Зрительное восприятие
Введение
Зрение эволюционно приспособлено к
восприятию электромагнитных излучений в определенной, весьма узкой части их
диапазона (видимый свет). Зрение - многозвеньевой процесс, начинающийся с
проекции изображения на сетчатку уникального периферического оптического
прибора - глаза. Затем происходят возбуждение фоторецепторов, передача и
преобразование зрительной информации в нейронных слоях зрительной системы, а
заканчивается зрительное восприятие принятием высшими корковыми отделами этой
системы решения о зрительном образе.
Результаты последних лет указывают
на существование преаттентивного механизма пространственного группирования
зрительной информации, образованного фильтрами второго порядка.
Исследование механизмов второго
порядка необходимо, чтобы понять, как человек воспринимает окружающий его мир в
целостных образах и конечно, для понимания того, как функционирует мозг
человека.
Этапы зрительного
восприятия
зрительный
кодирование восприятие группирование
Процесс зрительного восприятия может
быть условно разделен на два этапа. (Neisser, 1973). Первый осуществляется в пределах проекционной системы и
реализуется в виде параллельного локального описания входной информации с
помощью линейных фильтров. Данная операция осуществляется клетками,
формирующими пути от рецепторов сетчатки до простых нейронов стриарной коры.
Далее выделенная локальная информация должна каким-то образом группироваться в
блоки, которые затем используются при опознании. На ранней, преаттентивной
стадии анализа реализуется автономно действующий механизм объединения локальной
информации. Параллельная обработка начинается с разбиения сцены на множество
локальных фрагментов, каждый из которых анализируется в значительной степени
независимо. В результате выделяются так называемые примитивы - элементарные
признаки изображений. Если говорить о пространственных признаках, то речь может
идти о локализации градиентов яркости, их ориентации и пространственной
частоте. Затем из этих примитивов зрительная система должна «собрать» отдельные
объекты.
Следующий этап - это перевод
множественного локального описания входной информации в единое глобальное
представление о зрительной сцене.
Этап №1. Первичная
обработка зрительной информации и обработка на уровне наружного коленчатого
тела
В основу гипотезы Хьюбела и Визела о
функциональной организации зрительной коры положены два принципа: детекторный
принцип обработки сигнала и иерархический принцип построения системы
детекторов. Детекторный принцип позволил обнаружить, что нейроны зрительной
коры выделяют определённые признаки изображения. Иерархический принцип говорит
о том, что нейроны с рецептивными полями (простые-сложные-сверхсложные)
представляют собой последовательные, все более высокие стадии обработки
зрительной информации, на которых выявляются все более сложные признаки
изображения.
Первый уровень обработки зрительной
информации - это описание изображения множеством рецептивных полей зрительной
подкорки. Нейроны сетчатки и наружного коленчатого тела измеряют
интегрированную световую энергию в пределах центральной зоны суммации
рецептивных полей. Существование тормозной периферии рецептивных полей приводит
к тому, что нейроны этого уровня выполняют ряд операций по предварительной
переработке зрительного сигнала: выделение сигнала из шума, подчеркивание
контуров и высоких пространственных частот, пространственную и временную
декорреляцию изображения. Ретинотопическая и структурная упорядоченность в
организации наружного коленчатого тела, а также выявленные у рецепторных
нейронов свойства позволяют предположить существование на этом уровне
зрительной системы квазипоточечного описания зрительного мира, т.е. отображение
в пространственно-временном рельефе активности нейронов наружного коленчатого
тела всех свойств изображения, существующего на сетчатке. Пространственные
характеристики такого рельефа активности несут в себе сведения о
пространственных свойствах изображения и освещенности изображения; т.е.
предполагается, что подкорковые уровни служат для эффективного кодирования,
обработки и передачи в кору сведений об отдельных параметрах ретинального
изображения (интенсивность, размер, элементы формы и т.д.).
Таким образом, на уровне первичной
обработки информации в сетчатке, и на уровне обработки информации в наружном
коленчатом теле происходит подготовка информации (своего рода выделение сигнала
из шума) для её дальнейшей фильтрации в стриарной и экстрастриарной коре.
