Модель гетеротрофной сукцессии в культуре сенного настоя
ФЕДЕРАЛЬНОЕ
АГЕНТСТВО ПО РЫБОЛОВСТВУ
Федеральное
государственное БЮДЖЕТНОЕ образовательное учреждение высшего профессионального
образования
«Мурманский
государственный технический университет»
Биологический факультет
Кафедра «Биоэкология»
ДИПЛОМНАЯ РАБОТА
Модель гетеротрофной сукцессии в
культуре сенного настоя
Выполнила:
студентка 5 курса БФ группы Бэ-571,
А.Ю. Колгатникова
Научный руководитель:
Доцент кафедры «Биоэкология»,
к.б.н. Н.А. Пахомова
Мурманск 2012
Содержание
Введение
. Обзор литературы
.1 Общие понятия о сукцессиях
.2 Гетеротрофные сукцессии
.3 Биотестирование и биоиндикация
.4 Задачи и приёмы биотестирования
.5 Методы биотестирования
.5.1 Биохимический подход
.5.2 Морфологический подход
.5.3 Физиологический подход
.6 Биотестирование с использованием инфузорий
. Материалы и методы
.1 Приготовление сенного настоя
.2 Подготовка опыта на нефтепродукты
. Результаты и обсуждение
.1 Получение культуры сенного настоя
.2 Краткая характеристика классов встречающихся организмов
.2.1 Ресничные инфузории (Ciliata)
.2.2 Саркодовые (Sarcodina)
.2.3 Гастротрихи (Gastrotricha)
.2.4 Коловратки (Rotatoria)
.2.5 Турбеллярии (Turbellaria)
.2.6 Нематоды (Nematoda)
.3 Применение инфузорий в биотестировании
.3.1 Нефтепродукты
.3.2 Эпизодические наблюдения
Заключение
Список литературы
Введение
Сукцессия (от лат. successio
- преемственность, наследование) - это процесс саморазвития сообщества.
Экологическую сукцессию можно определить по следующим параметрам: развитие
видовой структуры во времени, изменение условий среды обитания под влиянием
предшествующих организмов, смена видов в экосистеме, которая контролируется
сообществом, и происходящие в системе обменные процессы. Кульминацией развития
является формирование стабильного сообщества, длительность существования
которого определяется поступлением (или отсутствием) энергии из внешней среды.
Возможность выращивания одноклеточных организмов и других видов
простейших гетеротрофов доказана давно, однако широкое производство их начали
всего 16 лет назад. Достаточно исследованы технологические аспекты
культивирования автотрофных микроводорослей и большое внимание уделяется
изысканию путей получения некоторых гетеротрофных беспозвоночных организмов.
Большая роль здесь принадлежит свободноживущим инфузориям. Особое внимание
уделяется подбору питательного субстрата, а также выявлению степени влияния
различных факторов среды на устойчивость культур и последовательность изменения
видового состава в поликультурах.
Однако, как показывает практика, получение культур сенного настоя требует
проведения предварительных исследований, так как скорость сукцессионных
процессов, особенно на ранних стадиях, высока, смена видов может произойти
очень быстро, и не всегда понятны причины, вызывающие гибель или исчезновение
организмов. Поэтому для длительного поддержания их жизнедеятельности необходимо
изучить оптимальные условия существования организмов.
Актуальность проведенной работы заключается в том, что получаемые в
культуре сенного настоя простейшие гетеротрофы могут быть использованы как
тест-организмы для оценки токсичности водной среды.
Цель: пронаблюдать и описать видовой состав различных вариантов сенных
настоев, динамику численности простейших во времени.
Для достижения цели были поставлены следующие задачи:
) определить таксономический состав встречающихся в сенном настое
простейших;
) исследовать структуру видового состава инфузорий в различных сенных
настоях;
) выстроить на основе полученных данных сукцессионный ряд.
Автор выражает признательность и благодарность научному руководителю
к.б.н., доценту кафедры «Биоэкология» Пахомовой Нине Андреевне и к.б.н.,
доценту кафедры «Биоэкология» Минченок Елене Евгеньевне за помощь в подготовке
и оформлении материалов дипломной работы, а также доктору Masatsugu Yamazaki из университета Нагоя (Япония) за
помощь в определении видов Turbellaria.
1. Обзор
литературы
1.1 Общие
понятия о сукцессиях
Впервые термин «сукцессия» был предложен Г. Каулсоном в 1898 г.
Теория сукцессии изначально разрабатывалась геоботаниками. Клементс
заложил основы этой науки. По Клеметсу, сукцессия - процесс, в течение которого
один и тот же участок земной поверхности последовательно занимается различными
растительными сообществами. Единицей сукцессии является серия, заключающая
растительные сообщества, последовательно развивающиеся на данном участке, от
пионерной, или инициальной стадии через сукцессионные стадии к конечному
климаксу.
Также по Клементсу (1916), процесс сукцессии состоит из следующих этапов:
) возникновение незанятого жизнью участка;
) миграция на него различных организмов или их зачатков;
) приживание их на данном участке;
) конкуренции их между собой и вытеснение отдельных видов;
) преобразование живыми организмами местообитания, постепенной
стабилизации условий и отношений.
Хотя термины, введённые Клементсом, широко используют, существует две
принципиально различные парадигмы, в рамках которых смысл этих терминов
различен: континуализм и структурализм. Сторонники структурализма развивают
теорию Клементса, сторонники континуализма в принципе отвергают реальность
сообществ и сукцессий, считая их стохастическими явлениями и процессами
(поликлимакс, климакс-континуум). Процессы, происходящие в экосистеме, в этом
случае упрощают до взаимодействия видов, встретившихся случайным образом, и
абиотической средой.
По предложению В.Н.Сукачева (1954) различают по два типа автогенных и
аллогенных сукцессий.
Автогенные сукцессии разделяются на сингенез и эндоэкогенез.
При сингенезе изменения происходят под влиянием взаимоотношений между
видами сообщества, дифференциации экологических ниш без существенного изменения
видами условий местообитания. Основная роль сингенеза отмечается для начальных
стадий и для очень быстрых по масштабу времени сукцессий. Сингенетические
изменения необратимы и постепенны, могут проявляться как на первичных, так и на
вторичных местообитаниях, протекать по типу прогрессивной и, реже, регрессивной
сукцессии. Это природный процесс, но его "пусковым механизмом" могут
быть и антропогенные факторы. Примерами сингенеза могут служить первые стадии
зарастания новых субстратов и восстановление растительности на нарушенных
человеком территориях.
Эндоэкогенез - это изменение сообщества путём изменения условий среды,
сукцессия в классическом (узком) смысле. Примером может служить формирование
почвы на скалах.
Оба типа сукцессий тесно связаны. Условия среды изменяются и при
сингенезе, но значимость этих изменений по сравнению с взаимоотношениями
организмов мала. При эндоэкогенезе взаимоотношения между организмами также
оказывают определённое влияние на изменение системы в целом, но оно слабее
влияния биотической трансформации среды обитания.
Аллогенные сукцессии разделяются на гейтогенез и гологенез.
Гейтогенез - локальные (т.е. не связанные с общими тенденциями развития
ландшафта) аллогенные смены. Это сравнительно быстрая по масштабу времени
сукцессия (что, естественно, связано с её локальностью), вызываемая чаще всего
антропогенными факторами. Необратимый характер гейтогенеза может наблюдаться,
как правило, в экологически ранимых ландшафтах экстремальных условий среды.
Регрессивный гейтогенез называется ретрогрессией. Примером локальных внешних
воздействий, приводящих к гейтогенезу, может служить хозяйственная деятельность
человека - пастбищная дигрессия.
Гологенез - это изменения целых ландшафтов с более или менее параллельным
изменением сообществ и экосистем. Природный гологенез - процесс масштаба
тысячелетий, антропогенный протекает значительно быстрее. Природный гологенез
всегда постоянен и необратим, но последствия антропогенного гологенеза могут оказаться
обратимыми. Примером можно считать изменение растительности в процессе развития
форм рельефа под влиянием строительства водохранилищ.
Наиболее часто термин «сукцессия» используют для смен, происходящих в
сообществах. Чтобы разобраться в многообразии возможных смен, предложено много
классификаций на разных логических основаниях: по масштабу времени (быстрые,
средние, медленные, очень медленные), по обратимости (обратимые и необратимые),
по степени постоянства процесса (постоянные и непостоянные), по происхождению
(первичные и вторичные), по тенденциям изменения продуктивности (прогрессивные
и регрессивные), по тенденции изменения видового богатства (прогрессивные и
регрессивные), по антропогенности (антропогенные и природные), по характеру
происходящих во время сукцессии изменений (автотрофные и гетеротрофные). По
мнению А.П. Шенникова (1964), подобные классификации - это «классификации смен
по их причинам». В действительности, как указывает С.М. Разумовский (1999),
одни из данных классификаций - это классификации «причин, но не смен»,
поскольку одно дело причина, вызвавшая процесс, а совсем другое - сам процесс.
Связь тут далека от однозначной функциональной. Нередко такие классификации
делают на основе произвольно выбранных показателей, которые могут меняться во
время сукцессии.
Варминг (1901) в своей работе «Борьба между растительными сообществами»
пишет: «Каждое небольшое изменение в жизненных условиях тотчас же нарушает
устойчивое до сих пор равновесие, т.е. вызывает немедленно перемещение и
изменение во взаимоотношениях групп».
Изменения, происходящие при сукцессиях, обусловлены изменениями во
взаимоотношениях видов растений, а они связаны с различиями в отношении видов к
«экологическим факторам... и с разнообразием биологических особенностей жизненных
форм».
В любой сукцессионной серии темпы происходящих изменений постепенно
замедляются. Конечным итогом является формирование относительно устойчивой
стадии - климаксового сообщества, или климакса. Термин "климакс" был
предложен американскими исследователями Г. Уитфордом и Г. Каулесом в 1901 г.
Одум определил тенденции, которые следует ожидать в процессе сукцессии
экосистем (Одум, 1975), которые представлены в таблице 1.
Существуют три гипотезы достижения климакса.
Гипотеза моноклимакса Клементса (Одум, 1975; Уиттекер, 1980; Миркин,
1985; Дедю, 1990) - признание возможности существования в каждой природной зоне
одного устойчивого климаксового состояния вне зависимости от того, где бы ни
начиналась сукцессия. Гипотеза была предложена Клементсом в 1904 г.
Сущность гипотезы моноклимакса Клементса сводится к четырём положениям
(три первых Уиттекер (1974) считал обязательными):
) устойчивость;
) конвергенция (сближение признаков сообщества в ходе сукцессии);
) превалирование (преобладание этих климаксов по площади);
) аналогии климаксового сообщества организму (факультативное).
Кроме основных климаксов, Клементс допускал также существование большого
числа вспомогательных климаксов - отклонений под воздействием того или иного
существенного в территории фактора: субклимакс («задержавшаяся» сукцессионная
стадия), дисклимакс (нарушенный климакс), постклимакс (климакс более высокой по
уровню организации жизненной формы), преклимакс (обратный постклимаксу). Все
климаксы одного района объединялись Клементсом в панклимакс (высшая
синтаксономическая единица - климакс-формация).
Таблица 1. Сравнение развивающихся и зрелых систем (по Одуму,
1975)

Гипотеза моноклимакса с момента появления подвергалась критике, так как
три основных её положения оказались несопряженными, и потому ей на смену пришли
представления о поликлимаксе. Таким образом, гипотеза моноклимакса сегодня
представляет чисто исторический интерес. Однако в монографии Разумовского
(1981) автор «возвращается» к этим представлениям в связи с отстаиванием
представлений о дискретности растительного покрова.
Гипотеза поликлимакса Нихолса-Тенсли (Уиттекер, 1980) - признание
существования в одном климатическом районе нескольких климаксов, различающихся
эдафически. На территории с одинаковым климатом может образовываться несколько
климакс-формаций, что и составляет суть гипотезы. Представления о поликлимаксе
были развиты путём критики концепции моноклимакса американским ботаником
Нихолсом (1917) и английским экологом Тенсли (1920).
Гипотеза климакс-мозаики Уиттекера (Дедю, 1990) - признание наличия
непрерывно варьирующих под воздействием эдафических факторов климаксом в
однородной в климатическом отношений территории; климакс в каждой точке
признается индивидуальным и трактуется как мозаика видовых популяций,
совмещённых условиями среды.
Ниже представлена схема, иллюстрирующая гипотезы достижения климакса
(рис. 1).

