Видовой состав летне-осеннего фито- и зоопланктона Чёрного моря у берегов Абхазии
Видовой состав летне-осеннего фито- и
зоопланктона чёрного моря у берегов Абхазии
Оглавление
Введение
Глава 1. Физико-географическая и гидрологическая характеристика
Чёрного моря
1.1 Современное состояние Чёрного моря
1.2 Рельеф дна и грунты
1.3 Берега и реки
1.4 Прозрачность и цвет
1.5 Атмосферная циркуляция
1.6. Течения
1.7 Солёность
1.8 Температурный режим
1.9 Концентрация растворённого кислорода
1.10 Биогенные элементы
1.11 Нефтяное загрязнение абхазской акватории Черного моря
Глава 2. Методы исследования планктона
2.1 Орудия для сбора планктонных организмов
2.1.1 Качественная сеть Апштейна
2.1.2 Планктонособиратель
2.1.3 Батометр
2.2 Консервирование и этикетирование проб планктона
2.3 Качественная обработка проб
2.4 Количественная обработка проб
Глава 3. История изучения планктона Чёрного моря
Глава 4. Аннотированный список летне-осеннего фито - и зоопланктона
Чёрного моря
4.1 Аннотированный список фитопланктона
4.2 Аннотированный список зоопланктона
Глава 5. Экология. Экологическое и биологическое значение фито - и
зоопланктона Чёрного моря
5.1 Миграции планктона
5.2 Пищевые цепи
5.3 Биомасса и продуктивность
5.4 Динамика численности и биомассы планктона в связи с влиянием
вселенцев - Mnemiopsis leidyi и Beroe ovatа
Заключение
Литература
Введение
Чёрное море населяют примерно 2900 видов организмов, из
которых около 850 видов относятся к особой группе пелагических гидробионтов -
планктону.
Планктон представляет собой группу водных организмов, которые
ведут свободно-плавающий, независимый от твердого субстрата, как опорного
элемента, взвешенный в толще воды образ жизни и отличаются или полным
отсутствием органов движения (многие представители фитопланктона), или
последние у них довольно слабы, поэтому эти организмы не могут противостоять даже
слабым течениям, а их активные перемещения происходят в относительно небольших
пределах. В состав данных гидробионтов входят фито-, зоо - и ихтиопланктонные
организмы.
Многие из этих видов средиземноморского происхождения - они
проникли сюда через Босфор после соединения Чёрного моря со Средиземным,
которое произошло 6 тыс. лет назад. Среди планктёров встречаются как
одноклеточные, так и многоклеточные организмы, относящиеся к разным
таксономическим группам. Они интересны по своему строению, морфологии, экологии
и по биологическому разнообразию.
Планктон Чёрного моря играет существенно важную роль в
морской экосистеме, так как является необходимым и незаменимым звеном в пищевых
цепях. Планктёры связаны тесными пищевыми связями со всеми другими обитателями моря.
Фито - и зоопланктонное сообщество - самое богатое и быстрее всего
восстанавливающееся звено пищевых цепей. Ими питаются личинки практически всех
видов рыб, планктоядные рыбы, многие виды моллюсков и различные другие
гидробионты.
Эти обстоятельства делают актуальным проблему изучения
видового состава планктона и динамики их численности. Этой проблеме и посвящена
данная работа, целью которой является выявление видового состава летне-осеннего
планктона.
Исходя из данной цели, нами были поставлены следующие задачи:
· идентификация фитопланктонных организмов.
· идентификация зоопланктонных организмов.
· выявление наиболее массовых видов
планктёров, имеющих кормовое значение.
· оценка литературных данных по динамике
численности и биомассы фито - и зоопланктона.
Работа выполнена на кафедре экологии и морфологии животных
Абхазского Государственного Университета, под руководством доцента Дбара Романа
Саидовича. При подготовке данной работы активную помощь оказывали сотрудники
отдела планктона Азовского Научно-исследовательского Института Рыбного
Хозяйства (АзНИИРХ). На разных этапах выполнения дипломной работы существенную
помощь оказывали сотрудники кафедры экологии и морфологии животных АГУ. Всем им
мы выражаем искреннюю благодарность.
планктон черное море экологический
Глава 1.
Физико-географическая и гидрологическая характеристика Чёрного моря
1.1
Современное состояние Чёрного моря
Чёрное море - типичное внутреннее море. На западе его воды
омывают восточные берега Балканского полуострова, на севере оно ограничено
Восточно-Европейской равниной, на востоке - Кавказским побережьем и Колхидской
низменностью, а на юге - побережьем Малой Азии. Длина береговой линии, по
данным различных авторов, составляет 4020-4100 км. Черноморская впадина имеет
вытянутую в широтном направлении, близкую к овальной, форму с довольно простым
общим очертанием и крутыми склонами.
Чёрное море является внутренним морем Атлантического океана.
На северо-востоке оно соединяется с Азовским морем Керченским проливом, на
юго-западе - Босфорским проливом с Мраморным морем.
Площадь Чёрного моря составляет 420325 км2, общий
объём воды 550000 км3. Средняя глубина Чёрного моря 1271 метр,
наибольшая 2245 метров (вблизи Синопа). Самая северная точка находится на 46○33'
с. ш. (у Березанского лимана, около Очакова); самая восточная - на 41°42' в. д.
(между Батуми и Поти); самая южная - на 40°56' с. ш. (у города Гиресун западнее
Трапзона); самая западная - 27°27' в. д. (в Бургасском заливе). (Экба и др.
2007).
Береговая линия Чёрного моря сравнительно слабо изрезана.
Наиболее расчленённые северо-западные и северные берега связаны с последним
этапом эволюции Черноморского бассейна. Здесь расположены обширные заливы -
Одесский и Каркинитский. На Кавказском берегу самый большой залив -
Новороссийская бухта, а на турецком - Синопский залив, бухта Самсун и Вона
залив, в европейской части побережья - залив Игнеада. На болгарской части самые
большие заливы - Бургасский и Варненский.
Черноморский берег беден полуостровами. Среди них самый
большой - Крымский полуостров (и связанный с ним Керченский). На побережье
Турции самые большие полуострова - Инджебурун и Ясун.
Чёрное море весьма бедно островами. Тремя самыми большими
островами являются Змеиный (Фидониси, у Сулинского гирла реки Дуная), Березань
(у одноимённого лимана) и Кефкен (в 92 км к востоку от Босфора).
Остальные черноморские острова незначительны по площади - в
большинстве случаев представляют собой большие или малые скалы.
В чёрное море впадают реки двух континентов - Европы и Азии.
Водосборная
площадь самой большой реки Дуная 820 тыс. км2, рек
Днестра, Дона (впадает в Азовское море), Днепра, Южного Буга - меньше. Самые
большие реки, текущие с востока, - Кубань и Риони, с юга - Кызыл-Ирмак,
Емиль-Ирмак, Сакарья и Чорох. На Болгарской территории самые большие реки,
впадающие в Чёрное море, - Камчия, Велека, Резовская.
1.2 Рельеф
дна и грунты
В рельефе черноморского дна чётко выделяются три основные
формы - шельф, материковый склон и глубоководная котловина.
Шельф начинается от современной береговой линии и
заканчивается там, где уклон подводного склона резко меняется. Шельф
представляет собой непосредственное продолжение суши, оказавшейся под водами
моря после последнего оледенения (10-12 тыс. лет назад). В Чёрном море шельф
заканчивается в среднем на глубине 90-110 м и только южнее Севастополя и Ялты -
на глубине 140-160 м. Шельф занимает 24% площади дна Чёрного моря. Ширина
черноморского шельфа различна в разных частях моря. На северо-западе, где в
море впадают реки Дунай, Днестр, Южный Буг, Днепр, максимальная ширина шельфа
достигает 200-250 км, а у Кавказского и Малоазиатского берегов ширина его лишь
несколько километров.
Шельф занимает значительную площадь на северо-западной,
западной и юго-западной перифериях Черноморской котловины, тогда как у Кавказского
и Анатолийского берегов он представлен узкой прерывистой полосой. Различные
участки шельфа образовались в разное время, и имеют разное геологическое
строение (Вылканов и др., 1983).
Материковый склон составляет 40% площади дна. Он является
продолжением шельфа. Согласно исследованиям советских геофизиков, В.Т.
Гончарова и др., в Чёрном море можно выделить 3 типа материкового склона. К
первому типу относится крутой изрезанный склон, расположенный близко к берегу.
Уклон его местами достигает 20-30°. Склоны этого типа обычны для Кавказского и
Анатолийского побережья Чёрного моря.
Ко второму типу относится склон прямолинейных сглаженных
очертаний, постепенно понижающийся, крутизной 1-3°. Это склон у Странджанского
побережья и к югу от Керченского пролива. Склон с более сложными очертаниями и
более разнообразным рельефом относится к третьему типу. Это участки между мысом
Калиакра и Крымским полуостровом, Анатолийское побережье
между Инеболу и Синопом.
Глубоководная котловина занимает центральную часть Черноморской
впадины и имеет форму овала, вытянутого с запада на восток. Большая часть дна
котловины находиться на востоке бассейна и занимает 1/3 площади Чёрного моря.
Глубина его колеблется между 2100 и 2200 м. Дно котловины представляет собой
равнину с идеально сглаженным рельефом. Самые большие глубины обнаружены вблизи
Анатолийского берега (2245 м).
Кавказский район отличается узким шельфом и крутым
расчленённым материковым склонам. Северо-западнее портового пункта Гагра
материковый склон начинается непосредственно у берега. Ширина шельфа 1-2 км,
порта Сочи до 12-18 км. Глубина внешнего края шельфа не превышает 110 м, а
юго-восточнее портового пункта Геленджик составляет 67-75 м.
Переход от шельфа к материковому склону чётко выраженный,
резкий, иногда обрывистый. Верхняя часть склона от внешнего края шельфа до
глубин 1200-1400 м крутая (угол наклона в среднем 15-17º, местами 30º и более), расчленена
подводными долинами и разделяющими их поднятиями. Подводные долины имеют V -
образный поперечный профиль и далеко вдаются в шельф. Наиболее крупные из них
прорезают шельф почти от самого берега и прослеживаются на склоне до его
подножия. Ряд подводных долин находится напротив устьев рек, но многие из них
никакой связи с реками не имеют.
Юго-восточный район. Шельф и материковый склон этого района
отличаются сложностью очертаний и строением рельефа. Юго-восточнее мыса Пицунда
и вблизи портового пункта Очамчира шельф образован выступами, его ширина
составляет 20-30 км. В остальной части района ширина шельфа колеблется от 3 до
8 км. Внешний край шельфа расположен на разных глубинах: южнее мыса Пицунда на
глубине 300-500 м, в остальной части района на глубинах 100-110 м. Поверхность
шельфа выровнена, но угол наклона её меняется от нескольких минут до полутора
градусов в местах сужения. Между мысом Пицунда и портом Сухум примерно на
глубине 100 м прослеживается уступ высотой 5-16 м. Ниже уступа на глубинах
130-140 м встречаются небольшие возвышенности и борозды.
Материковый склон от внешнего края шельфа до глубин 800-1000
м выражен в рельефе крутым уступом. Угол наклона колеблется от 3º до 10º и более. Поверхность
склона, за исключением участка между мысом Пицунда и портом Сухум, расчленена
многочисленными подводными долинами. Верховья некоторых из них подходят почти
вплотную к берегу. Крупные долины продолжаются до самого подножия склона.
Большинство форм рельефа вытянуты в широтном направлении.
Донные осадки Чёрного моря представлены всеми
гранулометрическими типами (валуны, галечник, гравий, песок, илистый песок,
песчанистый ил, ил и глинистый ил). Среди песчаных и илистых отложений в
качестве примеси встречается большое количество раковинного материала. А в
северо-западном районе на шельфе отмечаются большие площади чисто ракушечных
отложений.
Узкая полоса обнажённых скальных (коренных) пород отмечается
в прибрежной части у гористых берегов (между портом Новороссийск и портовым
пунктом Геленджик) и на материковом склоне. Среди гранулометрических типов
донных осадков на шельфе преобладают ил и глинистый ил.
В Северо-западном районе в отличие от других районов на общем
фоне песчанистых илов заметно выделяются участки илов и песков, среди
ракушечников встречаются участки песков и песчанистого ила. Такое распределение
осадков в значительной степени обусловлено рельефом дна. Пески соответствуют
мелководьям или отдельным поднятиям дна, илы и песчанистые илы - понижениям
дна. Границы и площади, занимаемые разными типами осадков на шельфе, подвижны.
Различия в условиях осадкообразования на разных участках
шельфа обуславливают и различия в содержании таких важных для продуктивности
компонентов, как органическое вещество и микроэлементы. С распределением
органического вещества связывается и распределение микроэлементов, играющих
важную роль для организмов.
В юго-восточном районе осадки на шельфе представлены в
основном илом и глинистым илом, на глубинах менее 30 м - песком и галькой.
Материковый склон покрыт илом.
В Турецком районе на шельфе и материковом склоне преобладают
илы и глинистые или, а в прибрежной части, на глубинах менее 30 м -
песчано-ракушечные отложения.
В Юго-западном районе донные осадки на шельфе представлены
песчано-ракушечными и илистыми отложениями. Песчано-ракушечные отложения
отмечаются в основном на глубинах менее 30 м на валообразных поднятиях шельфа,
илы - в понижениях дна. На глубинах 100 м и более донные осадки представлены
илом и глинистым илом. В центральном районе дно покрыто илом и глинистым илом.
1.3 Берега и
реки
Берега Чёрного моря составляют почти непрерывную линию, как у
озера. Такие моря называют замкнутыми или внутренними - они замкнуты внутри
суши, окружены ею со всех сторон. Чёрное море соединяется со Средиземным морем
и через него - с Атлантическим океаном двумя узкими проливами. Первый пролив -
Босфор, это 30-километровый водный коридор, разделяющий Европу и Азию, он ведёт
из Чёрного моря - в маленькое Мраморное море. Второй пролив - Дарданеллы, через
которые можно выйти из Мраморного моря в Средиземное море. Дарданеллы -
достаточно широкий и свободный водный путь, а Босфор - узкий (через него даже
перекинут мост) и неглубокий пролив. Поэтому свободного обмена водой,
перемешивания между Чёрным и Средиземным морями нет.
Нет ни одного другого внутреннего моря в мире, в которое
втекает столько пресной воды. Её приносят крупнейшие реки Европы: Дунай, Днепр,
Днестр, Буг, Дон, Кубань, Риони и тысячи мелких речек и ручьев. Всего 350 км3
речной воды в год (Вершинин, 2007).
Дунай - вторая по мощности после Волги река в Европе, он один
приносит больше половины воды в Чёрное море. Очень большой речной сток -
ключевой физический фактор, определяющий свойства вод Чёрного моря и его
биологическую структуру. Приток океанической воды через Босфор мал, а реки приносят
пресную воду, которая разбавляет морскую. В результате - в черноморской воде в
два раза меньше соли, чем в океане или в соседнем Средиземном море.
Восточную часть Чёрного моря можно рассматривать как
отдельную и самостоятельную область. Здесь однотипные берега, сформированные
под совместным влиянием речных наносов и волнений. По всей береговой дуге
преобладают волны, подходящие с запада. Современная береговая зона восточной
части Чёрного моря образовано в результате совместной деятельности рек, выносящих
песчаный, галечный и валунный материал, а также волнений, перемещающих речной
аллювий вдоль морского берега.
