными аппаратами (Гельгорн, Луфборроу, 1966). Это
спекулятивное в своей основе представление не находит экспериментального
подтверждения в современных исследованиях.
Поэтому проблема взаимодействия
между парасимпатической и симпатической иннервацией при регуляторных влияниях
на сердце в настоящее время может обсуждаться лишь в предположительном плане.
Основной предпосылкой к обсуждению проблемы естественно должны служить наиболее
экспериментально обоснованные представления о регуляторных возможностях того и
другого иннервационного аппарата, выявленные при изучении раздельного их
функционирования.
Можно предполагать поэтому, что
симпатический регуляторный аппарат играет вспомогательную роль, по крайней
мере, при адаптивном управлении частотой сердцебиений, и приобретает значение
самостоятельного регулятора лишь в особых условиях и. по отношению к активации
сократительного миокарда.
Решающее значение
парасимпатической иннервации в регуляторной адаптации гемодинамической
производительности сердца было установлено в хронических экспериментах на
животных с предварительно устраненной симпато-адреналовой системой. Поэтому
может оказаться, что парасимпатическая только в отсутствии симпатической
иннервации сердца приобретает способность дублировать функции последней.
Однако, система данных показывает, что парасимпатический аппарат сердца в
условиях сильных мышечных напряжений и при стартовых состояниях оказывает
ускорительные влияния, которые по своей мощности значительно превосходят
ускорения сердцебиений, обусловленные электрической стимуляцией симпатических
путей сердца, введением максимально эффективных доз катехоламинов или
интенсивным рефлекторным возбуждением симпатической иннервации сердца,
вызванным ваготомией или повышением внутричерепного давления.
Приведенные выше данные
показали, что количественные модификации парасимпатических импульсных влияний
элементов обусловливают переход от тормозных эффектов к ускорительным в
пределах одного и того же опыта, как в условиях предварительно удаленной, так и
при интактной симпатической иннервации. Эти экспериментальные данные составляют
убедительное доказательство того, что как тормозящие, так и ускорительные
влияния парасимпатической иннервации сердца определяются исходными свойствами
ее структурно-функциональной организации и зависят только от количественных
особенностей включения в активность элементов. Последнее вместе с высокой
эффективностью парасимпатических ускорительных влияний, явно превосходящей
потенциальные возможности симпатических ускорений, показывает, что в обычных
условиях физиологического напряжения организма парасимпатическая иннервация
составляет основной нервный аппарат, с помощью которого осуществляется не
только торможение, но и предельные степени ускорения сердца. О регуляторной
роли симпатической иннервации сердца, которую она играет при обычных условиях,
имеются лишь ограниченные сведения. И эти сведения очень нуждаются в контроле,
вследствие того что они, получены при патологических условиях, созданных
глубоким вивисекционным вмешательством.
Включение в активность
симпатической иннервации сердца в натуральных условиях целого организма,
несомненно, второе доказано только при чрезвычайных и стрессовых
обстоятельствах.
К ним относятся: общее
охлаждение организма, сильные эмоциональные напряжения, яростная борьба за
жизнь, геморрагия, высокоамплитудные колебания кровяного давления, угроза
ослабления активности вследствие утомления от крайнего и длительного
напряжения. Подобные условия требуют для своего преодоления повышенной
активности сердечнососудистой системы и общей мобилизации энергетических средств
организма, которые и осуществляются с помощью симпатической системы.
Влияние на энергетические
ресурсы и ускоренное их использование, вероятно, являются основной
регуляторно-трофической задачей симпатической иннервации. Медиаторы
симпатических нервов сердца - норадреналин, адреналин (Бинг, Ленци и др.,
1959), возможно, и предназначены для непосредственного вмешательства в
химическую динамику эффекторных клеток сердца (Коштоянц, 1951). Они ускоряют
протекание обменно-энергетических процессов при повышенной активности сердца.
Вероятно, в соответствии со своеобразием клеточного обмена пейсмекерной ткани в
зоне синоатриального узла освобождается и дофамин как фактор, непосредственно
ускоряющий генерацию автоматических возбуждений. Действительно, под влиянием
симпатических воздействий наблюдаются изменения в энергетических ресурсах
сердца (гликоген, креатин-фосфат и аденозинтрифосфорная кислота). Зависимость
обменных процессов в сердечной мышце от симпатической иннервации обнаружена в
более поздних исследованиях (Говырин, 1967). При стимуляции симпатических
нервов сердца выявлено повышение активности фосфорилазы.
Симпатическая иннервация сердца
при очень сильном возбуждении (например, электрическая стимуляция, рефлекторное
возбуждение при выключении жизненно важной афферентной информации- ваготомия)
сама по себе за счет стимуляторного влияния на энергетику эффекторных клеток
может повысить частоту наличного автоматического ритма сердца. Однако ранее
показано, что симпатическое учащение обычно не достигает предельно высоких
частот. Имея регуляторно-трофическое назначение, симпатическая иннервация
сердца в нормальных условиях рефлекторного возбуждения, вероятно, не оказывает
непосредственного ускорительного влияния на пейсмекер сердца, т. е. не является
пусковым ускорительным механизмом. Ее влияния на пейсмекер реализуются на фоне
учащения сердцебиений, обусловленных влиянием парасимпатической иннервации.
Своим влиянием на энергетику клеток симпатическая иннервация позволяет
удерживать на крайне высоких уровнях ритм разрядов в пейсмекере, а повышенные
скорости проведения возбуждений - в проводящей системе и усиливает сокращение
миокарда.
В отличие от вспомогательной
регуляторной роли по отношению к пейсмекеру инотропное действие симпатической
иннервации может проявляться при постоянном ритме и независимо от других
влияний на сердце. Парасимпатическая иннервация непосредственно, видимо, не
может оказывать положительно инотропное действие на миокард желудочков сердца
млекопитающих, а косвенные средства усиления сокращений (через активацию
диастолы и ритмоинотропную зависимость), вероятно, нуждаются в подкреплении
трофическим влиянием симпатической системы. С этой точки зрения отношения между
парасимпатической и симпатической иннервациями сердца при обеспечении высокой гемодинамической
производительности сердца в условиях очень сильных и длительных напряжений
представляются в следующем виде: парасимпатической иннервации принадлежит,
вероятно, пусковая ускорительная роль, симпатическая же иннервация своими
трофическими влияниями обеспечивает соразмерную интенсификацию энергетических
процессов в пейсмекере, в проводящей системе и сократительном миокарде. Этим
влиянием симпатической системы поддерживается на высоком уровне общая
ритмическая активность сердца при крайне высоких частотах сердцебиений.
Положительно инотропные влияния на желудочки симпатическая иннервация может
осуществлять и вне зависимости от парасимпатических влияний на сердце.
Оба нервных «прибора» сердца,
вероятно, дополняют друг друга и по своим интимным механизмам: регуляторные
влияния парасимпатической иннервации изменяют трансмембранную ионную
проницаемость (калиево-натриевые ионные токи) и, вероятно, межклеточные
отношения в пейсмекере и проводящей системе. Этим объясняются быстрые
двусторонние изменения в ритме, ритмо-инотропной и ритмо-диастолической
зависимости.
По своему внутреннему механизму
и регуляторному назначению симпатическая иннервация сердца, вероятно, близка и,
может быть, даже тождественна адаптационно-трофическому влиянию симпатической
иннервации скелетных мышц. Согласно Орбели, адаптационно-трофическое влияние
синоптической иннервации на скелетную мышцу моделирует ее сокращения, которые
вызываются импульсацией моторных нервов и этим приспосабливают их интенсивность
к текущим потребностям организма. Эти адаптационные изменения активности
скелетных мышц обусловливаются трофическим действием симпатической иннервации,
которые сопровождаются изменениями в скоростях химических реакций и самого
физико-химического состояния мышечной ткани ее функциональных свойств (Орбели,
1960, 1962). Исследованиями Орбели и других установлено, что симпатическая
система при совместном генерализованном распределении, располагая
специализированными гуморальными средствами конечного трофического влияния -
катехоламинами, простирает свою адаптивно-трофическую регуляцию на все
органы с соматической иннервацией. Вследствие регуляторных влияний
симпатической системы повышается интенсивность активности органов, которые
стимулируются соматической иннервацией, изменяется чувствительность сенсорных
аппаратов, синапсов ЦНС и вегетативных ганглиев.
Характерная особенность всех
регуляторных влияний симпатической иннервации - это то, что они функционально
проявляются в изменениях в ответ на пусковые соматические импульсы, адекватные сенсорные
стимулы или в изменениях при эффектах от текущих процессов интегративного
взаимодействия в ЦНС (пример сеченовского торможения спинномозговых рефлексов).
Следовательно, симпатические
влияния выявляются и реализуются через активность соматической нервной системы.
Особенно убедительно это продемонстрировано на примере симпатической регуляции
активности скелетной мышцы. Симпатические влияния сами по себе не только не
способны вызвать видимый эффект на скелетной мышце, но, как показано
исследованиями М. В. Кирзона, так называемый симпатический эффект зависит от
ритма моторной импульсации, характера моторного залпа, от состояния
мионевральной связи и реагирующей на моторную импульсацию самой мышцы (Кирзон,
1946; Кирзон, Каменская, 1965).
Симпатическая иннервация
скелетных мышц имеет своим назначением поддержание их работы на высоком уровне,
когда вследствие длительной напряженной деятельности возникает опасность их
ослабления. Таким образом, назначением симпатической иннервации сердца является
не побуждение пейсмекера сердца к учащенным разрядам, а поддержание общей
ритмической активности сердца на том высоком уровне, на который она была
сдвинута парасимпатической регуляцией. Такая роль симпатической регуляции
ритмики сердца весьма вероятна, если иметь в виду, во-первых, что одна
парасимпатическая иннервация способна адаптивно повышать ритм сердцебиений до
крайних пределов.
Однако мобилизация
энергетических резервов сердца при его крайних напряжениях представляется еще
более жизненно важной необходимостью, чем для скелетной мышцы.
Парасимпатическая иннервация сама по себе не может так глубоко изменить
обменно-энергетические процессы, поэтому необходимость в дополнительной
адаптивно-трофической регуляции сердца представляется очевидной.
Вместе с подобным регуляторным
значением по отношению к адаптации ритмической активности сердца симпатическая
иннервация может включаться и независимо от парасимпатического регуляторного
аппарата. Это может иметь место при необходимости нейрогенного повышения силы
сокращений рабочего миокарда независимо от изменений в ритме. У млекопитающих
при затрудненном выбросе крови (или при его временном исключении) возбуждение
симпатической иннервации сопровождается мощным положительным эффектом. Этот
механизм положительного инотропного действия, вероятно, включается в условия,
когда сердце работает в условиях затрудненного выброса или на предельном уровне
его гемодинамической производительности.
Поддержание деятельности
сердца, скелетных мышц и других форм функциональной активности на достаточно
высоком уровне при длительных и утомительных напряжениях организма, нередко
является жизненной необходимостью. Известно, что трудные и экстремальные
условия, требующие крайнего напряжения организма, преодолеваются с помощью
симпатоадреналовой системы.
Изложенное представление о
взаимодействии парасимпатических и симпатических регуляторных влияний на сердце
- это следствие логического синтеза экспериментальных данных, показавших что
одна парасимпатическая иннервация способна обеспечить и тормозящие и ускоряющие
влияния на сердце, с концепцией Кеннона н его школы, согласно которой
симпато-адреналовая система используется при чрезвычайных или стрессовых
условиях, требующих от организма крайних напряжений. В основу этого синтеза
положена теория Орбели об адаптивно-трофической функции симпатической системы.
Вследствие того что сведения о
взаимодействии отделов вегетативной системы и принадлежащих им отдельных
иннервационных аппаратов в условиях целого организма отсутствуют, в настоящее
время приходится ограничиться этим в известной мере спекулятивным
представлением.
Некоторое значение для
рассматриваемой проблемы имеют результаты опытов с одновременным электрическим
раздражением парасимпатических и симпатических нервов, в которых имитируется
взаимодействие между парасимпатическими и симпатическими влияниями на сердце.
