Выведение нового гибрида помидор
Содержание
1 Хромосомная теория
наследственности
Томаты как культура, её значение
Рабочая гипотеза
Доказательство рабочей гипотезы
Список используемой литературы
1 Хромосомная теория наследственности
· Сцепление. Группы сцепления.
Доказательство Т.Г. Морганом сцепления.
Т.Г. Морган родился в 1866 году, то есть ровно
через год после опубликования менделеевских закономерностей. Это весьма
примечательно, поскольку именно Моргану суждено было развить менделизм.
Человеку, прожившему 77 лет, а умер он в 1943 году, удалось сделать то, чего не
мог сделать сам Мендель и все его последователи. Это был удивительно
талантливый ученый-исследователь, генетик с мировым именем. Достаточно сказать,
что в 24 года он получает степень доктора философии и вскоре становится экстраординарным
(заштатным) профессором университета.
Вскоре Т. Морган уезжает из Кентукки в Италию,
но затем снова возвращается в Колумбийский университет и работает там, в
лаборатории Вильсона. Именно в этой лаборатории некоторыми учеными были
сформулированы основные положения хромосомной теории пола, но главные открытия
были впереди, поскольку научная атмосфера лаборатории способствовала
плодотворной работе.
Т.Г. Морган начал исследования в области
хромосомной теории наследственности в 1910 году. Для своих исследований ему был
необходим объект, но горох Менделя не устраивал, поскольку при работе с ним
приходится долго ждать результатов исследований, тогда как нетерпеливому
Моргану требовались не годы, а дни, десятки дней, и не более. Таким требованиям
отвечал единственный объект муха-дроздофила. Цикл развития этого двукрылого
насекомого от яйца до имаго составляет всего 10-12 дней, то есть 30 поколений в
год, что и нужно было Моргану. Кроме того, мушки хорошо размножаются, имеют
много мутаций. Это дает возможность получать большое количество рекомбинантных
форм. Работать с мухой весьма удобно, поскольку достаточно поднести ватку,
смоченную серным эфиром к открытой пробирке, как все ее наследие, а это 200-300
особей, полученных в результате скрещивания, «засыпает», с помощью шпателя,
разделяя их на рекомбинанты и нерекомбинанты.
В 1900 году, вторично были открыты законы
Менделя, и закон независимого комбинирования генов восторжествовал, однако у
некоторых исследователей в результате проводимых экспериментов, появились
сомнения в правильности этого закона. Это были В. Бэтсон и Р. Пеннет. Они
скрещивали две расы душистого горошка, различающиеся по окраске цветков и форме
пыльцы. Одна из них имела пурпурные цветки и пыльцу удлиненной формы, а другая-
красные цветки и пыльцу округлой формы. Пурпурная окраска Р доминировала над
красной р, а удлиненная форма пыльцы L
над округлой L.
Во втором поколении ожидаемого расщепления
9:3:3:1, получаемого обычно при независимом комбинировании генов, не
получалось. Ученые несколько изменили методику, но пришли к тому же результату.
Во втором поколении, особей, похожих на
родителей, было больше теоретически ожидаемого количества. Оба исследователя
для объяснения этого предложили гипотезу «притяжения-отталкивания»
наследственных факторов, которая потом оказалась несостоятельной. До 1910 года
такое несоответствие считали исключением из правил, но когда накапливается
много таких исключений из правил, то возникает вполне законное сомнение в
правильности теории, а Т.Г. Морган, работая с мушкой-дрозофилой, обнаружил
много таких фактов.
Морган скрещивал муху дикого типа, имеющую серое
тело и длинные крылья, с мухой мутантной, имеющей черное тело и зачаточные
крылья, в первом поколении получал фенотип мух с серым телом и длинными
крыльями, но когда исследователь прибегнул к анализирующему скрещиванию, то
одинакового количества гибридов не получил, как это предусматривало независимое
комбинирование генов. На основании экспериментов Морган пришел к выводу, что
причиной такого явления следует считать сцепление генов. Поскольку гены
находятся как бы в «связке», то и передаются в анафазе-1 вместе, а поэтому
других особей и не может быть. Вот эти скрещивания:
b+ vq+ bvq F1
b+vq+
Р
: x
b+ vq+ bvq
bvq
серое
тело черное тело серое тело
длинные крылья зачаточные крылья длинные крылья
гаметы
b+vq+ bvq Fa b+vq+
x
bvq
bvq
bvq
На основании этого Т. Морган сформулировал
первый постулат хромосомной теории наследственности, который звучал так: «Гены
расположены в хромосомах и образуют группы сцепления, количество которых равно
гаплоидному числу».
Генетическое доказательство кроссинговера.
Величина перекреста и линейное расположение генов.
Прошло некоторое время, и при повторении
эксперимента Морган вдруг обнаружил мух, не похожих на родительские формы, то
есть с черным телом, и длинными крыльями и с серым телом, но короткими
крыльями. Таких мух было 17%, и если это мутации, рассуждал Морган, то почему
их много, ведь они встречаются в количествах 1:10000 особей? Он объясняет такое
явление кроссинговером, то есть перекрестом хромосом и формулирует второй
постулат хромосомной теории наследственности: «Между генами в хромосомах
наблюдается обмен в результате кроссинговера, который идет тем чаще, чем больше
расстояние между генами».
Однако так и остается открытым вопрос, почему
рекомбинантных особей 17% и каждый раз при повторном скрещивании эта цифра
подтверждалась? При скрещивании мутантных мух по другим признакам он получал иное
отношение нерекомбинантов и рекомбинантам, но всегда результат был стабильным.
Могран высказывает догадку о том, что эта цифра зависит от расстояния между
генами. Это была правильная догадка, но ее следовало экспериментально
подтвердить.