В современной динамике морских берегов Причерноморья всё
более возрастающую роль играют антропогенные вмешательство в естественный
режим. В результате этого береговая зона испытывает сильный дефицит
пляжеобразующих наносов, и берег повсеместно размывается и отступает. Таким
образом, пляжевые наносы своей крупностью отображают особенности рельефа
прилегающей суши и морфодинамику речных русел.
1.4
Прозрачность и цвет
Черноморская вода по своим гидрофизическим особенностям
отличается от воды других морей и океанов.
Цвет поверхности моря зависит от содержания в воде
неорганических и органических взвесей, от цвета неба, глубины, а вблизи берега
и от цвета донных отложений. Чёрное море, называемое в древности тёмноцветным,
действительно имеет более тёмные воды в сравнении со светло-зелёными водами
мелкого Азовского моря, которые похожи скорее на озёрные воды. Черноморские
воды содержат значительно больше планктона, а в прибрежье - детрита и глинистых
осадков, чем мраморноморские и эгейские воды.
Цвет Чёрного моря не везде одинаков; он изменяется в
зависимости от сезона и погоды. В прибрежной мелкой опреснённой части моря воды
обычно зеленоватого цвета, на расстоянии 30-40 миль от берега - чисто синего, в
промежуточной зоне - сине-зелёного цвета различных оттенков. Прибрежным водам
зелёный цвет придаёт в основном обилие фитопланктона. В тихие летние дни, когда
развитие фитопланктона незначительно, вода даже вблизи берега сине-зелёного
цвета.
Бытовые и промышленные стоки различным образом изменяют цвет
воды, делают его чёрным или коричневым (Вылканов и др., 1983).
Цвет воды тесно связан с её прозрачностью. Жёлто-коричневые
воды, содержащие глинистые взвеси, ярко-зелёные воды, насыщенные
фитопланктоном, темнее, окрашенные детритными частицами воды малопрозрачны, а
сине-зелёные и особенно синие воды прозрачны.
Вблизи устьев рек и в мелководных районах после дождя или
волнения прозрачность воды, измеренная диском, составляет лишь сантиметры. В
открытых прибрежных водах прозрачность в большинстве случаев 2-8 м, в
глубоководных районах она увеличивается почти в 3 раза.
Для сравнения отметим, что в морях, бедных планктоном,
прозрачность значительно больше. В открытых районах Средиземного моря она
достигает 50-60 м, а в Саргассовом море - 67 м.
Разные слои Чёрного моря имеют разную прозрачность.
Вертикальное распределение вод различной прозрачности в Чёрном море связано с
положением слоя скачка плотности, в котором скапливается планктон, его останки,
и другие органические и неорганические взвеси.
1.5
Атмосферная циркуляция
Над Чёрным морем можно выделить 9 типов атмосферной
циркуляции, 7 из которых соответствуют основным направлениям ветровых потоков
(северо-восточный, восточный, юго-восточный, юго-западный вместе с южным,
западный и северный). Восьмой - циклонический, к девятому типу отнесены мало
градиентные барические поля.
Северо-восточный тип. Район Чёрного моря занят юго-восточной
периферией обширного антициклона с центром над западными районами европейской
территории бывшей СССР. На юго-востоке Чёрного моря развивается циклоническая
деятельность или наблюдается пониженное давление. Прохождение циклов часто
сопровождается восточными и северо-восточными ветрами со скоростью 12-15 м/с,
иногда до 32 м/с. Зимой наблюдается сильное похолодание, летом - жара при
большой сухости воздуха. Средняя годовая повторяемость этого типа 12,4 %,
максимальная повторяемость (август и сентябрь) 20 %.
Восточный тип. Центр антициклона расположен над центральными
районами европейской территории бывшей СССР. Над Средиземным морем и Турцией
развивается циклоническая деятельность. При смещении средиземноморских циклонов
на юг Чёрного моря над всем морем наблюдается усиление восточного ветра.
Средняя годовая повторяемость этого типа 6,6 %.
Юго-восточный тип. Антициклон расположен над Казахстаном и
восточными районами и восточными районами европейской территории бывшей СССР.
Над Средиземным морем и Балканским полуостровом находится депрессия. При
смещении средиземноморских циклонов на юго-запад Чёрного моря наблюдается
усиление юго-восточного ветра. Средняя годовая повторяемость 7,8 %,
максимальная повторяемость (ноябрь) 14,7 %.
Юго-западный тип. С Балтийского моря на Балканы направлена
барическая ложбина. При развитой в этой ложбине вблизи Чёрного моря циклонов
возникают сильные южные и юго-западные ветры. Средняя годовая повторяемость 7,4
%, максимальная повторяемость (декабрь, январь) 13-14 %.
Западный тип. При прохождении глубоких циклонов со
Скандинавского полуострова по югу Украины и ли в тылу средиземноморских
циклонов над Чёрным морем возникают очень сильные ветры. Средняя годовая
повторяемость 2-5 %.
Северо-западный тип. Над Западной Европой располагается
антициклон с отрогом, направленным на Балканский полуостров; на юго-востоке
европейской территории бывшего СССР наблюдается циклоническая деятельность. В
тылу глубоких циклонов, смещающихся со Скандинавского полуострова на юго-восток
европейской территории бывшей СССР, над Чёрным морем возникают очень сильные
северо-западные ветры. Повторяемость этого типа меняется от 3,3 % в марте до
10,7 % в июле и составляет в среднем за год 5,8 %.
Северный тип. Обширный антициклон занимает Западную Европу;
над Кавказом и восточной частью Чёрного моря развивается циклоническая
деятельность. При быстром вторжении на Чёрное море с Балканского полуострова
антициклона или его отрога наблюдается усиление северного ветра. Средняя
годовая повторяемость 8,4 %, максимальная повторяемость (июль - сентябрь) 11,6
- 12,7 %.
Циклонический тип формируется при прохождении над Чёрным
морем атмосферных фронтов или депрессий со Средиземного моря. Характеризуется
ветрами южных, западных и северо-западных направлений. Зимой при этом типе
погоды наблюдаются значительная облачность и обложные осадки, летом - грозы.
Средняя годовая повторяемость этого типа 7,3 %, максимальная повторяемость в
феврале 13,2 %, в апреле 10,5 %. Малоградиентные барические поля,
характеризующиеся неустойчивыми слабыми ветрами, имеют самую высокую по
сравнению с другими типами среднюю годовую повторяемость (41%), максимальная
повторяемость (май - июль) 51,8 - 63,3 %.
Температура воздуха в течении года имеет чёткий сезонный ход
с одним максимумом. Самыми холодными месяцами являются январь и февраль, когда
температура воздуха опускается в северо-западной части моря до -2º С, а в южной и юго-восточной частях моря до 8º С. В марте температура воздуха в северо-западной части моря
повышается до 2º С. В дальнейшем, с увеличением притока
солнечного тепла воздух прогревается, и к июлю формируется практически
однородное поле с температурой 23-24º С над всем морем, самый
тёплый месяц - август.
Наиболее часто резкие похолодания над Чёрным морем вызываются
северными, северо-восточными и северо-западными ветрами. В суточном ходе
температуры воздуха минимум наблюдается зимой в 7-9 ч, весной и осенью в 6-7 ч,
летом в 5-6 ч. Максимум в течении года отмечается в 14-16 ч.
1.6 Течения
Ветер у берегов и в открытом море,
неравномерное пространственное распределение плотности морской воды, очертания
берегов, характер подводного рельефа, отклоняющее действие вращения земли - все
эти факторы в той или иной степени, участвуют в формировании систем течений
Черного моря.
Кроме того, одним из важнейших факторов
является береговой сток, вливающий большое количество пресной воды в море, а
также выпадающие на его поверхности осадки, создающие разницу уровней между
отдельными частями моря. Так например, эти факторы между северо-западным
приодесским и прибосфорским участками создают разницу в уровне 30 см и вызывают
постоянное компенсационное течение с севера на юг. Основное течение Черного
моря имеет кольцевой характер и движется против часовой стрелки. В наиболее
узком месте моря (между Крымским полуостровом и выступом Малоазиатского побережья)
часть двигающихся с запада вод уходит на север и, таким образом, как бы делит
море на две части, каждая со своим круговым течением. Основная струя течений
располагается в расстоянии 3-9 км от берега и не подвергается влиянию местных
особенностей очертаний берегов и рельефа дна и имеет ширину 45 - 55 км.
Скорость течения в ней чаще 0,9 - 2,0 км в час, но при сильных штормовых ветрах
может достигать 2,5 - 5,5 км в час. Вдоль Абхазского побережья проходит струя
восточного круговорота течений, которая имеет скорости 0,3 - 0,7 км/час.
У берегов располагается зона прибрежных
круговоротов течений, направленных также, как и основное течение, против
часовой стрелки. Они в навигационном слое воды имеют скорость свыше 1 км в час.
Основное черноморское течение направлено против часовой
стрелки по всему периметру моря образуя два заметных кольца (очки Книповича, по
имени одного из русских гидрологов, описавших эти течения). В основе такого
движения вод и его направленности лежит ускорение, придаваемое воде вращением
Земли - Кориолисовой силой. Однако, на такой, относительно небольшой акватории,
как Чёрное море, направление и сила ветра имеют не меньшее значение, и по этой
причине основное черноморское течение очень изменчиво. Это течение
распространяется только на верхний слой моря, не глубже 100-150 м.
В прибрежье образуется вихри противоположной основному
черноморскому течению направленности - антициклонические круговороты, особенно
они выражены у Кавказского и Анатолийского берегов.
Хотя основное течение Черного моря не
оказывает никакого влияния на его берега, однако, оно имеет большое значение в
жизни Черного моря. Там, где идет основное течение, все изолинии опускаются
вниз, а во внутренних областях, наоборот, куполообразно приподняты. С этим
связано распространение сероводорода в Черном море. В первом случае верхняя
граница сероводорода, по данным Л.А. Зенкевича (1947), опускается до 155 м, а
во втором - поднимается до 100 м от поверхности моря. Таким образом, мощность
слоя воды Черного моря, населенного животной жизнью может претерпевать весьма
значительные изменения в разных частях моря. Характер передвижения водных масс
отражается и в распределении нижней границы распространения планктона.
1.7 Солёность
Морская вода представляет собой универсальный раствор, в
состав которого входят все известные элементы. Пропорции между концентрациями
главных компонентов остаются примерно постоянными, а различия в содержании
касаются только общего количества присутствующих солей.
Солёность моря определяется, как общее количество минеральных
веществ в 1 кг морской воды, т.е. сухой остаток. Солёность выражается в граммах
на килограмм морской воды (одна тысячная часть или промилле обозначается знаком
‰).
Изучение солевого баланса в том, или ином водоёме возможно,
если в нём установилось динамическое равновесие во всех его системах.
Современный уровень солёности вод Чёрного моря установился примерно около трёх
тысяч лет тому назад. Солевой баланс Чёрного моря определяется речным стоком,
водным обменом с другими морями, переносом и перемешиванием. Большую часть
речного стока (около 200 км3) приносит в него Дунай. Через
Керченский пролив в Чёрное море с поверхностным течением поступает ежегодно
около 53 км3 слабосолёных вод Азовского моря. Через тот же пролив с
глубинным течением из него выносится около 30 км3 более солёных вод.
Чем больше соли растворено в воде, тем она тяжелей. Поэтому, когда вытекающая
из Чёрного моря лёгкая вода с низкой солёностью сталкивается в Босфоре с
тяжёлой океанической водой, она поднимается над ней. Часть тяжёлой
мраморно-морской воды по дну Босфора стекает в Чёрное море, повышая солёность
его глубин. Этот поток называют Босфорским противотечением, оно в два раза
слабее основного, поверхностного, течения, направленного в Мраморном море.
Солёность поверхностных вод в центральной части Чёрного моря
составляет в среднем 18‰, в некоторых случаях она превышает 18,2‰ и даже
достигает 18,5‰. Сезонные колебания солёности в этой части моря малы. В
весенне-летний период минимум её достигает 17,5‰. Вблизи берегов под влиянием
речных вод солёность уменьшается, сезонные колебания её здесь значительны. В
большинстве случаев средняя солёность прибрежных вод изменяется от 16 до 17‰;
самая низкая солёность наблюдается на северо-западе (от 13 до 15‰,) где в море
поступает основное количество речной воды (Экба, Дбар, 2007).
С увеличением глубины солёность Чёрного моря увеличивается.
На глубине 200 м солёность составляет 20‰, а у дна моря, на глубине 2 км она
может достигать 30‰.
В Чёрном море во все сезоны наблюдаются три района с
пониженной солёностью вод:
) приустьевое - прибрежное от Приморского до косы Тендра
(здесь речной сток составляет 80% суммарного стока всего моря); прикерченское -
с притоком азовских вод с пониженной солёностью; прикавказскоге - прибрежного
от Туапсе до Батуми, где реки приносят более 13% объёма стока рек всего
бассейна (Гидрометеорология, 1992).
Солёность этих районов испытывает сезонные колебания,
синхронные с изменениями речного стока; от начала зимы к весне в приустьевом
районе отмечается постоянное снижение солёности вод с минимумом в мае, когда
проходит в среднем пик паводка.
1.8
Температурный режим
Температурный режим Чёрного моря типичен для водоёмов
умеренной зоны. Зимой поверхностные воды его северо-западного и
северо-восточного районов охлаждаются до нуля, и в мелководных участках шельфа
и в бухтах образуется ледовый покров. Глубоководные районы моря имеют зимой
температуру 5-9°. Летом поверхность моря прогревается до 24-27° у берегов и до
21-23° в глубоководных районах (Скопинцев, 1974, цит. по Экба, Дбар, 2007).
Наименьшая средняя месячная температура поверхностного слоя
воды в прибрежной зоне на большинстве станций наблюдается в феврале, что в
целом согласуется с общим ходом теплового баланса. Именно в феврале суммарное
выхолаживание моря достигает максимума практически по всем районам. Однако,
соотношение теплопотерь моря и значений температуры воды не всегда согласуются,
что связано с поступлением более тёплых вод, приносимых основным черноморским
течением и особенностями морфометрии прибрежных районов.
Самый тёплый район Чёрного моря - его юго-восточная часть.
Кавказские горы
не пропускают в этот район северные ветры, кроме того, здесь
проходит прибрежное течение северо-западного направления. Средняя годовая
температура поверхностной воды Чёрного моря у Кавказского побережья колеблется
в пределах 16,5-18° С. В открытых частях моря амплитуда температурных колебаний
значительно меньше, чем у побережий: зимний минимум поверхностной воды 6,6° С,
летний максимум составляет 27° С (Экба и др., 2007).
Годовые колебания температуры охватывают поверхностный слой в
150 м, а глубже температура остаётся почти неизменной и составляет 8-9° С.
Характерной особенностью термического режима прибрежной зоны
в летнее и осеннее время являются резкие и быстрые непериодические понижения
температуры поверхностных слоёв под воздействием сгонных ветров. Амплитуда
колебания температуры в таких случаях может достигать 10-15° С в течение суток
и даже за несколько часов. Нагонные ветры вызывают скопление тёплой воды у
берега и увеличение толщины прогретого слоя. Наименьшие зимние температуры воды
во всех пунктах Абхазии (за исключением Гагры). Минимальная февральская
температура наблюдается в очамчирской акватории и составляет 6,4° С,
максимальная - в гагрской акватории (9,2° С).