Поэтому результаты этих опытов
в их конкретном выражении нельзя переносить на возможные натуральные
регуляторные эффекты от взаимодействия иннервационных приборов сердца. Они дают
лишь возможность убедиться в том, что эффекты от комбинированных воздействий,
вызванных электрическими стимулами целых нервных аппаратов, очень разнообразны
и в большей части не обнаруживают антагонистических отношений между
парасимпатическими и симпатическими влияниями.
Присоединение раздражения
симпатических нервов к стимуляции ядер вагуса может усилить слабый тормозной
эффект. Активация симпатикуса на фоне вагусного ускользания, как правило, ведет
к углублению торможения. Вагусный ускорительно-усилительный эффект в зависимости
от степени его выраженности может быть ослаблен или, наоборот, усилен
присоединением стимуляции симпатической иннервации. Суммарный эффект зависит и
от степени выраженности обособленного симпатического эффекта. В относительно
редких случаях возможна и взаимная компенсация эффектов. Когда стимуляция
симпатических нервов опережает вагусную, преобладающим эффектом обычно является
симпатический.
Разнообразные варианты эффектов
от совмещения стимуляции парасимпатических и симпатических нервов сердца позволяют
убедиться в том, что регуляторные возможности нервных влияний на сердце могут
весьма широко разнообразиться и, вероятно, совершенствоваться, когда нервные
воздействия на сердце комбинируются из импульсации парасимпатических и
симпатических путей.
Вопрос о том, при каких
условиях и как организуется регуляторное взаимодействие между
парасимпатическими и симпатическими влияниями, является одним из наиболее
важных нерешенных вопросов проблемы нервной регуляции сердца.
Естественно, что организация
комбинированных регуляторных влияний, осуществляющихся по обоим иннервационным
приборам сердца, является функцией центральных аппаратов, координирующих
адаптивные нервные влияния на сердце и его активность.
1.2 Парасимпатическая регуляция
Парасимпатическая нервная система - часть
автономной нервной системы, связанная с симпатической нервной системой и
функционально ей противопоставляемая. В парасимпатической нервной системе
ганглии расположены непосредственно в органах или на подходах к ним, поэтому
преганглионарные волокна длинные, а постганглионарные - короткие. Термин
парасимпатическая - т. е. околосимпатическая был предложен Д. Н. Ленгли в конце
XIX - начале XX века.
Эмбриональным источником для парасимпатической
системы является ганглиозная пластинка. Парасимпатические узлы головы
образуются путем миграции клеток из среднего и продолговатого мозга.
Периферические парасимпатические ганглии пищеварительного канала происходят из
двух участков ганглиозной пластинки - «вагусного» и пояснично-крестцового.
У млекопитающих в парасимпатической нервной
системе выделяют центральный и периферический отдел. Центральный включает ядра
головного мозга и крестцового отдела спинного мозга.
Основную массу парасимпатических узлов
составляют мелкие ганглии, диффузно разбросанные в толще или на поверхности
внутренних органов. Для парасимпатической системы характерно наличие длинных
отростков у преганглионарных нейронов и чрезвычайно коротких - у
постганглионарных.
Головной отдел подразделяют на среднемозговую и
продолговатомозговую части. Среднемозговая часть представлена ядром
Якубовича-Эдингера-Вестфаля, расположенным вблизи передних бугров четверохолмия
на дне Сильвиева водопровода. В продолговатомозговую часть входят ядра VII, IX,
X черепно-мозговых нервов.
Преганглионарные волокна от ядра
Эдингера-Вестфаля выходят в составе глазодвигательного нерва, и заканчиваются
на эффекторных клетках ресничного ганглия. Постганглионарные волокна вступают в
глазное яблоко и идут к аккомодационной мышце и сфинктеру зрачка. VII (лицевой)
нерв тоже несет парасимпатическую компоненту. Через поднижнечелюстной ганглий
он иннервирует подчелюстную и подъязычную слюнные железы, а переключаясь в
крылонебном ганглии - слезные железы и слизистую носа.
Волокна парасимпатической системы также входят в
состав IX (языкоглоточного) нерва. Через околоушной ганглий он иннервирует
околоушные слюнные железы.
Основным парасимпатическим нервом является
блуждающий нерв, который наряду с афферентными и эфферентными
парасимпатическими волокнами включает чувствительные и двигательные
соматические, и эфферентные симпатические волокна. Он иннервирует практически
все внутренние органы до ободочной кишки.
Ядра спинномозгового центра располагаются в
области II-IV крестцовых сегментов, в боковых рогах серого вещества спинного
мозга. Они отвечают за иннервацию ободочной кишки и органов малого таза.
Преимущественно нейроны парасимпатической
нервной системы являются холинергическими. Хотя известно, что наряду с основным
медиатором постганглионарные аксоны одновременно выделяют пептиды (например,
вазоактивный интестинальный пептид (VIP)). Кроме того, у птиц в ресничном
ганглии наряду с химической передачей присутствует и электрическая. Известно,
что парасимпатическая стимуляция в одних органах вызывает тормозное действие, в
других - возбуждающий ответ. В любом случае действие парасимпатической системы
противоположно симпатической (исключение - действие на слюнные железы, где и
симпатическая, и парасимпатическая нервная система вызывают активацию желез).
Парасимпатическая нервная система иннервирует
радужную оболочку, слезную железу, подчелюстную и подъязычную железу,
околоушную железу, легкие и бронхи, сердце (уменьшение частоты и силы сердечных
сокращений), пищевод, желудок, толстую и тонкую кишку (усиление секреции
железистых клеток).
Преганглионарные парасимпатические сердечные
волокна идут в составе ветвей, отходящих от блуждающих нервов с обеих сторон в
области шеи. Волокна от правого блуждающего нерва иннервируют преимущественно
правое предсердие, и особенно обильно синоатриальный узел. К
атриовентрикулярному узлу подходят главным образом волокна от левого
блуждающего нерва. Вследствие этого правый блуждающий нерв влияет
преимущественно на частоту сокращений сердца, а левый - на атриовентрикулярное
проведение. Парасимпатическая иннервация желудочков сердца выражена слабо, и
функциональное значение ее представляется спорным.
Под действием ацетилхолина замедляется
спонтанная диастолическая деполяризация в клетках синусового узла и, как
следствие, снижается ЧСС. Ацетилхолин замедляет также проведение и укорачивает
эффективный рефрактерный период в предсердиях; оба эти эффекта способствуют
возникновению и поддержанию предсердных аритмий .
С другой стороны, ацетилхолин замедляет
проведение и укорачивает эффективный рефрактерный период в АВ - узле, уменьшая
тем самым частоту проходящих к желудочкам импульсов (и, следовательно,
сокращений желудочков) при мерцательной аритмии и трепетании предсердий.
Отрицательный инотропный эффект ацетилхолина
обусловлен тормозным действием на симпатические окончания и прямым влиянием на
миокард предсердий. Его эффект на желудочки слабо выражен в связи с их
незначительной холинергической иннервацией.
Маловероятна и прямая парасимпатическая
регуляция ОПСС - холинергическая иннервация сосудов также слабая. В то же время
возможно непрямое действие парасимпатических нервов на сосуды, обусловленное
торможением выделения норадреналина из симпатических окончаний.
1.3 Симпатическая регуляция
физиологический сердечный мышца парасимпатический
Исторически симпатическая часть возникает как
сегментарный отдел, поэтому и у человека она частично сохраняет сегментарный
характер строения. Симпатический отдел по своим основным функциям является
трофическим. Он осуществляет усиление окислительных процессов, потребление
питательных веществ, усиление дыхания, учащение деятельности сердца, увеличение
поступления кислорода к мышцам. Центральный отдел симпатической части
Центральный отдел симпатической части располагается в боковых рогах спинного
мозга на уровне С8, Th1-L3, в substantia intermedia lateralis. От него отходят
волокна, иннервирующие непроизвольные мышцы внутренних органов, органов чувств
(глаза), железы. Кроме того, здесь располагаются сосудодвигательные и
потоотделительные центры.
Считают (и это подтверждается клиническим
опытом), что различные отделы спинного мозга оказывают влияние на трофику,
терморегуляцию и обмен веществ. Периферический отдел симпатический части
Периферический отдел симпатической части образуется, прежде всего, двумя
симметричными стволами, trunci sympathici dexter, et sinister, расположенными
по бокам позвоночника на всем его протяжении от основания черепа до копчика,
где оба ствола своими каудальными концами сходятся в одном общем узле. Каждый
из этих двух симпатических стволов слагается из ряда нервных узлов первого
порядка, соединяющихся между собой посредством продольных межузловых ветвей,
rami interganglionares, состоящих из нервных волокон. Кроме узлов симпатических
стволов (ganglia trunci sympathici), в состав симпатической системы входят
указанные выше ganglia intermedia. Симпатический ствол, начиная с верхнего
шейного узла, содержит также элементы парасимпатической части вегетативной и
даже анимальной нервных систем. Отростки клеток, заложенных в боковых рогах
тораколюмбального отдела спинного мозга, выходят из спинного мозга через
передние корешки и, отделившись от них, идут в составе rami communicantes albi
к симпатическому стволу.
Здесь они или соединяются синапсом с клетками
узлов симпатического ствола, или же, пройдя через его узлы без перерыва,
достигают одного из промежуточных узлов. Это так называемый преганглионарный
путь. От узлов симпатического ствола или (если там не было перерыва) от
промежуточных узлов отходят безмиелиновые волокна постганглионарного пути,
направляющиеся к кровеносным сосудам и внутренностям. Поскольку симпатическая
часть имеет соматическую часть, она связана со спинномозговыми нервами,
обеспечивающими иннервацию сомы. Эта связь осуществляется посредством серых
соединительных ветвей, rami communicantes grisei, которые представляют собой
участок постганглионарных волокон на протяжении от узлов симпатического ствола
до n. spinalis. В составе rami communicantes grisei и спинномозговых нервов
постганглионарные волокна распространяются в сосудах, железах и мышцах,
поднимающих волосы кожи туловища и конечностей, а также в скелетной
мускулатуре, обеспечивая ее трофику и тонус. Таким образом, симпатическая часть
соединяется с анимальной нервной системой посредством двоякого рода
соединительных ветвей: белых и серых, rami communicantes albi et grisei. Белые
соединительные ветви (миелиновые) имеют в своем составе преганглионарные
волокна. Они идут от центров симпатической части через передние корешки к узлам
симпатического ствола. Поскольку центры лежат на уровне грудных и верхних
поясничных сегментов, то и rami communicantes albi имеются лишь в пределах от I
грудного до III поясничного спинномозгового нерва. Rami communicantes grisei,
постганглионарные волокна, обеспечивают вазомоторные и трофические процессы
сомы; они соединяют симпатический ствол со спинномозговыми нервами на всем его
протяжении. Шейный отдел симпатического ствола имеет связь и с черепными
нервами.
Следовательно, все сплетения анимальной нервной
системы содержат в составе своих пучков и нервных стволов волокна симпатической
части, чем подчеркивается единство этих систем. Симпатический ствол каждый из
двух симпатических стволов подразделяют на четыре отдела: шейный, грудной,
поясничный (или брюшной) и крестцовый (или тазовый). Шейный отдел простирается
от основания черепа до шейки I ребра; располагается позади сонных артерий на
глубоких мышцах шеи. В его состав входят три шейных симпатических узла:
верхний, средний и нижний. Ganglion cervicale superius является самым крупным
узлом симпатического ствола, имея длину около 20 мм и ширину 4-6 мм. Лежит он
на уровне II и части III шейных позвонков позади внутренней сонной артерии и
медиально от п. vagus. Ganglion cervicale medium небольшой величины,
располагается обыкновенно в месте перекреста a. thyroidea inferior с сонной,
артерией, нередко отсутствует или может распадаться на два узелка. cervicale
inferius довольно значительной величины, расположен позади начальной части
позвоночной артерии; нередко сливается с I, а иногда и II грудным узлом,
образуя общий шейно-грудной, или звездчатый, узел, ganglion cervicothoracicum
s. ganglion stellatum. От шейных узлов отходят нервы для головы, шеи и груди.