Этап №2. Обработка
зрительной информации на уровне стриарной коры
Рецептивные поля стриарной коры
(поле 17 или V1) - двумерные решетчатые фильтры пространственных частот. Поля
измеряют пространственно-частотную и ориентационную компоненты фрагментов
изображения. Нелинейные нейроны служат для сегментации зрительного поля на
отдельные фрагменты. Для каждого отдельного фрагмента эта операция может быть
описана как выделение фигуры из фона. Выполняется она сложными нейронами,
измеряющими кусочные мощностные спектры и выделяющими участки, занятые
однородной текстурой; дирекциональными нейронами, выделяющими площадки,
составленные элементами, движущимися в одном направлении; бинокулярными
нейронами, выделяющими участки, расположенные на разной глубине, а также
цветовыми нейронами. Эти участки могут быть объединены в более крупную фигуру
обратными влияниями из вышележащих отделов. Выделение участка анализируется
модулем, составленным линейными нейронами. Каждый модуль состоит из рецептивных
полей одной величины и разной ориентации. Для каждого выделенного изображения
подбирается модуль соответствующего размера и положения из сети модулей,
перекрывающей центральный участок поля зрения. Показано, что такая система
модулей может быть отождествлена с оптимальной системой передачи информации по
Габору, поскольку все модули могут быть соотнесены с логонами, имеющими
одинаковую площадь. Модель модулей объединяет концепции модулей Маунткасля,
ориентационных свойств Хьюбела-Визела и пространственные частоты каналов
Кэмпбелла.
Таким образом, легко заметить, что в
стриарной коре происходит выделение информационных фрагментов по признаку
фигура-фон (тело-поле, объект-взаимодействие (окружение)). Акцент делается либо
на выделение фигуры из фона, либо на выделение фона у фигуры.
Этап №3. Обработка
зрительной информации на уровне экстрастриарной коры
Нейронные процессы, происходящие в
первичной зрительной (стриарной) коре, - это в известной мере базовые,
принципиальные процессы, т.е. первичная зрительная кора, прежде всего,
предназначена для первичной регистрации таких локальных отличительных черт
раздражителя, как его края и ориентация (хотя не исключено, что за счет обратной
связи с другими зрительными слоями она способна и к кодированию иных, более
сложных визуальных параметров; Leeetal., 1998). Из слоя VI нейронные сигналы
поступают в другие слои коры, отличающиеся от слоя VI как анатомически, так и
функционально. Все вместе эти слои образуют экстрастриарную кору (или вторичную
кору, которую также иногда называют зрительной ассоциативной корой или полем
Бродмана 18). Каждый из кортикальных слоев, участвующих в образовании
экстрастриарной коры, обрабатывает такую специфическую информацию об
особенностях нейронных сигналов, посылаемых слоем VI, как информация о форме,
цвете и движении.
Различные слои экстрастриарной коры
имеют свои номера, отражающие их расположение относительно первичной зрительной
коры (слоя VI). Так, ближайший к слою VI слой называется слоем V2, затем
следуют слои V3, V4 и V5 (последний также называется слоем МТ, что отражает его
расположение на стыке теменной и височной долей коры головного мозга. В
соответствии с другой номенклатурой этот участок также называется полем
Бродмана 19).
Что касается функций, то слои V2 и
V3 обрабатывают информацию о форме и положении в пространстве, слой V4, судя по
всему, «специализируется» на восприятии информации о цвете, а слой V5 - на
анализе визуальной информации о движении. Нейронные сигналы, берущие свое
начало в слое VI, поступают не только в перечисленные выше кортикальные слои,
но в слой височной доли коры - слой IT (по начальным буквам латинских слов
inferior и temporalis «нижний» и «височный») - в нижнюю височную кору,
расположенную на вентральной части височной доли и называемую также полями
Бродмана 20 и 21), и в участок теменной доли коры (полеБродмана17). Полагают,
что в слое IT интегрируются результаты обработки информации о форме и цвете и
что он также принимает участие в относительно сложных аспектах
пространственного восприятия. Нейрофизиологические свидетельства функциональной
специализации экстрастриарных кортикальных слоев получены на основании изучения
индивидуумов, перенесших травмы тех или иных участков коры головного мозга.
