Анатомическое строение листовой пластинки двудольных растений. Типы корней и корневых систем
Содержание
Анатомическое
строение листовой пластинки двудольных растений
Типы
корней и корневых систем
Происхождение
цветка
Микроспорогенез
(образование микроспор в гнездах пыльника). Строение мужского гаметофита
(пыльцевого зерна)
Ботаническая
характеристика рода Паслен
Характеристика
семейства Маковые, представители и значение этого семейства
Характеристика
суккулентов, примеры
Список
использованной литературы
Анатомическое строение листовой
пластинки двудольных растений. Приведите рисунок
Лист - это один из основных органов растения,
занимающий боковое положение на стебле и выполняющий функции фотосинтеза,
транспирации (испарения воды растением) и газообмена с окружающей средой.
Анатомия листа. Типичное анатомическое строение
листовой пластинки отражает ее приспособленность к выполняемым функциям (рис.
1). С обеих сторон она покрыта эпидермисом, который регулирует газообмен и
транспирацию. В клетках кожицы нет хлоропластов, поэтому они беспрепятственно
пропускают свет к основным тканям листа. Наружные стенки клетки кожицы,
особенно с верхней стороны листа, утолщены и покрыты слоем воска или
воскоподобного вещества - кутина, что предохраняет лист от перегрева и
излишнего испарения воды. Этому способствует также погружение устьиц вглубь
листовой пластинки, формирование волосков, создающих разные виды опушения, и
др.
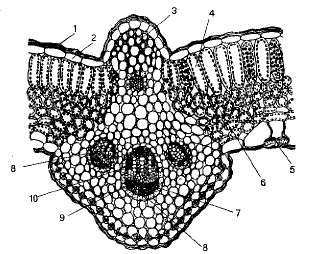
Рис. 1 - Анатомическое строение листа георгины
(поперечный срез): 1 - кутикула; 2 - эпидермис; 3 -- колленхима; 4 - палисадная
паренхима; 5-устьице; 6 - губчатая паренхима; 7 - флоэма; 8 - прово дящий
пучок; 9 - склеренхима; 10 - ксилема
Особенности внутреннего строения листа
определяются его главной функцией - фотосинтезом. Поэтому важнейшей тканью
листа является хлорофиллоносная паренхима (хлоренхима). Эта ткань образует
мякоть листа, или мезофилл, в клетках которого сосредоточены хлоропласты и
происходит фотосинтез. Остальные ткани обеспечивают нормальную работу
мезофилла. Система разветвленных проводящих пучков, которые пронизывают
листовую пластинку во всех направлениях, снабжает лист водой и обеспечивает
постоянный отток органических веществ от листа к другим органам растения.
Механические ткани (склеренхима, колленхима) совместно с живыми клетками
паренхимы (мезофилла) и эпидермиса обеспечивают определенную структуру и
высокую прочность листовой пластинки. Поэтому сравнительно тонкие и нежные
листовые пластинки способны занимать в пространстве такое положение, при
котором создаются наилучшие условия освещения и газообмена.
Мезофилл занимает все пространство между верхним
и нижним эпидермисом листа, исключая проводящие пучки и механи-- ческие ткани.
Клетки мезофилла имеют округлую или слегка вытянутую форму, с тонкими и
неодревесневшими стенками.
Мезофилл чаще всего дифференцирован на
палисадную (столбчатую) и губчатую паренхиму. Обычно палисадная паренхима
располагается под верхним эпидермисом, а губчатая прилегает к нижнему (см. рис.
8.13).
Клетки палисадной паренхимы вытянуты
перпендикулярно к поверхности листа и расположены в один или несколько слоев.
Они содержат примерно 75-80% всех хлоропластов листа и выполняют основную
работу по ассимиляции углекислого газа. Поэтому палисадная ткань располагается
в наилучших условиях освещения, непосредственно под верхней эпидермой.
Благодаря тому что ее клетки вытянуты перпендикулярно к поверхности листа, у
них имеется возможность регулировать направление и расположение хлоропластов
таким образом, чтобы избежать повреждающего действия прямой солнечной радиации
на фото синтезирующий аппарат, На сильном свету хлоропласты занимают в клетке
пристенное положение и становятся ребром к направлению лучей, в результате чего
большая часть светового потока проходит мимо хлоропластов или скользит по их
поверхности, не разрушая хлорофилл. При слабом освещении, наоборот, хлоропласты
распределяются в клетке диффузно или скапливаются в нижней ее части, что
способствует лучшему освещению каждого из них. В округлых клетках, характерных
для губчатой паренхимы, такое регулирование расположения хлоропластов при
разной освещенности (особенно на сильном свету) практически невозможно.
Под столбчатой паренхимой находится рыхлая
паренхима, клетки которой имеют округлую или продолговатую форму, содержат
меньше хлоропластов и расположены рыхло, поскольку между ними развиваются
крупные межклетники, заполненные воздухом.
В губчатой ткани интенсивность фотосинтеза ниже,
чем в столбчатой, но зато здесь активно идут процессы транспирации и
газообмена. Углекислый газ из воздуха через устьица, расположенные главным
образом в нижнем эпидермисе, проникает в большие межклетники и по ним поступает
ко всем ассимилирующим клеткам листа. Парообразная влага, кислород и углекислый
газ, образующиеся при фотосинтезе и дыхании клеток мезофилла, передвигаются в
обратном направлении и через устьица выделяются наружу. Таким образом, оба вида
ассимиляционной ткан и (палисадной и рыхлой) тесно связаны между собой не
только в структурном, но и в функциональном плане.
Расположение устьиц преимущественно на нижней
стороне листа имеет важное экологическое значение. Во-первых, нижняя сторона
листа меньше нагревается на свету, чем верхняя, поэтому потеря воды листом в
процессе транспирации происходит медленнее через устьица, расположенные в
нижнем, а не в верхнем эпидермисе. Во-вторых, главным источником углекислого
газа в атмосфере является «почвенное дыхание», т. е. выделение его в результате
жизнедеятельности почвенных микроорганизмов (бактерий, цианобактерий, грибов и
др.) и дыхания корней высших растений. Поэтому припочвенный слой воздуха обычно
обогащен углекислым газом, который по градиенту концентрации диффундирует вверх
и легко проникает через устьица в ткани листьев.
В центре листа находится крупный проводящий
пучок, а сбоку более мелкие пучки. В составе пучка ксилема повернута к верхней,
а флоэма - к нижней стороне листа. Проводящие пучки с окружающими их тканями
называют жилками. Они образуют в листе непрерывную систему, связанную с
проводящей системой стебля.
Типы корней и корневых систем
Корень - это осевой подземный орган.
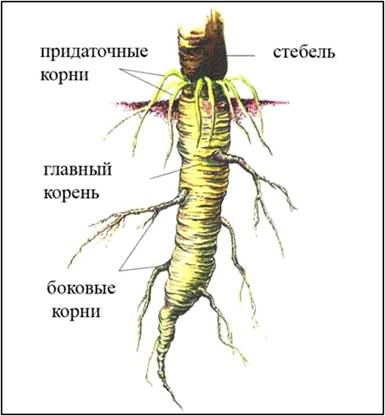
Рис. 2
Основные функции: обеспечивает закрепление
растения в почве, всасывание почвенного водного раствора солей и транспорт его
к надземным частям растения.
Дополнительные функции: запасание питательных
веществ, фотосинтез, дыхание, вегетативное размножение, выделение, симбиоз с
микроорганизмами, грибами. Первые настоящие корни появились у
папоротникообразных.
Зародыш корня называется зародышевым корнем и
закладывается одновременно с почкой в зародыше семени.
У растений различают:
Главный корень. Он образуется из зародышевого и
сохраняется на протяжении всей жизни. Всегда один.
Боковые корни. Ответвляются от корней (главного,
дополнительных, боковых). Образуют при ветвлении корни 2-го, 3-го и т. д.
порядка.
Придаточные корни. Образуются в любой части
растения (стебле, листьях).
Совокупность всех корней растения образует
корневую систему. Корневая система формируется в течение всей жизни растения.
Ее формирование обеспечивают преимущественно боковые корни. Различают два типа
корневой системы: стержневую и мочковатую.
Стержневая система имеет хорошо развитый главный
корень, который развивается из зародышевого и ветвится за счет боковых корней.
Главный корень способен проникать на значительную глубину.
Мочковатая корневая система образована
совокупностью придаточных корней, растущих от стебля, и боковых корней. Главный
корень в ней отсутствует. Формируется мочковатая корневая система во время
кущения. На подземной части стебля при этом образуется узел кущения, из которого
развиваются дополнительные побеги, то есть происходит подземное ветвление
стебля, с многочисленными дополнительными корнями.
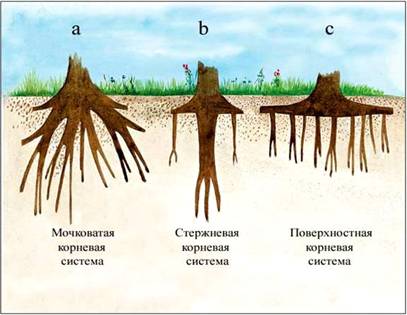
Рис. 3
Тип корневой системы является таксономическим
признаком: для двудольных характерна преимущественно стержневая корневая
система, для однодольных и некоторых травянистых двудольных - мочковатая.
У однодольных растений при прорастании семени
главный корень отмирает или развивается слабо, а из тканей стебля прорастают
дополнительные, которые образуют мочковатую корневую систему.
Поверхность корневой системы значительно больше,
чем поверхность надземной части. У разных видов растений различаются число и
длина корней. Особенно глубоко проникает в почву стержневая корневая система.
Например, корни осота проникают в почву на глубину 6 м, у люцерны посевной - на
10-12 м, у деревьев - еще глубже (свыше 20 м).
Человек использует знания по формированию
корневых систем при пересаживании рассады овощных и декоративных культур. Чем
лучше развита корневая система у растений, тем больше площадь питания,
надземная часть растения, то есть можно получить больший урожай. У проростка
отщепляется кончик главного корня, чтобы усилить ветвление за счет образования
новых, разрастания боковых корней. Этот способ получил название пикирования (от
франц. пика, пикетка- копье).
Свойство растений - разрастание корневых систем
в почве - используют для закрепления оврагов, подвижных песков.