Для экспериментального доказательства был удачно
применен известный дидактический метод трех точек, который в связи с
поставленной целью заключался в следующем. В лаборатории скрестили муху с
желтым телом J (Jellow)
с мухой, имеющей белые глаза w
(white) и получили 1,2 %
рекомбинантных особей. Затем муху с белыми глазами скрестили с мухой, имеющей
вильчатые щетинки bi
(bifild) и получили 3,5 %
рекомбинантов. Скрестили муху с желтым телом с мухой, имеющей вильчатые крылья,
и получили 4,7% рекомбинантов. Гипотеза подтвердилась экспериментально, и
Морган формулирует третий постулат хромосомной теории: «Расстояние между генами
равно проценту рекомбинантных особей».
Все эти доказательства Моргана свидетельствовали
о линейном расположении генов в хромосомах:
J w 4,7 bi.
1,2 3,5
Величину перекреста хромосом вычисляют в
процентах кроссинговерных особей к их общему числу в данном скрещивании.
Морганида - это генетическая длинна отрезка хроматиды, на которой в мейозе
происходит один обмен. Отсюда и более мелкая единица - сантиморган. Сантиморган
- это генетическое расстояние, на котором кроссинговер происходит с
вероятностью 1%. Таким образом, 1 сМ = 1%. В системе СИ 1 сМ = 1/100 морганиды.
В дальнейшем стали говорить более обобщенно, а
расстояние выражать в единицах кроссинговера (е.к),что по модулю равно проценту
рекомбинации.
· Цитологическое доказательство
кроссинговера. К. Штерн, Б. Мак-Клинток, Г. Крейтон.
После того, как было генетически доказано
явление кроссинговера, появилась необходимость доказать его цитологически. Это
было сделано в 30-х годах К. Штерном на дрозофиле, затем Г. Крейтон и
Мак-Клинток осуществили это на кукурузе.
К. Штерну удалось получить у самки и
самца половые хромосомы, отличные друг от друга цитологически и генетически. К одной
из Х-хромосом самки методом транслокации был присоединен бесцентромерный
фрагмент от У-хромосомы, что дало Г-образную Х-хромосому с генами сr
+ B + дикого типа.
Другая Х-хромосома была составной: один фрагмент с центромерой содержал
рецессивный ген В (ген сr
определяет окраску глаз цвета красной гвоздики - carnation,
а доминантный ген В - полосковидные глаза - Bar).
Бесцентромерный фрагмент Х- хромосомы прикрепился к 4 - аутосоме, что не
изменило хромосомного баланса зиготы, и самка была жизнеспособной. Поскольку
она гетерозиготна, то у нее в гаметогенезе мог происходить кроссинговер,
который учитывался как цитологически, так и генетически.
Для анализирующего скрещивания брали
самцов с нормальными Х и У хромосомами. Половая хромосома самца несла гены cr
и В + в гомозиготном состоянии. В потомстве были проанализированы только самки,
среди которых появилось два класса мух с некроссоверными хромосомами:
сr B
и cr+B+
сr B+
cr B+
Эти два последних класса мух могли
появиться только в результате кроссинговера между генами cr
и В.
Для доказательства на кукурузе были
использованы две линии, различающиеся по окраске и консистенции эндосперма.
Гены контролирующие эти признаки, локализованы в 9 хромосоме.
У одной из линии кукурузы эта пара
хромосом была помечена, то есть одна хромосома несла рецессивный ген (с)
неокрашенного и доминантный ген (wx+)
крахмального эндосперма, другая содержала доминантный ген окрашенного (c+)
и рецессивный ген восковидного эндосперма (wx).
Последняя хромосома имела булавовидную форму. Эта линия скрещивалась с линией,
имеющей морфологически нормальные хромосомы с рецессивными генами неокрашенного
и восковидного эндосперма в гомозиготном состоянии. В результате кроссинговера
появились формы:
С неокрашенным
восковидным: 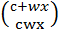
И окрашенным крахмалистым
эндоспермом: 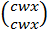 .
.
· Использование основных постулатов хромосомной
теории наследственности в селекции. Генетическое картирование. Дрозофильный
практикум.
Важнейший генетический объект - D.
Melanogaster является наиболее
подробно изученным организмом. На карте этого вида нанесено более 1000 генов.
При этом отмечается группировка генов в пределах ограниченных участков генома.
Над этим объектом не довлела необходимость решения глобальных проблем
практической селекции, а поэтому все генетики мира считали своим долгом
привлечь его для решения фундаментальных теоретических проблем. Эти
теоретические упражнения ученых и легли в основу составления карт, практическое
значение которых в селекции трудно переоценить. Что же дает знание карт? Взять
хотя бы томаты. Разработка частной селекции этого ценного овощного растения диктовалась
практикой и опирается на теоретические закономерности, открытие на дрозофиле.
Первые работы по картированию томатов были
известны еще в 1917 году, а к 1973 году было картировано 263 гена. Знание
локализации генов позволило сознательно планировать гибридизацию и в данном
случае оказывается полезной информация не только о локализации их в конкретной
хромосоме при независимом комбинировании, но и при сцепленном. Стало понятным,
что если сцепление не очень тесное, то при отборе требуется увеличение объема
выборки гибридов, чтобы достичь желаемых результатов по рекомбинации. И,
наоборот, при очень тесном сцеплении для выявления полезных рекомбинантов
требуется применение специальных рекомбинирующих факторов, особенно когда это
сцепление не желательно.
Рассмотрим конкретный пример. У томата в 9
хромосоме (22 локус) тесно сцеплены рецессивный ген nv
- преждевременного пожелтения листьев (нежелательный признак) и доминантный ген
Tm - 2 - устоичивости
к вирусу табачной мозаики (ВТМ). Они сцеплены настолько тесно, что на карте
показаны в одном и том же 22 локусе. Без применения рекомбинантных веществ
путем одной только селекции разделить эти гены не удается.
Однако следует с удовлетворением отметить, что в
других случаях оказываются сцепленными сразу несколько генов, контролирующих
ценные признаки. В данном случае это целый блок благоприятных генов и здесь
желательно избегать рекомбинаций, хотя при случае и это возможно. В таком
полезном для селекции блоке удачно сочетаются ценные признаки устоичивости к
кладоспориозу, корневой нематоде, детерминантный габитус куста, высокое
содержание каротина с темно-красной окраской плодов, характеризующей высокие
вкусовые качества плода.