Например, человек с поврежденным слоем V4 экстрастриарной коры может полностью
утратить цветовое зрение (Pearlmanetal., 1979; Sacks, 1995), а человек с
поврежденным слоем МТ (слоем V5) экстрастриарной коры может потерять возможность
воспринимать непрерывное движение (Zihletal., 1983).
Результаты параллельной
и последовательной обработки информации
Данные исследований последних лет
говорят о том, что параллельная обработка информации не заканчивается на уровне
выделения примитивов. Происходит дальнейшее преаттентивное объединение
пространственно-распределенной информации в так называемые когнитивные блоки.
На уровне последовательной обработки
включается внимание, с помощью которого происходит выявление различных свойств
предметов, выделенных на преаттентивном уровне, вследствие чего мы можем
сознательно оперировать полученными данными.
Кодирование признаков в
зрительной системе
На предварительных этапах
перцептивная система использует информацию с сетчатки глаза, в частности
вариации интенсивности, и описывает объект на языке элементарных составляющих,
таких как линии, края и углы. На основании этих элементарных составляющих
система составляет описание самого объекта. На завершающих этапах система
сравнивает это описание с описаниями форм разного рода объектов, хранящихся в
зрительной памяти, и выбирает наилучшее ему соответствие. Например, опознать
определенный объект как букву В-значит сказать, что его форма больше
соответствует форме буквы В, чем форме других букв.
Детекторы признаков в коре мозга.
Многое из того, что на сегодня известно об элементарных признаках объекта
восприятия, было получено в биологических экспериментах над другими видами
(кошками, обезьянами) с применением регистрации активности отдельных клеток
зрительной коры. В этих исследованиях изучалась чувствительность специфических
нейронов коры во время предъявления различных стимулов на те участки сетчатки
глаза, которые связаны с этими нейронами; такой участок сетчатки называют
рецептивным полем кортикального нейрона. Первые исследования с одноклеточной
регистрацией были проведены Хьюбелем и Визелем (Hubel&Wiesel, 1968),
которые получили за них Нобелевскую премию в 1981 году. Хьюбел и Визел выделили
в зрительной коре три типа клеток, различающихся по признакам, на которые они
реагируют. Простые клетки реагируют, когда глазу предъявляют стимул в виде
линии (тонкой полоски или прямой грани между темным и светлым участками),
имеющей определенную ориентацию и положение в рецептивном поле. На рис. 1
показано, как реагирует простая клетка на вертикальную полоску и на полоски,
наклоненные относительно вертикали. По мере отклонения ориентации от
оптимальной реакция снижается. Другие простые клетки настроены на другие
ориентации и положения. Сложные клетки тоже реагируют на полоску или край
определенной ориентации, но для них не обязательно, чтобы стимул находился в
определенном месте рецептивного поля. Они реагируют на стимул, находящийся в
любом месте их рецептивного поля, и реагируют непрерывно, пока стимул
перемещается по их рецептивному полю. Сверхсложные клетки реагируют на стимул
не только определенной ориентации, но и определенной длины. Если длина стимула
выходит за пределы оптимальной, реакция ослабляется и может совсем
прекратиться. Со времени публикации Хьюбелем и Визелем своих первых данных
ученые обнаружили клетки, реагирующие на другие формы стимулов, помимо полосок
и краев; например, они обнаружили сверхсложные клетки, реагирующие на углы и
кривые линии определенной длины (Shapley&Lennie, 1985; DeValois&DeValois,
1980).
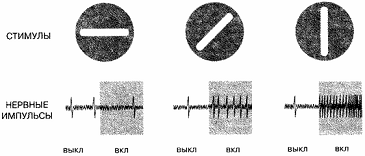
Рис. 1 Реакция простой клетки. На
рисунке показана реакция простой клетки коры на полоску света. Сверху показан
стимул, снизу - реакция; каждый большой всплеск на графиках внизу соответствует
одному нервному импульсу. При отсутствии стимула регистрируется только
случайный импульс. Когда стимуляция включена, клетка может реагировать или не
реагировать в зависимости от положения и ориентации полоски света. У данной
клетки предъявление горизонтальной полоски не меняет реакцию, полоска с
наклоном в 45° вызывает небольшое изменение реакции, а вертикальная полоска
вызывает очень большое изменение.