Рис. 1. Гипотезы достижения климакса (по Дедю, 1990)
Подразделение сукцессий на автотрофные и гетеротрофные имеет в своём
основании идею о том, что отношение первичной продукции (Р) к дыханию (R) в
ходе сукцессии «приближается к единице (равновесию)» (Одум, 1986).
Автотрофные сукцессии - это сукцессии, начинающиеся с сообществ, у
которых:
/R>1, (1)
гетеротрофные -
/R<1, (2)
при этом в гетеротрофной сукцессии «стационарного состояния достичь
невозможно» (Одум, 1986).
Представление о том, что:
/R=1, (3)
это состояние равновесия, является экстраполяцией представлений экологов,
изучавших некоторые наземные экосистемы, на всю биосферу. Каждое сообщество при
этом отождествляют с закрытой системой, основным, если не единственным,
источником энергии в котором является фотосинтез входящих в состав сообщества
растений. Между тем любое сообщество - открытое, что особенно относится к
водным сообществам, где доля аллохтонного органического вещества, энергию
которого использует сообщество, достигает 100%. У водных сообществ за пределами
фитали (за исключением сообществ хемобиоса) вообще нет первичной продукции.
Нередко и в пределах фитали первичная продукция очень незначительна, например,
во многих сообществах рыхлых грунтов. Такие сообщества находятся в состоянии
равновесия миллионы лет. Поэтому соотношение первичной продукции и дыхания как
показатель устойчивости применительно к ним оказывается совершенно
неинформативным. Теория, применимая лишь к ограниченному числу объектов, не
может претендовать на роль общей. Чтобы не менять теорию, такие сообщества
часто объявляют неполноценными. Странно считать сообщества, населяющие больше
половины поверхности Земли, неполноценными. Если придерживаться представлений,
описанных Одумом, то придётся отнести к первичным продуцентам любой низший
трофический уровень, поставляющий энергию в сообщество, в том числе и потребителей
детрита, хотя это и кажется непривычным. Более того, если начать копать глубже,
то принятая в настоящее время практика не вполне последовательна. Сейчас к
первичным продуцентам относят «автотрофные организмы, в основном зелёные
растения, которые могут производить пищу из простых неорганических веществ»
(Одум, 1986), т. е. получающие энергию для жизнедеятельности и синтеза нового
органического вещества за счёт любых источников, кроме окисления другого
органического вещества. При таком определении вестиментиферы чёрных
курильщиков, использующие энергию окисления восстановленных соединений
металлов, являются автотрофами (хемоавтотрофами). Тот же вид, живущий на трупе
кита или затопленном сухогрузе, использующий энергию окисления органического
вещества, становится уже гетеротрофом. В любом случае отношение P/R и деление
сукцессий на автотрофные и гетеротрофные имеют ограниченное применение.
Классический пример гетеротрофной сукцессии - динамика простейших в сенном
настое (Woodruff, 1912) - в действительности
является нормальным процессом потребления органики редуцентами.
1.2
Гетеротрофные сукцессии
Микробный микрокосм сыграл центральную, роль в развитии экологии, что
иногда недооценивается. Первая опубликованная работа в данной области
принадлежит Dallinger, который в своем выступлении в качестве президента
Королевского Общества Микроскопии в 1887 году, описал свою попытку определить,
можно ли путём изменения окружающей среды проследить адаптации на примере
короткоживущих организмов.впервые показал, что это возможно осуществить в
лабораторных условиях.
Почти 20 лет спустя, Woodruff провёл эксперименты с сенным настоем и
пришел к выводу, что взаимодействия между организмами были важной движущей
силой в сукцессионной последовательности простейших. Он отметил, что
определённая последовательность появления простейших хоть и произошла в данном
настое, но чёткая картина не была выявлена. Woodruff предположил, что воздух,
вода и сено могут быть источниками популяций простейших. Как только
бактерии действуют на сено, чтобы сделать его доступным для питания другим
организмам, начинается рост простейших.
Позже, Гаузе проведено ещё несколько экологических экспериментов по
конкуренции и хищничества с использованием микрокосма, в состав которого
входили бактерии, дрожжи и простейшие. Ряд ключевых принципов в области
экологии относятся к этим исследованиям. Например, по оценке параметров роста
для каждого вида, Гаузе был в состоянии предсказать, какой вид будет
конкурентоспособным и доминирующим. Интерпретация этой работы Хардином в
конечном счете привела к принципу конкурентного исключения. Кроме того, работа
Гаузе основывалась на динамике хищник - жертва с использованием Didinium
nasutum и Paramecium caudatum и продемонстрировала важное значение
пространственных убежищ и иммиграции на соотношение количества хищника и
жертвы.
Пионерские эксперименты Гаузе в микрокосме были использованы для изучения
различных тем в области экологии, например, сукцессии, разнообразии и
устойчивости отношений, динамики хищник - жертва, сосуществовании конкурентов и
сосуществование широкого спектра генералистов и специалистов.
Последовательность смены видов среди простейших в сенном настое была
определена Buchsbaum и Buchsbaum (1957). По их данным, после введения бактерий
в первоначально стерильный настой и последующего его брожения, среда стала
содержать подходящую для простейших пищу. Монады (примитивные жгутиконосцы)
появились первые, затем они были заменены Colpoda, которые впоследствии
стали доминирующим населением. Брюхоресничные и равноресничные инфузории
появились в небольшом количестве, их численность быстро снизилась. Поскольку
жизнь стабилизировалась, популяции коловраток и ракообразных стали
доминирующими.
Одум (1963) описал последовательность в сообществах планктона и сенного настоя.
В последнем случае наблюдения велись в течение месяца, жгутиконосцы появились
первые, на смену которым пришли небольшие кругоресничные инфузории, и далее
появились специализированные простейшие, такие как Hypotricha и
Vorticella и, наконец, Amoeba и коловратки возникли после того, как другие
организмы погибли.
Биотические взаимоотношения этих организмов также были рассмотрены.
Например, метаболический продукт одного организма может являться как объектом
питания, так и токсичным веществом для некоторых других организмов (Burkholder,
1952). Взаимосвязи, ведущие к увеличению или уменьшению видов, были обсуждены
Hariston и соавт. (1968).
Также на основе вышеназванных работ были проведены более глубокие
исследования гетеротрофной сукцессии в микроэкосистеме в целом (Gorden, 1969;
Lawton, 1995; Drake, 1996), и прослежено сосуществование микроорганизмов в этой
системе, к примеру, Paramecium aurelia и её хищника Didinium nasutum
(Luckinbill, 1973).
Сукцессии в природе чрезвычайно разномасштабны. Их можно наблюдать в
пробирках с сенным настоем, где сменяют друг друга разные виды микроорганизмов
и простейших (рис. 2), в лужах и прудах, в одной и той же порции листового
опада в лесу, где происходят закономерные смены сапрофитов и сапрофагов, на
стволах отмирающих деревьев, в пнях, на кротовинах, выбросах из нор сурков,
зарастающих отмелях, выветривающихся скалах, на заброшенных пашнях, насыпях, на
болотах, лугах, в лесах и т. д.
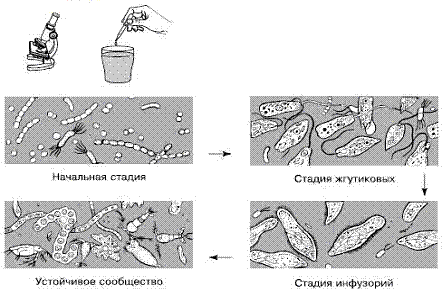
Рис.
2. Смена простейших в сенном настое (по Дажо, 1975)
Иерархичность
в организации сообществ проявляется и в иерархичности сукцессионных процессов:
более крупные преобразования биоценозов складываются из более мелких. Даже в
стабильных экосистемах с хорошо отрегулированным круговоротом веществ постоянно
осуществляется множество локальных сукцессионных смен, поддерживающих сложную
внутреннюю структуру сообществ.
Не
меньшие преобразования происходят и в энергетическом балансе системы (рис. 3).
С энергетических позиций сукцессия - такое неустойчивое состояние сообщества,
которое характеризуется неравенством двух показателей: общей продуктивности и
энергетических трат всей системы на поддержание обмена веществ.