В представленном блоке независимое
комбинирование возможно между генами Gf,
Mi (32, 35 локусы) и
В, sp (94 локус, 6-й
хромосомы). В таком случае возможна потеря генов В и sp,
что весьма нежелательно. Чтобы этого не происходило, вводят в генотип мутацию hp,
повышающую в плодах содержание В - каротина и ценность плода восстанавливается.
Для того, чтобы при гибридологическом анализе
можно было как то проследить за передачей нужных генов, используют тот же
эффект сцепления, но с генами выполняющими роль маркеров. Так, у томатов во 2 и
10 хромосомах локализованы гены, контролирующие форму плода, а во 2 хромосоме
имеется еще и ген скороспелости. По сути дела, гены скороспелости
сконцентрированы в двух блоках в тесном сцеплении с такими маркерами как m
(пятнистые листья), d
(укороченность всех частей растения), о (грушевидные плоды), s
(увеличение числа цветков). Все эти гены рецессивны и их используют в качестве
меток (маркеров) в селекции на скороспелость. Здесь открываются неисчерпаемые
возможности полезной рекомбинации с увеличенной или уменьшенной долей ценных
признаков. Отбор линий с увеличенной рекомбинацией достигается в результате
кроссинговера. Открытие половых хромосом и установление их роли в определении
пола послужило важным доводом в пользу того, что хромосомы определяют признаки
организма.
Определение пола
От чего же зависит рождение мужских и женских
особей? Рассмотрим это на примере определения пола у дрозофилы. В ходе мейоза у
самок образуется один тип гамет, содержащий гаплоидный набор аутосом и одну X-хромосому.
Самцы образуют два типа гамет, половина из которых содержит три аутосомы и одну
X-хромосому (ЗА+Х),
а половина - три аутосомы и одну У-хромосому (ЗА+У). При оплодотворении
яйцеклеток (ЗА+Х) сперматозоидами с X-хромосомами
будут формироваться самки (6А+ХХ), а от слияния яйцеклеток со сперматозоидами,
несущими У-хромосому, - самцы (6A+XY).
Поскольку число мужских гамет с X-
и У-хромосомами одинаково, то и количество самцов и самок тоже одинаково. В
данном случае пол организма определяется в момент оплодотворения и зависит от
хромосомного набора зиготы.
Сходный способ определения пола (XY-тип)
присущ всем млекопитающим, в том числе и человеку, клетки которого содержат 44
аутосомы и две X-хромосомы у
женщин либо XY-хромосомы у
мужчин.
Таким образом, XY-тип
определения пола, или тип дрозофилы и человека, - самый распространенный способ
определения пола, характерный для большинства позвоночных и некоторых
беспозвоночных. Х0-тип встречается у большинства прямокрылых, клопов, жуков,
пауков, у которых Y-хромосомы
нет вовсе, так что самец имеет генотип Х0, а самка - XX.
У всех птиц, большинства бабочек и некоторых
пресмыкающихся самцы являются гомогаметным полом, а самки -- гетерогаметным
(типа XY или типа
ХО). Половые хромосомы у этих видов обозначают буквами Z
и W, чтобы выделить
таким образом данный способ определения пола; при этом набор хромосом самцов
обозначают символом ZZ,
а самки - символом ZW
или Z0.
Доказательства того, что половые хромосомы
определяют пол организма, были получены при изучении нерасхождения половых
хромосом у дрозофилы. Если в одну из гамет попадут обе половые хромосом, а в
другую - ни одной, то при слиянии таких гамет с нормальными могут получиться
особи с набором половых хромосом ХХХ, ХО, ХХУ и др. Выяснилось, что у дрозофилы
особи с набором ХО - самцы, а с набором ХХУ - самки (у человека - наоборот).
Особи с набором ХХХ имеют гипертрофированные признаки женского пола
(сверхсамки). (Особи со всеми этими хромосомными аберрациями у дрозофилы
стерильны). В дальнейшем было доказано, что у дрозофилы пол определяется
соотношением (балансом) между числом X-хромосом
и числом наборов аутосом.
Наследование признаков, сцепленных с полом.
В том случае, когда гены, контролирующие
формирование того или иного признака, локализованы в аутосомах, наследование
осуществляется независимо от того, кто из родителей (мать или отец) является
носителем изучаемого признака. Если же гены находятся в половых хромосомах,
характер наследования признаков резко изменяется. Например, у дрозофилы гены,
локализованные в X-хромосоме,
как правило, не имеют аллелей в У-хромосоме. По этой причине рецессивные гены в
X-хромосоме
гетерогаметного пола практически всегда проявляются, будучи в единственном
числе.
Признаки, гены которых локализованы в половых
хромосомах, называются признаками, сцепленными с полом. Явление наследования,
сцепленного с полом, было открыто Т. Морганом у дрозофилы.
Х- и У-хромосомы у человека имеют гомологичный
(псевдоаутосомный) участок, где локализованы гены, наследование которых не
отличается от наследования аутосомных генов.
Помимо гомологичных участков, X-
и У-хромосомы имеют негомологичные участки. Негомологичный участок У-хромосомы,
кроме генов, определяющих мужской пол, содержит гены перепонок между пальцами
ног и волосатых ушей у человека. Патологические признаки, сцепленные с
негомологичным участком У-хромосомы, передаются всем сыновьям, поскольку они
получают от отца У-хромосому.
Негомологичный участок X-хромосомы
содержит в своем составе ряд важных для жизнедеятельности организмов генов.
Поскольку у гетерогаметного пола (ХУ) X-хромосома
представлена в единственном числе, то признаки, определяемые генами
негомологичного участка X-хромосомы,
будут проявляться даже в том случае, если они рецессивны. Такое состояние генов
называется гемизиготным. Примером такого рода X-сцепленных
рецессивных признаков у человека являются гемофилия, мышечная дистрофия Дюшена,
атрофия зрительного нерва, дальтонизм (цветовая слепота) и др.