Все вышеописанные типы клеток
называются детекторами признаков. Поскольку края, полоски, углы и изломы, на
которые реагируют эти детекторы, могут использоваться для аппроксимации
множества форм, есть основание рассматривать детекторы черт как кирпичики, из
которых строится воспринимаемая форма.
Таким образом, встриарной коре
человека кодируются следующие признаки изображения: контраст, пространственная
частота и ориентация.
Детектор полосы получается путем
сложения нескольких рецептивных полей НКТ. Происходит объединение пятен одного
размера и типа.
Рецептивные полястриарной коры
получаются в результате объединения рецептивных полей НКТ одной ПЧ и одного
типа - с on- или off - центром.
Пространственная частота кодируется
шириной, а ориентация - углом наклона. Чувствительность стриарных нейронов к
вертикальным и горизонтальным стимулам несколько выше, чем к наклонным.
Контрастной чувствительностью
определяется способность улавливать минимальные различия в освещенности двух
соседних областей, а также дифференцировать их по яркости.
Пространственная контрастная
чувствительность (ПКЧ) зрительного анализатора является функцией, которая
определяет минимальный контраст, необходимый для обнаружения изображений
различных размеров. Она отражает зависимость порогового контраста от
пространственной частоты стимула.
ПКЧ определяется как величина,
обратная минимальному контрасту решетки, при котором последняя различима при
данной пространственной частоте.
В основе яркостного контраста лежит
взаимное латеральное торможение зрительных нейронов.
Кодирование контраста в зрительной
системе начинается с сетчатки. На этом уровне из зрительной сцены выделяются
контрастные пятна. При этом размер рецептивных полей увеличивается от центра к
периферии. Представлено 2 типа рецептивных полей ганглиозных клеток сетчатки: с
on-
и с off-центром. Соответственно они кодируют светлые пятна на темном фоне
и наоборот. Контрастные границы пятен усиливаются на уровне НКТ за счет более
сильного латерального торможения.
В первичной зрительной коре
информация объединяется, кодируется большее количество характеристик за счет
строения детекторов, их взаимного расположения и взаимодействия.
Взаимосвязь признаков. Форма
описывается не только своими признаками: нужно определить также их взаимосвязь.
Важность связей между признаками иллюстрирует рис. 2. Признаки печатной буквы Т
включают вертикальную и горизонтальную линии, но если эти линии не соединены
правильно, в результате получится не Т. В описании Т следует учесть, что
горизонтальная линия своим центром касается верха вертикальной. Именно такую
связь признаков имели в виду гештальт-психологи, когда предупреждали
предшествующих психологов, что «целое отличается от суммы его частей».
Одно из таких отличий целого от его
частей проявляется в том, что целое создает новые перцептуальные
характеристики, которые невозможно объяснить за счет простого анализа отдельных
частей. На рис. 2 показаны четыре такие возникающие характеристики. Все они
возникают за счет специфических пространственных взаимоотношений между более
элементарными характеристиками. Тем не менее такие возникающие характеристики
часто ведут себя точно так же, как более простые характеристики, при выполнении
таких задач, как обнаружение цели и визуальный поиск (Enns&Resnick, 1990;
Enns&Prinzmetal, 1984; He&Nakayama, 1992). Эти факты свидетельствуют о
том, что в зрительной системе осуществляются различные типы сложного анализа
формы, прежде чем результаты этих анализов становятся доступны сознанию.
Отношения между признаками
При сочетании двумерных признаков,
таких как линии, углы и геометрические формы, результирующий паттерн в значительной
степени зависит от пространственных отношений между компонентами-признаками.
Помимо этого создаются (формируются) новые признаки. Эти возникающие признаки
обладают перцептуальной реальностью, несмотря на то, что они включают сложные
пространственные отношения.