Рис.
3. Изменение энергетических показателей сообществ при развитии соснового леса
(вверху) и лабораторной культуры простейших в сенном настое (внизу) (по Одуму,
1975):
Пв
- продукция валовая; Д - траты на дыхание; Пч - продукция чистая
(заштриховано); Б - биомасса
В
ходе сукцессии общая биомасса сообщества сначала возрастает, но затем темпы
этого прироста снижаются, и на стадии климакса биомасса системы
стабилизируется. Это происходит потому, что на первых этапах сукцессии, когда
видовой состав сообществ ещё беден и пищевые цепи коротки, не вся часть
прироста растительной массы потребляется гетеротрофами. Таким образом,
относительно высока чистая продукция сообщества, которая идёт на увеличение его
биомассы. Накапливаются как общая масса живых организмов, так и запасы
мёртвого, неразложившегося органического вещества. В зрелых, устойчивых
экосистемах практически весь годовой прирост растительности поступает и
расходуется в цепях питания гетеротрофами, поэтому чистая продукция биоценоза,
его «урожай», приближается к нулю.
С
удлинением цепей питания увеличивается эффективность использования поступающей
энергии, иначе - КПД всей системы, так как одна и та же порция энергии идёт на
поддержание большого количества биомассы. Знание этих закономерностей имеет
большое значение в практической деятельности человека.
Изымая
избыток чистой продукции из биоценозов, находящихся в начале сукцессионных
рядов, мы задерживаем сукцессию, но не подрываем основу существования
сообщества. Вмешательство же в стабильные, климаксные системы, с большой
полнотой расходующие энергию на «свои» нужды, неминуемо вызывает нарушения
сложившегося равновесия. Пока нарушения не превышают самовосстановительной
способности ценоза, возникающие демутационные смены могут вернуть его к
исходному состоянию. Этим пользуются, например, при рациональном планировании
рубок леса. Но если сила и частота воздействия выходят за рамки этих
возможностей, то первоначально устойчивое, богатое видами сообщество постепенно
деградирует, сменяясь производными с малой способностью к самовозобновлению.
Таким
образом, сообщество не может одновременно сочетать два противоположных
свойства: быть высокостабильным и давать большой запас чистой продукции,
который можно было бы изымать без вреда для самого ценоза.
Эксперименты
проводятся и в настоящее время. К примеру, Shamaporn Nivasaputra и
Thapana Chontananarth (2007) проследили за изменением сообщества простейших в
настое на основе рисового сена. Наблюдение в культуре рисовой соломы было
проведено в течение зимы 2007 года. Было установлено, что в 28 из 30 ёмкостей
(что составляет 93,33% от общего количества) наблюдалась некоторая
закономерность. Примерный сукцессионный ряд в 28 ёмкостях: первоначально
наблюдается обилие бактерий и жгутиковых простейших, в то время как инфузории и
саркодовые простейшие были отмечены в климаксном состоянии. Для простейших в
общем обнаружены 6 родов, которые можно разделить на 3 класса простейших, как
показано в таблице 2.
Таблица 2. Появление
различных классов простейших в настое
|
Дата обнаружения
|
Класс
|
Представители
|
|
11-20
|
Mastigophora
|
Chrysidella sp., Monas sp.
and Warnowia sp.
|
|
13-16
|
Ciliata
|
Paramecium
aularia
|
|
16-25
|
Sarcodina
|
Naegleria sp. and
Pyxidicular sp
|
В этом исследовании есть 2 пробирки, отличающиеся по видовому составу
(6,67%). Первоначально в настое развивались бактерии, после обнаружены
простейшие, первичные простейшие были замещены Mastigophora. Эти простейшие
жгутиковые часто встречаются в воде с высоким содержанием кислорода, но с
низким уровнем органических веществ. Sarcodina были обнаружены в воде с низким
содержанием кислорода, но с высоким уровнем органических веществ с рН в
диапазоне от 6,89 до 7,27 (Kudo,
1966). Для климаксного сообщества в этой последовательности простейших является
Sarcodina включают Naegleria sp., потому что этот вид устойчив в широком диапазоне рН
(7,20 - 7,84) и высокой концентрации органических соединений.
Изучение культуры, проведенное Eddy в начале ХХ века (Eddy,
1912) показало такие же результаты с теми же организмами. Существует разница с Eddy на стадии развития инфузорий. В
работах Shamaporn Nivasaputra и Thapana Chontananarth (2007) были
получены только ресничные простейшие в 8 ёмкостях из 28 ёмкостей, подъём и спад
их численности произошёл очень быстро (3 дня). На рис. 4 и 5 представлены
результаты исследования Shamaporn Nivasaputra и Thapana Chontananarth.
Количество простейших (103шт. /40 см3)