Гемофилия - это наследственная болезнь, при
которой кровь теряет способность свертываться. Ранение, даже царапина или ушиб,
могут вызвать обильные наружные или внутренние кровотечения, которые нередко
заканчиваются смертью. Это заболевание встречается, за редким исключением,
только у мужчин. Было установлено, что обе наиболее распространенные формы
гемофилии (гемофилия А и гемофилия В) обусловлена рецессивными генами,
локализованными в X-хромосоме.
Гетерозиготные по данным генам женщины (носительницы) обладают нормальной или
несколько пониженной свертываемостью крови.
Фенотипическое проявление гемофилии у девочек
будет наблюдаться в том случае, если мать девочки является носительницей гена
гемофилии, а отец - гемофиликом. Подобная закономерность наследования
характерна и для других рецессивных, сцепленных с полом признаков.
Сцепленное наследование.
Независимое комбинирование признаков (третий
закон Менделя) осуществляется при условии, что гены, определяющие эти признаки,
находятся в разных парах гомологичных хромосом. Следовательно, у каждого
организма число генов, способных независимо комбинироваться в мейозе,
ограничено числом хромосом. Однако в организме число генов значительно
превышает количество хромосом. Например, у кукурузы до эры молекулярной
биологии было изучено более 500 генов, у мухи дрозофилы - более 1 тыс., а у
человека - около 2 тыс. генов, тогда как хромосом у них 10, 4 и 23 пары
соответственно. То, что число генов у высших организмов составляет несколько
тысяч, было ясно уже У. Сэттону в начале XX
века. Это дало основание предположить, что в каждой хромосоме локализовано
множество генов. Гены, локализованные в одной хромосоме, образуют группу
сцепления и наследуются вместе.
Способ наследования сцепленных генов отличается
от наследования генов, локализованных в разных парах гомологичных хромосом.
Так, если при независимом комбинировании дигетерозиготная особь образует четыре
типа гамет (АВ, Ab,
аВ и ab) в равных
количествах, то при сцепленном наследовании (в отсутствие кроссинговера) такая
же дигетерозигота образует только два типа гамет: (АВ и ab)
тоже в равных количествах. Последние повторяют комбинацию генов в хромосоме
родителя.
Было установлено, однако, что кроме обычных
(некроссоверных) гамет возникают и другие (кроссоверные) гаметы с новыми
комбинациями генов - Ab
и аВ, отличающимися от комбинаций генов в хромосомах родителя. Причиной
возникновения таких гамет является обмен участками гомологичных хромосом, или
кроссинговер.
Кроссинговер происходит в профазе I
мейоза во время конъюгации гомологичных хромосом. В это время части двух
хромосом могут перекрещиваться и обмениваться своими участками. В результате
возникают качественно новые хромосомы, содержащие участки (гены) как
материнских, так и отцовских хромосом. Особи, которые получаются из таких гамет
с новым сочетанием аллелей, получили название кроссинговерных или
рекомбинантных.
Частота (процент) перекреста между двумя генами,
расположенными в одной хромосоме, пропорциональна расстоянию между ними.
Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они
расположены. По мере увеличения расстояния между генами все более возрастает
вероятность того, что кроссинговер разведет их по двум разным гомологичным
хромосомам.
Расстояние между генами характеризует силу их
сцепления. Имеются гены с высоким процентом сцепления и такие, где сцепление
почти не обнаруживается. Однако при сцепленном наследовании максимальная
частота кроссинговера не превышает 50 %. Если же она выше, то наблюдается
свободное комбинирование между парами аллелей, не отличимое от независимого
наследования.
Биологическое значение кроссинговера чрезвычайно
велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не
существовавшие комбинации генов и тем самым повышать наследственную
изменчивость, которая дает широкие возможности адаптации организма в различных
условиях среды. Человек специально проводит гибридизацию с целью получения необходимых
вариантов комбинаций для использования в селекционной работе.
Карты хромосом.
Т. Морган и его сотрудники К. Бриджес, А. Г.
Стертевант и Г. Дж. Меллер экспериментально показали, что знание явлений
сцепления и кроссинговера позволяет не только установить группу сцепления
генов, но и построить генетические карты хромосом, на которых указаны порядок
расположения генов в хромосоме и относительные расстояния между ними.
Генетической картой хромосом называют схему
взаимного расположения генов, находящихся в одной группе сцепления. Такие карты
составляются для каждой пары гомологичных хромосом.
Возможность подобного картирования основана на
постоянстве процента кроссинговера между определенными генами. Генетические
карты хромосом составлены для многих видов организмов: насекомых (дрозофила,
комар, таракан и др.), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Наличие генетической карты свидетельствует о
высокой степени изученности того или иного вида организма и представляет
большой научный интерес. Такой организм является прекрасным объектом для
проведения дальнейших экспериментальных работ, имеющих не только научное, но и
практическое значение. В частности, знание генетических карт позволяет
планировать работы по получению организмов с определенными сочетаниями
признаков, что теперь широко используется в селекционной практике. Так,
создание штаммов микроорганизмов, способных синтезировать необходимые для
фармакологии и сельского хозяйства белки, гормоны и другие сложные органические
вещества, возможно только на основе методов генной инженерии, которые, в свою
очередь, базируются на знании генетических карт соответствующих
микроорганизмов.
Генетические карты человека также могут
оказаться полезными в здравоохранении и медицине. Знания о локализации гена в определенной
хромосоме используются при диагностике ряда тяжелых наследственных заболеваний
человека. Уже теперь появилась возможность для генной терапии, то есть для
исправления структуры или функции генов.
Сравнение генетических карт разных видов живых организмов
способствует также пониманию эволюционного процесса.
Основные положения хромосомной теории
наследственности.
Анализ явлений сцепленного наследования,
кроссинговера, сравнение генетической и цитологической карт позволяют
сформулировать основные положения хромосомной теории наследственности:
Гены локализованы в хромосомах. При этом
различные хромосомы содержат неодинаковое число генов. Кроме того, набор генов
каждой из негомологичных хромосом уникален.
Аллельные гены занимают одинаковые локусы в гомологичных
хромосомах.