Их можно разделить на восходящую группу, направляющуюся к голове, на нисходящую
- опускающуюся к сердцу, и группу для органов шеи. Нервы для головы отходят от
верхнего и нижнего шейных узлов и делятся на группу, проникающую в полость
черепа, и группу, подходящую к голове снаружи. Первая группа представлена n.
caroticus interims, отходящим от верхнего шейного узла, и n. vertebralis,
отходящим от нижнего шейного узла. Оба нерва, сопровождая одноименные артерии,
образуют вокруг них сплетения: plexus caroticus interims и plexus vertebralis;
вместе с артериями они проникают в полость черепа, где анастомозируют между
собой и дают ветви к сосудам мозга, оболочкам, гипофизу, стволам III, IV, V, VI
пар черепных нервов и барабанному нерву. Plexus caroticus intemus продолжается
в plexus cavernosus, которое окружает a. carotis interna на участке прохождения
ее через sinus cavernosus. Ветви сплетений распространяются, кроме самой
внутренней сонной артерии, также по ее разветвлениям.
Из ветвей plexus caroticus internus следует
отметить n. petrosus profundus, который присоединяется к n. petrosus major и
вместе с ним образует n. canalis pterygoidei, подходящий через одноименный
канал к ganglion pterygopalatinum. Вторая группа симпатических нервов головы,
наружная, составляется двумя ветвями верхнего шейного узла, nn. carotid
externi, которые, образовав сплетение вокруг наружной сонной артерии,
сопровождают ее разветвления на голове. От этого сплетения отходит стволик к
ушному узлу, gangl. oticum; от сплетения, сопровождающего лицевую артерию,
отходит ветвь к поднижнечелюстному узлу, gangl. submandibulare. Через
посредство ветвей, входящих в сплетения вокруг сонной артерии и ее ветвей,
верхний шейный узел дает волокна к сосудам (вазоконстрикторы) и железам головы:
потовым, слезной, слизистым и слюнным, а также к мышцам волос кожи и к мышце,
расширяющей зрачок, m. dilatator pupillae.
Центр расширения зрачка, centrum ciliospinale,
находится в спинном мозге на уровне от VIII шейного до II грудного сегмента.
Органы шеи получают нервы от всех трех шейных узлов; кроме того, часть нервов
отходит от межузловых участков шейного отдела симпатического ствола, а часть -
от сплетений сонных артерий. Веточки от сплетений следуют по ходу ветвей
наружной сонной артерии, носят одноименные названия и вместе с ними подходят к
органам, в силу чего число отдельных симпатических сплетений равно числу
артериальных ветвей. Из нервов, отходящих от шейной части симпатического
ствола, отмечают гортанно-глоточные ветви от верхнего шейного узла - rami
laryngopharyngei, которые частью идут с n. laryngeus superior (ветвь n. vagi) к
гортани, частью спускаются к боковой стенке глотки; здесь они вместе с ветвями
языкоглоточного, блуждающего и верхнего гортанного нервов образуют глоточное
сплетение, plexus pharyngeus. Нисходящая группа ветвей шейной части симпатического
ствола представлена nn. cardiaci cervicales superior, medius et inferior,
отходящими от соответствующих шейных узлов.
Шейные сердечные нервы спускаются в грудную
полость, где вместе с симпатическими грудными сердечными нервами и ветвями
блуждающего нерва участвуют в образовании сердечных сплетений. Грудной отдел
симпатического ствола располагается впереди шеек ребер, прикрыт спереди
плеврой. В его состав входят 10-12 узлов более или менее треугольной формы.
Грудной отдел характеризуется присутствием белых соединительных ветвей, rami
communicantes albi, соединяющих передние корешки спинномозговых нервов с узлами
симпатического ствола.
Ветви грудного отдела: 1) Nn. cardiaci thoracici
отходят от верхних грудных узлов и участвуют в образовании plexus cardlacus; 2)
rami communicantes grisei, безмиелиновые - к межреберным нервам (соматическая
часть симпатического отдела); 3) rami pulmonales - к легким, образуют plexus
pulmonalis; 4) rami aortici образуют сплетение на грудной аорте, plexus
aorticus thoracicus, и частью на пищеводе, plexus esophageus, а также на
грудном протоке (во всех указанных сплетениях принимает участие и п. vagus); 5)
nn. splanchnici major et minor, большой и малый внутренностные нервы; n.
splanchnicus major начинается несколькими корешками, отходящими от V-IX грудных
узлов; корешки n. splanchnicus major идут в медиальном направлении и сливаются
на уровне IX грудного позвонка в один общий ствол, проникающий через промежуток
между мышечными пучками ножек диафрагмы в брюшную полость, где он входит в
состав plexus coeliacus; n. splanchnicus minor начинается от X-XI грудных узлов
и также входит в plexus coeliacus, проникая через диафрагму с большим
внутренностным нервом.
В этих нервах проходят сосудосуживающие волокна,
как это видно из того обстоятельства, что при перерезке этих нервов сосуды
кишечника сильно переполняются кровью; в nn. splanchnici содержатся волокна,
тормозящие движение желудка и кишок, а также волокна, служащие проводниками
ощущений от внутренностей (афферентные волокна симпатической части).
Поясничный, или брюшной, отдел симпатического ствола состоит из четырех, иногда
из трех узлов. Симпатические стволы в поясничном отделе расположены на более
близком расстоянии один от другого, чем в грудной полости, так что узлы лежат
на переднебоковой поверхности поясничных позвонков вдоль медиального края m.
psoas major. Rami communicdntes albi имеются только с двумя или тремя верхними
поясничными нервами. От брюшного отдела симпатического ствола на всем
протяжении отходит большое количество ветвей, которые вместе с nn. splanchnici
major et minor и брюшными отделами блуждающих нервов образуют самое большое
непарное чревное сплетение, plexus coeliacus.
В формировании чревного сплетения участвуют
также многочисленные спинномозговые узлы (С5-L3), аксоны их нейроцитов. Оно
лежит на передней полуокружности брюшной аорты, позади поджелудочной железы, и
окружает начальные части чревного ствола (truncus coeliacus) и верхней
брыжеечной артерии. Сплетение занимает участок между почечными артериями,
надпочечниками и аортальным отверстием диафрагмы и включает - парный чревный
узел, ganglion coeliacum, и иногда непарный верхний брыжеечный узел, ganglion
mesentericum superius. От чревного сплетения отходит ряд меньших парных
сплетений к диафрагме, надпочечникам, дочкам, а также plexus testicularis
(ovaricus), следующих по ходу одноименных артерий. Имеется также ряд непарных
сплетений к отдельным органам по стенкам артерий, название которых они носят.
Из последних верхнее брыжеечное сплетение, plexus mesentericus superior,
иннервирует поджелудочную железу, тонкую и толстую кишку до половины протяжения
поперечной ободочной. Вторым главным источником иннервации органов полости
живота является сплетение на аорте, plexus aorticus abdominalis, составленное
из двух стволов, отходящих от чревного сплетения, и веточек от поясничных узлов
симпатического ствола.
От аортального сплетения отходит нижнее
брыжеечное сплетение, plexus mesentericus inferior, для поперечной и нисходящей
части ободочной кишки, сигмовидной и верхних отделов rectum (plexus rectals
superior). У места отхождения plexus mesentericus inferior располагается
одноименный узел, gangl. mesentericum inferius. Его постганглионарные волокна
идут в тазе в составе nn. Hypogastrici. Аортальное сплетение продолжается
вначале в непарное верхнее подчревное сплетение, plexus hypogastricus superior,
которое у мыса раздваивается и переходит в сплетение таза, или нижнее
подчревное сплетение (plexus hypogastricus inferior s. plexus pelvinus).
Волокна, происходящие из верхних поясничных сегментов, по своей функции
являются сосудодвигательными (вазоконстрикторами) для полового члена,
двигательными для матки и сфинктера мочевого пузыря.
Крестцовый, или тазовый, отдел имеет обычно
четыре узла; располагаясь на передней поверхности крестца вдоль медиального
края передних крестцовых отверстий, оба ствола книзу постепенно сближаются друг
с другом и затем оканчиваются в одном общем непарном узле - ganglion impar,
находящемся на передней поверхности копчика. Узлы тазового отдела, как и
поясничного, связаны между собой не только продольными, но и поперечными
стволиками. От узлов крестцового отдела симпатического ствола отходит ряд
ветвей, которые соединяются с ветвями, отделяющимися от нижнего брыжеечного
сплетения, и образуют пластинку, протягивающуюся от крестца к мочевому пузырю;
это так называемое нижнее подчревное, или тазовое, сплетение, plexus
hypogastricus inferior s. plexus pelvinus.
Сплетение имеет свои узелки - ganglia pelvina. В
сплетении различают несколько отделов: 1) передненижний отдел, в котором
выделяют верхнюю часть, иннервирующую мочевой пузырь - plexus vesicalis, и
нижнюю, снабжающую у мужчин предстательную железу (plexus prostaticus),
семенные пузырьки и семявыносящий проток (plexus deferentialis) и пещеристые
тела (nn. cavernosi penis); 2) задний отдел сплетения снабжает прямую кишку
(plexus rectales medii et inferiores). У женщин выделяют еще средний отдел,
нижняя часть которого дает ветви к матке и влагалищу (plexus uterovaginal),
пещеристым телам клитора (nn. cavernosi clitoridis), а верхняя - к матке и
яичникам. От узлов крестцового отдела симпатического ствола отходят
соединительные ветви, rami communicantes, присоединяющиеся к спинномозговым
нервам, иннервирующим нижнюю конечность. Эти соединительные ветви составляют
соматическую часть симпатического отдела вегетативной нервной системы,
иннервирующую нижнюю конечность. В составе rami communicantes и спинномозговых
нервов нижней конечности находятся постганглионарные волокна, которые
распространяются в сосудах, железах и мышцах волос кожи, а также в скелетной
мускулатуре, обеспечивая ее трофику и тонус.
1.4 Гуморальная регуляция
Каждый организм, безразлично - одноклеточный или
многоклеточный, является единым целым. Все его органы тесно связаны друг с
другом и управляются общим, точным, слаженным механизмом. Чем выше развит
организм, тем сложнее и тоньше устроена, тем большее значение имеет для него
нервная система. Но в организме существует и так называемая гуморальная
регуляция, и координация работы отдельных органов и физиологических систем. Она
осуществляется при помощи особых высокоактивных химических веществ,
накопляющихся в крови и тканях в процессе жизнедеятельности организма.
Клетки, ткани, органы выделяют в окружающую
тканевую жидкость продукты своего обмена веществ, так называемые метаболиты. Во
многих случаях это - простейшие химические соединения, конечные продукты
последовательных внутренних превращений, протекающих в живой материи. Образно
выражаясь, это «отходы производства». Но нередко такие отходы обладают
необычайной активностью и способны вызвать целую цепь новых физиологических
процессов, образование новых химических соединений и специфических веществ.
К числу более сложных продуктов обмена относятся
и гормоны, выделяемые в кровь железами внутренней секреции (надпочечниками,
гипофизом, щитовидной железой, половыми железами и т.д.), и медиаторы -
передатчики нервного возбуждения. Это сильнодействующие химические вещества,
обычно довольно сложного состава, участвующие в подавляющем большинстве
жизненных процессов. Они оказывают самое решительное влияние на разные стороны
деятельности организма: действуют на психическую деятельность, ухудшают или
улучшают настроение, стимулируют физическую и умственную работоспособность,
возбуждают половую активность. Любовь, зачатие, развитие плода, рост,
созревание, инстинкты, эмоции, здоровье, болезни проходят в нашей жизни под
знаком эндокринной системы.
Вытяжки из желез внутренней секреции и химически
чистые препараты гормонов, искусственно полученные в лаборатории, применяются
при лечении различных заболеваний. Инсулин, кортизон, тироксин, половые гормоны
продаются в аптеках. Очищенные и синтетические гормональные препараты приносят
огромную пользу людям. Учение о физиологии, фармакологии и патологии органов
внутренней секреции превратилось за последние годы в один из важнейших разделов
современной биологии.