1.9
Концентрация растворённого кислорода
Наблюдения за содержанием растворённого кислорода в
поверхностном слое Чёрного моря, выполненные в юго-восточной части черноморской
акватории показали, что в осенний сезон средние концентрации кислорода
изменялись в пределах 8,86-11,23 мг/л, в летний - в пределах 6,66-9,02 мг/л, в
весенний - в пределах 7,22-9,59 мг/л, в зимний сезон - в пределах 8,97-13,49
мг/л (Экба и др., 2007).
Кислород проникает в воду через поверхность моря из воздуха,
а также образуется в верхнем освещённом слое воды (фотическая зона) при
фотосинтезе водорослей планктона. Для того, чтобы кислород попал в глубины,
море должно перемешиваться - за счёт волн и вертикальных течений. В Чёрном море
вода перемешивается очень слабо; нужны сотни лет, чтобы вода с поверхности
достигла дна.
Наибольшего размаха колебания средних абсолютных значений
содержания кислорода (4,52 мг/л) достигали в зимний период, что связано с
неравномерностью охлаждения поверхностного слоя воды по акватории моря в этот
сезон. В летний сезон размах колебаний средних концентраций по акватории моря
уменьшается до 2,34 мг/л. Наблюдается сезонная изменчивость концентрации
кислорода, увеличение их в осенне-зимний период и уменьшение в весенне-летний.
Наибольшие насыщения кислородом - зимой, наименьшие - летом. Из анализа
сезонного распределения кислорода в период 1998-2002 следует, что максимальное
значение концентрации кислорода наблюдалась в марте и составляет 11,48 мг/л,
минимальная концентрация кислорода приходится на август месяц и равна 8,35
мг/л. Из этого следует, что между концентрацией растворённого кислорода и
температурой существует обратная зависимость. Наименьшие значения температуры
воды приходится на зимние месяцы, в том же сезоне наблюдаются наибольшие значения
растворённого кислорода.
Колебания средних годовых концентраций кислорода лежат в
пределах 8,49-9,97 мг/л, минимальных в пределах 10,53-20,01 мг/л.
Распределение концентрации растворённого кислорода в основном
зависит от процесса фотосинтеза и температуры воды.
1.10
Биогенные элементы
Вертикальное распределение биогенных элементов (фосфатов и
кремнекислоты) в Чёрном море характеризуется минимальным содержанием в слое
фотосинтеза (0-50 м): концентрация фосфатов 0,16-0,45 мкг-ат/л, кремнекислоты
52-60 мкг-ат/л. С увеличением глубины происходит её постепенное повышение.
Содержание фосфатов в аэробной зоне ниже лсоя минимума достигает 4,3 мкг-ат/л,
кремнекислоты 120 мкг-ат/л.
Внутригодовые изменения концентрации биогенных веществ в слое
фотосинтеза носят ярко выраженный сезонный характер. В феврале в поверхностном
слое на большей части акватории Чёрного моря концентрация фосфатов изменяется в
пределах 0,25-0,30 мкг-ат/л.
Концентрация кремнекислоты максимальна на шельфе в
Северо-западном районе и составляет 30-45 мкг-ат/л. В центральной части шельфа
Северо-западного района и в Юго-восточном районе она составляет 14 мкг-ат/л. На
остальной акватории содержание кремнекислоты не превышает 10 мкг-ат/л.
В мае на всей акватории Чёрного моря наблюдается значительное
снижение концентрации биогенных веществ, вследствие интенсивной вегетации
фитопланктона. На большей части акватории моря концентрация фосфатов изменяется
от 0 до 0,15 мкг-ат/л. На западе Северо-западного района и в юго-восточной
части моря она несколько выше и достигает 0,2-0,3 мкг-ат/л.
Общей тенденцией в распределении кремнекислоты весной
является её постепенное уменьшение от 25 мкг-ат/л на севере Северо-западного
района до 5 мкг-ат/л в южной его части. В августе содержание фосфатов в слое
фотосинтеза примерно такое же как, как и весной. На большей части акватории
Чёрного моря оно составляет 0,10-0,20 мкг-ат/л. В Северо-западном районе
максимальная концентрация фосфатов наблюдается на севере района, минимальная -
на западе. В восточной части моря концентрация фосфатов изменяется в пределах
0,15-0,20 мкг-ат/л.
Концентрация кремнекислоты на большей части акватории моря в
августе не превышает 5 мкг-ат/л и лишь на севере Северо-западного района и в
прибрежной части в Юго-восточном районе она повышается до 10 мкг-ат/л.
Распределение кремнекислоты аналогично распределению фосфатов. В приустьевых
районах концентрация кремнекислоты максимально, в восточной половине моря
содержание фосфатов около 10 мкг-ат/л, на остальной акватории 5-10 мкг-ат.
Таким образом, в весенний и летний сезоны наблюдается
значительное уменьшение содержания биогенных веществ, в слое фотосинтеза
вследствие потребления их фитопланктоном. С ноября по март происходит
восстановление биогенной базы биологической продуктивности. В многолетнем плане
наблюдается тенденция снижения концентрации биогенных элементов во всём слое
фотосинтеза Чёрного моря.
За последние 20 лет концентрация фосфатов снизилась в 2 раза,
а кремнекислоты - почти в 10 раз. Исключение составляют приустьевые районы
Днестра, Днепра и Дуная, где в результате значительного выноса биогенных
веществ с речными водами концентрация фосфатов возросла в 2-4 раза. Кроме того,
следует отметить, что со второй половины 70-х гг. наблюдается увеличение
случаев нулевых концентраций фосфатов на горизонтах 100-150 м.
1.11 Нефтяное
загрязнение абхазской акватории Черного моря
По различным оценкам из почти 110 тыс. т нефтепродуктов,
которые ежегодно поступают в Черное море, 48% из них выносится Дунаем. Вместе с
тем, приведенных в документе данных явно недостаточно, чтобы делать однозначные
выводы о роли различных черноморских стран в загрязнении моря нефтяными
углеводородами, так как практически отсутствует информация о количестве
загрязняющих веществ, выносимых с территории Грузии, недоучтены данные по Турции,
Румынии, России. Наиболее полно представлены сведения о "вкладе"
Украины в загрязнение Черного моря. Так, ежегодно с её территории в акваторию
моря поступает 38,3 тыс т нефтепродуктов, что составляет 66,7 % от общего
загрязнения моря всеми черноморскими странами. Причем, главными источниками
загрязнения являются бытовые и промышленные стоки, в меньшей степени - реки.
Основными источниками загрязнения, поверхностных вод абхазской акватории
Черного моря являются: городская канализация, автотранспорт, АЗС. Наибольшее
количество нефти с суши поступает в море по рекам вместе с грязью, смываемой с
городских территорий. Также источником загрязнения поверхностных вод Черного
моря является морской транспорт и основное черноморское течение.
Наблюдения за загрязнением морской акватории Абхазии
нефтепродуктами проводится с 1998 года (Экба, Дбар, Гицба, 2004). Содержание
нефтепродуктов в морской воде определяется гравиметрическим способом (ПДК
составляет 0,05 мг/л).
Наблюдения за содержанием нефтеуглеводородов (НУ) в морской
воде проводились с 1998 по 2005 гг. в следующих пунктах прибрежной зоны
Сухумской бухты: порт, район Центральной спасательной станции, морской вокзал,
медицинский пляж (Синоп), м. Маяк. Пункты наблюдений выбраны исходя из степени
загрязненности в результате антропогенной деятельности. Сухумская бухта (район
Центральной спасательной станции и мор. вокзал, т.е. промышленная зона бухты)
наиболее сильно подвержены загрязнению сточными водами и морским транспортом.
Точки отбора проб Медицинский пляж (Синоп) и м. Маяк находятся в
противоположных концах города и наименее подвержены антропогенному воздействию.
Распределение НУ в 1998 и 1999 гг. определялось также на
некотором расстоянии от берега в центральной части акватории Сухумской бухты
(Экба и др., 2004). Наблюдения за распределением НУ в разных прибрежных пунктах
Сухума проводившиеся в 2001-2002 гг. (Дбар и др., 2001; Экба и др., 2003; Дбар
и др., 2005) показывают, что наименьшая средняя межгодовая концентрация НУ
наблюдается в водах медицинского пляжа (0,13 мг/л). Наибольшая - концентрация
НУ наблюдается в районе Центральной спасательной станции г. Сухум (0,25 мг/л).
Распределение нефтепродуктов рассматривалось кроме Сухумской
бухты также в некоторых других пунктах побережья Абхазии. Наименьшее количество
нефтеуглеводородов наблюдается в водах центрального пляжа п. Пицунда (1 ПДК).
Наибольшее количество НУ концентрируется в устье реки Гудоу (г. Гудаута),
максимальное количество которого приходится на август (20 ПДК), минимальное на
октябрь - 4 ПДК (Экба и др., 2004).
Наибольшие концентрации наблюдаются в летние месяцы, а
наименьшие в осенние. Максимальное значение НУ наблюдается в июне и составляет
29,8 ПДК, минимальное - в ноябре (4,8 ПДК). В летний период, по-видимому,
происходит наибольшее загрязнение акватории морским транспортом в связи с его
интенсивным использованием в курортный сезон. Значение концентрации НУ также
существенно зависят от метеоусловий: сгонных и нагонных ветров.
Пространственное распределение НУ в морской акватории Сухума
показывает, что наибольшая средняя концентрация наблюдается у береговой черты -
5 ПДК, наименьшая концентрация в более отдаленной от берега части, т.е. в 5 км
от берега и составляет 2,4 ПДК (Экба и др. 2004).
За весь период наблюдений наибольшая концентрация нефтеуглеводородов
приходится на 1998г. (среднее значение составляет 10 ПДК). В последующем
количество нефтеуглеводородов уменьшается и достигает минимального значения в
2002 г. (2,8 ПДК). В промежутке с 1982 по 1986гг. минимальное значение
наблюдалось в 1983-1984гг.0,14 мг/л), максимальное - в 1985 г. (0,35 мг/л).
Исходя из анализа представленного материала, можно сделать
вывод, что четко выраженного сезонного распределения НУ в акватории Черного
моря не наблюдается (Экба и др., 2004). Однако, чаще всего, максимальные
значения НУ у побережья Абхазии отмечаются в феврале-марте, июне-июле и
октябре-ноябре. Минимальные концентрации наблюдаются зимой - декабрь-январь и
весной - в апреле.
Глава 2.
Методы исследования планктона
2.1 Орудия
для сбора планктонных организмов
В зависимости от размеров планктонные организмы принято
делить на следующие категории:
мегалопланктон ― организмы, размеры
которых больше 5см;
макропланктон включает в себя организмы длиной от 5мм до 5см;
мезопланктон - организмы, видимые простым глазом, размеры их
варьируют от 0,5мм до 5мм;
микропланктон - организмы микроскопические, не проходящие
через отверстия самого плотного шелкового газа (№ 25/77); их размеры от 50мкм
до 0,5мм.
наннопланктон - организмы, свободно проходящие через
отверстия самого плотного шелкового газа и потому не улавливаемые планктонными
сетями, их размеры варьируют от 5 до 50мкм (Приложения, рис.4).
пикопланктон ― мельчайшие организмы,
длина которых меньше 5мкм (Константинов, 1986).
В связи с широким разнообразием размеров планктёров для их
отлова применяют различные методы с использованием разных орудий.
Все методы, обычно применяемые в практике планктонных
исследований, могут быть сведены к следующим категориям:
) сетной или сетяной метод, при котором улавливаются мезо - и
микропланктон;
) метод химического осаждения или отстойный метод;
) метод механического осаждения или центрифужный метод;
) метод плотных (мембранных) фильтров;
) камерный метод;
) метод, представляющий собою комбинацию камерного и
отстойного методов.
При исследовании планктона следует помнить о тесной связи
организмов с внешней средой, о единстве организма и среды, поэтому изучение
планктона должно производиться одновременно с учетом факторов водной среды
(температуры, света, течений, растворенных в воде газов, рН, минеральных солей,
N, P, Fe, органических веществ и пр.). Только изучение планктона в тесном
комплексе с другими гидробиологическими работами на водоеме даст возможность
понять те факторы, от которых зависит состав, распределение, периодичность,
количественное развитие планктона и прочие стороны его жизнедеятельности, и
выявить те влияния, которые сам планктон оказывает на окружающую его водную
среду.
Методы сбора планктона в большинстве случаев слагаются из
двух процессов:
а) взятия (зачерпывания) пробы воды, содержащей планктон и
доставки ее на поверхность;
б) отделения (отцеживания) планктона от воды. Оба эти
процесса могут осуществляться одновременно или отдельно в зависимости от
конструкции используемой аппаратуры.
Самым распространённым и простым методом сбора планктона
является метод сетной, при котором вода, содержащая планктон, фильтруется через
особую сетку из шёлкового газа, пропускающего воду и задерживающего организмы
планктона. Планктонные сетки могут быть качественные и количественные.
Качественными называют сетки с большим отверстием и малой фильтрующей
поверхностью (т.е. малой длиной сетки). Они дают представление о составе
планктона, но не годятся для определения его количества. У качественных сеток
делают не только маленькое входное отверстие, но и газовый конус длиной не
менее 2-3м.
Универсального метода, одинаково пригодного для всех
категорий организмов, для всех типов водоемов, независимого от места, от
времени сбора и обработки материала, отличающегося простотой и удобством,
требующего минимальной затраты времени и труда, лишенного потерь и дающего
надежные, точные, близкие к реальным количественные показатели,
удовлетворяющего всем задачам исследования, такого метода нет и быть не может.
Поэтому в дальнейшем дается описание различных методов сбора и обработки
планктона с указанием присущих каждому из них преимуществ и недостатков и
степени пригодности каждого в тех или иных конкретных случаях.
2.1.1
Качественная сеть Апштейна
Качественная планктонная сетка Апштейна состоит из латунного
кольца и пришитого к нему конической формы мешка из шёлкового газа,
заканчивающегося внизу стаканчиком, в котором собирается осадок планктона. Для
изготовления планктонной сетки употребляют так называемый мельничный газ,
отличающийся прочностью и равномерностью размеров его отверстий (ячей).
Существует много разных номеров такого газа, причём в русской нумерации номер
газа соответствует количеству отверстий, приходящийся на 10 мм.
Так как размеры планктонных организмов разные, то для более
полного сбора всех категорий организмов приходится употреблять сетки из газа
разных номеров. Для микропланктона №17/64-25/77, для мезопланктона №
12/49-14/55, для крупных ракообразных № 3/23-8/34.
Сетяной конус изготовляется по выкройке, вырезаемой из бумаги
и равной боковой поверхности развёрнутого конуса или половины её (Приложения,
рис.1). Для сшивания берутся тонкие иголки и тонкие прочные нитки. Конус из
шёлкового газа прикрепляется к металлическому кольцу не непосредственно, а при
помощи узкой полоски из плотной материи, ширина которой вместе с образующей
шёлкового конуса и должна равняться общей длине образующей боковой поверхности
усечённого конуса. Нижний, обшитый плотной материей конец конуса прикрепляется
к стаканчику с помощью плоского латунного кольца, снабжённого зажимным винтом;
если же стаканчик с утолщением у края, то нижний конец привязывается к
стаканчику с помощью нескольких оборотов прочной бечевки.