Гены расположены в хромосоме в линейной
последовательности.
Гены одной хромосомы образуют группу сцепления,
то есть наследуются преимущественно сцепленно (совместно), благодаря чему
происходит сцепленное наследование некоторых признаков. Число групп сцепления
равно гаплоидному числу хромосом данного вида (у гомогаметного пола) или больше
на 1 (у гетерогаметного пола).
Сцепление нарушается в результате кроссинговера,
частота которого прямо пропорциональна расстоянию между генами в хромосоме
(поэтому сила сцепления находится в обратной зависимости от расстояния между
генами).
Каждый биологический вид характеризуется
определенным набором хромосом - кариотипом.
2 Томаты как культура, её значение
История томата.
Название помидор происходит от итал. pomo
d'oro
- золотое яблоко. Настоящее название было у ацтеков - матль, французы
переделали его в фр. tomate
(томат).
Родина - Южная Америка, где до сих пор
встречаются дикие и полукультурные формы томата. В середине XVI
века томат попал в Испанию, Португалию, а затем в Италию, Францию и другие
европейские страны, Самый ранний рецепт блюда из томатов опубликован в
кулинарной книге в Неаполе в 1692 при этом автор ссылается на то, что это
рецепт родом из Испании. В XVIII
веке томат попадает в Россию, где вначале возделывался как декоративное
растение. Овощной продовольственной культурой растение было признано благодаря
русскому учёному-агроному А. Т. Болотову (1738-1833). Долгое время томаты
считались несъедобными и даже ядовитыми. Европейские садоводы разводили их как
экзотическое декоративное растение. В американские учебники по ботанике вошла
история, как подкупленный повар пытался отравить блюдом из помидоров Джорджа
Вашингтона. Будущий первый президент США, отведав приготовленное кушанье, пошёл
дальше заниматься делами, так и не узнав о коварном предательстве.
Томат сегодня - одна из самых популярных культур
благодаря своим ценным питательным и диетическим качествам, большому
разнообразию сортов, высокой отзывчивости на применяемые приёмы выращивания.
Его возделывают в открытом грунте, под плёночными укрытиями, в теплицах,
парниках, на балконах, лоджиях и даже в комнатах на подоконниках.
Состав плодов томатов
Зрелые плоды томата богаты сахара́ми
и витамином C, содержат белки,
крахмал, органические кислоты, клетчатку и пектиновые вещества, минеральные
вещества (кальций, натрий, магний, железо, хлор, фосфор, серу, кремний, йод), а
также каротиноиды каротин и ликопин (они определяет жёлто-оранжевый или красный
цвет плодов), витамины группы B,
никотиновую и фолиевую кислоты, витамин K. В
этом разделе не хватает ссылок на источники информации.
Информация должна быть проверяема, иначе она
может быть поставлена под сомнение и удалена.
Свежие томаты и томатный сок полезны
при сердечно-сосудистых заболеваниях из-за большого содержания в них железа и
калия, а также томаты полезны при гастритах с пониженной кислотностью, общем
упадке сил, ослаблении памяти, малокровии. Томатный сок снижает кровяное
давление, кроме того, повышенное содержание пектиновых веществ в томате
способствует снижению холестерина в крови. Благодаря высокому содержанию
биологически активных веществ томаты регулируют обменные процессы и
деятельность желудочно-кишечного тракта, усиливают работу почек и половых
желез. Применяют томат и как слабительное средство. Кашицу красных томатов
прикладывают к вздувшимся венам (прибинтовывают на ночь ежедневно или через
день в течение месяца). Консервированные томаты подвергаются молочно-кислому
брожению и присутствующая в них молочная кислота благоприятно сказывается на
микрофлоре кишечника. Однако, при консервировании томатов всегда используется
соль, поэтому соленые и маринованные помидоры не рекомендуется есть при
заболеваниях почек и сердечно-сосудистой системы, в том числе при гипертонии
(повышенном давлении).
Биологические особенности
Томат имеет сильноразвитую корневую систему
стержневого типа. Корни разветвленные, растут и формируются быстро. Уходят в
землю на большую глубину (при безрассадной культуре до 1 м и более),
распространяясь в диаметре на 1,5-2,5 м. При наличии влаги и питания
дополнительные корни могут образовываться на любой части стебля, поэтому томат
можно размножать не только семенами, но также черенками и боковыми побегами
(пасынками). Поставленные в воду, они через несколько суток образуют корни.
Стебель у томата прямостоячий или полегающий,
ветвящийся, высотой от 30 см до 2 м и более. Листья непарноперистые,
рассеченные на крупные доли, иногда картофельного типа. Цветки мелкие,
невзрачные, желтые различных оттенков, собраны в кисть. Томат - факультативный
самоопылитель: в одном цветке имеются мужские и женские органы.
Плоды - сочные многогнёздные ягоды различной
формы (от плоско-округлой до цилиндрической; могут быть мелкими (масса до 50
г), средними (51-100 г) и крупными (свыше 100 г, иногда до 800 г и более).
Окраска плодов от бледно-розовой до ярко-красной и малиновой, от белой,
светло-зелёной, светло-жёлтой до золотисто-жёлтой.
Семена мелкие, плоские, заострённые у основания,
светло- или тёмно-желтые, обычно опушённые, вследствие чего имеют серый
оттенок. Физиологически зрелыми становятся уже в зелёных, сформированных
плодах. Всхожесть сохраняют 6-8 лет.
При благоприятных температурных условиях и
наличии влаги семена прорастают через 3-4 суток. Первый настоящий лист
появляется обычно через 6-10 суток после всходов, последующие 3-4 листа - ещё
через 5-6 суток, в дальнейшем каждый новый лист образуется через 3-5 суток.
Начиная с молодого возраста в пазухах листьев отрастают боковые побеги
(пасынки). Продолжительность периода от всходов до цветения растения 50-70
суток, от цветения до созревания плода 45-60 суток.
По строению куста, толщине стебля и характеру
листьев различают 3 разновидности томатов: нештамбовый, штамбовый,
картофельный.