Но в живом организме клетки эндокринных желез
выбрасывают в кровь не химически чистый гормон, а комплексы веществ, содержащие
сложные продукты обмена (белкового, липоидного, углеводного), тесно связанные с
активным началом и усиливающие или ослабляющие его действие.
Все эти неспецифические вещества принимают самое
активное участие в гармоническом регулировании жизненных функций организма.
Поступая в кровь, лимфу, тканевую жидкость, они играют важную роль в
гуморальной регуляции физиологических процессов, осуществляемой через жидкие
среды.
Гуморальная регуляция тесно связана с нервной и
образует совместно с ней единый нейрогуморальный механизм регуляторных
приспособлений организма. Нервные и гуморальные факторы столь тесно
переплетаются друг с другом, что всякое противопоставление их недопустимо, как
и недопустимо расчленение процессов регуляции и координации функций в организме
на автономные ионные, вегетативные, анимальные компоненты. Все эти виды
регуляции настолько тесно связаны друг с другом, что нарушение одного из них,
как правило, дезорганизует и остальные.
На ранних этапах эволюции, когда нервная система
отсутствует, взаимосвязь между отдельными клетками и даже органами
осуществляется гуморальным путем. Но по мере развития нервного аппарата, по
мере его совершенствования на высших ступенях физиологического развития
гуморальная система все больше и больше подчиняется нервной.
Образующиеся под влиянием нервных импульсов
разнообразные продукты обмена веществ (метаболизма), известные под названием
метаболитов, в свою очередь могут действовать как раздражители на клетки
органов или окончания чувствительных нервов, вызывая рефлекторным путем определенные
физиологические, а иногда и патологические процессы.
Влияние нервной системы на химические
превращения в органах и на образование биологически активных веществ подробно
изучено и ни у кого не вызывает сомнений, но далеко не всегда учитывается
влияние, оказываемое химическими соединениями, образующимися в организме, на
состояние самой нервной системы. Деятельность головного и спинного мозга
зависит от кровоснабжения и обмена веществ в самих нервных клетках и нервных
волокнах, от химического состава и физико-химических свойств их микросреды.
Здесь имеет место теснейшая связь, взаимная обусловленность жизненных явлений.
Медиаторы
Катехоламины. Ацетилхолин.
Мысль о том, что передача возбуждения с нервного
окончания на клетки органов осуществляется при помощи химических веществ,
возникла уже давно. Но доказано это было только в 20-х годах нашего столетия.
Вещества, образующиеся при возбуждении, получили название медиаторов
(трансмиттеров) или передатчиков нервного возбуждения. Место их накопления -
окончания нервных волокон, где они появляются в тот момент, когда нервный
импульс приходит в рабочий орган, например в мышцу или железистую клетку. Они
образуются в синапсах, связывающих между собой нервные клетки центральной
нервной системы, в периферических нервных узлах, а также в нервных стволах.
Переход возбуждения с одной клетки на другую
является необычайно сложным процессом, тонкий механизм которого довольно
подробно изучен.
При электронно-микроскопическом исследовании
четко обнаруживается, что синапс состоит из двух соприкасающихся поверхностей,
одна из которых принадлежит аксону, другая - дендриту или телу клетки. При
увеличении в несколько десятков тысяч раз синапс представляется в виде щели,
шириной примерно в 200 А (ангстрем - одна стомиллионная доля сантиметра).
Поверхность аксона, обращенная к синапсу, получила название пресинаптической
мембраны (оболочки), а дендрита - постсинаптической.
В окончании аксона электронный микроскоп
обнаруживает целое скопление крошечных пузырьков (везикулов), наполненных определенным
химическим веществом. Вещество это - передатчик, медиатор, посредник нервного
возбуждения, осуществляющий переход импульса через синапс.
Чаще всего это ацетилхолин или норадреналин,
иногда серотонин, гамма-аминомасляная кислота и т.д. Вопрос о химической
регуляции функций требует специального рассмотрения. Пока что констатируем
факт: передача нервного возбуждения с нейрона на нейрон, с нервного окончания
на орган-исполнитель происходит при участии медиаторов. Это очень важное
обстоятельство, поверить в реальность которого очень долго не могли, а может
быть и не желали физиологи и биохимики.
Без всякого преувеличения можно сказать, что
открытие химической медиации явилось одним из наиболее блестящих, как принято
говорить, «делающих эпоху», открытий биологии XX века.
Различные нейроны - в зависимости от их
расположения, физико-химических свойств, обмена веществ, физиологических
функций - возбуждаются или, наоборот, прекращают свою деятельность
(затормаживаются) под влиянием различных медиаторов.
Отсюда и возникло представление, что существуют
возбуждающие и тормозящие медиаторы. Этому до сих пор окончательно не решенному
вопросу было посвящено немалое количество экспериментальных работ и
теоретических споров. Надо думать, что одни и те же химические вещества, в
зависимости от условий, могут вызывать как возбуждение, так и торможение
функций.
Нервный импульс представляет сложнейший
физико-химический процесс, связанный с перемещением некоторых минеральных
веществ, в частности ионов калия и натрия. В состоянии покоя ионы калия
находятся преимущественно внутри нервной клетки, ионы натрия на ее наружной
поверхности. В протоплазме нервных клеток ионов калия примерно в 30-40 раз
больше, чем в окружающей клетку тканевой жидкости, ионов же натрия - в 8-10 раз
меньше. В соответствии с этим внутри клетки преобладают отрицательные
электрические заряды, вне ее - положительные. В тот момент, когда нервный
импульс приходит в окончание аксона (так называемую синоптическую бляшку),
пузырьки, содержащие медиатор, лопаются. Ацетилхолин или норадреналин
изливаются в синоптическую щель и изменяют проницаемость постсинаптической
мембраны. Это ведет к тому, что ионы калия устремляются из клетки и
располагаются на ее поверхности, обращенной к щели, а ионы натрия входят в
клетку. Электрический заряд мембраны мгновенно изменяется, возникает разница
потенциалов, и импульс переходит с аксона одной клетки на дендрит другой. Как
только импульс прошел синапс, медиатор разрушается, ионы калия снова поступают
в клетку, а ионы натрия выходят из нее.
Для того чтобы понять, как действуют медиаторы,
предпримем небольшую прогулку в физиологическую лабораторию и проделаем
несколько простых, но весьма показательных опытов.
Проще всего использовать для этой цели лягушку.
Не случайно ряд законов жизнедеятельности организма был изучен именно на этом
неприхотливом и очень удобном для эксперимента животном. Деятельность сердца
лягушки можно изучать в течение нескольких суток, если питать его вместо крови
искусственным раствором солей (так называемой жидкостью Рингера), по составу
своему напоминающим плазму крови.
Эту жидкость после того, как она прошла через
сердце, можно собрать в стаканчик и подействовать ею на сердце другой лягушки.
Напомним, что сердцем управляют два нерва: один,
замедляющий его деятельность,- блуждающий нерв, другой, усиливающий и
ускоряющий его,- симпатический.
При раздражении блуждающего нерва слабым
электрическим током сила сердечных сокращений уменьшается, ритм их замедляется,
в то время как раздражение симпатического нерва усиливает их и учащает
деятельность сердца.
Теперь, после этих предварительных замечаний
перейдем к опыту.
Начнем с раздражения блуждающего нерва. Мы сразу
заметим, что сердце стало сокращаться медленно, что сила отдельных сокращений
уменьшилась. Все это открыто много лет назад. Но имеется и кое-что новое в этом
опыте. Если жидкостью Рингера, оттекающей от такого сердца, подействовать на
свежее сердце другой лягушки, оно тоже начнет медленнее и слабее сокращаться.
По-видимому, в жидкости появились вещества, подавляющие работу сердца.
Изменим условия опыта. Будем раздражать
симпатический нерв. Сердце ускорит и усилит свою деятельность, а под
воздействием оттекающей от него жидкости свежее сердце тоже начнет сильнее и
быстрее сокращаться.
Следовательно, медиаторы, образовавшиеся в
нервных окончаниях, передают возбуждение с нерва на рабочий орган. Поэтому они
и называются передатчиками нервного возбуждения. Эти опыты были поставлены в
начале 20-х годов нашего столетия австрийским ученым Леви, впоследствии
Нобелевским лауреатом, и послужили началом учения о химической передаче
нервного возбуждения.
В настоящее время установлено, что вещества,
накапливающиеся в физиологическом растворе поваренной соли, или в жидкости
Рингера, при раздражении блуждающего нерва близки к ацетилхолину, а вещества,
образующиеся при раздражении симпатического нерва,- к адреналину.
Наряду с другими биологически активными
веществами, медиаторы, поступая в кровь, принимают участие в регуляции и
координации физиологических процессов. Из этого следует, что необходимо
различать их роль в медиации и регуляции.
Ацетилхолин - медиатор парасимпатической системы
является сложным эфиром холина и уксусной кислоты. Он образуется при участии
синтезирующего фермента - холинацетилазы, активность которого в клетках
изменяется под влиянием условий среды и тканевого обмена. Ацетилхолин нестоек,
и срок его существования крайне ограничен. Выполнив свою задачу, ацетилхолин,
образовавшийся в нервных окончаниях, мгновенно расщепляется под влиянием
фермента холинэстеразы на свои составные части (уксусную кислоту и холин). До
сих пор принято было считать, что ацетилхолин приспособлен для выполнения
ограниченных задач и действие его сводится к передаче возбуждения с нерва на
эффекторную клетку. Но теперь, в значительной степени работами нашей
лаборатории, установлено, что неиспользованный при передаче возбуждения
ацетилхолин поступает из органов и тканей в кровь и принимает активное участие
в гуморальной регуляции функций. Его действие на клетки сходно с действием
парасимпатических нервов. В крови он захватывается эритроцитами и разносится
током крови по всему организму. При определенных физиологических ситуациях
ацетилхолин переходит из эритроцитов в жидкую часть крови и вызывает
парасимпатические реакции. Количество свободного, активного ацетилхолина в
жидких средах организма характеризует состояние (тонус и реактивность)
парасимпатической нервной системы.
Иначе обстоит дело с медиаторами симпатического
ряда - симпатинами. Доказано, что симпатические реакции в организме протекают
при участии целой группы веществ, известных под общим названием катехоламинов.
Катехоламины - в одно и то же время гормоны и медиаторы симпатоадреналовой
системы. С каждым годом все отчетливее выявляется их роль в приспособительных
реакциях организма, в обмене веществ, в деятельности мышц и сердца, в
кровоснабжении органов, эмоциональном возбуждении, возникновении и степени
болевого ощущения.
Основной, ведущий представитель катехоламинов,
наиболее известный и подробно изученный,- адреналин. Он образуется в мозговом
слое надпочечников и содержание его во внутренней среде организма характеризует
состояние этой важнейшей эндокринной железы нашего организма. Его
непосредственный предшественник, отличающийся от него отсутствием одной
метильной группы (CH3),- норадреналин - обладает функциями гормона мозгового
слоя надпочечников и медиатора центральных и периферических отделов
симпатической нервной системы. Долгие споры о природе симпатинов можно считать
законченными. Норадреналин выделяется окончаниями симпатических нервов и после
выполнения своей функции - передачи нервного возбуждения - вновь захватывается
этими окончаниями.
В 1933 г. бельгийский ученый Бакк высказал
предположение, что симпатины в одних случаях являются адреналином, в других -
норадреналином. Советский биохимик А. М. Утевский предположил, что симпатины -
сложная система адреналина, норадреналина и промежуточных продуктов их обмена.
Но в настоящее время установлено, что симпатическая медиация осуществляется с
помощью только норадреналина. Предшественник норадреналина дофамин - медиатор
симпатических образований в центральной нервной системе. Его отсутствие или
недостаточное образование в определенных участках головного мозга приводит к
тяжелому заболеванию, известному под названием паркинсонизма. Катехоламины
образуются в организме из аминокислот, путем последовательного превращения
фенилаланина в тирозин и дигидрооксифенилаланин (ДОФА). Помимо прямого
медиаторного действия, и катехоламины, и ацетилхолин, поступая в кровь и
тканевую жидкость, принимают самое активное участие в гуморальной регуляции
функций. Они оказывают необычайно сильное влияние на ход физиологических
процессов в организме, действуя и на специфические хеморецепторы и на клетки
органов и тканей. При этом содержание их в крови ничтожно, а активность
необычайно высока.