К металлическому кольцу на равном расстоянии друг от друга
прикрепляются 3 прочные бечевки, свободные концы которых связываются узлом над
входным отверстием сетки, или привязываются к небольшому колечку, к которому
присоединяется при помощи чекеля петля или колечко крепкого пенькового или
металлического троса, служащего для спуска сетки. Во избежание того, чтобы
шёлковый газ не подвергался разрыву от тяжести плохо фильтрующейся воды, веса
стаканчика и груза, служащего для отяжеления сетки, рекомендуется бечевки, привязанные
к металлическому кольцу.
Стаканчики для планктонных сеток бывают разной конструкции.
Особенно удобны металлические стаканчики с краном (Приложения, рис.2). Их
размеры: высота 40 мм, диаметр 28 мм, для средней модели сетки соответственно -
80 мм и 50 мм. Не менее удобен металлический стаканчик с глухим дном без крана,
состоящий из двух половин: короткой передней и более длинной задней,
соединённых друг с другом посредством штыкового затвора или винтовой нарезки.
Можно сделать стаканчик из алюминия в виде воронки с небольшой закраиной или
кольцевым утолщением на переднем конце для прикрепления его к сетке. Для
небольшой закидной сетки удобны стаканчики из толстого стекла разного объёма и
формы, снабжённые у входного отверстия закраиной или кольцевым утолщением для
прикрепления к нему нижнего конца сетного конуса, а внизу оттянутые в короткую
трубку, на которую надевается кусок резиновой трубки с зажимом Мора или которая
просто закрывается пробкой. Чтобы не потерять зажима и пробки, последние шнурком
привязываются к кольцевому утолщению стаканчика. Такой стаканчик можно сделать
из химической воронки, имеющей форму шарика с закраиной и с длинной выводной
трубкой, обрезав последнюю до размеров короткого придатка, на который и
надевается резиновая трубка с зажимом.
В такие планктонные сетки внутрь в раструб вставлены
напоминающие спидометр приборы.
По показанию приборов можно определить, какое количество воды
прошло сквозь сеть, и затем рассчитать численность организмов в единице её
объёма.
Современные планктонные сети имеют сложное устройство: они
состоят из нескольких сачков, прикреплённых к кабелю на различных уровнях;
глубина погружения регистрируется специальным прибором, и когда она достигает
нужной отметки, устье сетей по поступающей с судна радиокоманде открываются. По
истечении установленного времени другой сигнал закрывает их, и вся система
поднимается на поверхность. Это позволяет собирать образцы планктона с разных
глубин.
2.1.2
Планктонособиратель
Помимо сеток, которые протягивают на большие расстояния, для
получения данных по уровню развития планктёров, используются
планктонособиратели ― большие цилиндры
(ёмкостью 25-50 л), стенка которых сделана из сетного газа, а сверху и снизу
имеются две металлические крышки (Приложения, рис.3).
Прибор опускается в открытом виде: сетка спущена к нижней
крышке. Опустив планктонособиратель на нужную глубину, по тросику посылают
гирьку, которая, ударив по верхней крышке прибора, приводит в действие особый
механизм. Сетка подтягивается к верхней крышке и задёргивает прибор с боков.
Когда прибор поднят на палубу, содержимое его выпускают в банку через кран,
находящийся в центре нижней крышки. Преимущество планктонособирателя в том, что
он даёт возможность точно учесть количество планктонных организмов в определённом
объёме воды (Пряхин и др., 2006).
2.1.3
Батометр
Батометр представляет собой прибор, с помощью которого берут
пробы для химического анализа воды и сбора наннопланктона осадочным методом. На
батометре укреплены два опрокидывающихся глубоководных термометра. Батометр
(или их серия) укрепляется на соответствующих расстояниях на тросе и с помощью
специальной глубоководной лебедки погружается на необходимую глубину.
Так как краны батометра при спуске открыты, вода свободно
проходит через цилиндры приборов. Через некоторое время, необходимое для
отражения термометром температуры окружающей среды, с борта корабля по тросу
спускается специальный сигнальный или посыльный грузик. Ударом грузик
освобождает защелку у верхней подвески, и батометр опрокидывается, поворачиваясь
на оси у нижнего основания рамы. При этом краны закрываются, и вода данного
уровня оказывается запертой в батометре.
Одновременно с этим в опрокидывающемся термометре, в месте
сужения капилляра, у резервуара, происходит отрыв столбика ртути, вошедшего в
капилляр и соответствующего измеряемой на данной глубине температуре. При
опрокидывании первого батометра от нижней части отделяется установленный там
грузик, который, следуя по тросу вниз, обеспечивает опрокидывание второго
батометра. И так же последовательно опрокидывается вся серия приборов (Пряхин и
др., 2006).
Так называемый осадочный метод для сбора наннопланктона
заключается в следующем. Взятый с определённой глубины при помощи
гидрологического батометра образец воды в 1 литр целиком переливается в большую
бутыль и фиксируется формалином. Затем все организмы, в том числе и самые
мельчайшие, оседают на дно, после чего их осторожно выбирают стеклянной
пипеткой, переносят в баночку, определяют и пересчитывают.
2.2
Консервирование и этикетирование проб планктона
Проба планктона, взятая тем или иным способом, если она не
исследуется сразу же в живом виде, должна быть законсервирована. Самой
распространенной консервирующей жидкостью, пригодной для сохранения как
зоопланктона, так и фитопланктона, служит формалин.40% -й формалин прибавляется
к улову с таким расчетом, чтобы получился 2% -й (зимой) или 4% -й (летом)
раствор.
Другим консервирующим средством, но пригодным только для
зоопланктона, служит спирт, крепость которого для сохранения планктона в нём
должна быть не менее 70°. Из других фиксаторов можно указать на водный раствор
йода в йодистом кали (раствор Люголя), жидкость Флемминга.
Каждая проба снабжается этикеткой. Этикетку надо чётко писать
на узкой полоске бумаги или пергамента и вкладывать внутрь банки. В пробах с
осадочным планктоном следует прищемить чистый конец этикетки пробой.
Запись на этикетке следующая:
№ пробы
№ станции
Дата
Название водоёма
Место сбора
Глубина
Орудие лова
Продолжительность лова
Подпись
Очередной номер надо повторить на пробке чернильным
карандашом. Этикетки, написанные карандашом, заменяются постоянными,
написанными несмывающейся тушью. Необходимо вести журнальную запись проб, куда
записывают всё, что потребуется при обработке проб и оформлении работы:
состояние погоды, данные по прозрачности, глубине водоёма, скорости течения,
химизму воды, характер лова, продолжительность работы и др.
Пробы хранятся в порядке номеров в защищённом от света месте.
Пробы заливаются парафином или смесью воска и парафина, причём перед заливкой,
в случае необходимости пересылки проб, они доливаются до пробки
профильтрованной водой.
2.3
Качественная обработка проб
Методы обработки планктона могут быть качественные и
количественные, могут производиться над планктоном как в живом, так и в
консервированном виде.
При качественной обработке в живом виде собранный материал
следует исследовать вскоре после лова или на месте взятия пробы, или на
ближайшем стационарном пункте (лаборатории, станции и пр.). Ввиду быстрого
отмирания планктона при переносе проб на место обработки, следует защищать
пробы от нагревания солнцем, не закупоривать их наглухо, уложить их во влажный
мох или завернуть в мокрую тряпку.
Исследование планктона в консервированном виде производится
следующим образом: часть осадка берется пипеткой (не той, которая употребляется
при изучении планктона в живом виде) и переносится на предметное стекло,
накрывается покровным стеклом и исследуется при малом увеличении микроскопа или
под бинокуляром для зоопланктона и при большом увеличении для фитопланктона
(также для всего наннопланктона). Пипетка для взятия капли не должна быть
слишком узкой. При определении отмечается на глаз частота встречаемости
отдельных форм, которая в записях регистрируется теми или иными условными
обозначениями по пяти-шестибалльной шкале (Бурнева, 2001).
Для оценки встречаемости устанавливается такая шкала:
). Не в каждом препарате - обозначается дробью, числитель
которой показывает количество найденных экземпляров, а знаменатель число
препаратов.
). В каждом препарате в количестве не более 10 экземпляров -
обозначается цифрой 1.
). В каждом препарате в количестве 10 экземпляров, но не в
каждом поле
зрения - цифрой 2.
). Н каждом поле зрения в количестве от 1 до нескольких
экземпляров - цифрой 3.
). В массе - знаком ∞
.6). Если в препарате организмов 2-3, то при преобладающем из
них после ∞ ставится восклицательный знак (!) (Бурнева, 2001).
2.4
Количественная обработка проб
Количественные методы обработки планктона довольно
разнообразны и тоже могут производиться над планктоном, как в живом, так и в
консервированном виде. Самым старым и очень трудоемким является счетный метод.
При этом методе наиболее крупные организмы просчитываются целиком, удобно
использовать для этого камеру Богорова. В большинстве же случаев для подсчета
всех групп планктона, собранного мелкоячеистой сетью, необходимо ограничиться
подсчетом небольшой порции планктона, взятой микропипеткой.
Самую пробу планктона перед взятием из нее порции для
подсчета доводят до определенной концентрации, разбавляя ее, если она слишком
густа (количество просчитанных организмов в порции больше 1000), или сгущая ее
путем отсасывания воды, если она очень бедна (количество организмов в порции
меньше 100). Объем пробы, доведенной до требуемой концентрации, измеряют. Перед
взятием из нее порции пипеткой пробу взбалтывают в особой щирокогорлой банке и,
не дав организмам осесть, быстро погружают в пробу нижний конец пипетки и с
помощью ее берут порцию - 0.1 мл для фитопланктона и от 0.2 до 1 мл и даже до 5
мл для зоопланктона. Если взята порция в 0.1 мл, то ее накрывают большим
покровным стеклом (24 х 32 мм) и просматривают от края до края
последовательными движениями крестообразного столика микроскопа. Просчитывают
две такие порции. Если расхождение результатов двух просчетов не больше 5 %, на
этом просчет и заканчивают; если же расхождение более 5%, необходимо взять для
просчета еще 1-2 порции, пока расхождение не снизится до 5%. В случае
отсутствия микропипетки ее можно заменить обычной градуированной пипеткой.
Для сокращения времени просчета при очень больших продукциях,
в целях получения лишь сравнимых результатов можно ограничиться просчетом
организмов не во всей порции в 0.1 мл, а в пределах нескольких полей зрения.
Для этого сначала определяется площадь поля зрения, подсчитывается количество
организмов в пределах одного поля зрения, произвольно взятого, потом другого и
т.д. до 10-20 полей. Полученные числа суммируются и результат сложения делится
на число просчитанных полей зрения. Полученное частное будет представлять собою
среднее количество организмов в одном поле зрения. Определив, сколько таких
полей зрения содержится во всей порции площадью в 24х32 мм, и помножив на
найденный коэффициент среднее количество организмов в одном поле зрения, мы
получаем число, показывающее, сколько организмов содержится в 0.1 мл; помножив
еще на 10, получаем количество организмов в 1 мл, а, помножив на объем всей
пробы, откуда бралась порция для подсчета, определяем, сколько организмов
содержится во всей пробе воды, т.е., в случае сетного сбора, во всем столбе
воды, профильтрованном через сетку. Подсчет планктона, отфильтрованного через
мембранный фильтр или осажденного отстойным методом, лучше производить в
счетных камерах определенного объема и толщины.
Для проведения более точного подсчета мелких организмов под
микроскопом в большинстве случаев бывает безусловно необходимо разделить поле
зрения на части, что обычно достигается применением разграфленных счетных
пластинок. Введенный широко в американскую практику сетчатый окуляр-микрометр и
в некоторой модификации используемый и в России, хотя и бывает часто вполне
удовлетворителен, но не при всех условиях. Лучшие результаты получаются при
использовании так называемых счетно-полосчатых окуляров с параллельно
натянутыми в поле зрения двумя нитями, расстояние между которыми подбирается в
зависимости от количества подсчитываемых организмов.
Так как иметь несколько таких окуляров не экономно и не
всегда доступно, лучше приобрести один окуляр, с приспособлением, позволяющим
произвольно менять расстояние обеих параллельных нитей. Для достижения еще
большей точности, неподвижно и несколько эксцентрически натягивается третья
нить, пересекающая поперек обе параллельные нити. При подсчете перемещают
предметное стекло посредством крестообразного столика попеременно в
направлении, параллельном нитям, до конца объекта, потом точно на ширину одной
полоски.
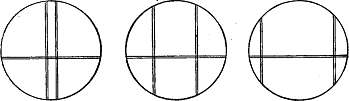
Рис.1. Счетно-полосчатые окуляры с натянутыми нитями.
При обработке проб планктона, собранных сетью из крупноячеистого
шелкового газа, пробу подсчитывают целиком, и только в случае большого
количества организмов ограничиваются просмотром части пробы.
При количественном подсчете планктона очень важно уметь
различать в фиксированном материале живые организмы от мертвых компонентов
планктона. Это различие устанавливается по изменению структуры при отмирании
организмов. Для живых диатомовых водорослей и динофлагеллят характерна целость
протопласта, целиком заполняющего клетку, для мертвых же - нарушение целости
последнего. Для живых сине-зеленых - зернистость клеток и четкость границ как клеток,
так и отдельных колоний, для мертвых же - гомогенность колоний и разбухание
оболочек клеток. У зоопланктона (ракообразных и коловраток) отличительным
признаком живых служат хорошо выраженная мускулатура и четкость границ между
внутренними органами; для мертвых надо отметить - на первой стадии распад
мускулатуры с появлением зернистости и последующее стирание границ между
органами.
От определения числа организмов в пробе переходят к
определению численности.
Данные по численности должны быть представлены, как
количество организмов в единице объёма. Если проба отобрана путем процеживания
объема воды через сеть Апштейна, то расчет производится следующим образом:
= 1000 n / v,
где x - количество организмов в 1 м3 воды, экз/м3;
n - количество организмов в пробе, экз.; v - объем воды, процеженной через
сеть, л.
Если отбор проб произведен количественной сетью Джеди, то
прежде всего рассчитывают коэффициент планктонной сети (или множитель перевода
в м), исходя из радиуса ее входного отверстия. Коэффициент сети рассчитывается
следующим образом:
= 1000000/ (S H),
где S - площадь входного отверстия сети, cм2, H - горизонт,
слой облова, см. Вычислив таким образом коэффициент сети при горизонте облова
0-1 м, находим коэффициенты при горизонтах 0-2, 2-5, 5-10 м и т.д. простым
делением значения k при 1 м соответственно на 2, 3 и 5. Численность организмов
N находится путем перемножения количества организмов в пробе n на коэффициент
сети k. Следующим этапом количественной обработки проб планктона является
получение данных по биомассе. Биомасса планктона определяется путем умножения
индивидуальной массы каждого организма на его численность. Однако следует
учитывать, что длина и масса, например, зоопланктеров одного и того же вида
может значительно варьировать в разных водоемах, климатических зонах, а также в
зависимости от сезона. Метод определения массы организмов путем
непосредственного взвешивания очень трудоемок. Поэтому уже достаточно
продолжительное время широко используется способ, при котором учитывается
соотношение между массой и длиной тела особи. Однако многочисленные данные,
опубликованные в литературе, часто плохо согласуются между собой. Это
объясняется недостаточным количеством данных и погрешностями методик. Е.В.