Видоизменения корня. Вследствие обретения новых
функций органы способны видоизменяться.
Корнеплод - утолщение главного корня, связанное
с отложением в нем запаса питательных веществ (морковь, свекла, редис и т. п.).
Корневые клубни (корнеклубни) - утолщение
боковых или дополнительных корней, связанное с отложением запаса питательных
веществ (батат, георгин и т. п.).
Корни-присоски. Характерны для растений
паразитов или полупаразитов. Такие корни проникают в толщу стебля других
растений и потребляют их соки. Повилика - это бесхлорофилльное
растение-паразит, которое питается благодаря сокам растения хозяина. Омела -
полупаразит. Это зеленое растение, способное к самостоятельному питанию
(фотосинтезу), но водные растворы солей она образует от растения, на котором
живет.
Дыхательные корни (пневматофоры). Это боковые
корни, которые растут вверх и поднимаются над поверхностью воды, почвы.
Формируются у растений (мангровые деревья), которые растут на чрезмерно
увлажненных почвах, болотах, с низким содержанием кислорода. Поэтому растения с
помощью таких корней получают кислород непосредственно из воздуха. Дыхательные
корни богаты аэренхимой.
Корни-прицепки.Это дополнительные короткие
корни, которые развиваются на растениях с вьющимся стеблем (плющ, фикус цепкий
и т. п.), которые плетутся вверх. Корни растут на стебле. С их помощью растение
цепляется за трещины, опоры и поднимается выше.
Ходульные корни. Образуются на надземных
побегах. Они закрепляются в почве и помогают растению (баньян, кукуруза и т.
п.) удерживаться.
Воздушные корни. Развиваются у растений
(орхидея), которые поселяются на деревьях, но не паразитируют. Воду и
минеральные соли они получают из воздуха с помощью корней, которые свисают в
воздухе.
Опорные корни. Встречаются у больших деревьев
(вяз, бук, тополь, тропические и т.п.). Представляют собой боковые корни. На
боковых корнях, которые проходят возле поверхности почвы, развиваются плоские
треугольные и прилегающие к стволу вертикальные надземные отростки, которые
напоминают доски, прислоненные к деревьям.
Втяжные, или контрактильные корни. У некоторых
растений происходит резкое сокращение корня в продольном направлении у его
основания (например, у растений, которые имеют луковицы). Втяжные корни
распространены у покрытосеменных растений. Они обусловливают плотное
приляганиек земле розеток (например, у подорожника, одуванчика и т. п.),
подземное положение корневой шейки и вертикального корневища, обеспечивают
некоторое углубление клубней. Таким образом, втяжные корни помогают побегам
находить наилучшую глубину залегания в почве. Втяжные корни в Арктике
обеспечивают переживание неблагоприятного зимнего периода цветочными почками.
Корневые системы способны улучшать свое питание
благодаря взаимодействию с микроорганизмами - грибами, бактериями, водорослями.
Симбиоз корней цветочных растений с грибами называется микоризой, с бактериями
- бактериоризой. Почвенный слой толщиной 2-3 мм вокруг корней растений образует
ризосферу. Корни выделяют в ризосферу вещества, которые привлекают
микроорганизмы.
В клетках корней некоторых растений (бобовые,
березовые и т. п.) поселяются клубеньковые бактерии, которые своими выделениями
вызывают разрастание паренхимы и образование клубеньков на корнях. Клубеньковые
бактерии способны фиксировать атмосферный азот в виде соединений, которые могут
усваиваться растениями (нитратов, нитритов). Часть азотных соединений
усваивается растением, а часть остается в почве. Бобовые растения используют в
сельском хозяйстве для обогащения почв азотистыми соединениями.
Происхождение цветка
Из попыток понять происхождение наиболее
типичного для покрытосеменных обоеполого цветка с так или иначе устроенным
околоцветником родились основные гипотезы происхождения покрытосеменных
растений как таксона.
Псевдантовая теория:
Время: начало XX века. Основатели: А. Энглер, Р.
Веттштейн.
Теория основана на представлении о происхождении
цветковых от эфедроподобных и гнетоподобных голосеменных предков. Была
разработана оригинальная концепция происхождения цветка - идея о независимом
возникновении частей цветка как органов «sui generis». Предполагалось, что
первичными у покрытосеменных были раздельнополые опыляемые ветром цветки с
небольшим и строго фиксированным числом частей, а дальнейшая их эволюция шла по
линии от простого к сложному.
Стробилярная, или эвантовая теория:
Время: конец XVIII века - начало XX века.
Основатели: И.В. Гете, О.П. Декандоль, Н. Арбер и Дж. Паркин.
Согласно этой теории, наиболее близки к искомым
предкам покрытосеменных мезозойские беннеттиты, а исходный тип цветка
представляется сходным с тем, что наблюдается у многих современных
многоплодниковых: обоеполый энтомофильный цветок с удлиненной осью, большим и
неопределенным числом свободных частей. Дальнейшая эволюция цветка в пределах
покрытосеменных имела редукционный характер.
Теломная теория:
Время: с 30-х годов XX столетия. Основатель: В.
Циммерман.
Согласно этой теории, все органы высших растений
происходят и независимо развиваются из теломов; высшие растения с настоящими
корнями и побегами происходят от риниофитов, тело которых было представлено
системой дихотомически ветвящихся простых цилиндрических осевых органов -
теломов и мезомов. В ходе эволюции в результате перевершинивания, уплощения,
срастания и редукции теломов возникли все органы покрытосеменных растений.
Листья семенных растений возникли из уплощённых и сросшихся между собой систем
теломов; стебли - благодаря боковому срастанию теломов; корни - из систем
подземных теломов. Основные части цветка - тычинки и пестики - возникли из
спороносных теломов и эволюционировали независимо от вегетативных листьев.
Микроспорогенез (образование микроспор в гнездах
пыльника). Строение мужского гаметофита (пыльцевого зерна).
Микрогаметогенез - процесс образования мужских
половых клеток (спермиев), происходит в пыльцевом зерне, которое является
мужским гаметофитом покрытосеменных растений. Развитие мужского гаметофита
происходит также в гнездах пыльников тычинок и сводится к одному митотическому
делению микроспоры и формированию оболочек пыльцевого зерна. Оболочка
пыльцевого зерна состоит из двух слоев: интины (внутренней, тонкой) и экзины
(наружной, толстой). Каждое пыльцевое зерно содержит две гаплоидные клетки:
вегетативную и генеративную. Из генеративной (спермагенной) далее образуются
два спермия. Из вегетативной (сифоногенной) впоследствии образуется пыльцевая
трубка.
Тычинка образуется из бугорка конуса нарастания
цветоносного побега. Меристематический тычиночный бугорок вначале вполне
однороден, но в дальнейшем его рост сопровождается дифференцированием.
Дифференцируются и микроспорангии, входящие в
состав тычинки.
Из периферических стерильных клеток образуется
стенка микроспорангия. Эта первоначально однородная стенка со временем
разделяется на три слоя:
внешний слой эндотеций,
средний слой,
самый внутренний выстилающий слой - тапетум.
Клетки эндотеция несут на своей внутренней
стороне специальные подковообразные утолщения, способствующие вскрыванию
зрелого пыльника.
Назначение среднего слоя не совсем понятно.
А вот внутренний слой тапетум (от лат. tapetum -
покрывало) имеет большое значение. Он состоит из таблитчатых тонкостенных
клеток с густой цитоплазмой. Обычно тапетум одноряден. В клетках тапетума
происходит деление путем митоза, вследствие чего они становятся многоядерными.
В дальнейшем ядра могут сливаться и образуются плотные полиплоидные ядра.
Возникает полиплоидная выстилающая ткань.
Тапетум физиологически чрезвычайно активная
ткань: его клетки содержат ферменты, гормоны и питательный материал,
используемый в процессах микроспорогенеза. Т.е. главная функция тапетума -
обеспечение спорогенной ткани питательными веществами.
Параллельно с дифференциацией клеток стенки
микроспорангия происходят события, ведущие к формированию микроспор.
Внутренность каждого пыльника занята первичной
спорогенной тканью. Первичные спорогенные клетки делятся несколько раз обычным
митотическим путем. В результате этих делений образуются материнские клетки
микроспор (или микроспороцисты).
Затем происходит редукционное деление (мейоз),
при этом из каждой материнской клетки образуется тетрада микроспор.
У многих растений при образовании микроспор
клетки тапетума расслизняются, образуют амебоидную ткань и распределяются между
клетками тетрад, способствуя их питанию.
В большинстве случаев в дальнейшем тетрады
распадаются и в гнезде находится беспорядочная масса пыльцы, состоящая из
одиночных, самостоятельных клеток - микроспор.
Правда, иногда тетрады не распадаются, а
остаются соединенными между собой и слипаются вместе в общий пыльцевой ком,
называемый поллиний. Также пыльцевые комки - поллинии характерны для орхидных.
Итак, процесс микроспорогенеза заканчивается
образованием микроспор. Зрелая пыльцевая клетка (микроспора) одноядерна и имеет
сложную оболочку, которая называется спородермой. Она состоит из двух главных
слоев - интины и экзины.
Внутренний слой - экзина представляют собой
утолщенную слоистую оболочку с различными скульптурными выростами в виде
шипиков, бугорков и ямок.
Основным веществом этой оболочки является
углевод - спорополленин. Благодаря споролленину экзина характеризуется
необычайной стойкостью: она не растворяется в кислотах, щелочах, выдерживает t0
до 3000С и сохраняется миллионы лет в геологических отложениях.
Характер скульптурированности поверхности экзины
является важным диагностическим признаком, позволяющим идентифицировать пыльцу
разных видов. Размеры, форма и поверхность пыльцевых зерен настолько
характерны, что по пыльце можно определять виды растений (споровопыльцевой
анализ) палинология. В биологическом плане, эти неровности помогают пыльце
удерживаться на рыльцах плодолистиков.
Как правило, в экзине пыльцевого зерна имеются
тонкие или даже перфорированные места. Эти места называются апертурами и имеют
очень важное значение - они служат для выхода пыльцевой трубки.
Интина представляет собой внутренний слой
оболочки пыльцевого зерна. Она облегает его содержимое и служит материалом,
образующим пыльцевую трубку. В ее составе преобладают пектиновые вещества.
Понятно, что эта оболочка весьма нестойкая и легко разрушается.