Сорта томатов.
Сорта томата характеризуют по различным
критериям:
По типу роста куста - детерминированные и
индетермированные.
По времени созревания - ранние, среднеспелые,
поздние.
По способу употребления - столовые, для
консервации, для производства сока и др.
Наиболее распространены сорта нештамбового
томата, имеющего тонкие стебли, полегающие под тяжестью плодов, и крупные,
слабогофрированные листья; кусты могут быть как карликовыми, так и
высокорослыми. Сорта штамбового томата достаточно многочисленны. Стебли у
растений толстые, листья среднего размера, с короткими черешками и сближенными
долями, сильногофрированные; пасынков образуется мало. Кусты компактные - от
карликовых до среднерослых. Выведены полуштамбовые сорта томата, занимающие
промежуточное положение между указанными группами. Сортов картофельного типа,
получившего название за сходство его листьев с картофельными, очень мало.
По типу роста куста сорта томата делятся на
детерминированные (слаборослые) и индетермированные (высокорослые). У
детерминированных сортов основной стебель и боковые побеги прекращают рост
после образования на стебле 2-6, иногда более кистей. Стебель и все побеги
заканчиваются цветочной кистью. Пасынки образуются только в нижней части
стебля. Куст небольшой или средних размеров (60-180 см). Кроме типично
детерминированных выделяют также супердетерминированные сорта, у которых
растения прекращают рост после формирования на основном стебле 2-3 кистей (все
побеги оканчиваются соцветиями и образуют сильноразветвлённый небольшой куст;
вторая волна роста отмечается после созревания большей части плодов; первое
соцветие образуется на высоте 7-8-го листа), а также полудетерминированные,
растения которых отличаются более сильным, почти неограниченным ростом -
формируют на одном стебле 8-10 кистей. У индетермированных сортов томатов рост
растений неограничен. Основной стебель заканчивается цветочной кистю (первая
кисть образуется над 9-12 листом), а пасынок, растущий из пазухи листа,
ближайшего к верхушечной кисти, продолжает рост основного стебля. После
образования нескольких листьев пасынок заканчивает свой рост заложением
цветочного бутона, а рост растения продолжается за счёт ближайшего пасынка. Так
происходит до конца вегетации, которая обычно завершается первым осенним
заморозком. Куст высокорослый (2 м и более), но темп цветения и
плодообразования ниже, чем у томатов детерминированных сортов, растянутый.
В России, среди не специалистов, широко известны
такие сорта помидоров, как: «бычье сердце», «дамские пальчики», другие. В
последние десятилетие получили распространение томаты «Черри».
Томат - теплотребовательная культура,
оптимальная температура для роста и развития растений 22-25 °C:
при температуре ниже 10 °C
пыльца в цветках не созревает и неоплодотворенная завязь отпадает. Томат плохо
переносит повышенную влажность воздуха, но требует много воды для роста плодов.
Растения томата требовательны к свету. При его недостатке задерживается
развитие растений, листья бледнеют, образовавшиеся бутоны опадают, стебли
сильно вытягиваются. Досвечивание в рассадный период улучшает качество рассады
и повышают продуктивность растений.
При внесении органических и минеральных
удобрений и поддержания грунта в рыхлом состоянии томат может расти на любых
(кроме очень кислых) почвах. Основные элементы минерального питания для
томатов, как и для других растений - азот, фосфор и калий. В азоте томат
особенно нуждается в период интенсивного роста плодов, однако переизбыток азота
нежелателен, поскольку это приводит к сильному нарастанию вегетативной массы
(т. н. жирование растений) в ущерб плодоношению, а также интенсивному
накоплению в плодах нитратов. При недостатке фосфора растения томатов слабо
усваивают азот, вследствие чего прекращается их рост, задерживается
формирование и созревание плодов, листья приобретают сине-зелёную, затем
сероватую, а стебли лилово-коричневую окраску. Фосфор особенно необходим
томатам в начале вегетации. Усвоенный растениями в этот период, он идёт затем
на формирование плодов. Калия томат потребляет больше чем азота и фосфора. Он
особенно нужен растениям в период роста плодов. При недостатке этого элемента
по краям листьев появлются жёлто-коричневые точки, они начинают скручиваться, а
затем отмирают. Томатам также необходимы микроэлементы, влияющие на рост и
развитие растений: марганец, бор, медь, магний, сера и др. Их вносят в виде
микроудобрений.
Технология выращивания.
Всходы томатов.
Рассада помидоров 1,5 месяцев после прорастания
семян.
Посев томатов производят в парники ещё зимой, с
таким расчётом, что через месяц после 2-й пикировки можно было высадить их
прямо в грунт, не боясь заморозков, или в полухолодные парники. При очень
ранней посадке растения могут быть готовыми к пересадке в грунт ещё в то время,
когда земля не готова к этому, и оставшиеся в парнике растения, будучи тесно
расположены, начинают вытягиваться и бледнеть, делаясь слишком чувствительными
к изменениям температуры. Ввиду этого время посадки должно быть строго
согласовано с местными климатическими условиями. В случае заморозков растения
необходимо покрывать старыми ящиками, рогожами или матами.
В первое время роста всходов в тёплом парнике
приходится наблюдать лишь за проветриванием парника и за ограждением всходов от
сорных трав и вредителей. Недели через 3-4 после посева, когда появится вторая
пара листьев с зубчиками, приступают к первой пикировке, пересаживая в тёплый
же парник, но с большим слоем земли; самая пикировка производится так же, как и
с капустой, причём под раму высаживают до 300 растений, если предстоит вторая
пикировка, или только до 200, если впоследствии растения будут высажены прямо в
грунт, без 2-й пикировки. Во втором парнике наблюдают за проветриванием
последнего не только во избежание сырости и плесени, но и в видах закаливания
растений.