Долгое время считалось, что нейроны
первичной проекционной области зрительной коры являются детекторами простых
признаков изображения (ориентаций линий или контрастных границ), а кодирование
признаков второго порядка в зрительном образе происходит в экстрастриарных
областях коры. Было обнаружено, что для половины нейронов первичной зрительной
коры кошки оптимальными являются крестообразные, угловые или звездчатые фигуры,
т.е. признаки изображения второго порядка. Причем часть клеток чувствительны,
только к одной из этих фигур, а другие отвечают одинаково на любую из них.
Ответы нейронов на подобные стимулы в два-три раза превышают ответы на
одиночную световую полоску. Большинство клеток обладают высокой
избирательностью к ориентации и форме (углу между полосками) этих фигур. Около
30% клеток инвариантны к одному из этих признаков изображения, а 10% - к обоим,
т.е. такие нейроны чувствительны именно к пересечениям линий - одному из
ключевых признаков зрительного образа.
Настройка большинства нейронов на
ориентацию и форму крестообразной фигуры динамична, т.е. клетка за время
развития своего импульсного ответа способна сканировать определенный диапазон
этих характеристик изображения. Это означает, что при выделении признаков
второго порядка зрительная система использует не только пространственное
позиционное кодирование, но и временное (паттерновое) кодирование признаков
зрительного образа. Оно является эффективным и гибким механизмом, дополняющим
более стабильное и жесткое позиционное кодирование.
Чувствительность к сложным фигурам у
трех четвертей нейронов первичной зрительной коры обусловлена механизмом
растормаживания, предполагающим наличие в рецептивном поле (РП) возбудительной,
торцевой тормозной и боковой растормаживающей зон. Когда рецептивное поле
нейрона стимулируется оптимально ориентированной фигурой с оптимальным углом
между полосками, то одна из сторон фигуры активирует возбудительную зону РП
клетки, а вторая - растормаживающую зону, которая в свою очередь блокирует
торцевое торможение и тем самым усиливает ответ по сравнению с ответом на
одиночную полоску (активация только возбудительной зоны). Этот механизм
обеспечивает чувствительность как к «полным», так и «неполным» фигурам. У
четверти клеток подобная чувствительность может определяться за счет того, что
они получают входы от двух нейронов с различными ориентациями, т.е.
конвергентным механизмом, или сочетанием двух механизмов. Это было подтверждено
модельными исследованиями К.А. Салтыкова.
Каналы описания
признаков. Избирательность каналов
Исследования по измерению
частотно-контрастной характеристики зрительной системы (в общем случае
модулирующей передаточной функции) начались в 60-х годах 20 века (Campbell, Green 1965). Зрительная
система рассматривалась тогда, как одноканальная и квазилинейная. Метод,
использовавшийся в исследованиях, - изменение глубины модуляции в пороговых
условиях работы зрительной системы. В результате исследования была получена
кривая частотно-контрастной чувствительности зрительной системы человека.
В начале 70-х годов были получены
экспериментальные данные, которые противоречили представлению о зрительной
системе как об одноканальной линейной системе. Было высказано предположение,
что работу зрительной системы можно рассматривать как функционирование совокупности
нескольких независимых пространственно-частотных каналов.
Кемпбелл, Робсон, Блейморк и
Куликовский выдвинули гипотезу о том, что зрительная система перерабатывает
информацию множеством независимых каналов, настроенных на различные
пространственные частоты (Campbell, Kulikowski, 1966; Campbell, Robson, 1968; Campbelletal., 1968, 1969; Blakemore, Campbell, 1969; Campbell, Maffei, 1970; Campbell, 1974).Зрительный анализатор надо рассматривать не как единый
канал обработки пространственной информации, а как систему параллельно
функционирующих каналов, каждый из которых обрабатывает определенную полосу
частот.
Кемпбелл и Робсон экспериментально
показали возможность Фурье-преобразования изображений в зрительной системе, и
показали наличие множества каналов, но пока эти каналы считались независимыми и
появился вопрос об их количестве.