Вслед за микроспорогенезом здесь же в пыльцевых
зернах протекает микрогаметогенез.
Ботаническая характеристика рода
Паслен
Семейство пасленовые (SOLANACEAE)
В семействе около 90 родов и не менее 2500
видов, широко распространенных в тропических, субтропических и умеренных
областях, главным образом в Центральной и Южной Америке. Представители
семейства - травы, кустарники или небольшие деревья с очередными (иногда в
области соцветия супротивными), простыми листьями. Цветки обычно в пазушных
верхоцветных соцветиях, обоеполые, актиноморфные или реже слегка зигоморфные.
Чашечка обычно 5-лопастная или 5-раздельная, остающаяся, часто при плодах
увеличенная. Венчик от колесовидного до трубчатого, 5-лопастный, редко
двугубый. Тычинок обычно 5 или в зигоморфных цветках меньше (4-2); пыльники
вскрываются продольно или верхушечными порами. Нектарный диск обычно развит.
Гинецей обычно из 2 плодолистиков, редко из 5 плодолистиков, обычно с
верхушечным простым столбиком с двулопастным рыльцем; завязь обычно двугнездная
(иногда ложно-3 или 5-гнездная) или редко 5-гнездная, обычно с многочисленными
семязачатками. Плод - ягода или септицидная коробочка, редко плод
распадающийся. Семена с эндоспермом. Цветки пасленовых опыляются различными
насекомыми, а в тропических странах также птицами, а иногда даже
млекопитающими.
Семейство пасленовых подразделяют на 2
подсемейства - нолановые (Nolanoideae) и собственно пасленовые (Solanoideae).
Первое нередко рассматривают как самостоятельное семейство нолановые
(Nolanaceae). Подсемейство нолановых относительно более примитивно, чем
пасленовые. Оно включает 2 близких рода - нолана (Nolana, около 75 видов,
распространенных от Перу до Патагонии и на Галапагосских островах) и алона
(Alona, 5-6 видов в Чили). Это травы или маленькие кустарнички с очередными,
цельными, более или менее суккулентными листьями, произрастающие главным
образом вдоль морских побережий. Гинецей у них из 5 плодолистиков.
Все остальные роды семейства пасленовых входят в
обширное подсемейство пасленовых. Оно, в свою очередь, подразделяется на 5
триб. Самой примитивной трибой считается триба никандровые (Nicandreae),
состоящая из одного монотипного рода никандра (Nicandra), обитающего в Перу и
Боливии. Это однолетняя трава с грубыми ребристыми жестко опушенными стеблями,
зубчатыми или лопастными листьями, одиночными цветками, 3-5-гнездной завязью и
довольно многосемянной ягодой, заключенной в сильно разрастающуюся при плодах
чашечку.
Самой большой трибой семейства пасленовых
является триба собственно пасленовых (Solaneae), куда входит несколько десятков
родов. Завязь у них 2-гнездная, редко многогнездная (например, у томата).
Триба, в свою очередь, подразделяется на несколько подтриб, из которых самая
примитивная - подтриба дерезовых. Сюда входят деревья, кустарники или травы.
Наиболее известный и самый крупный род дереза (Lycium) заключает около 100
видов листопадных или вечнозеленых прямостоячих или вьющихся, обычно колючих
кустарников, распространенных в тропических, субтропических и отчасти умеренных
областях, главным образом в Южной Америке. В нашей стране 7 видов в пустынях,
полупустынях и степях им юго-востоке европейской части, на Кавказе и в Средней
Азии. Некоторые виды разводят для изгородей и как декоративные. К подтрибе
дерезовых относится и такой широко известный род, как красавка, или атропа
(Atropa), состоящий из 4 видов, распространенных от Европы и Средиземноморья до
Индии. В трибе пасленовых отдельную подтрибу составляют также роды скополия
(Scopolia) и белена (Hyoscyamus). Многие виды скополии - скополия светло-желтая
(S. lurida), обитающая в Гималаях, и скополия тангутская (S. tangutica),
произрастающая в Тибете, содержат, кроме того, алкалоид атропин. Все 3 вида
возделывают как ценные лекарственные растения. В роде белена (Hyoscyamus) около
20 видов, произрастающих на Канарских островах, в Европе, Северной Африке (до
центральной части), Западной и Средней Азии. В России - 8 видов.
Подтриба пасленовых занимает центральное
положение в одноименной трибе семейства. Здесь нужно прежде всего упомянуть
довольно большой род физалис (Physalis), около 100 видов которого широко
распространены в тропических, субтропических и отчасти умеренных областях,
преимущественно в тропической Америке. Физалисы характеризуются очень крупной
пузыревидно вздутой при плодах, красной или оранжевой чашечкой.
Но, безусловно, самым важным для человека
является самый большой в семействе род паслен (Solanum), насчитывающий около
1700 видов, т. е. больше половины видового состава всего семейства. Он широко
распространен в тропических, субтропических и умеренных областях обоих
полушарий, но главным образом в Южной Америке. Это многолетние, реже однолетние
травы, полукустарники с прямостоячими или вьющимися стеблями, иногда небольшие
деревья. Плод 2-гнездная многосемянная ягода. В нашей стране около 20 дикорастущих
видов паслена. К роду паслен относится ряд важнейших для человека культурных
растении. Первое место среди них занимает картофель (от немецкого слова
Kartoffel). В культуре известно в основном 2 близких вида - картофель андийский
(S. andigena), издавна возделываемый на территории Колумбии, Эквадора, Перу,
Боливии и Северо-Западной Аргентины, и наш обыкновенный картофель.
И обширную трибу пасленовых, но в качестве
отдельной подтрибы входят южноамериканский род цифомандра (Cyphomandra) и
средиземноморско-азиатский род мандрагора (Mandragora). В роде мандрагора около
6 видов, распространенных от Пиренейского полуострова до Восточных Гималаев и
Тибета. Большинство видов мандрагоры представляют собой многолетние травы,
почти всегда бесстебельные, с очень крупными листьями в розетке, достигающей в
диаметре 1-2 м и более. Мясистые, богатые крахмалом корни мандрагор обладают
своеобразным ветвлением: подчас корень дает два вертикальных отростка и
несколько напоминает фигуру человека. Из-за этой своей особенности мандрагора с
древнейших времен овеяна легендами, приписывающими ей магическую силу. Кроме
того, она содержит алкалоид гиосциамин и в средние века считалась одним из
ценнейших лекарственных снадобий. В отличие от бесстебельных средиземноморских
видов мандрагоры гималайско-тибетская мандрагора стеблевая (М. caulescens)
имеет развитый стебель и более мелкие листья. В то же время ближайший
родственный ей вид -мандрагора тибетская (М. tibetica) - представляет собой
мелкое плотнорозеточное растение. Как можно предположить, в данном случае
приспособительная эволюция шла по пути фиксации ювенильной фазы вегетативной
сферы.
Далее в системе подсемейства пасленовых следует
небольшая триба дурмановых (Datureae), характеризующаяся тем, что в результате
развития двух ложных перегородок, разделяющих каждую из двух плацент
первоначально двугнездной завязи, завязь становится как бы четырехгнездной.
Плод у дурмановых - коробочка или ягода. Наиболее известным представителем этой
трибы является род дурман (Datura), насчитывающий около 10 видов многолетних
или однолетних трав, обитающих в тропических и тепло-умеренных странах, главным
образом в тропической Америке. Цветки у видов дурмана крупные, с белый
воронковидным венчиком длиной от 6 до 20 см и более. Все они отличаются крупными
длиннотрубчатыми цветками с дурманящим запахом и опыляются преимущественно
длиннохоботковыми ночными бабочками, которые отовсюду летят на запах дурмана,
пренебрегая другими растениями. В процессе эволюции удлинение трубки венчика у
многих представителей рода шло параллельно с увеличением длины хоботка
некоторых бабочек бражников (Sphingidae). В результате возникли виды дурмана,
которые могут опыляться лишь определенными видами бражников, попадая в полную
от них зависимость. Плоды дурманов - оригинальные шиповатые коробочки,
раскрывающиеся четырьмя створками. Эти ядовитые растения содержат ряд
алкалоидов и находят применение в фармакопее, а как анестезирующие были
известны еще древним перуанцам. Местами виды дурмана культивируют в качестве
декоративных. В южных районах бывшего СССР культивируются дурман безвредный (D.
innoxia) родом из Америки и дурман индийский (D. metel) родом из Юго-Западного
Китая, незрелые плоды которых содержат скополамин.
К дурману очень близок и нередко объединяется с
ним южноамериканский род бругмансия (Brugmansia), состоящий из 5 видов
кустарников или небольших деревьев с очень крупными длиной до 30 см кожистыми
листьями. Огромные яркие трубчатые цветки, висящие на длинных цветоножках,
утром раскрываются, а днем, в жару, снова захлопываются, что в полной мере
используют для себя виды дрозофилы. Мушки собираются в венчике сразу после его
раскрывания. Несмотря на то что дрозофилы находятся в цветке поблизости от
пыльников, они, по-видимому, участия в опылении не принимают. По истечении
нескольких дней венчик цветка, населенного мушками, опадает и вместе с ним
падают на землю подросшие личинки дрозофил, продолжая питаться оставшейся в
венчике пыльцой. Вероятно, плодовые мушки завершают свой жизненный цикл вместе
с цветком приютившей их особи. Г. К а р с о н, который наблюдал определенные
виды дрозофил, поселяющиеся в цветках бругмансии белоснежной (В. Candida),
считает, что эти виды плодовой мушки могут существовать только в сожительстве с
бругмансией. Опыляются цветки более крупными насекомыми, способными проникать в
цветок снизу вверх. Удлиненные мягкие или несколько деревянистые
нераскрывающиеся плоды бругмансии содержат крупные клиновидные семена. В
качестве декоративного растения широко известен «древовидный дурман»,
представляющий собой гибрид между бругмапсией белоснежной и бругмансией
древесной (В. arborea).