Рис. 4 Общая численность инфузорий

Рис. 5. Среднее
количество Sarcodina
.3
Биотестирование и биоиндикация
Биологический контроль окружающей среды включает две основные группы
методов: биоиндикацию и биотестирование. Применение в качестве биоиндикаторов
растений, животных и даже микроорганизмов позволяет проводить биомониторинг
воздуха, воды и почвы. Благодаря специальным индексам и коэффициентам
результаты биоиндикации оказываются достоверными и сопоставимыми.
Биотестирование как метод исследования используют специалисты различных
областей науки: в экологической токсикологии для анализа вод и почв, в
гуманитарной и ветеринарной медицине для исследования свойств внутренних сред
высших организмов, в сельском хозяйстве для экспресс-тестирования кормов на
общую токсичность, в химии для первичной оценки свойств новых веществ и так
далее. При этом само понятие "биотестирование" зачастую истолковывают
по-разному, приспосабливая этот термин к тому или иному узкому кругу задач и
объектов, тем самым, обедняя его содержание и даже вкладывая в него различный
смысл. Ещё сложнее обстоит дело с производными терминами в этой области знания
- они не только не имеют устоявшихся определений, но зачастую даже не являются
общепринятыми. Комплексное рассмотрение биологических тестов позволило
вычленить их общие черты и составить универсальную формулировку определения, в
котором преодолены проблемы узости сферы использования биотестирования и
адекватности описания биологической основы, на которой проводится тест. Итак,
биотестирование - это оценка свойств изучаемого объекта по его воздействию на
биологическую тест-систему (далее - тест-систему) в стандартных условиях. Данная
формулировка включает в сферу рассмотрения любые методы, в которых для изучения
какого-либо явления используются биологические объекты. Это позволяет
анализировать и сопоставлять биологические тесты, применяемые в разных
областях, с единой точки зрения.
Чтобы в дальнейшем различать очень близкие по целям применения и
смысловому использованию понятия, поясним также значение термина
«биоиндикация».
Биоиндикация (bioindication) - обнаружение и определение
экологически значимых природных и антропогенных нагрузок на основе реакций на
них живых организмов непосредственно в среде их обитания. Биологические
индикаторы обладают признаками, свойственными системе или процессу, на
основании которых производится качественная или количественная оценка тенденций
изменений, определение или оценочная классификация состояния экологических
систем, процессов и явлений. В настоящее время можно считать общепринятым, что
основным индикатором устойчивого развития в конечном итоге является качество
среды обитания.
Для оценки параметров среды используются стандартизованные реакции живых
организмов (отдельных органов, тканей, клеток или молекул). В организме,
пребывающем контрольное время в условиях загрязнения, происходят изменения
физиологических, биохимических, генетических, морфологических или иммунных
систем. Объект извлекается из среды обитания, и в лабораторных условиях
проводится необходимый анализ. Живой организм может тестироваться также в
специальных камерах или на стендах, где создаются условия изучаемого
загрязнения (что очень важно для выявления реакций организма на то или иное
доминирующее загрязнение или целый комплекс известных загрязняющих веществ на
данной территории обитания).
Хотя подходы очень близки по конечной цели исследований, надо помнить,
что биотестирование осуществляется на уровне молекулы, клетки или организма и
характеризует возможные последствия загрязнения окружающей среды для биоты, а
биоиндикация - на уровне организма, популяции и сообщества и характеризует, как
правило, результат загрязнения. Живые объекты - открытые системы, через которые
идёт поток энергии и круговорот веществ. Все они в той или иной мере пригодны
для целей биомониторинга.
Контроль качества окружающей среды с использованием биологических
объектов в последние десятилетия оформился как актуальное научно-прикладное
направление. При этом необходимо отметить дефицит учебной литературы по этим
вопросам и большую потребность в ней.
Реализация основных принципов устойчивого развития цивилизации в
современных условиях возможна лишь при наличии соответствующей информации о
состоянии среды обитания в ответ на антропогенное воздействие, собранной в ходе
проведения биологического мониторинга. Оценка качества среды является ключевой
задачей любых мероприятий в области экологии и рационального
природопользования. Сам термин «мониторинг» (от англ. monitoring - контроль) подразумевает проведение
мероприятий по непрерывному наблюдению, измерению и оценке состояния окружающей
среды. Комплексный подход в проведении биологического мониторинга (сочетание
методов биоиндикации и биотестирования, использование объектов разных уровней
организации) при систематическом наблюдении позволяет судить о перспективах
изменения структуры сообществ, продуктивности популяций и устойчивости
экосистем по отношению к антропогенным факторам.
Объектами мониторинга являются биологические системы и факторы,
воздействующие на них. При этом желательна одновременная регистрация
антропогенного воздействия на экосистему и биологического отклика на
воздействие по всей совокупности показателей живых систем. Необходимо
проведение многофакторного анализа с учётом наиболее типичных антропогенных
воздействий (например, химических веществ), а также изменений природных
факторов среды, уровень которых меняется вследствие антропогенного влияния. В
первую очередь учитывается изменение численности видов и видового состава
ценозов. Важно фиксировать также возможные изменения в природных популяциях,
например нарушения эмбрионального развития (уродств) и симметрии взрослых
особей в пределах популяции. Необходимо выявлять быстрый «отклик» организмов
или популяций и результаты стойких последствий, так как часть изменений может
быть отрегулирована биосистемами.
Примеры применения методов биоиндикации и биотестирования в практике
экологической экспертизы природных водоёмов и питьевых водоисточников
демонстрируют, что пороговые концентрации химических поллютантов, нарушающие
жизнедеятельность организмов-биотестов, находятся ниже принятых значений ПДК.
Постоянное присутствие поллютантов даже в низких концентрациях приводит к
снижению видового разнообразия гидробионтов за счёт исчезновения наиболее
чувствительных к качеству воды видов. Такие изменения в биоценозах
устанавливаются методами биоиндикации - определением индексов и показателей сапробности.
Состояние биологической системы (организм, популяция, биоценоз) в той или
иной степени характеризует воздействие на неё природных или антропогенных
факторов и условий среды и может применяться для их оценки.
Биоиндикаторы (от био и лат. indico - указываю, определяю) - организмы, присутствие,
количество или особенности развития которых служат показателями естественных
процессов, условий или антропогенных изменений среды обитания. Их индикаторная
значимость определяется экологической толерантностью биологической системы. В
пределах зоны толерантности организм способен поддерживать свой гомеостаз.
Любой фактор, если он выходит за пределы «зоны комфорта» для данного организма,
является стрессовым. В этом случае организм реагирует ответной реакцией различной
интенсивности и длительности, проявление которой зависит от вида и является
показателем его индикаторной ценности. Именно ответную реакцию определяют
методы биоиндикации. Биологическая система реагирует на воздействие среды в
целом, а не только на отдельные факторы, причём амплитуда колебаний
физиологической толерантности модифицируется внутренним состоянием системы -
условиями питания, возрастом, генетически контролируемой устойчивостью.
Многолетний опыт учёных разных стран по контролю состояния окружающей
среды показал преимущества, которыми обладают живые индикаторы:
- в условиях хронических антропогенных нагрузок могут реагировать даже на
относительно слабые воздействия вследствие кумулятивного эффекта; реакции
проявляются при накоплении некоторых критических значений суммарных дозовых
нагрузок;
- суммируют влияние всех без исключения биологически важных воздействий и
отражают состояние окружающей среды в целом, включая её загрязнение и другие
антропогенные изменения;
- исключают необходимость регистрации химических и физических параметров,
характеризующих состояние окружающей среды;
- фиксируют скорость происходящих изменений;
- вскрывают тенденции развития природной среды;
указывают пути и места скоплений в экологических системах различного рода
загрязнений и ядов, возможные пути их попадания в пищу человека;
- позволяют судить о степени вредности любых синтезируемых человеком
веществ для живой природы и для него самого, причем дают возможность
контролировать их действие.
Выделяют две формы отклика живых организмов, используемых в целях
биоиндикации, - специфическую и неспецифическую. В первом
случае происходящие изменения связаны с действием одного какого-либо фактора.
При неспецифической биоиндикации различные антропогенные факторы вызывают
одинаковые реакции.
В зависимости от типа ответной реакции биоиндикаторы подразделяют на чувствительные
и кумулятивные. Чувствительные биоиндикаторы реагируют на стресс
значительным отклонением от жизненных норм, а кумулятивные накапливают
антропогенное воздействие, значительно превышающее нормальный уровень в
природе, без видимых изменений.
В качестве биоиндикаторов могут быть использованы представители всех
«царств» живой природы. Для биоиндикации не пригодны организмы, поврежденные
болезнями, вредителями и паразитами. Идеальный биологический индикатор должен
удовлетворять ряду требований:
- быть типичным для данных условий;
- иметь высокую численность в исследуемом экотопе;
- обитать в данном месте в течение ряда лет, что даёт возможность
проследить динамику загрязнения;
находиться в условиях, удобных для отбора проб;
- давать возможность проводить прямые анализы без предварительного
концентрирования проб;
- характеризоваться положительной корреляцией между концентрацией
загрязняющих веществ в организме-индикаторе и объекте исследования;
- использоваться в естественных условиях его существования;
- иметь короткий период онтогенеза, чтобы была возможность отслеживания
влияния фактора на последующие поколения.
Ответная реакция биоиндикатора на определённое физическое или химическое
воздействие должна быть четко выражена, т.е. специфична, легко регистрироваться
визуально или с помощью приборов.
При выборе индикатора необходимо принимать во внимание соображения
экономии и учитывать характер использования тех или иных организмов. Например,
широко распространенные на исследуемой территории и не занесённые в «Красную
книгу».
.4 Задачи
и приёмы биотестирования
В выявлении антропогенного загрязнения среды наряду с
химико-аналитическими методами находят применение приемы, основанные на оценке
состояния отдельных особей, подвергающихся воздействию загрязненной среды, а
также их органов, тканей и клеток. Их применение вызвано технической
усложнённостью и ограниченностью информации, которую могут предоставить
химические методы. Кроме того, гидрохимические и химико-аналитические методы
могут оказаться неэффективными из-за недостаточно высокой их чувствительности.
Живые организмы способны воспринимать более низкие концентрации веществ, чем
любой аналитический датчик, в связи с чем биота может быть подвержена
токсическим воздействиям, не регистрируемым техническими средствами.
Биоиндикация предусматривает выявление уже состоявшегося или
накапливающегося загрязнения по индикаторным видам живых организмов и
экологическим характеристикам сообществ организмов. Пристальное внимание в
настоящее время уделяется приёмам биотестирования, т.е. использования в
контролируемых условиях биологических объектов в качестве средства выявления
суммарной токсичности среды. Биотестирование представляет собой методический
прием, основанный на оценке действия фактора среды, в том числе и токсического,
на организм, его отдельную функцию или систему органов и тканей.
Кроме выбора биотеста существенную роль играет выбор тест-реакции - того
параметра организма, который измеряется при тестировании.
Наиболее информативны интегральные параметры, характеризующие общее
состояние живой системы соответствующего уровня. Для отдельных организмов к
интегральным параметрам обычно относят характеристики выживаемости, роста,
плодовитости, тогда как физиологические, биохимические, гистологические и
прочие параметры относят к частным. Для популяций интегральными параметрами
являются численность и биомасса, а для экосистем - характеристики видового
состава, активности продукции и деструкции органического вещества.
С увеличением интегральности тест-реакции повышается «экологический
реализм» теста, но обычно снижаются его оперативность и чувствительность.
Функциональные параметры оказываются более лабильными, чем структурные, а параметры
клеточного и молекулярного уровней проигрывают в отношении экологической
информативности, но выигрывают в отношении чувствительности, оперативности и
воспроизводимости.
В оптимальных условиях организм реагирует на воздействие среды
посредством сложной физиологической системы буферных гомеостатических
механизмов. Эти механизмы поддерживают оптимальное протекание процессов
развития. Под воздействием неблагоприятных условий механизмы поддержания
гомеостаза могут быть нарушены, что приводит к состоянию стресса. Такие
нарушения могут происходить до появления изменений обычно используемых
параметров жизнеспособности. Таким образом, методология биотестирования,
основанная на исследовании эффективности гомеостатических механизмов, позволяет
уловить присутствие стрессирующего воздействия раньше, чем многие обычно
используемые методы.
1.5 Методы
биотестирования
Для того чтобы быть пригодными для решения комплекса современных задач,
методы биотестирования, используемые для оценки среды, должны соответствовать
следующим требованиям:
быть применимыми для оценки любых экологических изменений среды обитания
живых организмов;
характеризовать наиболее общие и важные параметры жизнедеятельности
биоты;
быть достаточно чувствительными для выявления даже начальных обратимых
экологических изменений;
быть адекватными для любого вида живых существ и любого типа воздействия;
быть удобными не только для лабораторного моделирования, но также и для
исследований в природе;
быть достаточно простыми и не слишком дорогостоящими для широкого
использования.
Одним из наиболее важных требований при оценке состояния среды является
чувствительность применяемых методов. Потребность в таких методах особенно
возрастает в настоящее время, когда в силу повышенного внимания к проблемам
охраны природы и в связи с развитием природоохранных мероприятий становится
необходимым оценивать не только и не столько существенные, как правило, уже
необратимые изменения в среде, но первоначальные незначительные отклонения,
когда ещё возможно вернуть систему в прежнее нормальное состояние.
Другое важное требование - универсальность как в отношении физического,
химического или биологического оцениваемого воздействия, так и типа экосистем и
вида живых существ, по отношению к которым такая оценка проводится. Причём это
необходимо как в отношении отдельных агентов, так и кумулятивного воздействия
любого их сочетания (включая весь комплекс как антропогенных, так и
естественных факторов).
Система должна быть относительно простой и доступной, пригодной для
широкого использования. В настоящее время существует ряд современных
молекулярно-биологических тестов качества среды, но в силу высокой
технологической сложности и стоимости их применение оказывается ограниченным.
При этом возникает вопрос: нужно ли прибегать к таким сложным методам при
решении общей задачи мониторинга состояния среды и нельзя ли получить сходную
информацию более доступным способом.
«Подходами» можно условно назвать группы методов, характеризующих сходные
процессы, происходящие с тест-объектами под влиянием антропогенных факторов.
1.5.1
Биохимический подход
Стрессовое воздействие среды можно оценивать по эффективности
биохимических реакций, уровню ферментативной активности и накоплению
определенных продуктов обмена. Изменения содержания в организме определённых
биохимических соединений (например, терпеноидов), показателей базовых
биохимических процессов (например, концентрации хлорофилла у фотосинтезирующих
растений) и структуры ДНК в результате биохимических реакций (например, при
оксидантном стрессе) могут обеспечить необходимую информацию о реакции
организма в ответ на стрессовое воздействие.
1.5.2
Морфологический подход
В условиях техногенного воздействия на природные экосистемы снижение
численности популяций происходит в значительной мере за счёт эмбриональной и
личиночной смертности. Эмбрионы и личинки - наиболее чувствительные к
повреждающим факторам фазы жизненного цикла гидробионтов. Воздействие на
организм стрессирующих факторов приводит к отклонениям от нормального строения
различных морфологических признаков. В московских и подмосковных водоёмах за
последние 20 лет катастрофически возрос процент уродливых личинок лягушек и
жаб. Отмечено появление рыб с нарушениями эмбрионального морфогенеза, т.е. с
различными аномалиями (асимметрия тела и т.д.). В пригородных водоёмах и малых
реках, по данным гидробиологического мониторинга, исчезли многие виды
гидробионтов. Процессы воспроизведения организмов - это сложная цепь
взаимообусловленных событий, любое из звеньев которой может быть нарушено
воздействием токсичной среды.
Для диагностики воздействия загрязнений на морфологические характеристики
применяются методы оценки флуктуирующей асимметрии.
Симметрия как вид согласованности отдельных частей живых организмов имеет
общебиологическое значение. При работе с биологическими объектами в настоящее
время используется классификация асимметрий (нарушения симметрии) по Л. Ван
Валену (Van Valen, 1962), согласно которой они подразделяются на три типа:
) направленная асимметрия, когда какая-то структура развита на одной
стороне больше, чем на другой (сердце млекопитающих);
) антиасимметрия - большее развитие структуры на одной из сторон (правша
и левша в популяции человека);
) флуктуирующая асимметрия - незначительные ненаправленные отклонения от
строгой билатеральной симметрии.
Флуктуирующая асимметрия является результатом неспособности организмов
развиваться по точно определенному плану. Различия между сторонами не являются
генетически детерминированными и не имеют адаптивного значения. Выступая в
качестве меры стабильности развития, флуктуирующая асимметрия характеризует
состояние морфогенетического гомеостаза - способности организма к формированию
генетически детерминированного фенотипа при минимальном уровне онтогенетических
нарушений. Таким образом, флуктуирующая асимметрия может быть охарактеризована
как одно из наиболее обычных и доступных для анализа проявлений случайной
изменчивости развития.
Возможность использования асимметрии в биоиндикации показана многими
авторами, которые убедительно доказали на примере различных видов растений и
животных, что величина асимметрии реагирует на различные стрессоры
антропогенного характера и может являться мерой нарушения развития организма.
Флуктуирующая асимметрия - это один из общих онтогенетических показателей,
характеризующий стабильность индивидуального развития, дающий оценку состояния
природных популяций и зависящий от состояния среды. Величина флуктуирующей
асимметрии и её зависимость от определенных факторов может быть определена лишь
на популяционном уровне. Кроме того, В. М. Захаровым показано, что
флуктуирующая асимметрия является практически единственной формой
фенотипической изменчивости с известной причиной обусловленности.
1.5.3
Физиологический подход
Одна из наиболее важных характеристик, высокочувствительная к стрессовому
воздействию среды, - энергетика физиологических процессов. Наиболее экономичный
энергетический обмен имеет место лишь при строго определённых условиях среды,
которые могут быть охарактеризованы как оптимальные. Интенсивность энергетического
обмена аэробного организма может быть определена посредством измерения скорости
потребления кислорода. При оптимальных условиях организм находится на самом
низком энергетическом уровне, при любых негативных изменениях среды обитания
потребность в кислороде будет увеличиваться.
Для характеристики энергетического обмена две величины являются
фундаментальными: основной обмен и максимальный обмен. Основной обмен отражает
минимальный уровень потребления энергии, необходимый для обеспечения
нормального функционирования организма при отсутствии каких-либо внешних
воздействий. Максимальный обмен соответствует предельному количеству энергии,
которое организм способен выработать в случае необходимости. Разность между
этими величинами представляет энергетический ресурс адаптации конкретного вида
животных, поскольку основной и максимальный уровни обмена являются
видоспецифическими величинами.
Другая базовая характеристика, перспективная для оценки стрессовых
воздействий, - темп и ритмика ростовых процессов.
Важной характеристикой физиологических процессов является поведенческая
активность живых организмов.
В качестве тест-функций применяются физиологические параметры
пресноводных беспозвоночных гидробионтов разных уровней филогенеза.
Свойства внешней среды, и в частности гидросферы, проявляются в
интенсивности воздействия на организм или популяцию отдельных факторов или их
комбинаций. Вещества, поступающие в водоём антропогенным путём, могут оказывать
регулирующее, трофическое, токсическое и информативное воздействие на
гидробионтов. При незначительных концентрациях в водоёме эти вещества можно
выявить, оценивая физиологический статус гомеостатических показателей
организма, которые могут изменяться при сдвигах в окружающей среде.
Вопрос о роли поведения в иерархии индикационных показателей состояния
окружающей среды поднимали в своих работах такие учёные, как Н.С. Строганов,
А.Д. Слоним, Б.А. Флёров, Л.П. Брагинский, Н.А. Тушмалова. Степень отклонения
от стандартных поведенческих реакций животных служит одним из первичных,
регистрируемых на визуальном уровне сигналов изменений в окружающей среде и,
несомненно, может быть отнесена к реакциям «первого ранга».
Наибольший интерес представляют типы поведения, относящиеся к
эволюционно-универсальным реакциям, свойственным всем эукариотам, включая
человека. К таким феноменам относятся спонтанная двигательная активность как
врожденная форма поведения и память - приобретенная форма поведения.
В опытах на инфузориях показателями исходного функционального состояния
служили объективно регистрируемые реакции: спонтанная двигательная активность,
уровень спонтанных сокращений, уровень пищевой возбудимости, состояние ядерного
аппарата. Была продемонстрирована применимость основных физиологических
показателей, используемых в опытах на позвоночных животных, для определения
функционального состояния организмов, лишённых нервной системы. Исследования
поведенческих реакций в ответ на внешнее воздействие проводились на
пресноводной гидре Hydra attenuate, имеющей примитивную нервную систему.
Оценивались реакции привыкания к раздражителю, при этом критерием выработки
привыкания служило сокращение щупалец гидры. В экспериментах на плоских червях
планариях Polycelis nigra, Euplanaria gonocephala, Dugesia
tigrina (примитивный мозг - зачатки цефализации) были изучены основные
реакции гидробионтов на экологически значимые раздражители. В дальнейшем для
оценки качества водной среды стал успешно применяться метод выработки условных
рефлексов у планарий.
Целостное поведение животных рассматривается как лабильное взаимодействие
врождённых и приобретённых элементарных реакций, необходимое для быстрой и
эффективной адаптации к условиям среды. Изучение поведения сложно и требует
тщательных наблюдений в природе, подкрепленных лабораторными экспериментами.
Возникшие на заре эволюции закономерности поведения у простейших сохраняются и
у более развитых животных.
Таким образом, поведение является эволюционно обусловленным показателем
физиологического состояния животного. На основании изменений в поведенческих
феноменах одного вида животных можно прогнозировать нарушения поведения и
других видов. Выбор форм поведения для биотестирования определяется их
чувствительностью к изменениям, происходящим в окружающей среде.
1.6
Биотестирование с использованием инфузорий
В числе организмов, на которых проводят биотестирование, присутствуют
представители подцарства простейших. История применения Protozoa в качестве
тест-организмов насчитывает не одно десятилетие. В то же время, развитие иных
методов токсикологического анализа, таких как химические, физико-химические,
иммунологические, а также использование биологической оценки на организмах из
других систематических групп не снижают актуальности биотестов на простейших.
Об этом говорит значительное число методических разработок, рекомендаций и
публикаций, постоянно появляющихся в специализированных изданиях, а также их
широкое использование на практике. Эти методы обладают высокой
чувствительностью, экспрессностью, надёжностью, универсальностью и малой
себестоимостью. Они просты в проведении, поддаются инструментализации и
автоматизации, а их результаты легко интерпретируемы. В отличие от химических и
физико-химических методов анализа, биотестирование на инфузориях позволяет
прогнозировать интегральное воздействие изучаемого объекта на живые организмы,
поскольку реакция биологической тест-системы зависит не только от отдельных
токсичных соединений, содержащихся в объекте исследования, но и от их
взаимодействия между собой, а также от присутствия веществ, обладающих ярко
выраженным влиянием на токсичность указанных соединений. А по сравнению с
биотестами на высших животных оно обладает значительными преимуществами в
экономической, методической и этической сферах.
Хотя в рассматриваемой области накоплен богатый фактический материал, в
биологической науке до сих пор отсутствует единая концепция, описывающая
биотестирование на инфузориях. Определения основных используемых терминов не
получили точных и исчерпывающих формулировок, недостаточно изучена культура
инфузорий, как система, по реакции которой производится оценка объекта внешней
среды, не выявлены закономерности её реакции на неблагоприятные воздействия, не
сформулированы принципы выбора тест-организмов. Как следствие, отсутствуют
единые методы подготовки проб и культуры инфузорий как тест-системы к
проведению анализа.
Недостаточность теоретического описания биотестирования на инфузориях
влечёт за собой разнообразные проблемы в практическом воплощении конкретных
методик. Тест-организмы зачастую подбираются без учета их биологических особенностей,
инфузории культивируют на нестандартных средах и в произвольных условиях, во
многих методиках процесс подготовки тест-системы к анализу отсутствует как
таковой, уровень чувствительности наблюдаемых тест-реакций далеко не всегда
соответствует поставленным задачам, используемые для оценки степени проявления
тест-реакции критерии в ряде случаев имеют малую информативность по сравнению с
альтернативными. Всё это приводит к огрублению результатов, неверной их
интерпретации, снижению объективности выводов, а зачастую и к дискредитации
биотестирования, как метода научного исследования.
По этим причинам в настоящее время особую важность приобретает вопрос о
комплексном подходе к биотестированию. В первую очередь необходим глубокий
теоретический анализ этой группы методов. Создание единой концепции оценки
свойств объектов окружающей среды по реакции культуры инфузорий должно
способствовать развитию, стандартизации и унификации уже разработанных методов,
а также созданию новых, более совершенных.
Таким образом, можно заключить, что исследования в указанной области
являются актуальными и имеют важное теоретическое и прикладное значение.
В первую очередь внимание исследователей привлекли те виды
свободноживущих инфузорий, которые имеют в своем жизненном цикле стадию покоя и
способны образовывать цисты покоя. Особый интерес в этом отношении представляют
почвенные виды инфузорий. Один из них, Colpoda steinii, был выбран в
качестве тест-организма. Среди признаков, присущих прочим инфузориям,
данный вид обладает рядом уникальных свойств. Способность образовывать цисты
покоя, устойчивые к высушиванию, и быстрый переход от стадии покоя к активной
жизнедеятельности позволяют получать на основе культуры Colpoda steinii
удобный в использовании сухой препарат-диагностикум с длительным сроком
годности. А отсутствие стадии конъюгации в жизненном цикле и фазы старения в
клональном цикле, специфика процесса деления макронуклеуса, в результате
которого генетический материал, в отличие от большинства инфузорий, точно
распределяется между дочерними вегетативными ядрами, а также тетраплоидность
макронуклеуса на постцистной стадии жизненного цикла определяют относительное
генетическое постоянство лабораторных штаммов данной инфузории и однородность
её культуры, полученной путём индуцированного проращивания цист покоя.
Принцип, лежащий в основе биотестирования как процесса преобразования
материального потока в информационный, представлен в виде схемы, показанной на
рис. 6.
Процесс постановки биологического теста начинается с того, что
исследуемый образец переводят в состояние, удобное для проведения эксперимента,
т. е. получают действующую форму, после чего осуществляют ее контакт с
тест-системой.