Возьмем обычную медицинскую пиявку и вырежем у
нее из спины кусочек мышцы. Если погрузить этот кусочек в раствор ацетилхолина
в разведении 1:200 000 000, пиявка начнет сокращаться. Она отвечает на
незначительное количество ацетилхолина, содержащееся в жидкости Рингера, в
крови, в вытяжках из тканей.
Какое же значение имеют медиаторы для передачи
нервного импульса? Этому вопросу посвящено бесчисленное количество
экспериментальных работ, выполненных во всех лабораториях мира. Еще в 1924 г.
казанский физиолог А. Ф. Самойлов высказал предположение, что нервы передают
возбуждение на мышцу посредством каких-то химических веществ. Вслед за ним к
такому же выводу пришел выдающийся английский физиолог Ч. Шеррингтон. То, что казалось
полстолетия назад лишь мало обоснованным предположением, сегодня излагается во
всех учебниках физиологии как установленный и не подлежащий сомнению факт. Мало
того, в дальнейшем удалось показать, что нервные стволы не являются пассивными
проводниками импульсов. При возбуждении они выделяют специфические активные
вещества, имеющие большое значение для передачи возбуждения. Медиаторы
образуются как при движении нервного импульса из нервного центра к органу
исполнителю, так и при сигнализации с периферии в центры. Они выделяются
нервными окончаниями при поступлении импульса в эффекторную клетку и аксонами
нейронов при синоптической передаче.
Центростремительные нервные импульсы, возникшие
в кожном рецепторе, проникают через задние корешки в спинной мозг, зрительные
бугры и кору головного мозга. Возбуждение одних клеток вызывает в свою очередь
последовательную активацию других. Возбужденная нервная клетка выделяет
специфические продукты обмена веществ (ацетилхолин, норадреналин, серотонин),
которые, действуя через соответствующие синапсы на соседние клетки, в свою
очередь усиливают или ослабляют их деятельность. Таким образом, возникает
длинная полисинаптическая цепь, по которой нервный импульс передается от клетки
к клетке, с рецептора в центральную нервную систему и из нее в эффектор. А
использованный медиатор разрушается, становится неактивным либо поступает в
кровь и принимает участие в регуляции функций.
Чрезвычайно важное значение для химической
регуляции функций имеет взаимодействие медиатора с рецептором. Рецептор,
принимающий центробежные нервные импульсы, можно рассматривать как устройство,
через которое специфическая информация поступает из нервных окончаний в
клетку-исполнительницу. Одни рецепторы отвечают на действие ацетилхолина
(холинорецепторы разного типа - М и Н), другие - катехоламинов (адренорецепторы
альфа и бета), третьи - серотонина и т.д. Работами многих исследователей, в том
числе и советских, установлено, что чувствительность рецепторов, их способность
приходить в состояние возбуждения под влиянием одного или нескольких медиаторов
в значительной мере определяет физиологические процессы, протекающие в клетках
и органах. Так, например, при экспериментальной гипертонии у животных
чувствительность адренорецепторов к адреналину увеличивается в 2,3 раза, а к
норадреналину - в 3,2 раза. Следовательно, одно и то же количество медиатора
может вызвать у животного, страдающего гипертонией, более значительное
повышение кровяного давления, чем у здорового.
В центральной нервной системе передача возбуждения
с одной клетки на другую также совершается при участии медиаторов. В различных
участках головного и спинного мозга в качестве передатчиков нервного
возбуждения действуют разнообразные химические соединения - адреналин,
норадреналин, дофамин, ацетилхолин, серотонин, гамма-аминомасляная кислота,
глютаминовая кислота и др. Набор определенных медиаторов характерен не для
отдельных структурных образований мозга, а для функциональных систем, в которые
могут входить различные по своему строению нервные образования, объединенные
для выполнения какого-либо действия.
На Международном физиологическом конгрессе в
Токио (1965 г.) возник вопрос, какие же вещества, образующиеся в центральной
нервной системе, следует считать медиаторами?
Доказательством медиаторной роли того или
другого химического вещества можно считать наличие его в телах нейронов и,
особенно, в окончаниях аксонов, способность синтезироваться внутри нервных
клеток, присутствие синтезирующих и расщепляющих его ферментов, существование
связанных, неактивных форм. Медиаторы должны освобождаться при нервных
импульсах, даже вызванных электрическим током.
Тонкие методы электронной микроскопии,
гистохимии, ультрацентрифугирования и т.д. позволили сделать важные выводы о
существовании в центральной нервной системе многочисленных ансамблей нейронов,
каждый из которых имеет не только особые, свойственные лишь ему биохимические,
но и физиологические свойства. В нервной ткани постоянно происходит образование
и распад разнообразных химических передатчиков. Одни из них обладают
возбуждающими, другие - тормозящими свойствами, т.е. существуют, по мнению ряда
исследователей, медиаторы, как усиливающие, так и подавляющие деятельность
отдельных нервных образований.
Доказано существование в мозгу, по крайней мере,
трех биохимических нейронных систем - адренергической, холинергической и
серотонинергической. В первой передача нервного возбуждения осуществляется
норадреналином и его предшественником - дофамином, во второй - ацетилхолином, в
третьей - серотонином.
Скандинавские исследователи составили даже
приблизительную схему распределения этих систем в ткани мозга. Они различают:
1) норадреналиновую нейронную систему, которая локализуется преимущественно в
ретикулярной формации ствола мозга, в гипоталамусе, лимбических структурах
переднего мозга и в коре больших полушарий; 2) дофаминовую систему - в
структурах среднего мозга и подкорковых образованиях (бледном шаре); 3)
серотониновую нейронную систему, проходящую через средний мозг к гипоталамусу и
лимбическим структурам переднего мозга.
Холинергические системы расположены в глубоких
слоях коры мозга, в подкорковых структурах, в гипоталамусе (преимущественно
переднем) и в ретикулярной формации мозгового ствола. Советский ученый И. В.
Орлов, используя тонкую методику отведения электрических потенциалов от
отдельных нейронов головного мозга, показал, что 35,7% обследованных клеток
ретикулярной формации отвечают лишь на болевое раздражение. При этом оказалось,
что 34% из них реагировали на действие ацетилхолина, норадреналина и серотонина,
20,6% отвечали на инъекцию ацетилхолина и норадреналина, 14,7 % - норадреналина
и серотонина, 8,8% - ацетилхолина и серотонина. Лишь 11,8% клеток не давали
реакции на химические раздражения, 2,8% реагировали только на ацетилхолин и
7,3% - только на норадреналин. Исследования эти представляют особый интерес.
Они показывают, что в ретикулярной формации ствола мозга существуют специальные
«болевые» клетки, возбуждающиеся под влиянием одного или нескольких медиаторов.
Гистамин
Одним из наиболее важных биологически активных
веществ, образующихся в организме и имеющих непосредственное отношение к
проблеме боли, является гистамин. Химическое строение его хорошо изучено. В
известной мере гистамин можно считать медиатором. Но действие его значительно
сложнее и шире, чем передача нервного возбуждения.
Интерес к гистамину необычайно возрос с тех пор,
как его удалось выделить почти из всех органов человека и животных. Он
постоянно содержится в крови, но количество его не превышает 0,05-0,06 мг на 1
л жидкости. Зато из 1 кг бычьего легкого удается извлечь 30 мг, а из 1 кг
печени - 2,5 мг гистамина. Некоторые авторы утверждают, что 1 кг легких взрослого
человека содержит до 70 мг гистамина, а 1 кг кожи человека - 30 мг. Много
гистамина в селезенке кролика, в сердце коровы, в нервах человека и животных.
Но этот гистамин неактивен. Он связан белками и не в состоянии проявить свое
действие, пока не освободится из связанной формы. Именно освобождение гистамина
играет важнейшую роль в возникновении многих болезненных состояний.
Гистамин образуется в организме из аминокислоты-
гистидина. Под влиянием фермента, гистидин-декарбоксилазы, аминокислота
гистидин превращается в гистамин. Чем активнее фермент, тем интенсивнее он
образует гистамин, тем большие количества этого продукта поступают в кровь и
ткани. По мере образования гистамин связывается тканями, превращаясь в
неактивную форму, либо разрушается ферментом-окислителем, известным под
названием диаминоксидазы, или гистаминазы.
Образование гистамина происходит во многих
органах и тканях, например в печени, почках, поджелудочной железе, но особенно
интенсивно в кишечнике, где оно осуществляется при весьма деятельном участии
кишечных бактерий.
Небольшое количество гистамина поступает в
организм с пищей - с молоком, мясом, некоторыми овощами (шпинатом, помидорами и
др.).
Хотя свободного гистамина в организме
сравнительно немного, действие его необычайно многообразно и охватывает самые
различные физиологические процессы и функции. Под влиянием гистамина повышается
проницаемость сосудистых стенок, расширяются кровеносные капилляры, суживаются
артерии, снижается кровяное давление, усиливается слезотечение, уменьшается выделение
мочи.
В здоровом организме гистамин участвует во
многих физиологических процессах, регулируя деятельность органов, стимулируя их
в одних случаях и ослабляя в других. Как неотъемлемая составная часть входит он
в комплекс биологически активных веществ, циркулирующих в крови или находящихся
в тканях. Без участия гистамина не может осуществляться гуморальная регуляция
функций.
Гистамин - один из сильнейших возбудителей
желудочной секреции. В клинике внутренних болезней широко применяется
гистаминовая проба, которая позволяет решить вопрос о состоянии желез желудка.
Если после введения гистамина в кровь желудочный сок не выделяется, то,
следовательно, слизистая желудка атрофирована, и железы ее, либо вовсе
отсутствуют, либо потеряли способность вырабатывать соляную кислоту и
переваривающие пищу ферменты; это позволяет врачу отличить органические
изменения в желудке от функциональных.
В последнее время много говорят о роли гистамина
в возникновении язвенной болезни желудка. По-видимому, повышенная кислотность
желудочного сока в значительной мере связана с высоким содержанием гистамина в
крови и тканях.
При подкожной инъекции гистамина резко
повышается функция мозгового слоя надпочечников. Гормон этих желез - адреналин
- поступает в кровь и вызывает ряд характерных сдвигов в деятельности
организма. В клинической практике для того, чтобы проверить, нет ли у больного
злокачественной опухоли надпочечника - феохромоцитомы, вводят небольшое
количество гистамина. Если действительно имеется феохромоцитома, она начинает
выбрасывать в кровь свои, во много раз превышающие норму запасы адреналина, что
позволяет поставить диагноз этого заболевания с большой долей вероятности.
Каждому из нас приходилось встречать людей,
особо чувствительных к некоторым обычным, ничем не примечательным воздействиям
на организм. Одни не выносят запаха хвои, другие - свежего сена, третьи -
масляной краски. Сколько раз мы слышим, что один из наших знакомых необычайно
чувствителен к творогу, другой - к землянике, третий - к ракам и т.д. Стоит им
только поесть блюдо, изготовленное из «неугодных» организму продуктов, как кожа
у них покрывается сыпью или волдырями, возникает мучительный зуд, отекают
отдельные участки тела (лицо, веки, кисти рук), а иногда начинаются приступы
какого-то странного беспокойства, крапивницы, мигрени, насморка, бронхиальной
астмы, лихорадки. Все эти состояния - разнообразные проявления аллергии,
связанные с нарушениями гистаминового обмена.
Под влиянием сложных и многообразных процессов,
совершающихся в организме, вызванных некоторыми воздействиями окружающего нас
мира, например охлаждением, перегреванием, ожогом солнечными лучами, введением
некоторых фармакологических препаратов, гистамин освобождается из связанной
формы. Переполненные гистамином тканевые депо - эти «склады», насыщенные
неактивным, связанным гистамином,- начинают опорожняться. В кровь поступает
свободный и весьма агрессивный гистамин. Он повышает проницаемость сосудов,
расширяет капилляры, снижает давление крови, усиливает секрецию желудочного
сока...