Балушкина и Г.Г. Винберг сопоставили и критически оценили все содержащиеся в
литературе уравнения и материалы, позволяющие по измерениям длины тела находить
массу планктонных животных. В результате было предложено в качестве общего
способа выражения зависимости между массой и предложено в качестве общего
способа выражения зависимости между массой и длиной тела особи уравнение:
= g lb,
где w - масса тела, мг; l - длина тела организма, мм; g -
масса тела при длине тела 1 мм, мг сырого вещества; b - показатель степени.
Биомассу планктона рассчитывают по формуле:
B=N×W, где
N - число организмов (экз/м3);
W - индивидуальный вес (мг/м3).
В заключение заметим, что статистика, базирующаяся
исключительно на количественном подсчете числа особей, не всегда может быть
удовлетворительной, так как, благодаря различным размерам планктонных форм, она
не может дать картины фактической продукции; поэтому является необходимость
определения объемов или весов планктонных организмов. Однако и определение
одних объемов или весов без учета численности организмов часто не дает
представления о степени относительного развития, например наиболее мелких и
наиболее крупных по объему или весу видов, что имеет большое значение для
многих вопросов планктонологии. Поэтому в диаграммах распределения планктона,
сконструированных на основе объемов, следует вписывать количества особей в 1
литре.
Глава 3.
История изучения планктона Чёрного моря
История человеческого общества неразрывно связано с морем.
Ещё на заре своего существования человек селился у берегов морей и океанов и
находил здесь обильную легко добываемую пищу. Изучение Чёрного моря началась
ещё в античное время, вместе с плаваниями греков, основавших на берегу моря
свои поселения. Людей издавна интересовал таинственный и загадочный мир моря.
Более серьёзное изучение флоры и фауны Чёрного моря началось
с конца XVIII века. С этого времени русские учёные приезжали на берега Чёрного
моря и, проводя на них некоторое время, изучали различные организмы. Эти
научные поездки были начаты академиком П. Палласом (1793-1794) и продолжены Э.
Эйхвальдом и К. Кесслером и многими другими. Для изучения фауны Чёрного моря
важную роль сыграли экспедиции Н. Андрусова, А. Остроумова, Ш. Шпиндлера и
Лебединцева, проведённые в 90-х годах XIX века, открывшие явление заражения
глубинных слоёв моря сероводородом (Зенкевич, 1956).
Все эти экспедиционные исследования конечно имеют важное
значение для изучения фауны Чёрного моря и планктона в частности, но в силу
того, что они носили кратковременный характер эти экспедиции не могли в полной
мере выявить всё многообразие живых организмов, населяющих Чёрное море.
Начало систематическим научным исследованиям Чёрного моря
положили два события конца XIX века - изучение Босфорских течений (1881-1882) и
проведение двух океанографических глубиномерных экспедиций (1890-1891).
Существенный вклад в изучение планктона Чёрного моря, и в
целом его фауны и флоры внесла, созданная в 1871 г. Севастопольская
биологическая станция (ныне Институт биологии южных морей). Это было крупное
самостоятельное учреждение, ведущее работу круглый год, имеющее свой
самостоятельный персонал, оборудованные лаборатории, обширные библиотеки,
морской аквариум, плавучие средства, и всё это на берегу моря, благодаря чему
постоянно имелась возможность получить в неограниченных количествах свежий материал
для исследований.
Работа Севастопольской биологической станции по изучению
флоры и фауны Чёрного моря приобрела более систематичный и регулярный характер.
Особенно успешными они стали с начало XX века, когда эту станцию возглавил
известный русский гидробиолог академик Зернов.
Кроме Севастопольской биологической станции, большой вклад в
изучение планктона Чёрного моря внесли такие учреждения, как Карадагская и
Новороссийская биологическая станции, Азово-черноморский институт рыбного
хозяйства в Керчи, Батумская рыбохозяйственная станция и др. (Зенкевич, 1956).
В это же время были созданы и другие биологические станции, такие, как
Солевецкая биологическая станция на Белом море и т.д.
В целом, советский период в изучении морей во всех областях
океанологии был чрезвычайно плодотворным. Всё это способствовало тому, что в
60-х годах прошлого столетия моря СССР были безусловно наиболее изученными
морями мира. Основной характерной особенностью советской океанологии являлось
комплексное изучение моря, т.е. всестороннее исследование явлений,
развёртывающихся в морских водоёмах. Применительно к биологическим явлениям
комплексность заключалась в охвате всего ряда явлений - от цикла биогенных
веществ, определяющих первичную продукцию, через все звенья пищевых цепей до
конечных их звеньев - промысловых рыб и морских млекопитающих.
В 60-70-х годах XX века огромный вклад в развитие
планктологии Чёрного моря внесли учёные: Сорокин, занимающийся зоопланктоном;
Павловская, исследующая микрозоопланктон; Павлова, которой принадлежат труды по
мезозоопланктону; Виноградов, Туманцева, Шушкина, Зайка, Павельева и др. Уже с
80-х годов прошлого столетия активное исследование планктонных организмов
северо-восточной части Чёрного моря начинает Азовский научно-исследовательский
институт рыбного хозяйства (АзНИИРХ). Сотрудники этого института Надолинский
В.П., занимающийся ихтиологией и, в частности ихтиопланктоном, Мартынюк М.Л.,
исследующий зоопланктон и др. внесли существенный вклад в изучении планктонного
сообщества Чёрного моря. Эти исследования и в настоящее время активно
проводятся АзНИИРХ.
Ныне функционируют и многие другие научно-исследовательские
учреждения, как например, Институт биологии южных морей НАН Украины, которая
активно занимается изучением флоры и фауны Чёрного моря. Все эти исследования
имеют не только теоретическое значение для биологической науки, но и большое
практическое значение, так как помогают решать многие вопросы, касающиеся
рыбохозяйственной деятельности.
Глава 4.
Аннотированный список летне-осеннего фито - и зоопланктона Чёрного моря
4.1
Аннотированный список фитопланктона
В составе фитопланктона Чёрного моря по данным Вершинина
насчитывается примерно 700 видов организмов (Вершинин, 2007).
У берегов Абхазии в результате наших исследований в составе
осенне-летнего фитопланктона были обнаружены и идентифицированы
следующие виды:
1. Сeratium tripos Nitzsch (Приложение, рис.3).
Данный вид относится к динофитовым водорослям (Dinophyta). Представлен в основном
организмами средних размеров. Длина и ширина тела без рогов почти одинаковы,
рога довольно крупные и грубые. Правый задний рог нередко меньше левого, часто
он параллелен переднему рогу. Задние рога изогнуты и направлены относительно
параллельно переднего.
Вид эвригалинный, эвритермный, распространён в холодных,
умеренных и тёплых водах морей и океанов. Вид сильно изменчив в форме и
размерах (Киселёв, 1950).
2. Ceratium furca Ehr (Приложение, рис.4).
Клетки в общем виде похожи на клетки C. tripos. Они примерно
палочковидной формы, передний рог конической формы. Отличительной чертой
данного вида является наличие прямых задних рог средней величины, направленных
параллельно друг другу. Один рог обычно более короткий. Клетки крупные, 360-400
мкм общей длиной, 32-60 мкм шириной (Киселёв, 1950).
3. Ceratium fusus Ehr (Приложение, рис.5).
Клетки веретеновидные или палочковидные, 300-800 мкм общей длиной, 60
мкм шириной. Эпитека высококоническая, постепенно переходящая в длинный
апикальный рог. Поясок кольцевидный. Гипотека короче эпитеки. Тека гладкая или
складчатая, рога с шипами или без шипов. Они более вытянуты, один задний рог
укорочен, второй длинный и прямой. Вид панталласный, возможно космополит
(Киселёв, 1950).
4. Pseudosolenia calcar-avis Schuitze, или псевдосоления
птичья шпора (Приложение, рис.6) - самая крупная планктонная диатомея. Кончики
этой длинной тонкостенной трубки загнуты, как когти птицы. Она может вырасти
длиннее 1 миллиметра - гигант среди фитопланктонных водорослей. Но, так как
стенки её панциря тонкие, а поверхность большая, то плотность клетки
уменьшается и она продолжает парить в пелагиали (Вершинин, 2007).
5. Proboscia alata Brighw (Приложение, рис.7). Общая
морфология данного представителя Diatomea уподобляется морфологии Pseudosolenia calcar-avis Schuitze, однако, при
детальном рассмотрении можно выделить отличительные черты: клетки более мелкие
и узкие, концы менее заострённые. Довольно распространённый вид в
Чёрном море (Забелина и др., 1951).
6. Prorocentrum micans Ehrenberg (Приложение, рис.8) -
представитель отдела Dinophyta. Клетки сильно сжатые с боков, наиболее широкие
почти на середине, с острым задним и тупым, с небольшой выемкой, передним
концом. Спинной край сильно изогнут, чем брюшной. Левая скорлупа несёт
различной длины массивный зубец с тонким крылом (перепонкой). Оболочка с
многочисленными порами (пороидами) и местами с нежными шипами. Два больших
жёлтых пластинчатых хроматофора. Длина 30-52 мкм (без шипа), толщина 20-24 мкм.
Широко распространённый неритический вид, встречающийся в
открытом море. Играет большую роль в питании рыб и моллюсков (Киселёв, 1950).
7. Chaetoceros sp. (Приложение, рис.9).
Клетки одиночные или соединённые щетинками в цепочки. Панцирь тонкостенный,
коротко цилиндрический, иногда с небольшим количеством вставочных ободков,
всегда без септ, с неразличимой структурой. Створки эллиптические, реже почти
круглые, с плоской выпуклой или вогнутой поверхностью, низким или более или
менее высоким загибом и длинной щетинкой на каждом из полюсов створки. В
колонии щетинки соседних клеток в основании перекрещиваются, образуя цепочки с
просветами (окнами) между створками смежных клеток. Хроматофоры в виде
одной-двух крупных пластинок или многочисленных мелких зёрен. У некоторых видов
образуются покоящиеся споры с толстостенным панцырем. Створки споры имеют
различную форму и различную структуру, реже створки гладкие, поясок отсутствует
(Забелина и др., 1951).
В черноморском планктоне встречаются довольно много видов из
данного рода, которых трудно идентифицировать.
4.2
Аннотированный список зоопланктона
В составе черноморского зоопланктона насчитывается примерно
150 различных видов. В составе зоопланктона у берегов Абхазии
нами обнаружены и идентифицированы следующие виды:
1. Penilia avirostris Dana (Приложение, рис.10). Вид
относится к отряду Листоногие раки (Phyllopoda), подотряду Ветвистоусых
(Cladocera). На краях створок по всей длине находятся шиповидные зубчики (около
40 на нижнем и около 10 на нижнем крае), между которыми расположены короткие
щетинки. Голова низкая, у самки вытянута в два направленные книзу выроста с
шипами; у самца снизу округлая. Антенны I у самки короткие, у самца очень
длинные, доходят до конца раковины. Антенны II у основания стебля с шиповидной
щетинкой и двумя 2-членистыми ветвями с восьмью щетинками на экзоподите и пятью
на эндоподите. У самцов позади шестой пары ног пара очень длинных
совокупительных органов. Самцы и самки чрезвычайно прозрачны, самцы бесцветны,
у самок могут быть скопления жёлто-коричневого пигмента. Длина самки 0,8-1,0
мм, самца - 0,65-0,80 мм.
Чисто морской вид. В Чёрном море встречается в тёплое время
года, в наибольшем количестве в августе - сентябре. Осенью (до ноября)
встречаются с латентными яйцами (Мордухай-Болтовский, 1968).
2. Oithona minuta Kricz (Приложение, рис.11, 12.)
является типичным представителем подотряда Cyclopoida, относящийся к семейству
Oithonidae. Тело относительно
узкое, стройное. Цефалоторакс широкий, с наибольшей шириной посредине, абдомен
очень узкий. Генитальный сегмент длинный, по длине равен двум последующим
вместе взятым, анальный сегмент короткий. Фуркальные ветви незначительно
длиннее анального сегмента. Пятая пара ног имеет вид бугорка с двумя концевыми
щетинками. Длина самки 0,5-0,7 мм, самца ― 0,4-0,6 мм
(Мордухай-Болтовский, 1968).
Массовая форма планктона Чёрного и Азовского морей, круглогодичная,
преимущественно поверхностная, эвригалинная.
3. Oithona similis Claus (Приложение, рис.13).
Цефалоторакс у самки одинаково широкий на всём протяжении. Рострум острый,
направлен вниз, со спины не виден. Последний торакальный сегмент маленький,
округлой формы, генитальный сегмент очень длинный, почти равен длине трёх
последующих вместе взятых. Фуркальные ветви такой же длины, как и анальный
сегмент. Мандибулы хорошо развиты. Цефалоторакс самца несколько расширен в
нижнем отделе, рострум отсутствует, абдомен 5-сегментный, генитальный сегмент
слегка расширен, анальный сегмент и фуркальные ветви маленькие. Геникулирующие
антенны самца 14-членистые. Пятая пара ног 2-членистая.
Широко распространенная круглогодичная, но не массовая форма
черноморского планктона. С мая по ноябрь предпочитает более глубокие горизонты,
остальное время держится в поверхностных слоях. Совершает вертикальные
миграции. Размножается в течение всего года (Мордухай-Болтовский, 1968).
4. Paracalanus parvus Claus (Приложение, рис.14) -
представитель подотряда Calanoida из семейства Paracalanidae. Форма тела
удлинённо-овальная, тело коренастое. Строение симметричное у обоих полов.
Головной конец широко-округлый у самки и незначительно выступает переднего края
у самца. Рострум в виде двух тонких нитей. Фуркальных щетинок четыре пары,
почти одинаковой длины. Антенны II с 6-членистым экзоподитом, их концевой членик
очень маленький, экзоподиты короче эндоподитов. Ноги пятой пары у обоих полов
одноветвистые. Самцы светло-жёлтые, иногда с красными пятнышками на голове и
третьем торакальном сегменте. Длина самки 0,8-1 мм, самца ― до 1 мм.
Один из наиболее распространённых неритических видов Чёрного
моря. Круглогодичный, эвритермный. Предпочитает поверхностные слои, однако,
встречается до глубины 150-175 м (Мордухай-Болтовский, 1968).
5. Acartia clausi Giesbrecht (Приложение, рис.15) ― представитель семейства Acartiidae. Тело очень стройное, удлинённое.
Цефалоторакс 5-сегментный. Передний край головы заметно притуплен, задние углы
последнего торакального сегмента значительно уже переднего. Абдомен самки 3-,
самца5-сегментный, с очень маленьким IV сегментом. Фуркальные ветви короткие, чуть
длиннее последнего сегмента абдомена. Фуркальных щетинок пять пар, они тонкие,
длинные. Геникулирующая антенна самца со слабым утолщением посредине, с
3-членистым концевым отрезком, на конце с локтеобразным изгибом. Длина самки
1,17-1,75 мм, самца ― 1-1,5 мм. Тело
прозрачное, непигментированное (Мордухай-Болтовский, 1968).