Рис. 6. Принципиальная схема биотестирования (по Виноходову, 2007)
На тест-систему действует три группы факторов. Первая группа объединяет
факторы исследуемого образца. К ним, например, относятся токсичные вещества,
которые содержатся в исследуемом объекте. Воздействие факторов этой группы и
подлежит оценке в процессе биотестирования, то есть является целевым. Вторая
группа факторов объединяет различные посторонние воздействия и “шумовые”
помехи. Это могут быть изменения температуры, освещения и прочих условий,
которые сказываются на состоянии тест-системы. Эти факторы не всегда удается
учитывать и устранять, тем не менее, необходимо стремиться к минимизации их
воздействия. К третьей группе факторов относятся разнообразные воздействия,
которые наносят на тест-систему намеренно с целью регулирования её чувствительности.
Эти воздействия играют роль физиологической нагрузки, и их применение может
повысить чувствительность тест-систем на один-два порядка.
В результате действия всех этих факторов тест-объекты и вся тест-система
в целом претерпевают некоторую деформацию, что проявляется в изменении
состояния тест-системы и появлении ряда реакций на различных уровнях её
функционирования. Эти реакции различаются по чувствительности, скорости
проявления, лёгкости наблюдения и другим параметрам. Одну или несколько из этих
реакций выбирают в качестве тест-реакции. Этот термин определяется следующим
образом.
Тест-реакция - это одна из закономерно возникающих ответных реакций
тест-системы на воздействие комплекса внешних факторов, выбранная для анализа
состояния этой тест-системы.
Для корректной постановки и адекватной интерпретации результатов
биологического теста необходимо создавать тест-системы определённого состава и
перед использованием приводить их в строго нормированное начальное состояние.
Говоря о составе тест-системы, в первую очередь необходимо упомянуть её
небиологическую составляющую - среду, в которой распределены тест-объекты. Её
химический состав оказывает заметное влияние на чувствительность организмов к
внешним воздействиям. Одни вещества, такие как белки, антиоксиданты, гумины и
некоторые микроэлементы могут оказывать протекторное действие на клетки в
процессе интоксикации. Другие, например, поверхностно-активные вещества и
некоторые органические растворители даже в незначительных концентрациях,
напротив, способны многократно усиливать воздействие токсичных веществ на живые
объекты. По этой причине Виноходов считает, что тест-системы должны включать
лишь те среды, которые готовят на основе растворов с известным и постоянным
химическим составом.
Правильный подбор тест-организма является не менее важным, чем
использование стандартной питательной среды. В экспериментах Виноходов (2007)
использовал инфузорий четырёх видов: Colpoda steinii, Paramecium
caudatum, Tetrahymena pyriformis и Stylonychia mytilus. Существенные
морфологические и физиологические различия этих организмов обуславливают
разнообразие сфер их использования в биологической практике. И, хотя до
настоящего времени не сделано обоснованного выбора универсального тест-объекта,
идеально пригодного для любых методик биотестирования (более того, существует
мнение, что таковой не может быть найден в принципе), анализ индивидуальных
особенностей перечисленных инфузорий, а также сравнение их чувствительности к
ряду токсичных веществ позволили учёным сформулировать общие требования,
которым должны отвечать инфузории, рассматриваемые в качестве кандидатов на
роль тест-организмов. Они таковы:
идентификация вплоть до вида и штамма (клона);
возможность обеспечения постоянства генетических и физиологических характеристик
штамма (клона);
простота культивирования; высокая чувствительность к основным
поллютантам;
- наличие легко наблюдаемых тест-реакций.
При подборе вида инфузории для каких-либо специфических биологических
тестов могут быть составлены и другие дополнительные требования, но указанные
выше сохраняют своё значение.
Помимо вопроса о составе тест-системы, существенное значение имеет
проблема нормирования её начального состояния, которое зависит от
физиологического состояния тест-объектов, входящих в неё, то есть от фазы роста
культуры и стадии жизненного цикла каждой отдельной инфузории. Эти два
параметра должны подвергаться стандартизации. В экспериментах Виноходов
показал, что чувствительность культуры Colpoda steinii, находящейся на
постцистной стадии развития, к мертиолату почти вдвое выше, чем в фазе
экспоненциального роста. В то же время, по данным ряда авторов,
чувствительность инфузорий Tetrahymena pyriformis, не имеющих в
жизненном цикле стадии цисты покоя, в экспоненциальной фазе роста выше, чем в стационарной.
Таким образом, можно заключить, что оптимальным для постановки биологического
теста состоянием культуры инфузорий, с точки зрения чувствительности ответной
реакции, является постцистная стадия жизненного цикла у цистобразующих видов и
фаза экспоненциального роста у инфузорий, которые не имеют стадии цисты покоя.
Нормирование тест-системы по стадии жизненного цикла, входящих в её
состав инфузорий, может быть осуществлено с использованием разнообразных
методов синхронизации. Как правило, применяют синхронизацию с помощью серии
температурных шоков. Этот метод имеет высокую эффективность, но его
практическое использование затруднено вследствие трудоёмкости и значительных
временных затрат на его проведение. Поэтому, хотя в ряде специальных биологических
тестов шоковую синхронизацию продолжают применять, при проведении масштабного
биотестирования от неё рекомендуют воздерживаться. Существует также иной метод
синхронизации. Он относится к группе механических или селективных методов и
заключается в том, что заранее подготовленные цисты покоя инфузории Colpoda
steinii подвергают индуцированному эксцистированию. Данный способ
достаточно эффективен и весьма прост в использовании, его пригодность в
практическом применении служит весомым аргументом при выборе тест-организма для
постановки биологического теста.
В результате вышеописанных манипуляций получали синхронизированную
культуру инфузорий, но оценку степени её синхронизированности проводили не по
общепринятому методу, который заключается в наблюдении за процессом
единовременного деления клеток культуры, а исходя из степени чувствительности
клеток к токсичным веществам. Полученный индекс синхронизированности
показывает, во сколько раз время гибели 100% клеток инфузорий превышает время
гибели 50% клеток, и может служить характеристикой генетической и
физиологической однородности тест-системы. На постцистной стадии жизненного
цикла культуры Colpoda steinii его величина составляла 1,07. Это означает, что
время, в течение которого погибают все клетки в культуре, превышает время
гибели половины из них на 7%.
Таким образом, проблема нормирования первоначального состояния
тест-систем, используемых в биотестировании, может быть решена с помощью
регулирования двух параметров - фазы роста культуры и стадии жизненного цикла
клеток, входящих в её состав.
В своих исследованиях Виноходов изучал две тест-реакции. Первая из них,
тест-реакция гибели, была подробно исследована на модели инфузории Colpoda
steinii. В качестве тест-критерия для оценки степени проявления этой тест-реакции
можно регистрировать либо количество погибших особей за определённый промежуток
времени, либо время, за которое погибает определённая часть задействованных в
опыте животных. Поскольку при работе с инфузориями Colpoda steinii
используют весьма значительное количество клеток, а также вследствие высокой
степени синхронизированности культуры этой инфузории на постцистной стадии
жизненного цикла, применяли второй из этих тест-критериев.
Была исследована реакция инфузорий на мертиолат, бутанол, глутаровый
альдегид, фенол, о-толуидин, фенилгидразин сульфат, 2,4-Д, гексахлоран,
хлорофос, метафос, этафос, а также на неорганические соли, содержащие катионы
Cd2+, Cu2+, Hg2+, Pb2+ и Zn2+. Во всех изученных случаях зависимость времени
гибели клеток инфузории от концентрации вызывающего эту гибель токсичного
вещества ("концентрация - время") имела характерный вид. Обобщенные
кривые, описывающие этот процесс, показаны на рис. 7.