Опустевшие депо быстро заполняются вновь
образовавшимся гистамином, который в свою очередь может легко освободиться и
перейти в кровь. Этому «гистаминовому наводнению» организм противопоставляет
мощную систему обороны. Но в некоторых случаях поступление превышает
разрушение, и тогда-то возникает многообразное болезненное состояние, которое
врачи называют аллергическим.
Разумеется, нельзя ставить знак равенства между
аллергией и гистамином. Проявления аллергии не сводятся к действию одного
только гистамина, к гистаминовому отравлению. Но, как правило, без его участия
не возникают аллергические явления.
Гистамин действует в организме при разведении в
десятки миллионов раз. Тысячные доли миллиграмма способны вызвать сокращение
изолированной кишки морской свинки. Накопление сравнительно небольших количеств
гистамина в крови человека вызывает у него тяжелые нарушения самочувствия,
возникновение самых неожиданных расстройств.
Фармакологическая промышленность наших дней
синтезировала несколько десятков препаратов противогистаминного действия
(антигистамины). При введении в организм они препятствуют проявлению его
токсических свойств. Это очень легко доказать в лабораторном опыте. Если
морской свинке ввести димедрол, а затем четырехкратную смертельную дозу
гистамина, свинка остается в живых.
В разных странах Европы и Америки можно
приобрести эти препараты. В СССР - это димедрол, диазолин; за границей -
антерган, супрастин, пипольфен, антистин. Механизм их действия сложен и не
всегда ясен. В основном противогистамины блокируют чувствительные к гистамину
тканевые элементы. Они как бы закрывают цель, в которую «бьет пуля» гистамина.
Разные препараты действуют по-разному. Одни из них подавляют ферменты,
образующие гистамин из гистидина, другие активируют разрушение гистамина, третьи
препятствуют выходу связанного гистамина «на свободу». В определенной степени
все антигистамины влияют на центральную и периферическую нервную систему.
Положив таблетку димедрола на язык, мы чувствуем легкую анестезию, а проглотив
ее - засыпаем глубоким сном, как от сильно действующего снотворного.
Противогистамины получили огромное значение в
связи с проблемой лучевой болезни. Работами многих ученых доказано, что под
влиянием ионизирующей радиации, в том числе и космических лучей, в крови и
тканях резко нарастает количество гистамина. А там, где имеется гистамин,-
нужны противогистамины.
Выбор препарата в каждом отдельном случае
зависит и от характера заболевания, и от наличия препарата в продаже и, в
известной степени, от опыта врача и индивидуальных особенностей больного.
Появление противогистаминных препаратов на
фармакологическом рынке сыграло огромную роль в лечении многих заболеваний.
Последние годы принесли неожиданное открытие. Оказалось, организм вырабатывает
собственные, естественные противогистамины. Тонкими лабораторными
исследованиями удалось показать, что кровь здорового человека способна
нейтрализовать, обезвредить добавленный к ней гистамин. Открытие это
принадлежит французскому ученому Парро, который дал описанному им явлению
название гистаминопексии, а самый эффект обезвреживания гистамина назвал
гистаминопексическим.
Феномен гистаминопексии обусловлен наличием в
нормальной сыворотке крови особого белка - плазмапексина I, который по своему
химическому строению относится к псевдо-гамма-глобулинам. Содержание его в
крови равно 0,4-0,7 % всех белков сыворотки. Плазмапексин связывает не только
гистамин, но также и другие биологически активные вещества (серотонин,
ацетилхолин, окситоцин). Однако он не связывает брадикинин - вещество, имеющее непосредственное
отношение к возникновению боли, о котором мы еще не один раз будем говорить.
В дальнейшем было установлено, что отсутствие
гистаминопексии в сыворотке больных с различными аллергическими заболеваниями
зависит не только от отсутствия плазмапексина I, но и от появления в крови
плазмапексина II, не способного связать гистамин в крови, и антипексина,
подавляющего связывание гистамина плазмапексином I.
В нашей лаборатории И. Л. Вайсфельд подробно
изучила гистаминопексический эффект при различных заболеваниях. Оказалось, что
при некоторых формах патологии (аллергических, нервных) сыворотка крови теряет
способность связывать добавленный к ней гистамин. Это наблюдается у больных
бронхиальной астмой, вазомоторным ринитом, крапивницей. И, хотя содержание в
крови свободного гистамина может быть ниже нормы, из-за отсутствия
гистаминопексического эффекта он отличается особой активностью и даже в
незначительных количествах способен вызывать аллергические явления.
Серотонин (5-окситриптамин)
Приблизительно 25 лет назад три американских
ученых - Рапорт, Грин и Пейдж - выделили из бычьей сыворотки вещество,
способное повышать кровяное давление. Оно и было названо ими серотонином, т.е.
веществом, выделенным из сыворотки (по-латыни serum) и повышающим кровяное давление.
За годы, прошедшие с того времени, свойства серотонина подробно изучены и сам
он синтезирован. Формула его хорошо известна, но роль в регуляции функций
представляется еще довольно спорной.
Можно считать, что серотонин - истинный
медиатор. Он отвечает всем требованиям, предъявляемым к этому типу веществ.
Подобно катехоламинам и ацетилхолину, серотонин осуществляет передачу импульсов
с одной нервной клетки на другую. В головном мозгу имеются группы нейронов,
особенно чувствительных к серотонину, деятельность которых связана с его
образованием и распадом. Нейроны эти сосредоточены преимущественно в ядрах
подбугорья и в среднем мозгу.
В одном литре крови нормального здорового
человека содержится приблизительно 0,06-0,2 г серотонина, причем основная масса
его находится в тромбоцитах.
В течение многих лет ученые разных стран
пытаются разгадать роль серотонина в осуществлении процессов жизнедеятельности
отдельных органов или всего организма. В настоящее время известно, что
серотонин принимает участие в регуляции деятельности головного и спинного
мозга, двигательной, сердечно-сосудистой, пищеварительной, выделительной и
многих других физиологических систем. Обычно серотонин находится в тканях в
виде связанной, неактивной формы. Под влиянием некоторых воздействий, и
особенно при введении различных лекарственных препаратов, например резерпина,
серотонин освобождается из связанной формы. Но жизнь его, как правило,
непродолжительна. Почти во всех тканях содержится моноаминоксидаза - фермент,
довольно быстро инактивирующий серотонин в организме.
В последние годы пристальное внимание
исследователей привлекает значение серотонина в возникновении и развитии
инфаркта миокарда. И хотя в этом вопросе еще далеко нет полной ясности, при
сердечных болях нередко назначают препараты, способные повысить уровень
серотонина в крови. Имеются указания, что накопление серотонина в мышце сердца
предотвращает развитие инфаркта, что, впрочем, требует проверки. Еще слишком
много белых пятен в этой области знания.
Недостаточно изучено также влияние серотонина на
вегетативную нервную систему. В одних случаях его действие подобно возбуждению
симпатической нервной системы, в других - парасимпатической. Не исключено, что
это зависит от дозы введенного препарата, а быть может - от исходного состояния,
вернее, настройки центральных и периферических отделов комплексной
вегетативно-гуморальной - гормональной системы.
Во многих случаях серотонин обладает
противосудорожными и успокаивающими свойствами. Накопляясь в центральной
нервной системе, серотонин подавляет ее активность. Не случайно так много
внимания уделяет медицинская наука изучению обмена серотонина у больных с
различными психическими заболеваниями.
Несомненно, также участие серотонина в
возникновении целого ряда других заболеваний. Видимо, избыточное содержание его
в организме способствует развитию язвенной болезни желудка и 12-перстной кишки.
Установлено, что в некоторых злокачественных опухолях, например, в
феохромоцитоме, можно обнаружить целые «залежи» серотонина. Нередко в моче
больных, страдающих злокачественными опухолями, обнаруживается в большом
количестве 5-окси-индолуксусная кислота - продукт превращения серотонина.
И, наконец, не последнюю роль играет серотонин в
возникновении и развитии болевого синдрома.
В заключение несколько замечаний об участии в
регуляции функций медиаторов, гормонов, различных химических соединений,
образующихся в процессе обмена веществ. Как они осуществляют гуморальную
регуляцию функций? Какова их роль в системе гомеостаза?
Исследования последних лет показали, что для
изучения состояния вегетативной нервной системы у человека и животных можно
использовать методы определения биологической активности крови.
Это значит, что, исследуя содержание в крови
некоторых гормонов и медиаторов, например катехоламинов, ацетилхолина,
гистамина, серотонина и др., можно получить представление о состоянии и
реактивности (т.е. готовности к действию) различных отделов вегетативного
аппарата. Высокое содержание в крови адреналина говорит о повышенной активности
гормонального отдела симпатоадреналовой системы, а высокий уровень
норадреналина - ее нервного отдела.
Накопление в организме ацетилхолина, инсулина,
отчасти гистамина и серотонина является показателем энергичной деятельности
вагоинсулярной (парасимпатической) системы.
Общая биологическая активность крови, т.е.
влияние, которое она оказывает на определенные функции, органы и ткани
(изолированное сердце и кишка лягушки, кровяное давление кошки и кролика,
спинная мышца пиявки, прямая мышца живота лягушки и т.д.), зависит от соотношения
в ней веществ, возбуждающих симпатические и парасимпатические тканевые
элементы.
У здоровых людей биологическая активность крови
волнообразно колеблется в довольно узких границах. При этом соотношение
гормонов, медиаторов, ферментов, различных солей в крови непрерывно меняется,
то повышаясь, то снижаясь. Оно зависит от потребностей организма, различных при
тех или иных условиях, а также от состояния последовательно включающихся по
мере необходимости регуляторных приборов, основная задача которых сводится к
сохранению постоянства внутренней среды.
Нарастание в крови содержания биологически
активных веществ одного ряда (например, симпатических) автоматически вызывает
накопление веществ противоположного действия (парасимпатических),
компенсирующих, уравновешивающих или сглаживающих действие первых. Это - все та
же испытанная и проверенная миллионами лет эволюционного развития система
гомеостаза.
Чрезвычайно сложные, постоянно меняющиеся
количественные и качественные соотношения биологически активных веществ в
жидких средах организма не только отражают, но и определяют состояние различных
отделов вегетативной нервной системы. Поэтому правильнее говорить о
вегетативно-гуморально-гормональном регуляторном комплексе. Совершенно
естественно, что накопление в организме ацетилхолина, вызванное его усиленным
новообразованием, освобождением из связанной формы либо недостаточным захватом
эритроцитами и белками, низкой активностью расщепляющих ферментов, особой
чувствительностью холинорецепторов, создает благоприятную почву для повышения
тонуса парасимпатической нервной системы. Напротив, высокое содержание
катехоламинов в крови и органах является несомненным показателем симпатической
«настройки» вегетативной нервной системы.
При различных заболеваниях регуляторные
механизмы начинают действовать с перебоями, своевременно не включаются и
вызывают извращенные реакции. Взаимоотношения между нервными, гуморальными и
гормональными механизмами нарушаются, результатом чего является возникновение
длительных или кратковременных состояний расстройства регуляции в виде
вегетативных приступов, нарушения сна и бодрствования, разнообразных
болезненных явлений, происхождение которых требует в каждом отдельном случае
специальной расшифровки.
Глава 2. Модель распознания
функционального состояния организма с помощью анализа сердечного ритма
2.1 Основные физиологические
свойства сердечной мышц
Сердечная мышца, как и скелетная, обладает
возбудимостью, способностью проводить возбуждение. Возбудимость сердечной
мышцы. Сердечная мышца менее возбудима, чем скелетная. Для возникновения
возбуждения в сердечной мышце необходимо применить более сильный раздражитель,
чем для скелетной. Установлено, что величина реакции сердечной мышцы не зависит
от силы наносимых раздражений (электрических, меха, химических и т. д.).
Сердечная мышца максимально сокращается и на пороговое, и на более сильное по
величине раздражение.
Проводимость. Волны возбуждения проводятся по
сердечной мышцы и так называемой специальной ткани сердца с неодинаковой
скоростью. Возбуждение по волокнам мышц предсердий распространяется со
скоростью 0,8-1,0 м/с, по волокнам мышц желудочков- 0,8-0,9 м/с, по специальной
ткани сердца-2,0-4,2 м/с.