Через месяц после первой пикировки, когда
растения станут слишком теснить друг друга, приступают ко второй пикировке,
перемещая растения уже более свободно (не более 200 растений под раму),
поднимая парниковый ящик и все менее и менее прикрывая растения рамами, с тем
чтобы приучить растения к наружному воздуху. Окончательную пересадку в грунт производят
приблизительно через месяц после 2-й пикировки, когда нет уже опасности перед
заморозками. В тех случаях, когда желают получить более ранние плоды, например
- в начале или середине июня, посев в парниках производят возможно раньше и
перед посадкой в грунт производят три пикировки.
Пересадку растений из парников производят в
горшки, причём их держат в открытых парниковых ящиках, прикрывая рогожами лишь
на ночь и при понижении температуры. Окончательную пересадку грунт из горшков
производят, не нарушая кома земли и закапывая его в заранее приготовленные
ямки. Пользование горшками позволяет садовнику не торопиться с пересадкой и
выждать наверняка благоприятного времени, так как в горшках растения продолжают
правильно развиваться. Что же касается месторасположения для томатов, то они
любят освещенный, сухой, хорошо орошаемый грунт. Свежего удобрения томат не
выносит, подвергаясь картофельной болезни; хорошо удаётся томат после капусты,
получившей богатое удобрение. Высаживают томаты рядами, тесная посадка вредна
во всех отношениях. Немедленно после посадки растения поливают, и эту поливку
продолжают до тех пор, пока растения не примутся.
В начале периода после посадки, когда ночи ещё
прохладные, следует избегать поливки после захода солнца, так как это вызвало
бы ещё большее охлаждение земли. Вдоль всей плантации проводят бороздки для
орошения растений. Томаты довольствуется орошением, и поливку из лейки самых
растений необходимо производить лишь при крайне сильных засухах, да и то раза
два в лето. При дальнейшем росте растений необходимо подвязывать и подрезать
растения (шпалерный способ разведения), что содействует равномерному освещению
растений, лучшему проветриванию, а следовательно, и более обильному, и более
раннему созреванию плодов. После обрезания растения таким образом, что остаются
лишь 2-3 сильных побега, промежуточные же удаляются, томаты подвязывается или к
шпалерам (решеткам, проволокам и т. п.), или к кольям, причем должно быть
наблюдаемо, чтобы каждый стебель развивался вполне свободно. Дальнейший уход
заключается в удалении жировых побегов и поправке подпорок.
Сбор плодов начинается с начала июня и
продолжается, смотря по местности, до середины сентября. Перед наступлением
холодов растения во избежание замерзания выдёргивают из земли вместе с плодами
и кладут в парниковые ящики, где и происходит дозревание плодов. Самый сбор
плодов производят с помощью ножа или ножниц. Собранные плоды переслаиваются
соломой. При пересылке их кладут в ящики не более, как в два слоя.
Вредители, болезни и методы борьбы с ними.
Вредителями томатов являются медведка,
комарики-сциариды чёрного цвета, тепличная белокрылка, картофельные тли и
некоторые другие насекомые: (хлопковая совка, колорадский жук).
Болезни томатов могут быть вызваны избытком или
недостатком азота, калия, фосфора или же грибками и вирусами:
фитофтороз
мозаика (вирус Nicotiana
virus J.),
бронзовость листьев (вирус Lycopersicum
virus),
корневая гниль (возбудитель - гриб Thielaviopsis
basicola),
ризоктониозная гниль плодов (гриб Rhizoctonia
solani Kuehn.),
розовая гниль плодов (гриб Fusarium
gibbosum Арр. et
Wr.),
серая гниль (гриб Botrytis cinerea
Pers.),
стеблевая гниль томатов (гриб
Didymella lycopersici),
фомоз (бурая гниль; гриб Phoma
destructiva Plowr.),
фузариозное увядание (гриб Fusarium
oxysporum f. lycopersici.),
антракиоз (гриб Colletotrichum
atramentarium (Berk. et Br.) Taub.),
бурая пятнистость листьев, кладоспориоз, или
листовая плесень (гриб Cladosporium
fulvum Cooke.),
вертициллезное увядание (грибы Verticillium
albo-atrum
и V. dahliae).
Также встречаются болезни разной природы,
проявляющиеся в растрескивании плодов, скручивании листьев томатов.
Использование
Плоды томата употребляют в пищу свежими,
варёными, жареными, консервированными, из них готовят томат-пасту, всевозможные
соусы, соки, лечо.
3 Рабочая гипотеза
Так как необходимо вывести ликопидосодержащий
гибрид помидор для диетического питания в санатории, то рассмотрим
климатические условия местности. Санаторий «Ласточка» находится в Тюменской
области. Для того чтобы решить вопрос о получении определенного сорта помидор
необходимо рассмотреть климат в котором они будут выращиваться.
Отличительными особенностями погоды Тюменской
области являются продолжительная, умеренно суровая снежная зима и теплое лето,
что характерно для континентального климата. Разница температур лета и зимы
составляет в среднем около 35 С (для примера, в Прибалтике в тех же широтах эта
разница соответствует 22-23 С). Средняя температура на юге Тюменской области в
январе составляет около -19-17 С, в июле - +19 С. Зимой в отдельные дни почти
ежегодно температура ночью понижается до -36-44 С, но в январе бывают и
оттепели до +2 С. А в летний период нередки заморозки до -2-3 С.
Количество осадков колеблется от 200 до 600 мм в
год, они выпадают преимущественно с мая по октябрь. Продолжительность
солнечного сияния здесь гораздо выше, чем в таких же широтах Европейской части
страны, а подчас и в городах, расположенных гораздо южнее (например, в Киеве),
она достигает 2017 часов. Зимой также часто наблюдаются сильные ветры с
метелями (до 15-20 м/с). На юге Тюменской области теплая погода наступает
довольно рано - в апреле и держится до середины или конца октября. Поэтому
сезон для познавательного туризма длится довольно долго. Для любителей
курортного отдыха подойдут, пожалуй, лучше всего летние месяцы. Для путешествий
по западной Сибири также можно порекомендовать короткий период «бабьего лета» в
сентябре - в это время пейзажи особенно красивы. Плоский, равнинный рельеф юга Тюменской
области расцвечивается осенними красками. Для фотографирования хорош также
достаточно солнечный месяц март. Для любителей охотничье-рыболовного туризма
может представлять интерес и сибирская зима, когда открыта охота на копытных.