Наиболее распространённый вид в планктоне Чёрного моря.
Круглогодичный, эвритермный. Вид встречается в двух формах ― большой и малой. Большая является основной формой черноморского
планктона.
По литературным данным с 1994 г. В Чёрном море обитает и
другой вид Acartia tonsa Dana из семейства Acartiidae.
6. Harpacticus sp. (Приложение, рис.16). Тело
немного сужено книзу. Антенны I 8-9-членистые. Экзоподит антенн II 2-членистый, максиллипед
обычно расширен. Экзоподит и эндоподит первой пары торакальных ног
2-3-членистые, эндоподиты с одной дистальной щетинкой нам первом членике.
Эндоподит второго и третьего торакальных ног с признаками полового диморфизма.
Пятая пара грудных ног у обоих полов двуветвистая, экзоподит с пятью щетинками.
Типичные представители морской пелагиали, некоторые виды
космополиты (Мордухай-Болтовский, 1968).
7. Centropages kroyeri pontica Karaw (Приложение, рис.17). Тело
крепкое, коренастое, цефалоторакс 6-сегментный, все сегменты почти одинаковой
длины. Генитальный сегмент самки по бокам с небольшими кнопкообразными
выростами, покрытыми мелкими шипиками. Фуркальные ветви симметричные,
относительно длинные, с оперёнными внутренними краями. Фуркальные щетинки густо
оперены. Второй членик 4-ой пары торакальных ног у самца с большим шипом на
наружном крае. Длина самки 1,25-1,65 мм, самца - до 1,4 мм. Тело Чётко
пигментировано.
Тепловодная, преимущественно летне-осенняя форма. Летом
держится в верхних горизонтах, зимой опускается глубже. Встречается главным
образом в прибрежной зоне, но в небольшом количестве отмечается и в открытом
море. В летнем кормовом планктоне играет очень большую роль. Отличается высокой
плодовитостью - до 250 личинок на одну самку (Мордухай-Болтовский, 1968).
8. Sagitta setosa Muller (Приложение, рис.18) -
представитель типа Chaetognatha. Планктонное животное с вытянутым телом и двумя
парами боковых плавников. Хвостовой плавник округло-треугольный. Длина тела до
22 мм, имеется передние, задние, боковые и хвостовой плавники. Число передних
зубов 5-10, задних - 9-16. Имеется два цикла половой зрелости: при длине тела
16-17 мм и около 22 мм. В первом случае яичник содержит 10-15 зрелых яиц, во
втором - 70-80яиц. Форма, созревающая при длине тела до 22 мм, была описана
Молчановым в качестве особого вида S. euxina несколько раз пересматривался. Фюрнестен (цит.
по Мордухай-Болтовский, 1968). считает, что оснований для выделения крупной
формы в самостоятельный вид недостаточно, однако, имеются сторонники и
противоположной точки зрения.
9. Coryne tubulosa M. Sars (Sarsia mirabilis L. Agassiz, S. tubulosa M. Sars) (Приложение, рис. 19).
Тело полипа веретеновидное, снабжённое 16-20 беспорядочно расположенными
головчатыми щупальцами. Медузы имеют глазки в основании щупалец. Колонии
стелющиеся или слабо разветвлённые, высотой 2-3 см. Скелет ножки полипа и
стволика гладкий, но в нижней части неясная кольчатость. Тело полипа высотой до
1-2 мм. Медузоидные почки развиваются на теле полипа под щупальцами. Зонтик
медузы несколько выше полусферы, иногда значительно вытянут в высоту. По краю
зонтика очень длинных с глазком в основании щупальца. Ротовой хоботок
червеобразный, способный далеко вытягиваются за пределы полости зонтика
(Мордухай-Болтовский, 1968).
Обычный, часто массовый вид. Полипы поселяются в литорали,
медузы держатся в поверхностных слоях воды вблизи берегов.
10. Oikopleura dioica Fol (Приложение, рис. 20). Вид
является представителем подтипа Оболочники (Tunicata), класса Appendicularia.
Ротовое отверстие направлено вверх. Дорсальная сторона туловища почти плоская,
у половозрелых форм задняя часть туловища значительно расширена за счёт гонады.
Домик покрывает только переднюю часть туловища, гонада всегда свободна. Отоцист
крупный. Пищевод открывается в левый желудок. Правый желудок крупнее левого и
разделён на переднюю и заднюю части. Полностью развитые гонады крупные, чащевидные,
вогнутой стороной прилегают сзади к желудку. Длина туловища до 1 мм. Хвост
примерно в 4 раза длиннее туловища. В хвосте две субхордальные клетки. Питается
мелкими жгутиками, диатомовыми, кокколитофоридами. Обитает в широком диапазоне
температуры и солёностей. До работы В.Н. Никитина (1929) черноморский вид
неверно определяли как O. Сophocerca (Мордухай-Болтовский, 1968).
Распространён в тёплых и умеренных водах многих морей
Атлантического, Тихого и Индийского океанов. Считается более обычным в прибрежных
полосах.
11. Aurelia aurita L (Приложение, рис.21) -
представитель класса Сцифоидные медузы (Sciphozoa). Эта медуза напоминает
по форме зонтик. Интересным в организации этого животного является строение
органов чувств, которые собраны в комплексы - ропалии. Это видоизменённые
укороченные щупальца медузы. Внутри ропалии находится один статоцист, а по
бокам два пузыревидных глазка. Органы зрения представлены глазными пузырьками с
роговицей и хрусталиком. У A. Aurita имеется восемь ропалий, под которыми находятся восемь ганглий.
Гонады образуются в энтодерме желудка, в его карманах, и имеют подковообразную
форму. Аурелия является крупным видом медуз (Шарова, 2004).
12. Rhizostoma pulmo Macri (Приложение, рис.22) также
является представителем Sciphozoa. Этот вид имеет мясистый грибовидный купол,
из-под которого свисают тяжёлые кружевные ротовые лопасти. Купол корнерота
может достигать 70 см в диаметре; в длину такая медуза больше метра (Шарова,
2004).
13. Beroe ovate (Приложение, рис.23) -
крупный гребневик, относящийся к отряду Морские огурцы (Веroida). Для него характерно
отсутствие щупалец, наличие крупного, овального тела, огромного рта и сильно
растягивающей глотки. Этот вид является активным хищником, который питается
гребневиками, в частности мнемиопсисом, заглатывая его целиком (Шарова, 2004).
14. Noctiluca miriabilis, или ночесветка
(Приложение, рис.24) - представитель представитель класса Растительные
жгутиконосцы (Phitomastigophora), который относится к отряду панцирные жгутиконосцы (Dinoflagellida). У этого организма
округлое тело до 2 мм в поперечнике, жгутики редуцированы. Они способны
светиться в темноте. При ударе вёсел по воде или при движении плывущего
предмета начинается искрящееся фосфорическое свечение. Это явление объясняется
окислением жировых включений в цитоплазме ночесветки (Шарова, 2004).
В составе меропланктона нами идентифицированы
следующие представители:
1. Науплиус Acartiа (Приложение, рис.25).
Длина тела на I-VI стадиях равна 0,12; 0,13; 0,14; 0,17; 0,2 и 0,24 мм. Антенны
I 3-членистые. Коксоподит и базиподит антенн II с одним изогнутым шипом и одной
тонкой щетинкой. Вооружение заднего конца тела ортонауплиусов состоит из одной
пары маленьких шипов. Верхняя губа квадратная и по нижнему краю оперена рядом
ровных мелких щетинок. Науплиальные стадии Acartia встречаются в планктоне
верхних слоёв моря в течение всего года (Мордухай-Болтовский, 1968).
2. Науплиус Centropages (Приложение,
рис.26). Длина тела на I-VI стадиях равна 0,08; 0,1; 0,12; 0, 19; 0,21 и 0,25
мм. Антенны I 3-членистые. Коксоподит антенн II с одним изогнутым шипом,
базиподит - с одним шипом и двумя щетинками. Вооружение заднего конца тела I ортонауплиуса состоит из
одной пары шипов. Верхняя губа округлая, на старших науплиальных стадиях
опушена по нижнему краю рядом неровных мелких щетинок. Науплиальные стадии
Centropages встречаются в верхних слоях моря в летние месяцы
(Мордухай-Болтовский, 1968).
3. Науплиус Paracalanus (Приложение, рис.27).
Науплии очень мелкие. Длина тела на I-VI стадиях равна 0,08; 0,1; 0,13; 0,15;
0,18; 0,21 и 0,23 мм. Встречается в верхних слоях моря в тёплые месяцы года
(Мордухай-Болтовский, 1968).
4. Науплиус Oithona (Приложение,
рис.28). Длина тела на различных науплиальных стадиях варьирует от 0,05 до 0,23
мм. Экзоподит антенн II 6-членистый. Жевательные шипы коксоподита антенн II
развиты хорошо. Вооружение заднего конца тела представлено одной парой длинных
тонких щетинок у науплиусов первых двух стадий и двумя парами - у науплиуса III
стадии. Личинки в живом состоянии прозрачные. Встречаются в верхних слоях моря
в холодное время года (Мордухай-Болтовский, 1968).
5. Науплиус Balanus (Приложение, рис.29).
Выделяют 6 науплиальных стадий в развитии Balanus. Науплиус легко распознаётся
по так называемым фронтолатеральным рожкам. Эти рожки являются характерным
признаком науплиусов Усоногих раков (Cirripedia) и не встречается у личинок
ракообразных других систематических групп. У науплиусов первой стадии
фронтолатеральные рожки обычно направлены назад, иногда они плотно прижаты к
головному щитку. Начиная со второй стадии у всех видов фронтолатеральные рожки
приобретают передне-боковое направление. В передней части тела личинки на всех
стадиях хорошо заметен простой непарный науплиальный глаз. На шестой стадии в
дополнение появляются парные сложные глаза. Верхняя губа (лабрум) прикрывает
рот личинки. Для личинки характерно наличие дорсальной торакальной иглы и
абдоминального отростка (Полтаруха и др., 2008).
7. Поздняя личинка Bivalvia (Приложение, рис.31).
Данная стадия развития двустворчатых моллюсков является переходной от личинки к
молодому взрослому организму. Личинка довольно крупная, створки сформированы.
Через некоторое время оседает на дно и превращается в ювенильную особь. Часто
встречается в неритической зоне (Вершинин, 2007).
8. Поздняя личинка Polychaeta (Приложение, рис.32).
Личинки многощетинковых червей могут встречатся в планктоне Чёрного моря
круглый год. Наиболее обильны они в летнее-осенний период. Причём они
встречаются на самых разных стадиях развития: трохофоры, метатрохофоры и
нектохеты. Имеют важное значение в черноморском меропланктоне
(Мордухай-Болтовский, 1968).
Глава 5.
Экология. Экологическое и биологическое значение фито - и зоопланктона Чёрного
моря
5.1 Миграции
планктона
Популяциям многих представителей планктона свойственны
массовые перемещения, регулярно повторяющиеся в пространстве и во времени. Они
совершаются в вертикальном и горизонтальном направлениях, в те участки ареала,
где в данное время наиболее благоприятные условия. Таким образом, миграции
позволяют популяции маневреннее использовать жизненные ресурсы в соответствии с
изменением их расположения в пространстве и времени, а также в связи с
потребностями самих организмов.
Вертикальные миграции. В вертикальных
миграциях наибольшее значение имеют суточные, сезонные и возрастные
(онтогенетические).
Суточные миграции, свойственные многим представителям фито -
и зоопланктона, отличаются многообразием своего проявления: сроками, амплитудой,
направлением, скоростью и другими особенностями. По-видимому, столь же
разнообразно и их экологическое значение, во многих случаях ещё не
расшифрованное, трактуемое на уровне гипотез. Особенно сложна картина суточных
миграций зоопланктонтов. Большинство их в тёмное время суток концентрируются у
поверхности, днём ― в более глубоких слоях.
Заметно реже наблюдаются инвертированные миграции, при которых животные
сосредоточиваются у поверхности днём и держатся на глубине ночью (Константинов,
1986).
Как указывалось выше, механизм суточных вертикальных миграций
и их экологический смысл до конца не ясны. Некоторые авторы рассматривают
миграционные перемещения как пассивный процесс, вызываемый изменением скорости
переноса организмов вертикальными токами воды, особенно внутренними волнами,
имеющими по некоторым компонентам 12 - и 24-часовую периодичность. Другие
авторы акцентируют внимание на изменение скорости погружения организмов при
циклическом чередовании дневных и вечерних температур, сопровождающемся соответствующей
модуляцией условий плавучести (изменение плотности и вязкости воды). Третья
гипотеза строится на представлении о периодичности локомоторной активности,
стимулируемой или ингибируемой различными внешними воздействиями, в первую
очередь светом и температурой; погружение происходит пассивно, подъём ― за счёт активных движений. Эта гипотеза имеет больше сторонников,
чем первая и вторая. Важно отметить, что вертикальные миграции у организмов с
нейтральной плавучестью неизвестны (Рудяков, 1977, цит. по Константинов, 1986).
Ещё не менее ясен экологический смысл миграций, их адаптивное
значение, которое некоторые авторы вообще отрицают, полагая, что эволюция шла
по пути утраты миграционного поведения - автоматического следствия
несовершенства регуляции плавучести, неспособности обеспечить её нейтральность
(Константинов, 1986).
М.М. Кожовым, Б.П. Мантейфелем и рядом других исследователей
развито представление о защитно-кормовом значении суточных вертикальных
миграций. В тёмное время суток планктонные животные-фитофаги питаются в верхних
слоях воды, где наблюдается наиболее высокая концентрация водорослей, а на
светлое время суток уходят вглубь, тем самым резко уменьшая свою доступность
для рыб - зоопланктофагов. Однако, рыбы и другие планктофаги в качестве контрадаптации
также поднимаются к поверхности в вечернее время и используют сумерки для
интенсивного питания.
Наряду с фактами, свидетельствующими в пользу представления о
защитном значении вертикальных суточных миграций, имеется много других,
противоречащих этой гипотезе (Виноградов, 1968, цит. по Константинову, 1986):
существуют миграции на небольшую глубину, не спасающие мигрантов от поедания
рыбами; некоторые формы поднимаются к поверхности ночью, другие - днём.
Ж. Маклареном (цит. по Константинов, 1986). высказано мнение,
что в условиях прямой температурной стратификации воды суточные миграции
выгодны энергетически, так как на глубине в условиях более расход энергии на
рост и развитие ниже. Пребывание в холодной воде ведёт к увеличению размера
мигрантов и повышению плодовитости. Это создаёт демографические преимущества
для тех форм, у которых размер тела и плодовитость находятся в обратной
зависимости от температуры. Для питания животные поднимаются в верхние слои,
где корма больше, а после откорма погружаются вглубь. Подъём к поверхности в
ночное время не случаен, так как даёт известные преимущества (снижение
выедаемости). Свет служит основным сигнальным фактором, регулирующим
периодичность миграционной активности, эндогенная ритмика имеет корригирующее
значение.
В водных экосистемах на распределение организмов по слоям
большую роль оказывают гидрологические особенности воды, в первую очередь это
солёность и температура. В связи с этим морские организмы можно подразделить на
три группы: психрофильные, обитающие при низких температурах, мезофильные - в
диапазоне средних температур и термофильные, обитающие при высоких
температурах.