Сократимость. Сократимость сердечной мышцы имеет
свои особенности. Первыми сокращаются мышцы передние, затем-папиллярные мышцы и
субэндокардиальный слой мышц желудочков. В дальнейшем сокращение охватывает и
внутренний слой желудочков, обеспечивает тем самым движение крови из полостей
желудочков в аорту и легочный ствол. Физиологическими особенностями сердечной
мышцы является удлиненный рефрактерный период и автоматия. Теперь о них
поподробнее.
Рефрактерный период. В сердце в отличие от
других возбудимых тканей имеется значительно выраженный и удлиненный
рефрактерный период. Он характеризуется резким снижением возбудимости ткани в
течение ее активности. Выделяют абсолютный и относительный рефрактерный период
(р.п.). Во время абсолютного р.п. какой бы силы не наносили раздражения на
сердечную мышцу, она не отвечает на него возбуждением и сокращением. Он
соответствует по времени систоле и началу диастолы предсердий и желудочков. Во
время относительного р.п. возбудимость сердечной мышцы постепенно возвращается
к исходному уровню. В этот период мышца может ответить на раздражитель сильнее
порогового. Он обнаруживается во время диастолы предсердий и желудочков.
Сокращение миокарда продолжается около 0.3 с, по времени примерно совпадает с
рефрактерной фазой. Следовательно, в период сокращения сердце неспособно
реагировать на раздражители. Благодаря выраженному р.п., который длится больше
чем период систолы, сердечная мышца неспособна к тетаническому (длительному)
сокращению и совершает свою работу по типу одиночного мышечного сокращения.
Автоматия сердца. Вне организма при определенных
условиях сердце способно сокращаться и расслабляться, сохраняя правильный ритм.
Следовательно, причина соизолированного сердца лежит в нем самом. Способность
сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом,
носит название автоматии.
В сердце различают рабочую мускулатуру, представ
поперечнополосатой мышцей, и атипическую, или специальную, ткань, в которой
возникает и проводится возбуждение.
У человека атипическая ткань состоит из:
синоаурикулярного узла, располагающегося на
задней стенке правого предсердия у места впадения полых вен;
атриовентрикулярного (предсердно-желудочкого)
узла находящегося в правом предсердии вблизи пере между предсердием и
желудочками;
пучка Гиса (председно-желудочковый пучок),
отходящего от атриовентрикулярного узла одним стволом. Пучок Гиса, пройдя через
перегородку между предсердиями и желудочками, делится на две ножки, идущие к
правому и левому желудочку. Заканчивается пучок Гиса в толще мышц волокнами
Пуркинье. Пучок Гиса-это единственный мышечный мостик, соединяющий предсердия с
желудочками. Синоаурикулярный узел является ведущим деятелем сердца (водитель
ритма), в нем возникают импульс, определяющие частоту сокращений сердца. В
норме атриовентрикулярный узел и пучок Гиса являются только передатчиками
возбуждения из ведущего узла к сердечной мышце. Однако им присуща способность к
автоматии, только выражена она в меньшей степени, чем у синоаурикулярного узла,
и проявляется лишь в условиях патологии. Атипическая ткань состоит из
малодифференцированных мышечных волокон. В области синоаурикулярного узла
обнаружено значительное количество нервных клеток, нервных волокон и их
окончаний, которые здесь образуют нервную сеть. К узлам атипической ткани
подходят нервные волокна от блуждающих и симпатических нервов.
2.2 Ритм сердца. Показатели
сердечной деятельности
Ритм сердца и факторы, влияющие на него. Ритм
сердца, т. е. количество сокращений в 1 мин, зависит главным образом от
функционального состояния блуждающих и симпатических нервов. При возбуждении
симпатических нервов частота сердечных сокращений возрастает. Это явление носит
название тахикардии. При возбуждении блуждающих нервов частота сердечных
сокращений уменьшается - брадикардии. Ритм сердца может изменяться под влиянием
гуморальных воздействий, в частности температуры крови, притекающей к сердцу.
Местное раздражение теплом области правого предсердия (локализация ведущего
узла) ведет к учащению ритма сердца при охлаждении этой области сердца
наблюдается противоположный эффект. Местное раздражение теплом или холодом других
участков сердца не отражается на частоте сердечных сокращений. Однако оно может
изменить скорость проведения возбуждений по проводящей системе сердца и
отразиться на силе сердёчных сокращений.
Частота сердечных сокращений у здорового
человека находится в зависимости от возраста.
Что же является показателями сердечной
деятельности?
Показатели сердечной деятельности. Показателями
работы сердца являются систолический и минутный объем сердца.
Систолический, или ударный, объем сердца - тот
объем крови, который поступает из желудочка за одну систолу. Величина
систолического объема зависит от размеров сердца, состояния миокарда и
организма. У взрослого здорового человека при относительном покое систолический
объем каждого желудочка составляет приблизительно 70-80 мл. Таким образом, при
сокращении желудочков в артериальную систему поступает 120-160 мл крови.
Минутный объем сердца - это количество крови,
которое сердце выбрасывает в легочный ствол и аорту за 1 мин. Минутный объем
сердца - это произведение величины систолического объема на частоту сердечных
сокращений в 1 мин. В среднем минутный объем составляет 3-5 л. Систолический и
минутный объем сердца характеризует деятельность всего аппарата кровообращения.
2.3 Оценка функционального состояния
организма с учетом уровня их двигательной активности
Известно, что существует этапность в становлении
механизмов регуляции сердечнососудистой системы, которая проявляется в
характере ее реакции на одно и то же воздействие в разных периодах
постнатального развития (Фролькис В.В., 1975). В связи с этим, в динамике
особенности показателей вегетативной регуляции СР у лиц младшего и среднего
школьного возраста в сформированных группах с разным уровнем двигательной
активности. Особенности изменения регуляции СР при разном уровне двигательной
активности преимущественно обусловлены не возрастом школьника, а тонусом ВНС.
Это согласовывалось с представлением о том, что исходный вегетативный тонус
является одной из одной из важных характеристик, определяющих тип реагирования
(Казначеев В.П., 1980). В силу этого, особенности изменения параметров СР в
группах у школьников разного возраста, были связаны в основном с тем, что в
старшем школьном возрасте среди лиц с несвойственной для них регуляцией
преобладают лица с симпатикотонией, а в младшем школьном возрасте - с
ваготонией.
Поскольку изменения регуляции СР имеют общую
динамику для лиц с одинаковым тонусом ВНС не зависимо от их возраста, то,
следовательно, если учитывать исходный тонус ВНС при анализе реагирования
организма на двигательную деятельность, нет необходимости для выделения
возрастных групп. Поэтому для анализа изменений ФС организма у школьников в
каждой из групп с разной двигательной активностью было выделено три подгруппы
лиц с разным исходным тонусом ВНС - эйтоники, симпатотоники и ваготоники.
В группе 1 (с меньшей нагрузкой) выяснилось, что
у лиц с эйтонией отсутствовали достоверные изменения ФС (рис. 1). При этом у
39% лиц с эйтонией оно характеризовалось удовлетворительной адаптацией, у 33% -
напряжением механизмов адаптации и у 28% - неудовлетворительной адаптацией.

Рис. 1. Соотношение лиц с разным
функциональным состоянием в группе 1 (с меньшей нагрузкой) в начале и в конце
исследования
Можно предположить, что мышечная
нагрузка в этой группе не оказала влияние на лиц с эйтонией вследствие своей
незначительности. Однако следует отметить, что по литературным данным (Искакова
З.Б., 1991; Антропова М.В. и др., 1997), к концу учебного года у школьников
развивается напряжение систем регуляции, а поскольку завершение наших
исследований произошло в середине второй половины учебного года, то можно
говорить о нивелировании данного напряжения за счет двигательной активности.
Это свидетельствовало о стабилизирующем влиянии двигательной активности на
характеристики вегетативной регуляции.
У большинства лиц с симпатикотонией
(73%) ФС организма достоверно улучшилось и стало характеризоваться
удовлетворительной адаптацией. Тоже наблюдалось у 50% лиц с ваготонией. Однако
у 30% лиц с ваготонией сохранилось ФС, характеризовавшееся напряжением
механизмов адаптации, и у 20% - неудовлетворительной адаптацией.
Проведенный анализ показал, что в
группе 1 (с меньшей нагрузкой) значительно изменилось по сравнению с началом
исследования соотношение лиц с разным ФС. Существенно увеличилась доля лиц с
удовлетворительной адаптацией, и значительно сократилось число лиц с
напряжением механизмов адаптации и неудовлетворительной адаптацией. Наблюдаемая
динамика ФС в группе с низкой мышечной нагрузкой, по-видимому, была связана не
с тренировочным эффектом, а с развитием в организме благоприятных
неспецифических адаптационных реакций. Это согласуется с исследованиями ряда
авторов (Гаркави Л.Х., Квакина Е.Б, Уколова М.А., 1990; Ульянов В.И., 1995; Fleshner M., 1999).
В результате особенностями ФС
организма в группе 2 (с большей нагрузкой) выяснилось, что достоверные
изменения ФС произошли только у лиц с эйтонией (рис. 2). Количество эйтоников с
удовлетворительной адаптацией увеличилось с 30% до 70%. Полностью исчезли лица,
характеризующиеся неудовлетворительной адаптацией.
Рис. 2. Соотношение лиц с разным функциональным
состоянием в группе 2 (с большей нагрузкой) в начале и в конце исследования
Среди лиц с симпатикотонией и ваготонией
достоверных изменений ФС не произошло. При этом у большинства лиц (74%) с
симпатикотонией сохранилось ФС, характеризующееся напряжением механизмов
адаптации. Выборка лиц с ваготонией состояли из трех, близких по размерам,
частей: лица с удовлетворительной адаптацией - 31%, с напряжением механизмов
адаптации - 29%, с неудовлетворительной адаптацией - 40%.
Отсутствие улучшения ФС у лиц с ваготонией и
симпатикотонией в группе 2 (с большей нагрузкой) указывало на то, что для них
требуется более тщательное планирование двигательной активности в зависимости
от ФС организма.
Таким образом, это свидетельствует о том, что
формирование адаптивных реакций существенно зависело от индивидуальных
особенностей вегетативной регуляции и объема мышечной нагрузки. Так в группе с
меньшими нагрузками формирование адаптивных реакций в меньшей степени зависело
от характера дифференцированности типа вегетативной регуляции. В то же время в
группе с большей нагрузкой удовлетворительная адаптация формировалась только у
лиц с достаточно пластичной вегетативной регуляцией, а у лиц с жестко определенным
типом регуляции адаптивные изменения наблюдались в значительно меньшей степени.
Полученные результаты развивают представление о
формировании механизмов вегетативной регуляции сердечного ритма в онтогенезе и
могут быть использованы для оценки адекватности различных видов воздействия
индивидуальным адаптационным возможностям организма.
Нарушения сердечного ритма
Нарушения сердечного ритма - это очень сложный
раздел кардиологии. Сердце человека работает всю жизнь. Оно сокращается и
расслабляется от 50 до 150 раз в минуту. В фазу систолы сердце сокращается,
обеспечивая ток крови и доставку кислорода и питательных веществ по всему
организму. В фазу диастолы оно отдыхает. Поэтому очень важно, чтобы сердце
сокращалось через одинаковые промежутки времени. Если укорачивается период
систолы, сердце не успевает полноценно обеспечить организм движением крови и
кислородом. Если сокращается период диастолы - сердце не успевает отдохнуть.
Нарушение сердечного ритма - это нарушение частоты, ритмичности и последовательности
сокращений сердечной мышцы. Сердечная мышца - миокард состоит из мышечных
волокон. Различают два вида этих волокон: рабочий миокард или сократительный,
обеспечивающий сокращение проводящий миокард создающий импульс к сокращению
рабочего миокарда и обеспечивающий проведение этого импульса. Сокращения
сердечной мышцы обеспечиваются электрическими импульсами, возникающими в
синоаурикулярном или синусовом узле, который находится в правом предсердии.