Из других представителей трибы следует отметить
чрезвычайно интересный род соландра (Solandra), около 10 видов которого
распространены в тропической Америке. Большинство соландр представляет собой
длинно- и толстостебельные лианы, обитающие в тропических горных лесах и
обвивающие крупные деревья. Взбираясь по их стволам на большую высоту, лиана
распластывает на верхушке кроны дерева простые цельнокрайние кожистые листья и
подставляет солнцу крупные красивые, слегка зигоморфные колокольчатые цветки на
толстых цветоножках. Многосемянные ягоды растений, свободно охваченные
разросшейся яркой чашечкой, хорошо заметны сверху на поверхности кроны
дерева-хозяина и поэтому охотно поедаются птицами, которые и разносят на
дальние расстояния их почковидные мелкие семена. Некоторые особи соландры в
преклонном возрасте почти или совершенно теряют связь с почвой, продолжая
функционировать как полуэпифиты или как настоящие эпифиты. Наиболее красиво
цветущие виды рода культивируют в тропических садах и оранжереях как
декоративные. Особенно популярна соландра крупноцветковая (S. grandiflora),
оплетающая живым ковром заборы и стены домов.
Интересно, что опыление некоторых представителей
трибы дурмановых осуществляется не только различными насекомыми, по также и
летучими мышами. Это прослежено для рода трианея (Trianaea), 3 вида которого
распространены в горных лесах Северных Анд. У этих растений довольно крупные
цветки располагаются на концах длинных висячих ветвей и выделяют обильный
нектар. Цветки раскрываются вечером, испуская резкий неприятный запах,
привлекающий летучих мышей.
Триба цестровых (Cestreae) получила наименование
по наиболее заметному роду цеструм (Cestrum), объединяющему около 150 видов,
обитающих в тропических и субтропических областях Америки. Это кустарники или
небольшие деревья с цельными, большей частью узкими, часто опушенными листьями.
Их цветки, собранные в верхушечные или пазушные соцветия с маленькой чашечкой и
длинным воронковидным или трубчатым венчиком, открываются в основном по ночам,
издавая сильный запах. Некоторые виды цеструма, как цеструм полевой (С.
campestre), опыляются колибри. Плоды цеструмов - продолговатые ягоды.
К данной трибе принадлежит также и род табак, или
никотиана (Nicotiana), насчитывающий 66 видов, из которых 45 обитают во
внетропических частях Америки, а 21 вид приурочен к Австралии и Полинезии. К
табакам близок род петуния (Petunia), представленный около 30 южноамериканскими
видами; некоторые из них повсеместно культивируются как декоративные. У нас
наиболее известна петуния гибридная (P. hybrida).
Крайне своеобразен относящийся к цестропым род
маркея (Markea), 18 видов которого обитают по преимуществу в лесах тропической
Америки - от бассейна реки Амазонки до Мексики. Некоторые виды маркеи
представляют собой лианы с длинными изящными стеблями и простыми цельнокрайними
листьями, собранными в пучки на концах веточек. Обвиваясь вокруг опоры, стебли
маркеи вползают по деревьям все выше и выше, образуя в верхней части кроны
дерева-хозяина терминальные соцветия с бархатистыми короткотрубчатыми
ак-тиноморфнымн, часто пурпурно-зелеными цветками. С. Фогель в 1958 г.
проследил, что цветки некоторых видов маркеи, такие, как маркея дресслери (М.
dressier!), закрыты в дневное время и открываются только по ночам, привлекая
своим запахом местных лесных крыс, которые и являются основными опылителями
этих растений.
Очень близок к роду маркея как в систематическом
отношении, так и по образу жизни род хуануллоа (Juanulloa). Растения,
принадлежащие к этому роду, отличаются от видов рода маркея своими узкими
трубчатыми опушенными цветками. Среди 12 видов хуануллоа, распространенных от
Мексики до Боливии, имеются не только эпифиты, но даже и полупаразиты, живущие
на деревьях. Обладая вполне развитыми и нормально функционирующими зелеными
листьями, полупаразитные растения образуют гаустории, которые постепенно
проникают в древесину молодых ветвей дерева-хозяина, высасывая из сосудов воду
с растворенными в ней минеральными солями.
Наиболее подвинутой трибой семейства является
триба сальпиглоссовых (Salpiglossideae). Растения, принадлежащие к ней, часто
обладают зигоморфными цветками с 2-4 фертильными тычинками и септицидными
раскрывающимися плодами типа коробочки с одревесневшей наружной эпидермой.
Одним из примечательных родов трибы является схизантус (Schizanthus) -
эндемичный чилийский род, насчитывающий около 10 видов. От остальных пасленовых
он отличается ясно выраженной двугубостью крайне зигоморфного венчика. Из 5
тычинок только 2 фертильные, 2 превращены в стаминодии, а пятая очень
редуцирована. Интересен механизм опыления цветков: две фертильные тычинки
располагаются внутри губы, образованной двумя нижними лопастями венчика. Когда
пчела или бабочка опускается на эту губу, пыльники энергично взрываются,
выстреливая пыльцу в воздух. Некоторые из пыльцевых зерен обычно попадают на
насекомое, которое переносит их к другому цветку. Некоторые виды этого рода
издавна культивируют как декоративные.
Другой, заслуживающий внимания род
сальпиглоссовых - брунфельсия (Brunfelsia) - состоит из 40 видов,
произрастающих в тропической Америке. Это широко распространенные в тропической
зоне декоративные деревья или кустарники, цветущие в сезон дождей и названные
поэтому «дождевыми деревьями». Листья брунфельсии целытокрайние, кожистые,
расположенные чаще на концах веточек, иногда собранные в пучки. Цветки слегка
зигоморфные, синие, пурпурные или белые, с 4 фертильными тычинками. С возрастом
цветки меняют окраску, как у бурачниковых. Плоды большинства видов - сочные или
кожистые ягоды, окруженные слегка разросшейся чашечкой и содержащие крупные
призматические семена с мясистым эндоспермом. Некоторые из видов весьма
декоративны. Род сальпиглоссис (Salpiglossis), давший название трибе, в количестве
5 видов обитает в Южной Америке. Салъпиглоссис выемчатый (S. sinuata)
встречается как декоративное в культуре.
Прекрасное и удивительное семейство пасленовых
объединяет в себе самые разнообразные и порой неожиданные виды, к которым
относятся как ядовитые и опасные растения, такие как мандрагора, белладонна,
дурман, белена и бругманзия - отравительница бабочек, так и обычные самые что
ни на есть съедобные овощи, такие как картофель, баклажан, помидор, сладкий и
острый перцы. Кроме того, семейство пасленовых включает в себя ароматные
декоративные цветы. Это и общеизвестные петуньи, скромняга - табак, брунфельсии
и цеструмы с чудесным удушающим запахом, красные и пурпурные колокольчики
иохромы, соландры, белые и лиловые солянумы и многие другие… Кроме этих душистых
растений существует еще множество бесчисленных пасленов - черный горько-сладкий
мексиканский табак, фонарики фезалисов и много прочих растений.
Свое название семейство пасленовых взяло от
крупного рода паслен Solanum, которое включает в себя примерно сто родов и
почти три тысячи видов растений, которые благополучно произрастают по всему
земному шару в разнообразных климатических зонах и условиях. Паслены
представляют собой травы, кустарники, небольшие деревья с самым разнообразным
внешним видом: с лежащими, стоящими, ползучими стеблями, с простыми или
перистыми листьями. Такие разные растения объединяет одна общая черта: все
пасленовые (не исключая и съедобные) в большей или меньшей степени ядовиты.v
Наше знакомство с этими удивительными созданиями
начнется с самых обыкновенных из них, растущих где придется и часто при
дорогах.
Паслен черный (Solanum nigrum) - травянистое,
покрытое волосками однолетнее растение, что растет на пустырях, свалках, на
обочинах дорог и на опушках лесов. Широко распространен в климатических зонах
от умеренного к южному. Листья овально-удлиненные незазубренные, бледноватые
белые цветы с желтым пыльником, собранные в небольшие кисти, примерно, по пять
цветков в каждой. Плоды похожи на ягоды, сначала зеленые, а при созревании - черные,
содержащие алкалоид соланин. При употреблении большого количества плодов
черного паслена возможно сильное отравление, однако, при правильном применении
может использоваться в медицинских целях. В народной медицине и аюрведе
используется при циррозе печени и хронических кожных заболеваниях, таких как
псориаз и лишаи. Имеющий противовоспалительный эффект, паслен черный
применяется также при лихорадках и для снятия болей. Обладает антисептическими
свойствами и может также использоваться в качестве глистогонного средства.
Паслен горько-сладкий (Solanum Dulcamara)
представляет собой полукустарник, с длинными ползучими ветвями, покрытыми
продолговатыми заостренными, у основания сердцевидными, листьями. В отличие от
паслена черного, его фиолетовые цветки довольно яркие и крупные, собраны в
слегка поникающие кисти, из которых впоследствии получаются красные, как
маленькие помидорчики, плоды. Растет горько-сладкий паслен на влажной почве, в
кустарниках. Отвар из его стеблей употребляют от лишаев, ревматизма и катаров.
Однако ягоды ядовиты и в фармакологии не используются.
Еще одни паслены, но не придорожники: паслен
ложноперечный и перцевидный.
Паслен ложноперечный (Solanum pseudocapsicum)
именуемый иногда иерусалимской вишней - уроженец Его острова Мадейра, где он
растет повсеместно, как у нас паслён чёрный. В диком виде у нас это растение
встречается в Крыму. Это вечнозеленый прямостоячий кустарник высотой больше
метра. Его листья длиной около десяти сантиметров, ланцетные или овальные,
слегка волнистые, без опушки. Цветы белые, одиночные или в малоцветковых
кистевидных соцветиях, плоды ягодообразные, округлые, оранжево-красные, реже
желтые, до полутора сантиметров в диаметре. Плоды его несъедобные, невкусные и
ядовитые, содержащие ядовитый алкалоид соланин. Паслен ложноперечный в народе
называют "ягоды любви", по поверью, если бесплодные супруги съедят по
ягодке, у них немедленно появятся дети.
Паслен перцевидный (Solanum capsicastrum), или
звездчатый перец, в целом, сходен с предыдущим видом, но меньше по размерам и
имеет гораздо меньшие плоды. Молодые побеги у этого растения имеют сероватую
опушку и более короткие листья. Родина паслена перцевидного - Южная Бразилия и
Уругвай, где он произрастет в диком виде в сельве. Обильно цветет летом
небольшими белыми цветочками, которые красиво покрывают все растение. К зиме
созревают ярко-красные плоды размером с вишню. Выведены также карликовые
разновидности этого растения с плодами разных оттенков: от ярко-оранжевого до
темно-красного, есть сорта с белыми и ярко-красными плодами.