Горизонтальные миграции. Горизонтальное
распределение черноморского зоопланктона неравномерное. Особенно богата им
северо-западная часть моря вследствие значительного притока пресных вод (Дунай,
Днестр, Днепр и др. большие реки), содержащих биогенные соли. Сравнительно
хорошо развивается планктон в заливах, а также к юго-востоку от Крымского
полуострова. Обилие зоопланктона в этой акватории связано с водами, богатыми
биогенными солями. Кроме того, в центральных областях моря обнаружены районы,
богатые зоопланктоном вследствие подъёма на поверхность глубинных вод (Вылканов
и др., 1983).
Значительное влияние на распределение зоопланктона, особенно
в мелководных районах и поверхностных слоях, оказывают ветры. В летние месяцы
часто сильные сгонные ветры относят верхний, богатый зоопланктоном слой на
десятки миль от берега, а на его место из глубин поднимаются холодные массы,
сравнительно бедные зоопланктоном, состоящим из холодноводных видов.
Вертикальное распределение зоопланктона хорошо выражено. Нижняя граница его
распространения в Чёрном море почти всюду определяется проникновением кислорода
на глубины 100-175 м и только на юго-западе под воздействием антициклонического
течения спускается до 225 м.
Зоопланктон имеет специфическое распределение по сезонам и
глубинам, которое обусловливается температурными и трофическими потребностями
его и изменением солёности вод. Одна часть зоопланктона не зависит от
температуры воды, т.е. эвритермна, и встречается круглый год чаще всего на
глубинах 15-50 м. Другая часть зоопланктона предпочитает более низкие
температуры, т.е. криофильна. Зимой криофилы встречаются на всех глубинах, а с
наступлением лета они спускаются ниже 50-60 м, где температура относительно
постоянная. Осенью в связи с похолоданием и перемешиванием воды они снова
поднимаются на поверхность. Третья группа видов зоопланктона теплолюбивая
(термофильная) - встречается только или преимущественно летом. С потеплением
воды количество термофильных зоопланктёров увеличивается, достигая максимума в
конце лета или в начале осени; зимой зоопланктон исчезает. К этой категории
относятся водные блохи и некоторые веслоногие рачки (рода Centropages).
5.2 Пищевые
цепи
Термин "пищевая цепь" означает ряды организмов,
одни из которых служат пищей для других. Пищевые цепи большинства водных
животных начинаются с живой растительной клетки или с продуктов её распада.
Первоисточником пищи всех водных животных являются органические существа,
созданные водными растениями. Растительный планктон является главной пищей
одной части животного планктона и молоди рыб. Другая часть зоопланктона и
большинство бентосных беспозвоночных и рыб потребляют животную пищу.
Таким образом, органические вещества, созданные растениями,
проходят через различные организмы и после смерти и гниения конечного
потребителя превращаются в минеральные соединения, которые становятся пищей
растительных организмов.
В Чёрном море, как и в Мировом океане, основным потребителем
фитопланктона становится зоопланктон, а точнее веслоногие рачки (Сopepoda). Они отлавливают из
воды фитопланктонные организмы, а также бактерии и мельчайший детрит, а сами
служат пищей для рыб.
В пелагиали Чёрного моря различают два сообщества:
) эпипланктонное - в тонком слое воды выше термоклина, или
метаталасса, (выше промежуточного водного слоя, расположенного ниже эпиталасса,
температура которого изменяется в зависимости от сезона, и выше гипоталасса,
температура которого не изменяется);
) батипланктонное, находящегося в значительно более мощном
слое воды ниже термоклина.
Все организмы планктонных сообществ можно разделить на шесть
трофических уровней:
) первичных продуцентов и сапрофагов,
) растительноядных организмов,
) потребителей смешанной пищи,
) хищников первого порядка,
) хищников второго порядка,
) хищников третьего порядка (Вылканов и др., 1983).
Основной поток вещества и энергии в эпипланктонную систему
проходит через мелкие и средние организмы всех трофических уровней. Водоросли
размером до 50 мкм и животные организмы размером до 1,5 мм имеют ничтожную
биомассу, однако темп образования вещества и энергетический обмен их высок.
Именно эти организмы составляют основную массу планктона, и используются в
качестве пиши организмами более высокого трофического уровня.
В батипланктонных сообществах основной поток вещества и
энергии проходит главным образом через крупные организмы всех трофических
уровней.
Трофический уровень объединяет в одну группу все организмы,
тип питания и состав пищи которых одинаковы независимо от таксономической
принадлежности организмов. Все трофические уровни представляют собой звенья
одной пищевой цепи, в которой организмы последующего звена поедают организмы
предыдущего звена, имеющего больший запас энергии. В каждом сообществе Чёрного
моря абсолютная величина потока энергии от первого трофического уровня к
последнему уменьшается в 20 раз.
Начальным звеном большинства цепей служат продуценты
(фитопланктон и фитобентос).
Последним звеном пищевых цепей в море оказываются рыбы или
морские млекопитающие, которые используются человеком. Лишь в редких случаях
последним звеном становится животное, не употребляющееся в пишу остальными
животными или человеком. Пищевые цепи могут быть длинными и сложными, но
известны и очень короткие, как, например, водоросли - человек. Вопрос о пищевых
цепях имеет большое практическое значение. С хозяйственной точки зрения,
наиболее выгодны короткие цепи с малым числом звеньев.
В Чёрном море известны три класса животных, имеющих большое
пищевое значение: ракообразные, моллюски и кольчатые черви. Они служат основной
пищей для рыб. Знать пищевые взаимоотношения между рыбами, ценными в
промысловом отношении, весьма важно для рыбной разведки, которая обеспечивает
успешный лов рыбы. С этой целью изучаются количественные распределения
планктонных и бентосных организмов, прямо или косвенно служащих пищей для рыб.
Планктон Чёрного моря - самое богатое и быстрее всего
восстанавливающееся звено пищевых цепей. Зоопланктон, как и детрит, служит
пищей множеству основных групп морских животных - губкам, значительной части
сидячих многощетинковых червей, большинству низших ракообразных, двустворчатым
моллюскам, оболочникам и др.
Каждый вид животных является звеном какой-либо пищевой цепи.
Растительный и животный мир в водной массе и на дне Чёрного моря связан в одно
целое: растения (водоросли), производящее живое вещество являющиеся
продуцентами, потребляются животными - консументами, которые превращают их в
другие органические вещества, в конечном счёте минерализующиеся
микроорганизмами - редуцентами. Так в море осуществляется вечный круговорот
органических веществ, в котором особую роль играют освобождённые при
минерализации биогенные соли, а от них зависит количественное развитие фитопланктона
и фитобентоса. Таким способом поддерживается равновесие в динамике жизни моря.
Круговорот органических веществ не замкнутая система - некоторая часть
органических веществ переходит в донные осадки (Вылканов и др., 1983).
Таким образом, планктон Чёрного моря является важным звеном
пищевой цепи морской экосистемы.
5.3 Биомасса
и продуктивность
Представление о том, что жизнь в Чёрном море качественно
однообразна и количественно бедна, зародилось в середине XIX века при сравнении
черноморской фауны с богатой и разнообразной фауной Средиземного моря, а также
с исключительно богатой фауной рыб Азовского моря. Это представление
утвердилось после открытия сероводорода в безжизненных глубинах Чёрного моря и
доминировало в науке до 50-х годов XX века.
Качественные исследования в Чёрном море, которые были
проведены в конце XIX и в течении всего XX века, способствовали увеличению списка
представителей его флоры и фауны. Исследования русских учёных опровергли
неверное представление о качественной и количественной бедности жизни в Чёрном
море. Особые заслуги в выяснении этого важного в теоретическом и практическом
отношении вопроса имеет советский учёный Водяницкий (Вылканов и др., 1983). На
оснований исследований бактериального населения, фитопланктона, фитобентоса, зоопланктона,
а также рыбного населения Чёрного моря, он сделал следующий вывод о
биологической продуктивности Чёрного моря: "По сравнению с другими морями
Чёрное море не имеет признаков ослабления продукционных процессов; по
продуктивности оно занимает одно из первых мест".
При оценке продукционных свойств моря употребляют термины
"биомасса" и "биологическая продукция". Под биомассой
понимается масса живых организмов, обитающих в определённый момент на единице
поверхности или в единице объёма местообитания. Прирост биомассы за
определённое время (сутки, месяц, год) представляют собой биологическую
продукцию, изменение которой отражает ход биологических процессов в водоёме.
Существует три вида продукции: первичная - органические
вещества, являющиеся продуктом жизнедеятельности фотосинтезирующих морских
растений (фитопланктон и фитобентос), вторичная - органическое вещество
животных, которое они усвоили из растений, конечная - та часть первых двух
видов продукции, которую потребляют люди. Ценные виды конечной продукции
образуют биологические ресурсы водоёма.
Величина первичной продукции определяется путём учёта
фотосинтеза водных растений, но интенсивность фотосинтеза нельзя
идентифицировать с количеством первичной продукции. Интенсивность первичного
продуцирования выражается двумя величинами: валовой первичной продукцией
(количество органического вещества, образованного при фотосинтезе) и чистой
первичной продукцией (часть органического вещества, не расходуясь на обмен
растений, накапливается в них или выделяется ими в воду).
Первичная продукция образуется в поверхностном слое моря, где
освещение и наличие биогенных солей обеспечивают протекание процессов
фотосинтеза в растительных организмах. В Чёрном море имеются предпосылки для
развития богатой первичной продукции: "живой слой" воды (0-200 м)
получает значительное количество биогенных солей из речного стока и при
вертикальном обмене.
По величине первичной продукции определяют величину вторичной
и конечной продукции водоёма. Одна часть чистой первичной продукции
растительных организмов включается в пищевые цепи животных и накапливается в
них, образуя вторичную продукцию, другая часть минерализуется.
Типичными потребителями первичной продукции в Чёрном море
служат представители зоопланктона. Подобно первичной, вторичная продукция также
бывает валовой и чистой - равной приросту водных животных. Вторичная продукция
рассчитывается, как и первичная, на единицу объёма или на единицу площади
водоёма. Её количество выражается в массе сырого или сухого вещества, в калориях,
в количестве белков, жиров и углеводов.
Для оценки суммарной чистой продукции водоёма используют
предложенный Зенкевичем Р/В - коэффициент - отношение годовой продукции (Р) к
их средней годовой биомассе (В). Произведение биомассы морских животных на их Р/В-коэффициент
равняется общей чистой вторичной продукции моря. Этот коэффициент для различных
обитателей Чёрного моря различен: для фитопланктона - 300, зоопланктона - 15,
зообентоса - 2,5, рыб ― 0,5.
Промысловые водные организмы составляют биологические ресурсы
водоёма, которые, в отличие от полезных ископаемых, самовоспроизводятся.
Биологические ресурсы океанов и морей определяются не количеством ценных в
промысловом отношении организмов, а приростом их, т.е. продукцией.
Средняя годовая биомасса кормового зоопланктона в восточной
половине Чёрного моря составляет примерно 50 мг на 1 м3, в западной
- примерно 80 мг на 1м3 (Вылканов и др., 1983). Биомасса
зоопланктона оценивается в 21 млн. тонн, а годовая продукция - 315 млн. тонн.
Самый богатый планктоном (31,1% общей биомассы) слой лежит до глубины 23 м, в
слое 50 м содержится 51,8% общей биомассы.
По данным Мартынюка биомасса веслоногих рачков (Copepoda) в
северо-восточной части Чёрного моря в 1993-1998 годах составлял 45,3 мг/м3,
в 2000-2003 годах - 26,4 мг/м3, а в 2005 г. она составила 14,8 мг/м3
(Мартынюк, 2004 - 2005). Что касается ветвистоусых рачков (Cladocera), то биомасса в
прибрежном районе в 93-98-х годах составляло 0,6 мг/м3, в 2000-2003
годах - 2,1 мг/м3, в 2005 - 0,1 мг/м3.
Интересно проследить и динамику биомассы меропланктона по
данным Мартынюка. В 1993-1998 годах она составила 1,5 мг/м3, в
2000-2003 годах опустилась до 0,3 мг/м3, а в 2005 г. уже возросла до
11,2 мг/м3 (Мартынюк, 2004-2005).
Если сравнивать в целом биомассу кормового зоопланктона, то с
1993 г. по 2005 г. наблюдается её снижение. Так, в 1993-1998 годах она в
среднем составила 56,2 мг/м3, в 2000-2003 годах она достигла 34,4
мг/м3, в 2005 г. - 28,3 мг/м3 (Мартынюк, 2004-2005). Из
выше изложенного следует, что половину биомассы кормового зоопланктона
формируют в основном Copepodа, меньшую часть, примерно 35-40 % от общей
биомассы формирует меропланктон.
По данным Вылканова биомасса фитопланктона Чёрного моря до
глубины 50 м оценивается в 3,6 млн. т, а до глубины 100 м - 4,3 млн. т;
продукция фитопланктона на тех же глубинах составляет соответственно 1080 и
1290 млн. т сырого вещества (Вылканов, 1983). Средняя биомасса фитопланктона в
1960-1984 годах составила 312 мг/м3. Самые высокие среднегодовые
биомассы отмечались в 76-82-х годах прошлого столетия. Во все годы среди
фитопланктонных организмов доминировали диатомовые водоросли (Diatomea) и
динофлагелляты (Dinophyta).
5.4 Динамика
численности и биомассы планктона в связи с влиянием вселенцев - Mnemiopsis
leidyi и Beroe ovatа
Закрытая на протяжении миллионов лет и недавно открывшаяся,
но всё-таки полузамкнутая экосистема Чёрного моря, до сих пор далека от
равновесного состояния и очень чувствительна к внешним воздействием. Поэтому
изменения биологической структуры Чёрного моря продолжаются по сей день -
происходит вселение новых видов и исчезновение ранее существовавших. Большая
часть современных черноморских видов - около 80% - средиземноморские вселенцы,
пришедшие сюда после появления Босфорского пролива. Вселение новых видов может
происходить как целенаправленно, так и случайно.
Видами, которые случайно оказались завезёнными в Чёрное море,
и в значительной степени повлиявшие на зоопланктонное сообщество оказались Mnemiopsis leidyi и Beroe ovatа.
В начале 80-х годов в Чёрном море появился экзотический
вселенец Mnemiopsis leidyi, представитель североамериканской фауны.
Адаптировавшись в новых для себя
условиях, он к концу 80-х годов резко увеличил биомассу до 1
млрд т и стал мощным конкурентом для всех потребителей зоопланктона, сдерживая
их биомассу на низком уровне. Кроме того, потребляя представителей
ихтиопланктона (икра и молодь рыб), он значительно снизил воспроизводство
многих черноморских видов. Особенно сильно пострадали массовые виды. Однако,
качественных изменений в соотношении различных групп ихтиопланктона не
произошло, как и в "догребневиковый" период, по данным Надолинского,
в уловах ИКС-80 продолжали доминировать икра и личинки хамсы, ставртды,
барабули, а также личинки морских собачек и бычков бубырей (Надолинский, 2005).