Рис. 7. Обобщённые кривые "концентрация - время" (по
Виноходову, 2007): 1 - кривая для ЛЭ50; 2 - кривая для ЛЭ100
Сравнение чувствительности к токсичным веществам тест-реакции гибели Colpoda
steinii и высших животных показывает, что инфузории реагируют на
концентрации на один-два порядка ниже, чем теплокровные. Об этом наглядно
свидетельствуют материалы таблицы 3. В последней графе указана концентрация
токсичного вещества в экстракте исследуемого корма при соотношении экстрагента
и навески пробы корма 5:1, полноте экстракции 100% и содержании токсина в пробе
в количестве, приводящем к гибели 50% подопытных теплокровных животных, считая
условно, что животное съедает в сутки количество корма, эквивалентное 10%
своего веса.
Таблица 3. Оценка чувствительности тест-реакции гибели инфузории Colpoda
steinii к токсичным веществам в сравнении с теплокровными животными (по
Виноходову, 2007)

Из материалов таблицы видно, что чувствительность данного тест-объекта к
изученным токсичным веществам выше, чем у теплокровных животных. Таким образом,
тест-реакция гибели инфузории Colpoda steinii может быть использована
для анализа объектов внешней среды в области острой токсичности.
Сопоставление чувствительности тест-реакции хемотаксиса инфузории Paramecium
caudatum со стандартными характеристиками токсичных веществ (таблица 4)
показывает, что данная тест-реакция может быть использована для оценки объектов
окружающей среды при работе с концентрациями некоторых токсичных веществ,
приближающихся к области предельно допустимых концентраций.
В процессе экспериментов обе тест-реакции показали постоянные уровни
чувствительности к изученным токсичным веществам, высокую экспрессность,
информативность и удобство в использовании, что даёт основания к рекомендации
их для практического применения.
Таблица 4. Оценка чувствительности тест-реакции хемотаксиса инфузории Paramecium
caudatum к токсичным веществам в сравнении с гигиеническими параметрами (по
Виноходову, 2007)

Для оценки качества морских вод 5-18‰, сточных вод морского
происхождения, растворенных в морской воде загрязняющих веществ, водных
экстрактов донных осадков предложен метод биотестирования с использованием
инфузорий Euplotes patella, основанный на определении выживаемости и
динамики численности инфузорий в тестируемых средах. Метод предложен Петросяном
и Дятловым (Петросян, Дятлов, 2000).
Е. patella lemani присущи характерные морфологические особенности семейства:
тело плоское, эллиптическое; боковые цирры немногочисленны; поперечные и
фронтовентральные цирры сильно развиты, хорошо заметны; ротовые реснички
выступающие, тянутся более чем на половину длины тела.
По данным Петросяна и Дятлова (Петросян, Дятлов, 2000), допустимый
диапазон солёности в эксперименте, длившемся 60 суток, - 5-18‰. Однако Корлис
(Corlis, 1979) признал представителей рода
Euplotes космополитами. Можно предположить, что этот вид может
адаптироваться к широкому диапазону солёности, в том числе выдерживать полное
опреснение.
Метод биотестирования с использованием инфузорий Е. patella lemani
основан на определении выживаемости тест-объекта в тестируемой среде, а также
сравнении скорости роста культуры в тестируемых и контрольных средах.
Кратковременное биотестирование (до 24 ч) по показателю смертности инфузорий
позволяет определить наличие острого токсического действия тестируемой среды.
Длительное биотестирование (96 ч) по показателям роста культур позволяет
обнаружить хроническое токсическое действие тестируемой среды.
В биотестировании использовали 5-7-дневную культуру инфузорий в
логарифмической фазе роста. Эксперименты проводили при комнатной температуре и
естественном освещении. Лабораторным контролем служили фильтрованная,
стерилизованная и аэрированная морская вода из чистых районов моря. Эта же вода
служила средой культивирования инфузорий. Использование аптечной соли для
приготовления морской воды нежелательно. Подготовку тестируемых проб воды,
сточных вод, донных осадков и растворов загрязняющих веществ проводили в
соответствии с общепринятыми методиками. Длительность экспериментов и
регистрируемые показатели определялись токсичностью среды.
Если тестируемая проба оказывает острое токсическое действие на
тест-объекты в течение 24 ч, то на лабораторной культуре инфузорий выполняют
хронический тест в течение 96 ч. Хронический эксперимент проводят в трехкратной
повторности.
Проведение массовых анализов различных природных сред с использованием
морских инфузорий предполагает не только точное следование правилам содержания
культур, процедуре биотестирования подготовки проб к анализу, но и
периодическую калибровку тест-функций самих культур. Инфузории, представляющие
более или менее однородную лабораторную популяцию и используемые в
экспериментальной работе, нуждаются в постоянной калибровке относительно
стандартных токсических веществ. Для калибровки наиболее часто используют
бихромат калия.
2.
Материалы и методы
2.1
Приготовление сенного настоя
гетеротроф сенной настой сукцессионный
Для получения культуры Protozoa использовалась несколько изменённая методика получения сенного настоя по
Плонскому (Плонский, 1997). По аналогии были приготовлены 3 различных типа
настоя: капустный, сенной и на банановых шкурках. Ниже приводится методика
получения сенного настоя.
В 500 мл воды добавлено 10 г сена и прокипячено в течение 20 минут,
охлаждено до комнатной температуры, профильтровано через фильтровальную бумагу
и доведено до объёма 1000 мл.
Для приготовления настоя использовалось сено различного состава, купленное
в зоомагазине. После образования бактериальной плёнки в настой добавлялась
почва или аквариумная вода. Почва приобреталась в цветочном магазине или
набиралась на улице (склон, расположенный недалеко от ММБИ). Аквариумная вода
была получена из аквариума с рыбами, находящегося в биоцентре кафедры
«Биология».
Чтобы предотвратить высыхание настоя, в ёмкости периодически добавлялась
отстоянная водопроводная вода. В дневное время обеспечивалось хорошее
освещение, температура помещения поддерживалась в диапазоне 19-24 оС.
В качестве подкормки использовали небольшое количество кипячёного молока
(около 30 капель) или сухие дрожжи. Подкормка вводилась аккуратно с помощью
пипетки, после чего содержимое ёмкости перемешивали.
Идентификацию простейших проводили под микроскопом на предметном стекле
или в чашке Петри при увеличении: об. 40, ок. 10. Подсчёт проводили в чашке
Петри при увеличении: ок. 10, об. 20. Объём пробы составлял 4 мл. Инфузории и
другие организмы подсчитывались по 10 полям зрения, затем находили среднеарифметическое
значение.
Для выявления таксономической принадлежности обнаруженных организмов
использовались следующие определители: определитель пресноводных
беспозвоночных Европейской части СССР (планктон и бентос) (под редакцией
Кутиковой Л.А., Старобогатова Я.И.), определитель пресноводных беспозвоночных
России и сопредельных территорий (под редакцией Цалолихина С.Я.).
Приведённые в дипломе фотографии выполнены автором дипломной работы.
Исследования проводились в течение трёх лет, в период с 2009 по 2011 гг.
Всего было получено 8 сенных настоев, 1 настой листьев капусты и 1 корок
банана.
2.2
Подготовка опыта на нефтепродукты
Проба воды, загрязнённая нефтепродуктами (НП), была отобрана у бетонной
стены ОАО «Мурманская судоверфь». В лаборатории морскую воду отделили от
сгустков тяжелых НП. Затем в пробу добавили отстоянную водопроводную воду.
Через 1 сутки в полученном водном экстракте определили содержание НП.
Определение проводилось методом бумажной хроматографии. Диапазон измеряемых
концентраций - от 0,5 до 35 мг/л. Для биотестирования использовали рабочий
раствор исходной концентрации НП, а также разведённый раствор в соотношении 1:2
и 1:3. Опыты проводили в 3-х кратной повторности. В чашках Петри в равных
пропорциях смешивали сенной настой, содержащий 10 экземпляров брюхоресничной
инфузории Oxytricha sp. и водный раствор НП разных концентраций. Объём
пробы составил 10 мл. Просмотр проб проводили через 5 мин., 10 мин., 20 мин.,
30 мин. при увеличении: об.10х, ок.10х, об.40х, ок.10х. Наблюдения сопровождалось
видеозаписью или фотографированием.
Критерием острой и хронической токсичности является статистически
достоверное различие с контролем или снижение численности инфузорий на 50% и
25% по сравнению с контролем в течение 24-х и 96-часовой и более экспозиции
соответственно (Временное методическое руководство…, 2002).
В качестве тест-объекта была использована брюхоресничная инфузория Oxytricha
sp.
В начале опыта определили концентрацию НП в рабочем растворе методом
бумажной хроматографии. Она составила 5,0 мг/л, что превышает норматив для
рыбохозяйственных водоёмов в 100 раз.
Работа выполнена в рамках ГБ НИР «Разработка методов биоиндикации и
биотестирования загрязнения пресноводных водоёмов и их применение в условиях
Кольского полуострова», № темы 4.32/10, №ГР 01201002677.
3.
Результаты и обсуждение
3.1
Получение культуры сенного настоя
Опыт 1.
.11.2009 г. был приготовлен сенной настой, в который после появления
бактериальной плёнки (19.11.2009) добавили 50 г сухой земли, купленной в
цветочном магазине и воду из Семёновского озера (20.11.2009). 24.11.2009 были
замечены признаки заплесневения, 25.11.2009 проведён первый подсчёт организмов
(Colpoda steini). 27.11.2009 замечены Oxytricha sp., 1.12.2009 - коловратки Dissotrocha sp., 7.12.2009 появились представители Amoeba sp., 9.12.2009 - плоские черви рода Stenostomum и 12.12.2009 впервые обнаружены Nematoda sp.
Вывод: анализ полученных результатов позволил установить следующий
сукцессионный ряд организмов:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки → голые амёбы →
ресничные черви → нематоды.
Последовательность появления видов простейших и их количественная
характеристика показаны на гистограмме (рис. 8).
Необходимо отметить, что состояние экосистемы достаточно динамично. Мы
наблюдаем постоянную смену одних организмов другими, причём один и тот же вид
может исчезнуть, появиться и снова пропасть. Численность организмов в сенном
настое уменьшается в обратном порядке установленного ряда.