В результатах исследований Блейморка
и Кемпбелла (Blakemore, Campbell, 1969) с помощью метода адаптации были выявлены
пространственно-частотные каналы, отвечающие на любую частоту. Для объяснения
обнаружения одиночных полос, имеющих синусоидальную функцию освещенности и
различную ширину, Вилсону и Бергену (Wilson, Bergen, 1979) понадобилось всего 4 пространственных элемента,
настроенных на различные частоты. Мостафави и Сакрисон, использовавшие в
качестве стимулов двумерные текстуры также получили 4 канала.
Вилсон с соавторами показал и
доказал экспериментально существование ограниченного и малого количества
каналов в 1983 году. В качестве стимула использовались вертикальные решетки с
яркостными функциями схожими с элементами Габора, настроенные на различные
частоты. С помощью метода маскировки было показано существование 6 ПЧ каналов.
Ширина пропускания каналов оказалась неодинаковой, и наиболее чувствительный
канал был настроен на частоту 4.5 цикл / град. (Wilson, 1978).
В более поздней работе Вилсон и Гелб
(Wilson,
Gelb,
1984) восстановили из пространственно-частотных характеристик весовые функции
фильтров и уточнили их частотную настройку. Два самых низкочастотных канала
представляли собой разность 2 гауссиан, остальные - разность 3 гауссиан.
Кривая контрастной чувствительности,
построенная Кемпбеллом, полностью была заменена дискретными каналами,
найденными и математически описанными Вилсоном и Гелбом.
В итоге были представлены все
соотношения относительных фазовых сдвигов между тестом и маскировкой. В этих
условиях было обнаружено 6 каналов, настроенных на различные частоты.
Можно сделать вывод, что методики и
полученные результаты исследований, верны и могут использоваться в нашем
исследовании.
Проблема
пространственного группирования и зрительные механизмы второго порядка
В литературе проблема
пространственного объединения локальной информации получила название «проблема
связывания». Предполагалось, что объединение локальных признаков происходит
последовательно с помощью избирательного внимания (Treisman, Gelade, 1980;
Wolfeetal., 1990). Однако появлялись все новые факты, свидетельствующие о том,
что процесс группирования некоторых признаков также может осуществляться параллельно
(Humphreysetal., 1989; Carrascoetal., 1998 и др.). Было высказано предположение, что
автоматизированная зрительная обработка включает не одну, а две стадии (Chubb,
Landy, 1991; Wilson, 1993; Graham, 1994). Сначала механизмы первого порядка
выделяют локальные признаки, а затем механизмы второго порядка объединяют эти
признаки. В экспериментах было показано, что группирование локальной информации
действительно осуществляется параллельно (Бабенко и др., 2006). Таким образом,
были получены доказательства существования преаттентивных механизмов второго
порядка. Параллельная обработка не завершается выделением локальных признаков,
а включает механизм их пространственного группирования. Автоматическое
формирование некоторых когерентных форм вполне реально. Об этом заявляли еще
Гештальт - психологи. Именно наличие такого преаттентивного механизма
группирования может объяснить эффекты латеральных взаимодействий, способность
человека разделять текстуры и обнаруживать градиенты, оценивать общий размер и
ориентацию объектов, воспринимать иллюзорные контуры, корректно оценивать
движение и т.д. В доказательство существования такого механизма была предложена
двустадийная модель, в которой фильтры первого порядка избирательны к
ориентации, пространственной частоте и изменению яркости (простые нейроны
стриарной коры), а фильтры второго порядка (постстриарные сложные нейроны)
избирательны к пространственным модуляциям простых признаков. Это свойство
приобретается ими благодаря объединению выходов фильтров первого порядка. Сегодня
факт существования преаттентивного пространственного группирования можно
считать доказанным.
Свойства механизмов
второго порядка
Механизмы второго порядка
осуществляют объединение пространственно-распределенной информации в
когнитивные блоки.
Пространственное объединение
происходит по определенным жестким правилам:
Чем выше пространственная частота
входного сигнала, тем меньше пространственный интервал, на котором происходит
объединение информации.
Объединяются примитивы одной
ориентации и пространственной частоты по оппонетному принципу.
Механизмы второго порядка
чувствительны к модуляциям контраста, ориентации и пространственной частоты.
Одно субполе механизма второго
порядка, чувствительного к модуляции контраста объединяет 8 фильтров первого
порядка.