Данное растение очень популярно во многих
странах и в огромных количествах продается перед Рождеством, когда кустик
покрывается россыпью ярких ягод и становится необыкновенно нарядным. Хотя
паслен перцевидный считается чисто декоративным растением, существуют сведения
о его целебных свойствах и даже встречается его народное название -
"ангинник". Целители рекомендуют столовую ложку толченых сухих ягод
настаивать два часа в стакане кипятка, а затем этим настоем полоскать горло.
И еще один ярчайший представитель данного
семейства - паслен сосочковый (Solanum mammosum), известный как Коровье вымя
или Содомово яблоко. Как все пасленовые, любит тепло, от умеренного до весьма
жаркого, однако не против и небольшой тени. Вырастает высотой более метра, листья
мягкие, напоминающие небольшие лопухи, на довольно толстых стеблях имеются ярко
выраженные колючки. Цветет небольшими цветочками, чаще фиолетовыми, но
встречаются разновидности с белыми цветами. После цветения образует восковидные
желтые или оранжевые плоды, напоминающие небольшие вытянутые помидоры.
Окончания плодов похожи на коровье вымя с сосками. Ветви этого паслена,
украшенные необычными золотистыми плодами, очень декоративны, их часто срезают
для аранжировки букетов, тем более что в срезанном состоянии они сохраняют свой
внешний вид довольно долго. Паслен сосочковый растение редкое, поэтому
заполучить его в букет считается у флористов особым шиком. Коровье вымя -
растение несъедобное, однако, имеет применение в гомеопатии. Там его используют
в виде микстуры и применяют при кровохарканье и просто в качестве
отхаркивающего средства.
Список сорняков
Семейство Злаковые (Gramineae Juss.).
Овсюг, овес пустой (полетай) (Avena fatua L.).
Просо куриное, ежовник, петушье просо
(Echinochloa crus galli L. Beauv; Panicum crus galli L.).
Щетинник зеленый (обыкновенный), мышей зеленый.
(Setaria viridis (L.) Beauv).
Щетинник сизый, или малый, мышей сизый, брица
(Setaria glauca L. Beauv.; Setaria pallidefusca Schum.
Костер ржаной (Bromus secalinus L.).
Пырей ползучий (Agropyrum repens, Elytrigia
repens (L.) Nevski).
Свинорой пальчатый (пальчатая трава) (Cynodon
dactylon (L.) Pers.).
Гумай, сорго алепское (Sorghum halepense (L.)
Pers.).
Тростник обыкновенный, камыш (Phragmites
australis (Cav.) Trin. Ex Steud.).
Семейство Гречишные (Ро1уgоnасеае Juss.).
Горец шероховатый, гречишка развесистая (горчак,
плюшка) (Ро1уgonum lapathifolium (L.) Moench; P. tomentosum Schrank.).
Горец вьнковый, гречишка вьнковая, повитель
(Ро1уgonum convolvulis (L.).
Горец птичий, гречиха птичья (спорыш, гусятник,
конотоп, трава-мурава) (Р. aviculare L.).
Гречиха татарская, курлык (Fagopyrum tataricum
L. Gaertn.).
Семейство Маревые (Сhenopodiaceae Vent.).
Марь белая, марь обыкновенная (лебеда)
(Сhenopodium album L.).
Семейство Гвоздичные (Саryophyllaceae Juss.).
Торица полевая, обыкновенная (Spergula arvensis
L.).
Куколь обыкновенный (посевной) (Agrostemma
githago (L.).
Семейство Мареновые (Rubiaceae Juss.).
Подмаренник цепкий (Galium aparine L.).
Семейство Мальвовые (Ма1vaceae Juss.).
Канатник Теофраста (Abution theophrasti Medik.).
Просвирник обыкновенный (пренебреженный),
«калачики» (Malva neglecta Wallr.).
Гибискус тройчатый
(вздутый)
(Нibiscus
trionum L.; H. ternatus Cav.; H. vesicarius Bess.).
Семейство Фиалковые
(Violaceae Batsch).
Фиалка полевая
(Viola arvensis Murr.).
Семейство Амарантовые
(Аmаrаnthасеае
Juss.).
Щирица запрокинутая
(обыкновенная,
колосистая),
подсвекольник,
краснуха
(Amaranthus retroflexus L.).
Семейство Пасленовые (Sо1аnасеае Juss.).
Паслен черный (Solanum nigrum L.).
Белена черная (Hyosciamus niger (L.).
Семейство Лютиковые (Ranunculaceae).
Живокость полевая (посевная), консолида
великолепная, сикирки, василек рогатый (Delphinium consolida L.).
Семейство Крестоцветные (Cruciferae Juss.), или
Капустные (Brassicaceae Burnett).
Горчица полевая
(Sinapis arvensis L.).
Редька полевая
(дикая)
(Raphanus raphanistrum L.).
Пастушья сумка
обыкновенная
(Capsella bursa-pastoris (L.) Medik).
Желтушник левкойный (лакфиолевый) (Erysimum
cheiranthoides L.).
Ярутка полевая, денежник (Thlaspi arvense L.).
Дескурайния (дескурения) Софьи, гулявник
струйчатый (Descurainia Sophia (L.) Webb ex Prantl; Sisymbrium Sophia L.).
Сурепка обыкновенная (Barbarea vulgaris R. Br.).
Семейство Гераниевые (Geraniaceae).
Аистник цикутовый (цапельник, журавельник)
(Erodium cicutarium L. L` Her.)
Семейство Губоцветные (Labiatae Juss.) или
Яснотковые (Lamiaceae Linal.).
Пикульник заметный (красивый), зябра (Galeopsis
speciosa Mill.).
Пикульник обыкновенный, жабрей (Galeopsis
tetrahit L.).
Семейство Сложноцветные (Compositae Gisee) или
Астровые (Asteraceae Dumort).
Василек синий (Centaureae cyanus L.).
Мелколепестник канадский (Erigeron сanadensis
(L.).
Трехреберник непахучий, ромашка непахучая,
продырявленная (Matricaria inodora L., Tripleurospermum perforatum (Merat) M.
Lainz).
Ромашка душистая, пахучая, хамомилла, лепидотека
(Matricaria matricarioides (Less.) Porter, Lepidotheca (Chamomilla) suaveolens
(Pursh). Nutt.).
Циклахена (ива) дурнишниколистная (Ciclachaena
xanthiifolia (Nutt.) Fresen.
Крестовник обыкновенный (Senecio vulgaris L.).
Дурнишник обыкновенный, (зобовидный) (Xanthium
strumarium L.).
Одуванчик лекарственный (Taraxacum officinale
Wigg.).
Бодяк полевой (розовый) (Cirsium arvense (L.)
Scop.).
Осот полевой ( желтый) (Sonchus arvensis L.).
Осот огородный (Sonchus oleraceus (L.)
Латук татарский, молокан (Lactuca tatarica (L.)
C.A. Mey; Mulgedium tataricum D.C.).
Полынь обыкновенная, чернобыльник (Artemisia
vulgaris L.).
Полынь горькая (Artemisia absinthium L.)
Цикорий обыкновенный (Cichorium intybus L.).
Семейство Заразиховые (Оrobanchaceae Juss.).
Заразиха подсолнечная (кумская), волчок
(Orobanche cumana Wallr.).
Семейство Бобовые (Fabaceae (Lindley);
Leguminosae (Juss).
Донник желтый (лекарственный) (Melilotus
officinalis (L.) Pall.; Melilotus arvensis Wallr.).
Горошек мышиный (Vicia cracca (L.).
Семейство Сельдерейные (Зонтичные) (Apiaceae
(Umbelliferae).
Резак обыкновенный
(Falgaria vulgaris Bernh.).
Семейство Бурачниковые
(Boraginaceae).
Чернокорень лекарственный
(Cynoglossum officinale (L.).
Семейство Хвощовые
(Eguisitaceae Rich. ех
DС)
Хвощ полевой (Eguisetum arvense L.).
Семейство Вьюнковые (Convolvulaceae Juss.).
Вьюнок полевой, березка (Convolvulus arvensis
L.).
Щавель малый (кисловатый), щавелек (Rumex
acetosella L.).
Семейство Губоцветные (Labiatae Juss.) или
Яснотковые (Lamiaceae Linal.).
Мята полевая (Mentha arvensis L.).
Чистец болотный (Stachys palustris L.).
Характеристика семейства Маковые,
представители и значение этого семейства
Семейство маковые (Papaveraceae)
Семейство маковых объединяет около 45 родов и до
700 видов, распространенных главным образом в северной умеренной зоне.
Наибольшее количество видов (более 300) насчитывается в роде хохлатка
(Corydalis, табл. 31). Несколько меньшим по объему (около 100 видов) является
род мак (Papaver). Наиболее богаты родами и видами маковых умеренные и
субтропические области Старого и Нового Света. Небольшое число видов растет в
Южной и Юго-Восточной Африке, а также в Австралии. Значительно реже маковые
встречаются в Арктике, а в тропиках они приурочены, как правило, к
высокогорьям. Среди маковых много эндемичных родов. Так, сангинария
(Sanguinaria) является эндемиком Атлантическо-Северо-Американской
флористической области, эндемики Восточно-Азиатской области - гиломекон
(Hylomecon), маклея, птеридофиллум (Pteridophyllum). В Мадреанской
флористической области произрастают ромнея (Romneya) и дендромекон
(Dendromecon). Только в Голарктической области встречается род гипекоум
(Hypecoum). Виды этого рода произрастают в Средиземноморье, Юго-Восточной
Европе, Средней Азии, Сибири, Гималаях и Китае.
Представители семейства маковых, независимо от
того, в какой климатической зоне они встречаются, часто предпочитают места с
недостаточным увлажнением. Чаще всего они растут в степях, полупустынях и
пустынях. В Арктике и высокогорьях, где степень увлажненности значительно выше,
маковые чаще всего селятся на сухих пригорках, по каменистым склонам с хорошо
дренированными почвами.
В то же время в семействе есть растения и более
влажных местообитаний. Это чистотел (Chelidonium) и сангинария. Лесной мак
гиломекон, некоторые виды хохлатки и дымянки (Fumaria, табл. 33) произрастают
под пологом леса, а также на альпийских лугах и у тающего снега (виды
хохлатки).