Вселение мнемиопсиса негативна сказалась и на представителей Bivalvia; вообще численность
практически всех видов животных и растений в Чёрном море стала резко
уменьшаться.
Причиной массового развития мнемиопсиса стало отсутствие
хищников, способных контролировать их численность - никто не ел мнемиопсиса.
Положение исправилось в 1990-х годах - в Чёрном море появился другой гребневик
из Атлантики - Beroeovatа.
B. ovatа завезён сюда случайно с балластными водами, также, как и
мнемиопсис. Этот вид питается мнемиопсисом, и только мнемиопсисом. Интересно,
что больше он не ест никого - редкий случай очень узкой пищевой специализации.
Теперь каждый год происходит следующее: появляются мнемиопсисы в прибрежных
водах в апреле; к середине лета их становится очень много, однако, после этого
появляется берое. К октябрю мнемиопсисов в море практически не остаётся, но
зато численность берое сильно возрастает.
Наибольший положительный эффект от вселения берое
проявляется, как было сказано выше, в летне-осенний период. Являясь генетически
сезонной формой, это животное начинает развиваться в Чёрном море только со
второй половины августа, когда количество мнемиопсисов становится достаточно
высоким. С этого момента берое активно потребляет мнемиопсиса и тем самым
контролирует его численность. В этот период у зоопланктонного сообщества
появляется возможность максимально реализовать свои продукционные способности,
в результате чего биомасса его существенно возрастает. Так, в летне-осенний
период 2004 г биомасса кормового зоопланктона, изменяясь от 55 до 1778 мг/м3,
в среднем составила 379 мг/м3.
Из выше сказанного следует, что в случае берое можно говорить
о полезной интродукции нового вида - этот хищный гребневик остановил
катастрофическое оскудение морского сообщества. Характеризуя состав
зоопланктона в летне-осенний период, отметить, что до появления в водах Чёрного
моря берое одну вторую часть его биомассы составляли копеподы. На долю
кладоцер, наиболее активно выедаемых мнемиопсисом, приходилось не более 30%.
После вселения берое доля их в общей массе планктонной фауны значительно
возросла, и они стали ведущей группой. Так, в 1993-1998 г биомасса кладоцер в
летне-осенний период составлял 14,4 мг/м3, а в 2004 г - 265,3 мг/м3.
Второе место по своей значимости по данным исследований 2005
г занимает меропланктон. Его биомассу формировали личинки двустворчатых
моллюсков -велигеры, которые присутствуют в планктоне практически весь год, но
особенно их много в тёплое время года (август-сентябрь), когда происходит
интенсивное размножение моллюсков. Немаловажную роль в увеличении численности
личинок моллюсков сыграло также и снижение численности хищного моллюска - Rapana thomasiana, который активно поедает
взрослых моллюсков, снижая тем самым численность размножающихся особей.
Развитие копепод в последние годы характеризовалось невысоким
уровнем. Однако, видовое разнообразие летне-осеннего комплекса этих животных
возросло. Вместо встречающихся ранее 6-7 видов, в 2004-2005 гг. отмечено 12
видов (Мартынюк, 2004-2005). В 2005 г её средняя численность составила 379
экз/м3.
Сезонные изменения количественного и качественного состава, а
соответственно и биомассы характерно и для фитопланктона. Так, зимой
преобладают диатомовые водоросли (в основном виды рода Chaetoceros), весной также
диатомовые, летом ― динофлагелляты, а осенью
снова преобладают диатомовые. Количественное развитие диатомовых водорослей
имеет зимне-весенний максимум и редко ещё и осенний, динофлагелляты же
развиваются преимущественно летом и реже осенью, но биомасса их вследствие малых
размеров многих видов остаётся меньшей. Фитопланктон обильно развивается и
цветёт в определённый сезон, в случае если поверхностный слой моря содержит
большое количество биогенных солей. Когда количество этих солей сильно
уменьшится или их нет совсем, цветение уменьшается и прекращается. Затем
усиливаются процессы гниения, снова высвобождающие биогенные соли и
подготавливающие условия для нового цветения воды.
Уровень развития весеннего планктона в значительной мере
определяется абиотическими факторами, такими, как характер зимы, ветровая
активность, устойчивость атмосферных процессов, теплонакопление водных масс
(Мартынюк, 2004-2005).
Таким образом, сезонное изменение численности и биомассы
планктёров происходит под активным влиянием не только абиотических, но и
биотических факторов.
Заключение
Чёрное море - типичное внутреннее море с общей площадью
420325 км2 и общим объёмом воды 547015 км3. Чёрное море
является средиземным (внутренним) морем Атлантического океана, которое на
северо-востоке соединяется с Азовским морем Керченским проливом, а на
юго-западе - Босфорским проливом с Мраморным морем.
Чёрное море является уникальным морем - нет ни одного другого
внутреннего моря в мире, в которое втекает столько пресной воды - её приносят
крупнейшие реки Европы и Азии, среди которых самые крупные это: Дунай, Днестр,
Днепр, Буг, Кубань, Риони и др.
В рельефе черноморского дна чётко выделяются три основные
формы - шельф, материковый склон и глубоководная котловина с наибольшей
глубиной 2245 м.
Черноморская вода по своим гидрофизическим особенностям
отличается от воды других морей и океанов. Средняя солёность поверхностных вод
составляет 18‰, на глубине 200 м - 20‰, а у дна моря -30‰. Температурный режим
Чёрного моря типичен для водоёмов умеренной зоны: зимой поверхностные воды его
северо-западного и северо-восточного районов охлаждаются до нуля, а в
мелководных участках шельфа и в бухтах образуется ледовый покров. Самый тёплый
район Чёрного моря - его юго-восточная часть. Концентрация растворённого
кислорода в поверхностных слоях моря меняется в зависимости от сезона: в тёплое
время года его содержание уменьшается, а в холодное - увеличивается. Показатели
концентрации растворённого кислорода в различные сезоны варьируют от 6,66 мг/л
до 13,49 мг/л. Распределение концентрации растворённого кислорода в основном
зависит от процесса фотосинтеза и температуры воды.
Для исследований планктонных организмов применяют различные
методы с использованием разнообразных орудий для сбора материала. Самым простым
и широко распространённым методом для сбора планктёров является сетяной метод,
при котором применяют качественные и количественные сети разных конструкций
(сеть Апштейна, сеть Джеди и др.). Кроме сетей наиболее распространёнными
орудиями для сбора материала по планктону являются различные
планктонособиратели, батометры и планктоносборщики.
Гидробионты, относящиеся к экологической группе планктонных
организмов, включают в себя как одноклеточных, так и многоклеточных организмов,
относящиеся к разным таксономическим единицам. Эти гидробионты характеризуются
конвергентными морфолого-экологическими адаптациями, которые обеспечивают им
нахождение в пелагиали моря во взвешенном состоянии.
В состав фитопланктона Чёрного моря входят более 700 видов,
среди которых наиболее важное значение имеют представители диатомовых
водорослей (Bacillariophyta) и динофлагеллят (Dinophyta). Среди зоопланктёров есть представители
типа Sarcomastigophora, Ciliata, Coelenterata, Acnidaria и представители других типов животных; однако, наиболее
важными представителями черноморского зоопланктона являются ракообразные, в
основном веслоногие рачки (Copepoda) и ветвистоусые (Cladocera), среди которых есть
такие виды, как Oithona sp., Oithona similis, Penilia avirostris, Evande spinifera и др.
На горизонтальное и вертикальное распределение планктона
большую роль оказывают гидрологические особенности воды, в первую очередь
солёность и температура. Одна часть планктона предпочитает низкие температуры
(криофильна), а другая часть вовсе не зависит от температуры воды (эвритермна).
Планктон Чёрного моря является важным звеном пищевой цепи
морской экосистемы, так как планктонное сообщество - самое богатое и быстрее
всего восстанавливающееся звено пищевых цепей. Планктонные организмы, как и
детрит, служат пищей множеству основных групп морских животных - губкам,
большинству низших ракообразных, двустворчатым моллюскам, оболочникам и др.
Значительное влияние на планктёров Чёрного моря оказывают Mnemiopsisleidii и Beroeovata - вселенцы, которые проникли
сюда случайно с балластными водами какого-то судна от Атлантического побережья
Северной Америки. Мнемиопсис оказал негативное влияние на экосистему Чёрного
моря, вызвав здесь экологическую катастрофу. Положение исправилось в 1990-х
годах с появлением в море другого гребневика - берое. В случае берое мы можем
говорить о полезной интродукции нового вида.
Выводы
1. В составе черноморского фитопланктона в летне-осенний
период могут встречаться представители следующих видов: Сeratium tripos Nitzsch, Ceratium furca Ehr, Ceratium fusus Ehr, Pseudosolenia calcar-avis Schuitze, Proboscia alata Brighw, Prorocentrum micans Ehrenberg, Chaetoceros sp.
. В фитопланктоне Чёрного моря доминирующими, наиболее
массовыми формами, служащими кормовой базой являются представители динофитовых
(Dinophyta) и диатомовых водорослей
(Diatomea), в основном
представители родов Proboscia, Pseudosolenia, Ceratium и Chaetoceros.
. В результате исследования обнаружены некоторые особенности
сезонного распространения фитопланктонных организмов. Так, в летних пробах
основу фитопланктона составляли представители родов Proboscia и Pseudosolenia, а в осенних пробах
доминировали различные представители хетоцеросов (Chaetoceros).
. В составе зоопланктонного сообщества Чёрного моря в
летне-осенний период встречаются следующие представители: Penilia avirostris Dana, Oithona minuta Kricz, Oithona similis Claus,. Paracalanus parvus Claus, Acartia clausi Giesbrecht, Harpacticus sp., Centropages kroyeri pontica Karaw, Sagitta setosa Muller, Coryne tubulosa M. Sars, Oikopleura dioica Fol, Aurelia aurita L., Rhizostoma pulmo Macri. Beroe ovatа, Noctiluca miriabilis.
. В Чёрном море основу кормового зоопланктона составляют
такие массовые формы, как веслоногие (Copepoda) и ветвистоусые (Cladocera) раки
и представители меропланктона (временного планктона).
. В состав черноморского меропланктона входят личинки
различных организмов, среди которых нами были обнаружены следующие: науплиус Acartiа, науплиус Centropages,
науплиус Paracalanus, науплиус Oithona, науплиус Balanus, Cipris (циприсовидная
личинка Balanus), поздняя личинка Bivalvia, поздняя личинка Polychaeta.
. Исходя из оценки литературных данных по биомассе планктона
Чёрного моря, следует, что наибольшей биомассой обладают веслоногие
ракообразные и меропланктон: в 2003 г. биомасса копепод составляла 26,4 мг/м3,
а меропланктона - 11,2 мг/м3. Если сравнивать в целом биомассу
кормового зоопланктона, то с 1993 г. по 2005 г. наблюдается её снижение.
Литература
1.
Вершинин А.О. Жизнь Чёрного моря. - Краснодар-Москва, 2007. С.67-80, 145-191.
.
Воловик С.П., Корпакова И.Г., Афанасьев Д.Ф., Федяева В.В., Громов В.В. Флора
водных и прибрежно-водных экосистем Азово-Черноморского бассейна. ФГУП
"АзНИИРХ" Краснодар, 2008. С.168-198.
.
Вылканов А. Чёрное море. - Ленинград, 1983.С. - 36-215.
.
Дбар Р.С., Страничкина Г.А., Пустоварова О.В., Янкова Г.А. Результаты
мониторинга питьевой и морской воды в 1998-2000 гг. а Республике Абхазия //
Сухум: ГСЭБ и ООСРА. 2001.
.
Дбар Р.С., Экба Я.А. Дендроклиматические методы определения цикличности
метеоэлементов на территории Абхазии // Тр. Межд. конф. "Горные экосистемы
и их компоненты". - Нальчик, КБНЦ РАН. 2005.Т.I. С.104-111.
.
Забелина М.М., Киселёв И.А., Прошкина-Лавренко А.И., Шешукова В.С. Определитель
пресноводных водорослей СССР. Диатомовые водоросли. - М.: издательство
"Советская Наука". 1951. Выпуск 4. С.108-111.
.
Зенкевич Л.А. Моря СССР. Их фауна и флора. - М.: издательство Министерства
просвещения РСФСР. 1956. С.5-11, 313-316.
.
Киселёв И.А. Панцырные жгутиконосцы. - М.: издательство Академии Наук СССР.
1950. С.51-54, 62-115.
.
Константинов
А.С. Общая гидробиология. ― Москва, 1986. С.124-131.
.
Мартынюк М.Л. Состояние зоопланктонного сообщества в прибрежном районе
северо-восточной части Чёрного моря // Основные проблемы рыбного хозяйства и
охраны рыбохозяйственных водоёмов Азово-Черноморского бассейна // Сб. Науч. тр.
Ростов-на-Дону, 2004-2005.С. - 107-111.
.
Мордухай-Болтовский Ф.Д. Определитель фауны Чёрного и Азовского морей. - Киев:
издательство "Наукова Думка". 1968. т.I. С.56-72, 360-372.
.
Мордухай-Болтовский Ф.Д. Определитель фауны Чёрного и Азовского морей. - Киев:
издательство "Наукова Думка". 1969. т. II. С.12-53, 152-163.
.
Мордухай-Болтовский Ф.Д. Определитель фауны Чёрного и Азовского морей. - Киев:
издательство "Наукова Думка". 1972. т. III. С.250-271, 292-301.
.
Надолинский В.П. Оценка состояния ихтиопланктона Азовского и северо-восточной
части Чёрного морей и причины, его определяющие в современный период // Основные проблемы рыбного
хозяйства и охраны рыбохозяйственных водоёмов Азово-Черноморского бассейна //
Сб. Науч. тр. Ростов-на-Дону, 2004-2005.С. - 107-111.
.
Полтаруха О.П., Корн О.М. Атлас-определитель усоногих раков (Cirripedia:
Thoracica) прибрежных вод России. - М.: Товарищество научных изданий КМК. 2008.
С.11-18.
.
Пряхин Ю.В., Шницкий В.А. Методы рыбохозяйственных исследований. ― Краснодар, 2006. С.36-43.
.
Скопинцев Б.А. Формирование современного химического состава вод Чёрного моря.
- Л.: Гидрометеоиздат, 1975.
.
Шарова
И.Х. Зоология беспозвоночных. - М., 2004.С. - 239-257.
.
Экба Я.А., Дбар Р.С., Ахсалба А.К. Дендроклиматическая оценка воздействия
глобального потепления на хвойные лесные ценозы Абхазии // Тр. третьей Межд.
конф.: Биоразнообразие Кавказа. - Нальчик: издательство КБНЦ РАН. 2004.
С.92-107.
.
Экба Я.А., Дбар Р.С., Ахсалба А.К. Экологическая роль атмосферных и их влияние
на продуктивность экосистем // Проблемы экологии горных территорий. - Нальчик:
издательство КБНЦ РАН. 2003.С. 193-200.
.
Экба Я.А., Дбар Р.С., Гицба Я.В. Загрязнение нефтепродуктами Сухумской
акватории Чёрного моря // Труды третьей региональной конференции
"Биоразнообразие Кавказа". - Нальчик. 2004. С.109-114.
.
Экба Я.Л., Дбар Р.С. Экологическая климатология и природные ландшафты Абхазии.
- Сухум, 2007. С.224-271.