Рис. 8. Динамика смены первых организмов в сенном настое
Опыт 2.
.11.2009 по схожей методике был приготовлен настой на капустных листьях.
Однако даже при наличии бактериальной плёнки и небольшого плесневого налёта
(что аналогично опыту 1), простейшие не наблюдались, и 25.11.2009 опыт был
прекращён.
Опыт 3.
.12.2009 был приготовлен сенной настой с использованием того же сена, что
и в опыте 1, куда сразу же было добавлено 4 мл из существующего настоя с
большим количеством цист и одним представителем Oxytricha sp. Уже 12.01.2010 в настое наблюдались Colpoda steini и Oxytricha sp., у поверхности и на стенках скопилась слизь.
Вероятно, слизь являлась питательной средой для простейших и бактерий.
13.01.2010 уже обнаружены Stenostomum sp., Nematoda sp. и Dissotrocha sp.
Вывод: сукцессионный ряд сходен с представленным в опыте 1, однако
имеются некоторые расхождения. Так, в опыте не присутствовали представители
рода Amoeba. Организмы сменяют друг друга в
следующем порядке:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки → ресничные черви →
нематоды.
Опыт 4.
.10.2010 по аналогии с опытом 1 был приготовлен сенной настой, куда после
образования бактериальной плёнки добавили сухую землю в количестве около 50 г,
взятую на склоне около ММБИ.
Вывод: сукцессионный ряд представлен следующей последовательностью:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки.
Вероятно, ресничные черви и нематоды не были обнаружены из-за небольших
сроков проведения опыта.
Динамика изменения численности инфузорий представлена на рис. 9.
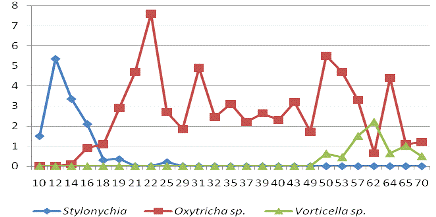

Рис. 9. Среднее чисто особей на одно поле зрения микроскопа
Опыт. 5.
.10.2010 поставлен опыт, аналогичный опыту 4, после образования
бактериальной плёнки добавлена сухая земля (50 г) и вода из аквариума.
Вывод: аналогично опыту 4.
Опыт 6.
.10.2010 по схожей методике был приготовлен настой на банановых корках.
Однако наблюдалась картина, аналогичная опыту 2. Впоследствии опыт был
прекращён.
Опыт 7.
.09.2011 по аналогии с опытом 1 был приготовлен сенной настой, в который
12.09.2011 добавили 20 г сухой почвы. 14.09.2011 в настое появились единичные Colpoda steinii, 30.09.2011 впервые обнаружены Euplotes sp., 13.10.2011 появились коловратки рода Adineta и Amoeba, инфузории рода Paramecium найдены 24.11.2011.
За время проведения опыта проведено 2 подкормки кипячёным молоком:
5.11.2011 и 30.12.2011.
Вывод: в ходе опыта прослеживается следующий сукцессионный ряд:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки → саркодовые →
брюхоресничные черви.
Динамика численности организмов представлена на рис. 10.
В отличие от предыдущих опытов, в данном варианте сенного настоя впервые
появились и долгое время присутствовали в настое раковинные амёбы, солнечники и
гастротрихи. Их численность была невысокой, но достаточно стабильной. Эти
представители беспозвоночных являются типичными почвенными организмами.

Рис. 10. Динамика численности организмов сенного настоя (с добавлением
почвы)
Опыт 8.
.09.2011 был приготовлен аналогичный опыту 7 сенной настой с добавлением
почвы. Colpoda steinii были обнаружены 14.09.2011,
19.09.2011 были обнаружены инфузории Euplotes sp., 5.10.2011 появились Adineta sp., представители Amoeba впервые замечены 13.10.2011,
инфузории Paramecium обнаружены 24.11.2011.
За время проведения опыта было осуществлено 2 подкормки кипячёным
молоком: 5.11.2011 и 30.12.2011.
Вывод: сукцессионный спектр данного опыта аналогичен полученному в опыте
7, но претерпевает некоторые изменения:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → солнечники → коловратки → голые
амёбы → брюхоресничные черви.
Динамика численности представлена на рис. 11.

Рис. 11. Динамика численности организмов в сенном настое (с добавлением
почвы)
Опыт 9.
.09.2011 был приготовлен аналогичный опыту 1 сенной настой, в который
12.09.2011 было добавлено 20 мл воды из аквариума. 14.09.2011 появились первые Colpoda steinii, 4.10.2011 - Euplotes sp., 18.10.2011 - Paramecium caudatum. 25.10.2011 наблюдались Amoeba, 15.12.2011 представители рода Adineta, 11.01.2012 обнаружены ресничные
черви рода Stenostomum, 18.01.2012 - Nematoda sp.
Вывод: сукцессионный ряд вышеописанного настоя можно представить
следующим образом:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → голые амёбы → коловратки →
ресничные черви → нематоды.
Динамика численности представлена на рис. 12.
Как видно из рисунка, в сенном настое с добавлением только воды из
аквариума из саркодовых присутствовали только голые амёбы, а также было
отмечено появление ресничных червей рода Stenostomum и нематод. Черви были обнаружены в
настое через 5 месяцев после начала опыта, и на протяжении всего времени
численность их была стабильной. Нематоды после появления вскоре исчезли.

Рис. 12. Динамика численности организмов в сенном настое (с добавлением
воды из аквариума)
Опыт 10.
.09.2011 был приготовлен сенной настой, аналогичный опыту 9 с добавлением
воды из аквариума. 14.09.2011 обнаружены первые Colpoda steinii, 18.10.2011 одновременно наблюдались
Euplotes sp. и Paramecium caudatum, 13.01.2012 появились Stenostomum sp. и 18.01.2012
- Adineta sp.
Вывод: сукцессионный ряд представлен следующей последовательностью:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки → ресничные черви.
Динамика численности представлена на рис. 13.
Таким образом, результаты наблюдений за сенными настоями с добавлением
аквариумной воды (опыты 9 и 10) были аналогичны.

Рис. 13. Динамика численности организмов в сенном настое (с добавлением
воды из аквариума)
3.2
Краткая характеристика классов встречающихся организмов
3.2.1
Ресничные инфузории (Ciliata)
Ресничные инфузории или Ciliata - класс типа инфузорий
(Ciliophora), типа простейших.


Ресничные инфузории характеризуются, главным образом,
присутствием мерцательных ресничек, покрывающих сплошь всё тело или
сгруппированных на определённых участках тела, затем присутствием ротового и
заднепроходного отверстия (отсутствующих лишь у весьма немногих форм) и, по
крайней мере, двух ядер, различных по величине, строению и функции. Это
микроскопические организмы от 0,03-1 мм в длину, но встречаются формы, видимые
и невооруженным глазом и достигающие 4 мм в длину.
Не менее разнообразна и форма тела ресничных
инфузорий, обладающих, в отличие от других простейших, постоянством очертаний:
наиболее просто организованные (примитивные) виды являются одноосными, будучи
шаровидной, эллипсоидной или продолговато-цилиндрической формы, с
расположенными на противоположных полюсах тела ротовым и заднепроходным
отверстиями; другие вследствие перенесения рта на брюшную сторону являются
билатерально-симметричными, тогда как третьи, и это громадное большинство
ресничных инфузорий, представляются вполне асимметричными.
Реснички, составляющие характерную
принадлежность ресничных инфузорий, суть эктоплазматические образования,
которые дифференцировались при уплотнении протоплазмы на её поверхности как
постоянные непреходящие отростки. Они имеют вид тонких, совершенно однородных и
прозрачных нитей, обыкновенно заострённых к концу, и служат не только для
передвижения ресничных инфузорий, но и для привлечения добычи. Реснички сидят
обыкновенно на маленьких возвышениях, или папиллах, эктоплазмы и расположены
продольными рядами, обусловливающими так называемую полосатость тела. Они сидят
или в неглубоких бороздках, или прямо на поверхности тела, причём у примитивных
форм ряды ресничек идут меридионально и лишь у немногих винтообразно от переднего
к заднему полюсу тела. При перемещении ротового отверстия на брюшную сторону у
более высокоорганизованных инфузорий ресничные полоски на спинной стороне и на
обоих боках сохраняют своё меридиональное расположение, тогда как на брюшной
стороне медианные ресничные полоски доходят лишь до нижнего края ротового
отверстия, или перистомы, и упираются в него, а боковые огибают рот и
сталкиваются под углом между собою в передней части тела. Вообще распределение
ресничек по поверхности тела находится в тесной связи с положением и
устройством рта и его придатков, равно как и с другими наружными и внутренними
органами, обусловливающими степень высоты организации ресничных инфузорий.