В пределах семейства наблюдается большое
разнообразие жизненных форм: от травянистых одно- и многолетников, составляющих
подавляющее большинство, до кустарников (виды бокконии - Bocconia и маклеи) и
даже небольших деревьев (например, боккония древовидная - В. arborea - в горах
Мексики). Некоторые виды рода адлюмия (Adlumia) являются лианами. У них тонкий,
извилистый стебель, длиной до 3 м, цепляющийся за опору с помощью вьющихся
черешков листьев.
Листья у маковых простые, очередные или самые
верхние почти супротивные либо мутовчатые, без прилистников. Прикорневые листья
часто образуют густую розетку. Форма листовой пластинки весьма разнообразна.
Например, у меконопсиса (Meconopsis, табл. 33) листья почти цельнокрайние,
иногда городчатые, продолговатые; у большинства маков - тонко
перисторассеченные; листья бокконии и маклеи глубоко выемчатые, пальчатые. Род
птеридофиллум обладает перисторассеченными листьями, напоминающими по форме
вайи некоторых папоротников, чем и объясняется латинское название этого рода.
Цветки маковых иногда собраны в верхоцветные
соцветия. Кистевидные соцветия встречаются у дымянок и хохлаток, метельчатые -
у бокконии, маклеи, птеридофиллума, гипекоума. Однако большинство
представителей семейства имеет одиночные цветки на длинных, прямостоячих,
безлистных цветоносах, обоеполые, актиноморфные или реже зигоморфные (например,
у дымянки, хохлатки, адлюмии и дицентры - Dicentra).
У всех маковых имеется опадающая чашечка,
состоящая из 2 или 3 чашелистиков. В некоторых случаях она образует до начала
цветения замкнутое вместилище, в котором находятся сморщенные, черепитчато
уложенные лепестки бутона. Это наблюдается у всех маков и близких к ним родов.
У дымянок чашелистики более мелкие, треугольные и замкнутого вместилища не
образуют. Бутоны чаще всего до цветения поникающие.
Венчик цветков маковых, если он есть, состоит из
4, 6 или 8-12 (до 10) лепестков, расположенных в два круга. Исключением
являются цветки бокконии и маклеи, лишенные лепестков. У маков, аргемоны,
ромнеи, эшольции, меконопсиса и многих других маковых лепестки наружного и
внутреннего кругов цельные, без нектарников, не отличаются друг от друга по
форме и размерам. Иногда лепестки внутреннего круга несколько мельче, как это
бывает, например, у чистотела. У гипекоума наружные лепестки обычно цельные или
слабо трехлопастные, внутренние лепестки трех раздельные, крайние доли гладкие,
а средняя с бахромчатым краем. Четыре лепестка зигоморфных цветков дымянки
(табл. 33), хохлатки (табл. 31), адлюмии и дицентры также расположены в два
круга. У дицентры и адлюмии наружные лепестки одинаковые, несущие при основании
шпоровидный или мешковидный отросток. У хохлатки, дымянки и дымяночки только
один верхний лепесток имеет резко выраженную шпору; лепестки внутреннего круга
обычно слипаются между собой верхушками, а на спинках имеют выделяющиеся,
прижатые кили.
Нектарники примитивного строения имеются у
гипекоума. Это нектарная ямка, находящаяся у основания средней доли внутренних
лепестков, свободная для доступа любых насекомых. Более специализированные
нектарники присутствуют у дицентры, адлюмии, хохлатки, дымянки и дымяночки.
Здесь нектар скапливается в шпоровидных или мешковидных отростках наружных
лепестков, причем сами железки, выделяющие нектар, расположены у основания
тычинок.
Тычинки чаще всего многочисленные, редко их
6-12, очень редко - 4 (птеридофиллум, гипекоум). У большинства маковых тычинки
свободные, лишь у дымянок и близких к ним родов 6 тычинок срастаются по 3 в два
пучка, образуя так называемые «фаланги». Средние из них, супротивные боковым
лепесткам, с нормально развитыми 4-гнездными пыльниками, а 4 краевые,
чередующиеся с лепестками, лишь с половинками пыльников. Пыльники экстрорзные,
вскрываются продольно.
Пыльца трехбороздная, рассеянно-многобороздная
или многопоровая. Экзина зернистая, сетчатая или бугорчатая.
Гинецей паракарпный, из 2 или 3-20
плодолистиков. Два плодолистика характерны для гипекоума, дымянки, хохлатки и
близких к ним родов; многочисленные - для остальных маковых. Завязь верхняя или
почти полунижняя (эшольция), одногнездная или ложномногогнездная в результате
врастания внутрь полости и соединения плацент, редко двугнездная благодаря
образованию ложной перегородки, с двумя (хохлатка, дымянка) или многочисленными
семязачатками.
Подавляющее большинство представителей семейства
маковых являются насекомоопыляемыми растениями. Как уже упоминалось,
большинство маковых имеет крупные цветки с обильной пыльцой. Пыльники в цветке,
как правило, созревают раньше завязи, что обеспечивает перекрестное опыление.
Пыльца, продуцируемая в огромных количествах, высыпается на лепестки. Это
привлекает множество насекомых, особенно мух и мелких жучков. При этом
сравнительно тяжелые насекомые - шмели и жуки - предпочитают садиться на
своеобразную посадочную площадку - широкие сидячие рыльца, такие, как у маков и
аргемоны. Кроме того, цветки маков, аргемоны и некоторых видов ромнеи
используются многими жуками и мухами как ночное убежище от холода.
В других случаях насекомых привлекает нектар.
Например, цветки гипекоума посещают пчелы, мухи и некрупные бабочки. Нектар в
цветках хохлатки, дымянки, дицентры, адлюмии доступен только насекомым с
длинным хоботком - медоносной пчеле, садовому шмелю и некоторым видам бабочек.
Хохлатка, дымянка и дымяночка, цветущие ранней весной, чаще всего опыляются
самцами комаров и ранневесенними мухами-журчалками.
Так протекает процесс перекрестного опыления в
хорошую погоду и в случае изобилия опылителей. Однако при неблагоприятных
условиях у многих маковых может происходить и самоопыление. Это хорошо известно
на примере видов мака, аргемоны, эшольции и некоторых других растений.
После раскрывания пыльников пыльца попадает на
лепестки. С наступлением сумерек или перед дождем цветки маковых, как правило,
закрываются. Благодаря этому механизму внутренние части цветка защищаются от
ночной сырости или дождя, и, во-вторых, лепестки с налипшей на них пыльцой,
тесно прилегая к рыльцу, отдают ему пыльцу, и происходит самоопыление.
У чистотела и ремерии (табл. 32) можно наблюдать
другое любопытное приспособление к автогамии. В самом начале цветения - а
продолжается оно от одного до трех дней - тычиночные нити сильно отогнуты по
направлению к краю венчика. По мере старения цветка они выпрямляются и, в конце
концов, приближаются к рыльцу и опыляют его.
Иногда, в связи с плохой погодой, цветки
некоторых растений не раскрываются вообще. Однако это не мешает плодоношению:
самоопыление в таких случаях происходит внутри нераскрывшегося цветка.
Зрелые пыльники гипекоума раскрываются кнаружи,
и в самом начале цветения пыльца из них полностью высыпается на средние доли
внутреннего круга лепестков. Под тяжестью пыльцы эти доли несколько
прогибаются, открывая свободный доступ к нектару и пыльце насекомым-опылителям.
Рыльце с двумя нитевидными ветвями располагается перпендикулярно к этим долям
цветка. Поэтому даже при вечернем закрывании цветка в первый день цветения
автогамия произойти не может, так как в этом случае непосредственно с рыльцем
соприкасаются части цветка, не несущие пыльцы. Самоопыление совершается в самом
конце цветения, на втором или третий день. К этому времени в результате
неоднократного открывания и закрывания цветка часть пыльцы просыпается на
середину наружных лепестков. Ветви рыльца на этой стадии дугообразно изгибаются
книзу, и, когда с наступлением темноты цветок снова закрывается, эта
просыпавшаяся пыльца неизбежно попадает на кончик рыльца и опыляет его.
Анемофилия среди маковых - явление, скорее
всего, вторичное и встречается только у двух родов - маклеи и бокконии. Их
безлепестные цветки собраны в кистевидные соцветия. Пыльники расположены на
очень длинных нитях и раскачиваются при малейшем движении воздуха.
Наиболее распространенный тип плода у маковых -
сухая коробочка округлой или стручковидной формы (рис. 106). Округлая коробочка
маков при высыхании либо растрескивается, либо раскрывается порами в верхней
части. Топкие стручковидные коробочки, раскрывающиеся створками или
разламывающиеся на членики, имеются у чистотела, сангинарии, гиломекона,
гипекоума, эшольции, хохлатки, дицентры. Исключением является дымянка, плод
которой - маленький одно- или двусемянный орех.
Семена большинства маковых мелкие, с обильным
маслянистым эндоспермом и маленьким, слабо дифференцированным зародышем,
октаэдрической или округлой формы, часто с придатками.
Весьма разнообразны способы распространения
семян у маковых и связанные с ними биологические приспособления. Семена,
лишенные придатков, чаще всего разбрасываются разными способами, т. е. являются
баллистами. Так, многочисленные мелкие семена маков высыпаются из зрелой коробочки
и падают довольно далеко от материнского растения. Особенно интенсивно это
происходит в ветреную погоду, когда сухие коробочки мака, как погремушки,
начинают качаться на сильном ветру. У эшольции при созревании коробочка
растрескивается, а семена, снабженные своеобразными пружинками, с силой
отскакивают от плацент, иногда в радиусе нескольких метров вокруг плодоносящего
растения.
Семенные придатки маковых - элайосомы - являются
выростами наружного интегумента. Такой вырост окружает микропиле (дендромекон)
или располагается вдоль семенного шва в виде гребешка (сангинария). Элайосомы
дицентры и чистотела видны под лупой, они напоминают диковинные восковые цветы,
сидящие на семенном шве. Сочная и маслянистая ткань элайосом привлекает
муравьев, которые и являются распространителями семян этого типа.
Кроме баллистов и мирмекохоров, в семействе
маковых имеются и эндозоохоры. Семена многих маков и близких к ним родов
поедаются птицами или другими животными и распространяются с экскрементами.