Одно субполе механизма второго
порядка, чувствительного к модуляции пространственной частоты объединяет 16
фильтров первого порядка.
Одно субполе механизма второго порядка,
чувствительного к модуляции ориентации объединяет 12 фильтров первого порядка.
Благодаря механизмам второго
порядка, оппонентным по ориентации, мы способны разделять многие текстуры.
Механизмы, оппонентные по пространственной частоте, позволяют получать
информацию о глубине. Оппонентность по контрасту дает возможность
сегментировать поле зрения и выделять объекты из фона. При этом универсальность
группирования на разных пространственных шкалах делает этот механизм
инвариантным к размеру изображения: приближение к объекту или удаление от него
не приводит к искажениям в восприятии.
Вывод на основе
физиологических данных по обработке зрительной информации
При прохождении информации от
сетчатки до проекционных зон больших полушарий последовательно срабатывают
фильтрационные механизмы. На этапе первичной обработки информации и прохождения
через наружное коленчатое тело информация готовится для первой стадии
фильтрации в стриарной коре. На втором этапе (стриарная кора) непосредственно
запускается механизм фильтрации по признаку фигура-фон (тело-поле). На третьем
этапе (этапе прохождения информации через экстрастриарную кору) работает
фильтрационный механизм по признаку статика - динамика. Далее, на этапе
попадания информации в нижневисочную и заднетеменную кору происходит обращение
к эталону признака внешнее - внутреннее. После этапа фокусировки слоя сознания,
по всей видимости, результирующая информация после прохождения всех
распознающих фильтрационных механизмов собирается в единый образ.
Список литературы
1. Campbell, F.W. &Green, D.G. (1965).Optical and retinal factors in visual resolution. Journal of
Physiology, 181, 576-593.
2. Campbell, F.W. &Gubisch R.W. (1966) Optical quality
of the human eye. J. Physiol., 186, pp. 558-578
. Campbell F.W., Robson J.G. Application of Fourier
Analyses to the Visibility of Gratings // J. Physiol. 1968. V. 197. P. 551-566.
. Campbell F.W. The transmission of spatial information
through the visual system // Neurosciences. Boston: MITpress, 1974. P. 95-103.
. Chubb, Landy, 1991; Wilson, 1993; Graham, 1994.
. Enns&Resnick, 1990; Enns&Prinzmetal, 1984; He
& Nakayama, 1992.
. Humphreys et al., 1989; Carrascoetal., 1998.
. Hubel & Wiesel, 1968.
. Lee et al., 1998.
. Pearlman et al., 1979; Sacks, 1995.
. Shapley &Lennie, 1985; DeValois&DeValois, 1980.
. Treisman, Gelade, 1980; Wolfe et al., 1990.
. Wilson, Bergen, 1979.
. Zihl et al., 1983.
16. Бондаренко В.М., Данилова М.В., Красильников Н.Н., Леушина
Л.И., Невская А.А., Шелепин Ю.Е. «Пространственное зрение». СПб.: Наука, 1999.
17. http://www.glazamed.ru/oftalmologiya
. Бабенко В.В., Кураев Г.А., Кульба С.Н. Механизмы
повышения контрастной чувствительности при увеличении протяженности
синусоидальной решетки // Сенсорные системы. - 2001. - Т.15, №1. - С. 3-10.
. Бабенко В.В., Кураев Г.А., Кульба С.Н. Проблема
зрительной сегментации и пространственно-частотная фильтрация // Российский
физиологический журнал им. И.М. Сеченова. - 2003. - Т.89, №10. - С. 1300-1309.
. Бабенко В.В. «Сегментация в зрении». Ростов-на-Дону: ООО
«ЦВВР», 2004
. Бабенко В.В., Божинская М.А. Проблема объединения
локальной зрительной информации в когнитивные блоки // Матер. IV Всероссийского съезда
Российского психологического общества. Ростов-на-Дону, 2007. - Т. 1. - С. 73.
. Хьюбел Д. «Глаз, мозг, зрение», М.: Мир, 1990.
. Шамшинова А.М. и Волков В.В. «Функциональные методы
исследования в офтальмологии», 1999.