Затем электрические импульсы распространяются по проводящим волокнам предсердий
к атриовентрикулярному узлу, расположенному в нижней части правого предсердия.
Из атриовентрикулярного узла начинается пучок Гиса. Он идет в межжелудочковой
перегородке и делится на две ветви - правую и левую ножки пучка Гиса. Ножки
пучка Гиса в свою очередь делятся на мелкие волокна - волокна Пуркинье по
которым электрический импульс достигает мышечных волокон. Мышечные волокна
сокращаются под действием электрического импульса в систолу и расслабляются при
его отсутствии в диастолу. Частота нормального (синусового) ритма сокращения
около от 50 сокращений во время сна, в покое, до 150-160 при физической и
психоэмоциональной нагрузке, при воздействии высоких температур.
Регулирующее влияние на активность
синусового узла оказывают эндокринная система, посредством содержащихся в крови
гормонов и вегетативная нервная система - ее симпатический и парасимпатический
отделы. Электрический импульс в синусовом узле возникает благодаря разнице
концентраций электролитов внутри и вне клетки и их перемещению через клеточную
мембрану. Основные участники этого процесса - калий, кальций, хлор и в меньшей
степени натрий. Причины нарушений сердечного ритма изучены не полностью.
Считается, что основными двумя причинами служат изменения нервной и эндокринной
регуляции или функциональные нарушения, и аномалии развития сердца, его
анатомической структуры - органические нарушения. Часто это бывают комбинации
этих основных причин. Увеличение частоты сердечных сокращений более 100 в
минуту называется синусовой тахикардией. Сокращения мышцы сердца при этом
полноценные и сердечные комплексы на электрокардиограмме не изменяются, просто
регистрируется учащенный ритм. Это может быть реакция здорового человека на
стресс или физическую нагрузку, но может быть и симптомом сердечной
недостаточности <#"564309.files/image003.gif">
Диаграмма 1. Амплитуда моды (АМо)

Диаграмма 2. Индекс напряжения регуляторных
систем (ИНрс)
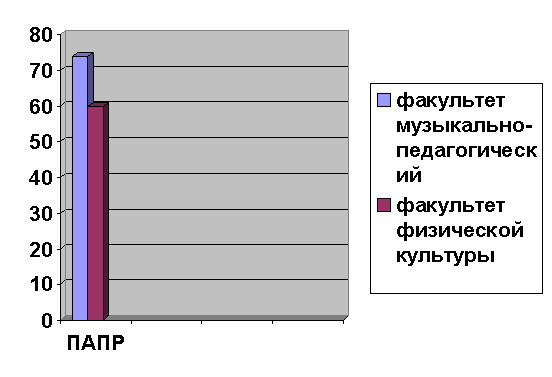
Диаграмма 3. Показатель адекватности
процессов регуляции (ПАПР)
Диаграмма 4. Индекс вегетативного равновесия
(ИВР)
*р < 0,001 - статистический достоверный сдвиг
по критерию Стьюдента
Анализируя полученные результаты мы пришли к
следующим выводам:
. Амплитуда моды у студентов
музыкально-педагогического факультета достоверно превышает на 4% факультет
физического воспитания, доказывает, что у данных студентов частота сердечных
сокращений выше нормы. (74 норма)
. Индекс напряжения регуляторных систем у
студентов музыкально-педагогического факультета превышает на 30% факультет
физического воспитания, что отражает повышенную степень централизации
управления ритмом сердца за счет симпатической нервной системы.
. Показатель адекватности процессов
регуляции у студентов музыкально-педагогического факультета превышает на 14
пунктов, что отражает превалирование активной регуляции симпатической нервной
системы над парасимпатической нервной системой синусного узла.
. Индекс вегетативного равновесия у
студентов музыкально-педагогического факультета превышает на 24 пункта по
сравнению с факультетом физического воспитания, что отражает повышение
активности симпатической нервной системы над парасимпатической вегетативной
нервной системой.
Таким образом, типологический подход позволил
выявить некоторые негативные особенности регуляции сердечного ритма студентов
музыкально-педагогического факультета. Студенты с симпатикотоническим типом
вегетативных регуляций музыкально-педагогического факультета характеризуются
выраженным преобладанием симпато-адреналового контура регуляции ритма сердца
над ваготоническим. По-видимому, это свидетельствует о значительном напряжении
механизмов адаптации сердечнососудистой системы обследованных к учебным
нагрузкам. Эффективность регуляции сердечного ритма у студентов факультета
физической культуры выше.
Заключение
Начальный период обучения в институте является
очень ответственным как в социальном, так и в физическом отношении моментом в
жизни студента. Новые условия обучения, высокая суммарная учебная нагрузка,
большой объём, новизна и сложность материала, которым должен овладеть студент,
предъявляют к организму повышенные требования. Существенная система обучения
изначально подавляет физиологические возможности организма, воздействуя на него
в прессинговом режиме однонаправленного характера, не учитывающем
морфофункциональных особенностей и индивидуальных темпов биологического
созревания. Это приводит к напряжению и срыву механизмов адаптации, существенно
снижает уровень здоровья интеллектуальный и физический потенциал студентов.
Ведущую роль в обеспечении адаптационной деятельности организма играет система
кровообращения.
Эта роль определяется, прежде всего, её функций
транспорта питательных веществ и кислорода - основных источников энергии для
клеток и тканей. Таким образом, изменение параметров сердечнососудистой системы
может расширять или же наоборот, лимитировать адаптационные возможности
организма, так как энергетический механизм занимает главное место в процессах
адаптации. И поэтому цель работы это использование показателей
сердечного ритма для оценки функционального состояния студентов. Для этого мы
поставили перед собой две задачи. Первая - выявить особенности показателей
сердечного ритма, влияющие на функциональное состояние студентов, и вторая -
изучить зависимость показателей сердечного ритма от функционального состояния
учащихся. Чтобы добиться результата мы использовали метод вариационной
пульсометрии по Р.М. Баевскому. В работе применялся автоматизированный способ
регистрации кардиоинтервалов с помощью электронного кардиографа посредством
связи блютус (Поли-Спектр-8/EХ
фирмы Нейрософт) и кардиотренажер (KETTLER).
Алгоритм математической обработки массива включает в себя вычисление моды (Мо),
амплитуды моды (АМо), индекс вегетативного равновесия (ИВР), вегетативный
показатель ритма (ВПР), показатель адекватности процессов регуляции (ПАПР),
индекс напряжения регуляторных систем (ИНрс). Методами математической
статистики определяем среднее арифметическое (М), среднее квадратическое
отклонение (r), ошибку выборки (m).
Проверку гипотезы достоверности различий параметров ритма сердца осуществляем
по t - критерии
Стьюдента.
С помощью этой методики мы пришли к следующим
результатам: типологический подход позволил выявить некоторые негативные
особенности регуляции сердечного ритма студентов КГУ им. Н.А. Некрасова.
Студенты с симпатикотоническим типом вегетативных регуляций
музыкально-педагогического факультета характеризуются выраженным преобладанием
симпато-адреналового контура регуляции ритма сердца над ваготоническим.
По-видимому, это свидетельствует о значительном напряжении механизмов адаптации
сердечнососудистой системы обследованных к учебным нагрузкам. Эффективность
регуляции сердечного ритма у студентов факультета физической культуры выше, что
косвенно доказывает положительное влияние физической нагрузки на регуляцию
сердечного ритма и функционирование сердечнососудистой системы в целом.
Список литературы
1.
Баевский, Р.М., Оценка адаптационных возможностей организма и риск развития
заболеваний / Р.М. Баевский, А.П. Берсенева. - М.: Медицина. 1997. - 265 с.
.
Вариабельность сердечного ритма: Теоретические аспекты и практическое
применение // Тез. докл. IV всерос. симп. / Отв. ред. Н.И. Шлык., Р.М.
Баевский. - УдГУ. Ижевск, 2008. - 344 c.
.
Вегетативные расстройства: Клиника, диагностика, лечение / Под.ред. А.М. Вейна.
- М.: ООО «Медицинское информационное агенство», 2003. - 752 с.
.
Влияние оздоровительного бега на состояние здоровья школьников // Мат. XVII
съезда физиологов России: тез. науч. сообщ. Ростов-на-Дону, 1998, с.345-346 (соавт.
Игишева Л.Н.)
.
Возрастные особенности регуляции сердечного ритма у детей и подростков //
Тезисы докладов 2-го съезда физиологов Сибири и Дальнего Востока. Новосибирск,
1995. С.227-228 (соавт. Игишева Л.Н., Ботин С.В.)
.
Граевская, Н.Д. Спортивная медицина: Курс лекций и практические занятия.
Учебное пособие / Н.Д. Граевская, Т.И. Долматова - М.: Советский спорт, 2004. -
304 с.
.
Двигательная деятельность - необходимый компонент валеологического образования
// в кн. Проблемы валеологизации образовательной среды. Кемерово, 1999, с.
44-90 (соавт. Казин Э.М., Игишева Л.Н.)
.
Зависимость вида адаптационной реакции, вызываемой женьшенем, от его дозировки
// в сб. Медико-биологические проблемы, №4, Кемерово, 1998 (соавт. Куликова
С.А., Баранова А.В.)
.
Зависимость состояния эритрона от типа адаптационной реакции // в сб.
Медико-биологические проблемы, №5, Кемерово, 1998 (соавт. Минин В.В., Драпезо
Р.Г.)
.
Использование автоматизированной кардиоритмографической программы
"ОРТОПЛЮС" для донозологических обследований // В сб.:Валеологические
аспекты образования: тез. докл. межрегиональной науч.-практ. конф. Барнаул,
1996. С.31-32 (соавт. Игишева Л.Н., Тарасова О.Л.)
.
Использование валеологических подходов к проблеме физического воспитания в
образовательных учреждениях // Валеология, № 4, 1998 (соавт. Казин Э.М.,
Петухов С.И., Игишева Л.Н.)
.
Использование ритмографии для оценки адаптационных возможностей организма
новорожденных с асфиксией в родах // Материалы к докладам научно-клинической
конференции посвященной 30-летию педиатрического факультета КГМА. Кемерово,
1998. (соавт. Цой К.Г., Царева В.А.)
.
Макаров, Л.М. Холтеровское мониторирование / Л.М. Макаров. - М.: Медпрактика-М.
- 2008. - 456 с.
.
Михайлов, В.М. Вариабельность ритма сердца: опыт практического применения
метода / В.М. Михайлов. - Иваново, 2002. - 290 с.
.
Оздоровление учащихся с использованием модели валеологического режима
двигательной активности // сб. Валеологические аспекты образования: тез. докл.
межрегиональной науч.-практ. конф. Барнаул, 1998 (соавт. Игишева Л.Н., Иванов
В.И.)
.
Оржоникидзе, З.Г. Физиология футбола / З.Г. Оржоникидзе, В. И. Павлов. - М.:
Человек, 2008. - 240 с.
.
Особенности регуляции сердечного ритма у подростков с повышенным артериальным
давлением. // Педиатрия №6, 1995, с. 17-21 (соавт. Игишева Л.Н., Ботин С.В.)
.
Платонов, В.Н. Система подготовки спортсменов в олимпийском спорте. Общая
теория и ее практические приложения / В.Н. Платонов. - К.: Олимпийская
литература, 2004. - 808 с.
.
Поиск взаимосвязей между параметрами кинетики кислотного гемолиза эритроцитов и
функциональным состоянием организма //Физиология человека, Т.22, № 4, 1996
(соавт. Казин Э.М., Голенда И.Л.)
.
Приходько В.И. Особенности функционального состояния сердечно-сосудистой
системы юных пловцов, достигших высоких спортивных результатов / В.И.
Приходько, Л.М. Беляева // Теория и практика физ. культуры. - 1996. - № 9. - С.
2-5.
.
Рябыкина, Г.В. Вариабельность ритма сердца / Г.В.Рябыкина А.В. Соболев.- М.:
Оверлей, 2001. - 200 с.
.
Физиология человека: Учебник / Под ред. В.М.Смирнова. - М.: Медицина, 2002. -
608 с.: ил. (Учеб. лит. Для студентов мед. вузов).