Некоторые грызуны - полевки, суслики, тушканчики, песчанки - распространяют
семена, запасая впрок коробочки мака и ремерии.
Маковые делятся на 3 подсемейства: маковые
(Papaveroideae), гипекоумовые (Hypecoideae) и дымянковые (Fumarioideae).
Подсемейство маковых состоит из 26 родов и более
450 видов, распространенных главным образом в северном полушарии,
преимущественно в умеренных и субтропических областях, с немногочисленными
видами в тропической Америке. Это многолетние или однолетние травы с окрашенным
или реже бесцветным соком, иногда полукустарники или кустарники (дендромекон,
боккония, маклея) или небольшие деревья (боккония древовидная). Листья
очередные или мутовчатые, как у платистемона (Platystemon). В листьях и стеблях
имеются млечные трубки или млечные мешки. Членики сосудов с простой
перфорацией. Цветки большей частью одиночные, реже в кистевидных соцветиях,
обычно крупные, актиноморфные, двухчленные или реже трехчленные. Околоцветник
состоит из 2-3, обычно опадающих чашелистиков и 4-6-8-12 (10) лепестков. Лепестки
свободные, двурядные, в бутоне черепитчатые и часто скомканные, иногда
отсутствуют (боккония, маклея). Нектарников нет. Тычинки обычно многочисленные,
редко их 6-12, очень редко 4 (птеридофиллум), свободные. Пыльца чаще всего
трехбороздная, реже рассеянно-многобороздная или многопоровая. Гинецей состоит
из двух или более плодолистиков, завязь верхняя или редко полунижняя
(эшольция), одногнездная или ложномногогнездная, редко двугнездная, с
многочисленными семязачатками. Плод - коробочка, стручковидная или
раскрывающаяся порами. Основное число хромосом, равно 5, 6, 7, 8, 9, 10, 11.
Это подсемейство - самое большое по объему. Из
его представителей наиболее широко распространенным и самым известным является
мак. Крупными по объему родами являются аргемона (около 50 видов,
распространенных в Северной и Центральной Америке), ромнея (около 30 видов),
глауциум (Glaucium, табл. 33, около 30 видов). Довольно изолированное положение
в подсемействе занимает монотипный род птеридофиллум (Япония), который часто
вместе с гипекоумом рассматривают в составе подсемейства (или семейства)
гипекоумовых. Иногда этот род выделяют в отдельное подсемейство или семейство
птеридофиллумовых (Pteridophyllaceae). От остальных маковых этот род отличается
числом хромосом, сильно редуцированным андроцеем, перисторассеченными листьями
и отсутствием млечного сока.
Монотипное подсемейство гипекоумовых
представлено одним родом гипекоум, 18 видов которого распространены в
Средиземноморье, Юго-Восточной Европе, Передней и Средней Азии, Сибири, Монголии,
Гималаях и Китае. Как правило, это травы с густой розеткой трижды
перисторассеченных листьев. Млечные трубки и млечные мешки отсутствуют. Цветки
собраны в многоцветковые верхоцветные соцветия, актиноморфные, двухчленные.
Околоцветник состоит из 2 мелких треугольных чашелистиков, которые не полностью
обволакивают венчик до момента его раскрывания, и 4 лепестков, расположенных в
двух кругах. Четыре тычинки супротивны лепесткам, две внутренние часто с двумя
проводящими пучками. Пыльца двубороздная. Гинецей из 2 плодолистиков. Плод
стручковидный, обычно разламывающийся поперек на членики, реже раскрывающийся
двумя створками. Основное число хромосом равно 6, 8.
Род гипекоум иногда включают в подсемейство
маковых, реже - в подсемейство дымянковых. Однако данный род сильно отличается
от представителей обоих этих подсемейств и заслуживает выделения в отдельное
подсемейство. Это подтверждается данными эмбриологии и наличием у гипекоума
двубороздной пыльцы, которая в виде исключения встречается у птеридофиллума.
Подсемейство дымянковых очень близко к
подсемействам маковых и особенно гипекоумовых. Это высокоспециализированное
подсемейство, состоящее из 16 родов и около 400 видов, распространенных главным
образом в северной умеренной зоне. Небольшая часть видов встречается в
Юго-Восточной Африке. Представители подсемейства - травы с прикорневыми,
очередными или редко почти супротивными, обычно сильно рассеченными листьями.
Млечные трубки и млечные мешки отсутствуют, но имеются гомологичные им
секреторные клетки. Цветки в кистевидных соцветиях, обоеполые,
билатерально-симметричные или чаще зигоморфные, с горизонтальной плоскостью
симметрии. Околоцветник состоит из 2 мелких, обычно опадающих чашелистиков, не
образующих замкнутого вместилища (иногда чашелистики совершенно незаметные или
отсутствуют), и 4 лепестков, расположенных в два круга, с нектарниками.
Андроцей из 6 тычинок, сросшихся по три в два боковых пучка. Оболочка пыльцы
трехбороздная (дицентра), рассеянно-многобороздная или многопоровая (дымянка и
дымяночка). Гинецей из 2 плодолистиков, со сплюснутым лопастным рыльцем и с
двумя - многими семязачатками. Плод - стручковидная коробочка, открывающаяся
двумя створками, с сохраняющимися плацентами, без перегородки, реже
односемянный орешек. Основное число хромосом равно 6, 8.
Самый большой род этого подсемейства - хохлатка
Хохлатка, как и ближайшие ее сородичи - дымянка и дымяночка, - являются
типичными представителями ранневесенней флоры умеренной зоны и предпочитают
мезофитные условия местообитания. Приспособлением к ранней вегетации у хохлаток
являются клубневидные утолщения корня с запасом питательных веществ. Некоторые
из дымянок иногда появляются в посевах и на газонах как сорняки.
Представители семейства маковых содержат
многочисленные и разнообразные алкалоиды - производные изохинолина. Названия
этих веществ - папаверин, адлюмин, бикукулин, глауцин, фумарин и т. д.- ясно
указывают источники сырья. Лекарственные свойства маковых были известны
издавна. Первым из маковых еще в доисторические времена был введен в культуру
мак снотворный (Papaver somniferum), вероятно в Западном Средиземноморье.
Оттуда эта культура проникла в Восточную, Южную и Среднюю Азию. Эти
предположения основываются на неоднократных находках коробочек мака в
палеолитических свайных постройках в этих районах. Второй древний центр
культурных форм - Северо-Западный Китай.
Семена мака используют для добывания масла и
приготовления различных кулинарных изделий. Гораздо большее значение имеет
добыча опиума из незрелых маковых коробочек. Он является ценным медицинским
сырьем для изготовления различных наркотиков. Основные современные районы
произрастания и культивирования опийного мака - Китай, Средняя. и Малая Азия, а
также Индия. Делались попытки выращивать опийный мак в Европе, но без; особого
успеха. Опийный мак обычно разводят в горах, на высоте 1500-2900 м, масличный -
на равнинах. Опийный мак сеют после периода осенних дождей в три приема - с
ноября по март. После опадения лепестков и тычинок молодую коробочку надрезают
острым ножом, а затем по мере загустения выступившего сока его собирают, сушат
и прессуют. Все операции по добыче опийного сырья производят вручную, чем и
объясняется большая его дороговизна. Опиум (от греч. opos - сок) был известен
уже в Древней Греции как лекарственное средство. В средние века курение опиума
в качестве одуряющего и галлюциногенного снадобья начало распространяться из
Малой Азии далее на восток. Особенно этот пагубный обычай распространился в
Китае, в связи с чем в 1796 г. китайское правительство запретило курение
опиума, а в 1820 г. и ввоз этого продукта. Это запрещение касалось главным
образом английского ост-индского торгового общества, тотчас же организовавшего
оживленную контрабандную торговлю. Ряд столкновений между Китаем и Англией
привел, наконец, к известной «опийной» войне 1841 г., после победоносного для
Англии окончания которой Китай вынужден был дать официальное разрешение на ввоз
английского опиума.
В настоящее время, несмотря на медицинскую
пропаганду и полицейские меры, наркомания в США, ФРГ, странах Латинской Америки
приобрела столь колоссальные размеры, что может быть приравнена к национальному
бедствию.
Однако со временем опиум не утратил своего
значения и в медицине. В чистом виде в качестве обезболивающих средств
используют морфий и пантопон, различные их производные входят в состав более
пятисот медицинских препаратов.
Лекарственными свойствами, широко используемыми
в народной медицине, обладает также другой представитель семейства - чистотел.
Сильно ядовитый сок этого растения содержит 6-7 алкалоидов: хелидонин,
гомохелидонин, протонин, сангуинарин и др. Его применяют для выведения
бородавок, отсюда и русское его название. Средневековые алхимики пытались в
«золотом» корне чистотела найти средство для изготовления золота из менее
ценных металлов. Согласно Плинию, лекарственным, это растение считалось и в
Древнем Риме. В средние века, особенно в Германии, чистотел применяли против
подагры, золотухи и при лечении переломов костей. В зоотехнике употребляют
свежую траву чистотела, пересыпанную солью, при вздутии живота у овец.
Виды рода дымянка также издавна используют в
медицине как источник лекарственного сырья. Во всех частях растений содержится
до 0,5% алкалоидов, витамины А, Е и С. Эссенцию из цветущих растений применяют
в гомеопатии. Молодые листья употребляют в пищу как салат. В народной медицине
сок используют наружно при сыпях и чесотке.
Ценным лекарственным растением является также
хохлатка. Наиболее богат алкалоидами ее клубень, что приводит к массовому
уничтожению растений. Один из видов, а именно хохлатка Северцова (Corydalis
severtzovii), внесен в «Красную книгу». Это эндемик Западного Тянь-Шаня и
Памиро-Алая (Южный Казахстан, Киргизия, Узбекистан, Северный Таджикистан).
Все маковые очень декоративны. Многие из них
давно введены в садовую культуру. Хорошо известен, например, мак восточный
(Papaver orientale) с крупными огненно-красными цветками и
щетинисто-волосистыми листьями, а также изящный мак альпийский (P. alpinum,
табл. 32) - украшение любого альпинария. Некоторые виды дицентры, адлюмии и
хохлатки также известны как декоративные растения и хорошие медоносы.