После Нового года, с установлением прочного снегового покрова, возможны
охотничьи сафари на снегоходах.
Так как Тюменскую область отличает
продолжительная зима, то для развития производства необходимо на территории
санатория создать теплицу для выращивания помидор.
Из поставленной задачи известно, что в хромосоме
2 расположен рецессивный ген (у) - 29 локус и ген доминант (Lr)
- 53 локус. Первый определяет высокое содержание ликопина и желтые плоды, а
второй ген - густое опушение.
Исходя из условий можно изобразить группы сцепления
томатов:
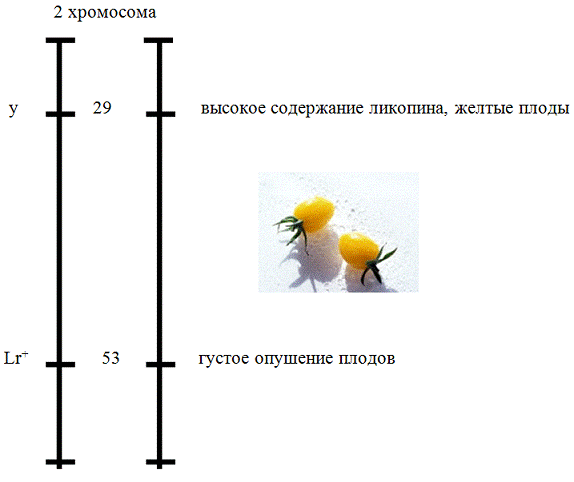
Соответственно у+, характеризуется красными
плодами, а Lr -
неопушенными плодами.
Находим расстояние между генами: 53 локус - 29
локус = 24, это значение меньше 50% таким образом скрещивание является
дигибридным.
Первое поколение растений будет иметь вид:
y Lr+ х
y+Lr F1 y Lr+
y
Lr+
y+Lr
y+Lr
4 Доказательство рабочей гипотезы
Для доказательства рабочей необходимо
использовать данные третьей главы.
Рассмотрев их видим, что из первого поколения
растений:
Некроссоверы y
Lr+ и y+Lr
их 76%, делим на два и получаем 38% на каждый.
Кроссоверы y
Lr и Lr+
y+ их 24%, делим на
два и получаем 12% на каждый.
Исходя из выше перечисленных данных необходимо
составить решетку Пинета.

Составив решетку Пинета получили:
гибридов - густоопушенных красноплодных
гибрида - густоопушенных желтоплодных
гибрида - неопушенных красноплодных
гибрид - неопушенный желтоплодный
Во втором поколении было получено 1000 гибридов.
Необходимо вывести густоопушенные желтоплодные
гибриды.
Желтоплодные плоды содержат пектиновые и
азотистые вещества, сахара, аскорбиновую, лимонную, яблочную, щавелевую, винную
кислоты, флавоноиды, алкалоиды, тиамин, рибофлавин, каротин, пурины, клетчатку,
минеральные соли (особенно много калия).
У желтоплодных и оранжевых сортов каротина
значительно больше. Как показали исследования последних лет, ликопин также
способен в организме человека и животных превращаться в витамин А (ретинол).
Помидоры - поливитаминное растение. Благодаря
витаминной ценности, высокому содержанию солей калия, железа, магния, кобальта
и цинка, их включают в пищу больных сердечно-сосудистыми заболеваниями, с
нарушением обмена веществ, что необходимо для здорового питания в санатории. Их
рекомендуют при заболеваниях желудочно-кишечного тракта, так как они бедны
клетчаткой, а их нежная мякоть легко переваривается и усваивается. Они
проявляют нежное слабительное действие, усиливая перистальтику кишок. Имеются
данные о способности свежих томатов и томатного сока умеренно понижать кровяное
давление и уменьшать уровень холестерина в крови. Свежая плодовая мякость и
томатный сок тормозят рост и развитие ряда микроорганизмов.
В народной медицине плоды и свежий томатный сок
применяют при нарушениях пищеварения, язвенной болезни желудка, истощении.
почечнокаменной болезни. Кашицу из свежих помидоров прикладывают к язвам.
Из расчетов следует:
- 100%
- 10,56%
Таким образом во втором поколении вывели 106
гибридов густоопушенных желтоплодных для диетического питания в санатории
«Ласточка».
Растения выращивались в теплице, которая
рассчитана 500 кустов.
Для того, чтобы рассчитать полученную прибыль от
выращивая гибридов, выполним действия:
кустов растений х 10 (среднее количество
килограмм полученное с одного куста) = 5000 кг.
х 80 рублей (средняя стоимость одного килограмма
помидор) = 400000 р.
Таким образом при рациональном ведении хозяйства
и содержания теплицы возможно получить прибыль в 400000 р.
Дополнительные вопросы:
.Сколько типов гамет образуют растения первого
поколения?
кроссоверные
некроссоверные
.Сколько растений второго поколения имеют густое
опушение и красные плоды?
- 100%
- 51,44%
гибрида или 51,44%.
.Сколько растений второго поколения неопушенные
красноплодные?
- 100%
- 23,56% 236 растений или 23,56%
.Сколько процентов растений неопушенные
желтоплодные?
,44%
.Сколько генотипов во втором поколении? 9
генотипов
Список используемой литературы
гибрид помидор сцепление
наследование
1. Бочкова, Н.П. Генетика /
Под ред. Бочкова Н.П. - М.: Мастерство, 2001.
.Голощапов, А.П. Генетика: курс
лекций и практических занятий. Курган, ГИПП «Зауралье», 2001. - 350 с.
. Орехова, В.А., Лажковская Т.А.,
Шейбак М.П. Генетика. - Минск: Высшая школа, 1999.
. Чебышева, Н.В. Биология / Под ред.
Чебышева. Н.В. - М.: ГОУ ВУНМЦ,2005.
. Ярыгин, В.Н., Волков И.Н. и др.
Биология. - М.: Владос, 2001.