Дикорастущие представители семейства маковых
благодаря декоративности усиленно истребляются. Несколько видов из этого
семейства, произрастающие на территории нашей страны, внесены в «Красную
книгу». Например, эндемик предгорий Северного Кавказа мак прицветниковый (P.
bracteatura) с огромными, диаметром до 20-25 см, цветками. Сейчас он
встречается крайне редко, естественное возобновление практически отсутствует.
Характеристика суккулентов, примеры
микроспорогенез суккулент паслен
цветок
Название «суккулент» происходит от латинского
«succus» - сок. Под названием «суккуленты» объединены растения из разных
семейств с сочными листьями и стеблями. Такая похожесть объясняется схожими
условиями существования - эти растения живут в засушливых областях, в пустынях
и даже в придорожной пыли. Они не привыкли к частым дождям. Более того, частый
полив их губит. Многие суккуленты обходятся влагой, которую получают всего пару
раз в год. В этот короткий сезон дождей они запасают воду в листьях и стеблях.
Восковой налет, благодаря которому многие суккуленты кажутся сизыми, защищает
листья от палящих солнечных лучей и испарения влаги.
Суккуленты встречаются в нескольких десятках
семейств растительного царства. Толстянковые и кактусовые полностью состоят из
суккулентных растений. В других семействах (ластовневые, сложноцветные,
агавовые, перечные, асфоделовые, молочайные) суккуленты представлены многими
видами. В некоторых семействах насчитывается по 1-2 рода суккулентных растений.
Суккуленты обитают на всех материках земного
шара, но в подавляющем большинстве они произрастают в пустынях и полупустынях
Южной Африки (Карру, Калахари и прилегающая Капская земля) и засушливых районах
Центральной Америки. В природе они растут на скалах, в песках, влажных
тропических и горных субтропических лесах и т.п.
Суккуленты бывают разного облика и размеров -
деревья, кустарники, травы, среди них есть эпифиты, лианы. Различают стеблевые
и листовые (листо - стеблевые) суккуленты. У стеблевых функцию редуцированных
листьев выполняют стебли. Типичными их представителями являются кактусовые.
Листовые имеют мясистые, сочные листья. У подавляющего числа видов (выходцев из
пустыни) листья собраны в розетку, что способствует меньшему испарению воды. В
комнатных условиях многие виды выращивают ради цветков (виды каланхоэ, хойи и
др.). Однако в первую очередь суккуленты привлекают удивительно красивыми и
причудливыми формами.
Это весьма неприхотливые, легкие в культуре
растения. Важно только обеспечить привычные для них в естественной среде
обитания условия - солнечный свет и свежий воздух летом, прохлада и сухая почва
в период покоя зимой. С весны до осени им нужен регулярный полив, но только
тогда, когда почва станет сухой. Ведь благодаря своим суккулентным свойствам их
толстые мясистые листья и стебли накапливают достаточно влаги, и в наших
квартирах они, так же как у себя на родине, могут обходиться без воды, по
крайней мере, несколько недель.
При низких зимних температурах полив опасен, а
при температуре ниже 10 С это верная гибель для растений. Весной суккуленты
надо подкормить, причем удобрения лучше добавлять микродозами в поливную воду.
Суккуленты солнце-любивые, но требуют частичного притенения, особенно в
полуденное время. Только в хорошо проветриваемых помещениях они переносят и
прямые солнечные лучи.
На зиму одни виды суккулентов можно оставить на
подоконнике, другие лучше поместить в более холодные условия. Идеальные условия
для «колыбельного» периода суккулентов - сухой холод - предоставляются им в
зимнем саду; хорошие условия - при содержании в холодной комнате, с диапазоном
температур от 5 до 10 'С (максимум 15 'С) и освещением не менее 12 часов в
сутки.
Анатомическое строение суккулентов таково, что
большая часть тканей листа и стебля состоит из крупных тонкостенных клеток,
загасающих воду. Одревеснение стеблей столь незначительно, что некоторые
суккуленты получили название «масляное дерево» (Cotyledonpaniculata), «сальное
дерево» (Portulacariaafra), потому что их толстые «стволы» легко режутся ножом
- как масло.
Воду у суккулентов запасают разные органы.
Например, у алоэ, агавы, хавортии, гастерии, очитка - листья. Такие растения
называют листовыми суккулентами. У стеблевых суккулентов сочный стебель. Ярким
примером последних служат кактусы и молочаи.
Иногда резервуарами для воды служат листья и
стебли одновременно, как у цилиндрических опунций, пеперомий, крестовников,
некоторых бегоний. К суккулентам можно отнести и такие растения, у которых
влага запасается в сильно разросшейся нижней части ствола. Это так называемые
«бутылочные» деревья из рода адениум (семейство кутровых), адения (семейство
пассифлоровых), из семейства Виноградовых - цифостема и многие другие. У
отдельных видов кислиц (род Oxalis) тонкая и мягкая пластинка листа быстро
опадает, а оставшийся черешок становится сочным и долгое время служит
вместилищем влаги. В природе встречаются суккулентные лобелии из семейства
колокольчиковых, родственные тыкве мадагаскарские ксеросициусы.
Из приведенных примеров видно, что к суккулентам
относятся растения из разных, не родственных друг другу групп. Более 33
семейств, а по данным В. Pay, не менее 50, имеют «сочных» представителей.
Что же роднит суккуленты, такие разные, далекие
в систематическом отношении? Прежде всего среда обитания, т.е. жизнь в условиях
экологически очень сходных - засушливых областях земного шара; затем
приспособленность растений к условаиям крайней сухости. Таким образом, можно
сделать заключение, что Суккулент - понятие эколого-морфологическое.
Интерес людей к суккулентным растениям проявился
очень давно Уже в середине XVI в испанцы и португальцы завезли в Европу первые
суккулентные растения, такие, как опунция, цереус, алоэ, агава.
Выращивали их в монастырских садах или в садах
крупных вельмож. Внимание к суккулентным растениям проявляли многие ученые.
Так, например, швейцарские ботаники Иоган Баухин (Bauhin J) и Иоган Херлер
(Cherler J.) в работе «Historiae Plantarum Generalis» (1619) приводили список
растений с мясистыми стеблями и листьями, способными долгое время оставаться
живыми после того, как их сорвешь.
Профессор ботаники в Кембридже Р. Бредли
(Bradley R.) в своей работе «История суккулентных растений» (1717-1727) писал о
невозможности гербаризировать Суккуленты. К. Линней в работе «Философия
ботаники» (1751) выделял группу суккулентных растений из 25 родов.
В XIX и XX вв огромное число ученых занималось
изучением этих растений. Только одно перечисление их имен заняло бы целую
страницу.
В начале XIX в. богатыми коллекциями суккулентов
обладали А. Хаворт (Англия), принц Сальм-Дик (Германия). В России в 30-х годах
XVIII в. в ботаническом саду Петра Великого в Санкт-Петербурге наряду с
тропическими растениями выращивали кактусы, алоэ, агавы, молочаи. Э.Л. Регель -
крупный ботаник того времени, возглавлявший Ботанический сад почти 25 лет,
описал несколько видов Суккулентов - толстянку Купера толстяку Шмидта
(Crassulacooperi, С schmidtii), агаву Максимовича и агаву опушенную
(Agavemaximowicziana, A. pubescens), а также некоторые кактусы.
В середине XIX в. коллекция растении из
засушливых областей в Ботаническом саду Санкт-Петербурга состояла из 550
наименований, в том числе 40 видов алоэ 23 видов агав, 15 видов хойи, 12 видов
гастерий. К концу XIX в. (1897 г.) она насчитывала 1700 таксонов (видов,
разновидностей и культиваров). Коллекцию удалось сохранить и в годы революции,
гражданской войны и разрухи в стране, однако объем ее несколько уменьшился. По
архивным данным удалось установить, что на 1941 г. она состояла из 1200
таксонов. Огромный ущерб саду был нанесен во время второй мировой войны.
Усилиями садоводов, работавших в блокадном Ленинграде, удалось сохранить
большое число крупных экземпляров суккулентов - всего около 300 видов.
После Великой Отечественной войны начинается
активное восстановление коллекции. Интерес любителей к суккулентным растениям в
СССР особенно проявился в 50-60-е годы, когда в разных городах страны были
созданы общества любителей кактусов.
Однако коллекционированием Суккулентов из других
семейств в настоящее время занимаются сравнительно немногие.
Современная архитектура зданий с обилием света
позволяет расширить ассортимент комнатных растении, и Суккуленты представляют
большой интерес в этом отношении. Небольшие размеры, компактность,
сравнительная легкость размножения, несложность культуры делают их
перспективными для озеленения комнат, создания микрокомпозиций, использования в
качестве компонентов в так называемых каменистых горках при озеленении
интерьеров. Одним из положительных свойств суккулентных растений является их
устойчивость к сухости воздуха. Сухой воздух в современных квартирах - главный
враг большинства комнатных растений тропического и субтропического
происхождения. Суккуленты же могут переносить длительную пересушку земляного
кома, даже летом.
Список использованной литературы
1.
Андреева Ивелина Ивановна. Ботаника: учебник для студентов вузов по агрон.
специальностям / И.И. Андреева, Л.С. Родман.- 3-е изд., перераб. и доп. - М.:
КолосС, 2007. - 528 с.
.
Тихомиров Ф.К. Ботаника: Учебник для с.-х. вузов. / Ф.К. Тихомиров.- 3-е изд.,
перераб. - М.: Высш. шк., 1974.- 397 с.
.
Жуковский П.М. Ботаника: Учебник по агрономическим специальностям / П.М.
Жуковский.- 5-е изд., перераб. и доп. - М.: Колос, 1982.- 623 с.
.
Лотова, Л.И. Ботаника: морфология и анатомия высших растений / Л.И. Лотова. -
Изд.4, доп. - М.: Книжный дом "ЛИБРОКОМ", 2010.- 512 с.
.
Суворов В.В. Ботаника с основами геоботаники: учебник для подготовки
бакалавров, обучающихся по направлениям 110400 "Агрономия" и 110500
"Садоводство" / В.В. Суворов, И.Н. Воронова. - 3-е изд., перераб. и
доп. - М.: АРИС, 2012.- 520 с.
.
Методические указания по изучению курса ботаники и задания для контрольных
работ студентам заочной сокращенной формы обучения агрономических
специальностей / Воронеж. гос. аграр. ун-т; [подгот.: А.В. Никулин, Е.М.
Олейникова]. - Воронеж: ВГАУ, 2010. - 20 с.