Как общее правило можно выставить, что ресничный
покров, одевающий равномерно всё тело примитивных ресничных инфузорий,
претерпевает редукцию у высших форм. Эта редукция выражается в простейшем
случае в исчезновении ресничек на заднем или на переднем полюсе тела, или на
обоих одновременно, так что вместо равномерно одевающего всё тело ресничного
покрова появляется один или несколько поясков или даже один венчик или спираль
ресничек. У других ресничных инфузорий (Hypotricha) при перемещении рта на
брюшную сторону ресничный покров ограничивается лишь брюшной поверхностью тела,
тогда как спинная сторона является совершенно лишённой ресничек или снабжённой
небольшими щетинками. Одновременно и в связи с редукцией ресничного покрова
вместо ресничек появляются более сложные эктоплазматические образования,
которые дифференцируются морфологически: одни - для передвижения, другие - для
привлечения пищи, третьи - для защиты и т. д. К таким эктоплазматическим
образованиям принадлежат: мерцательные пластинки и перепонки, щетинки,
щупальцевидные отростки, крючья и шиповидные отростки.
Громадное большинство ресничных инфузорий является
одиночными, и лишь немногие образуют колонии.
Некоторые ресничные инфузории находятся в постоянном
движении, тогда как другие нередко приостанавливаются и лежат неподвижно более
или менее продолжительное время, а третьи ведут неподвижный образ жизни и
только при наступлении конъюгации или образовании новых колоний временно
переходят в свободноподвижное состояние. Различные движения обусловливаются, главным
образом, пищей; так, например, хищники, находясь в непрестанной погоне за
добычей, плавают безостановочно, тогда как инфузории, питающиеся бактериями,
нередко подолгу лежат неподвижно на одном месте, заглатывая бактерии.
Большинство ресничных инфузорий движется равномерно и одинаково искусно и
быстро вперёд и назад, причем движения сопровождаются большей частью вращением
тела вокруг продольной оси. Hypotricha, снабжённые крючьями и щетинками на
брюшной стороне, очень искусно и быстро бегают при помощи этих приспособлений
по различным предметам, а также и цепляются ими.


По характеру питания инфузории чрезвычайно разнообразны. Большое число их
питается бактериями, некоторые «предпочитают» одноклеточные водоросли.
Существуют немногие растительноядные виды с ограниченным числом пищевых
объектов. Например, одна из пресноводных инфузорий (Nassulla ornata) питается
только нитчатыми сине-зелеными водорослями, которые она скручивает спиралями в
эндоплазме.
Рацион инфузорий-хищников разнообразен. Некоторые охотятся за добычей,
которая порой больше их самих. Например, дидинии (Didinium) питаются
значительно более крупными туфельками (Paramecium). Они поражают добычу
хоботком, а затем засасывают её, раздуваясь при этом, как шар.
Свободноживущие инфузории встречаются как в пресных водах, так и в морях.
Образ жизни их разнообразен. Часть инфузорий - планктонные организмы,
пресноводные и морские. Имеется большое число морских и пресноводных видов
бентических инфузорий. Они ползают по дну, водным растениям и т. п. Очень богат
видами, относящимися к разным отрядам, экологический комплекс инфузорий,
населяющий толщу прибрежного морского песка (псаммофильная фауна). Кроме
свободноплавающих инфузорий в пресной и морской воде существуют многочисленные
виды (отряд кругоресничных), прикрепляющиеся к субстрату несократимыми или
сократимыми стебельками. Нередко эти сидячие инфузории поселяются на подвижных
объектах- моллюсках, насекомых, ракообразных.


В настоящее время доказано, что некоторые мелкие виды инфузорий
(Colpoda), жгутиконосцы и очень мелкие амёбы могут жить в почве как в активном
состоянии, так и в состоянии цист. Исследования показывают, что фауна
простейших играет немаловажную роль в жизни почвы. Сложные взаимоотношения
устанавливаются между фауной простейших и бактериальной почвенной флорой.
Бактерии служат пищей для простейших. Однако простейшие не только поедают их,
но и выделяют некоторые вещества, стимулирующие размножение бактерий (особенно
азотфиксирующих). Показано, что в ряде случаев простейшие способствуют
повышению плодородия, особенно южных хорошо увлажняемых почв (например, в зоне
культуры хлопчатника).

Ресничные инфузории размножаются бесполым и половым
путем: первый заключается в делении и почковании и происходит в
свободноподвижном или покоящемся (инцистированном) состоянии, последний
наступает время от времени и называется конъюгацией.
Кроме конъюгации у инфузорий существуют и другие формы реорганизации
ядерного аппарата, связанные с резорбцией старого и образованием нового
макронуклеуса. Одна из них, наблюдаемая довольно часто и имеющая место у широко
встречающегося вида инфузории туфелек Paramecium aurelia,- автогамия. В
отношении судьбы и поведения ядер она протекает сходно с конъюгацией. Отличие
от конъюгации в том, что при автогамии не происходит временного объединения
двух особей и все процессы протекают в пределах одной особи. Автогамия - это
процесс самооплодотворения, когда сливаются вновь только что разделившихся два
сестринских ядра.

Опыты показывают, что длина периода бесполого размножения у инфузорий при
разных условиях варьирует. Голодание, действие некоторых солей ускоряют
наступление конъюгации, обильное и разнообразное питание задерживает её. Но всё
же для большинства видов периодическая реорганизация ядерного аппарата
необходима. При отсутствии её инфузории впадают в депрессию и погибают.
По-видимому, сложное ядро - макронуклеус требует обновления. Существуют,
однако, некоторые виды инфузорий, которые при оптимальных условиях культуры
могут размножаться бесполым путем неограниченно долгое время. Кроме обновления
макронуклеуса половой процесс у инфузорий, как и у всех организмов, имеет
большое значение как источник повышения наследственной изменчивости, дающий
материал для естественного отбора.
Класс ресничных инфузорий включает 36 семейств, около
160 родов и свыше 450 видов. До последнего времени их подразделяли, по
предложению Штейна, на 4 отряда: Holotricha, Heterotricha, Hypotricha и
Peritricha. На основании новейших исследований инфузории подразделяются на
группы Aspirotricha и Spirotricha.
В наших опытах мы определили следующие роды и виды: Colpoda steinii (рис. 17), Colpoda colpidiopsis, Stylonichia sp. (рис. 18), Euplotes sp.
(рис. 15), Paramecium
caudatum (рис. 16), Paramecium
bursaria, Vorticella sp. (рис. 14), Oxytricha sp.(рис.
19). Некоторые виды инфузорий, встречающихся в культуре сенного настоя, мы не
сочти необходимым определять, так как эти животные встречались единично и
быстро исчезали.
3.2.2
Саркодовые (Sarcodina)
Организмы, имеющие непостоянную форму тела. Обладают изменчивыми по форме
выростами клетки, называемыми псевдоподиями (ложноножками), сохраняющимися в
течение всего или большей части жизненного цикла.
Главная масса саркодовых - обитатели морей. Наряду с морскими видами
имеется немало пресноводных, почвенных и небольшое число паразитов. В пределах
класса различается 3 подкласса: корненожки (Rhizopoda), лучевики (Radiolaria) и
солнечники (Heliozoa).
Корненожки (Rhizopoda)
характеризуются разнообразной формой, подвижными псевдоподиями и отсутствием
дифференцировки цитоплазматического тела на более или менее постоянные зоны. В
подклассе Rhizopoda различают 3 отряда:
амёбы (Amoebina);
раковинные корненожки (Testacea);
фораминиферы (Foraminifera).


Амёбы (Amoebina). Низшие, наиболее просто устроенные корненожки, лишённые
скелета. Большинство амёб - обитатели пресных вод. Некоторые виды живут в море,
а также в почве. Небольшое число - паразиты.
Передвигаясь, амёбы наталкиваются на различные мелкие объекты:
одноклеточные водоросли, клетки бактерий, частицы органического детрита, мелких
простейших и т. п. Если объект достаточно мал, амёба обтекает его со всех
сторон и он вместе с небольшим количеством жидкости оказывается внутри
цитоплазмы простейшего. Таким образом в эндоплазме образуются пищеварительные
вакуоли. Для Amoeba proteus, например, пищей служат другие мелкие
простейшие и одноклеточные водоросли. Описанный способ заглатывания оформленной
пищи при посредстве псевдоподий называется фагоцитозом.
Многие виды свободноживущих амёб (особенно те, которые живут в почве)
способны при наступлении неблагоприятных условий (например, подсыхании)
инцистироваться. При этом амёба втягивает псевдоподии, округляется и выделяет вокруг
себя прочную двойную оболочку белковой природы. В таком неактивном состоянии
амёбы могут сохраняться долгое время (месяцами), а затем вновь переходить к
активной жизни.


Амёбам свойственно бесполое размножение, осуществляемое путём деления
надвое. Этот процесс начинается с митотического деления ядра. Наблюдения
авторов, считающих, что ядра амёб делятся путем амитоза, оказались ошибочными.
На теле появляется легкий перехват, постепенно врезающийся в тело простейшего и
перешнуровывающий её на две дочерние особи. В период деления прием пищи обычно
прекращается.


Раковинные амёбы (Testacea). Представители этого отряда отличаются от
амёб защитной раковиной, одевающей тело. Раковина Testacea обычно имеет вид
округлого или овального мешочка с отверстием (устьем), из которого выдаются
псевдоподии, имеющие у разных видов неодинаковую форму и длину. Раковина у одних
форм представляет тонкий слой плотного органического вещества, у других она
образуется посторонними частицами (песчинки и т. п.), склеенными выделениями
цитоплазмы.
Размножение раковинных амёб совершается, как и у представителей
предыдущего отряда, делением надвое, причём одна из половин выходит наружу
через устье и окружается новой раковиной.
Раковинные корненожки распространены в пресных водах, встречаются
преимущественно среди прибрежной растительности, на дне вблизи берега.
Значительное число видов Testacea живёт в торфяных болотах.


Солнечники (Heliozoa). К
солнечникам относятся всего несколько десятков видов пресноводных и морских
простейших. От радиолярий они отличаются отсутствием центральной капсулы. Тело,
чаще всего шаровидное, распадается на широкую зону эктоплазмы и эндоплазму.
Имеются многочисленные радиально расположенные аксоподии. В эндоплазме одно или
несколько ядер. В эктоплазме пресноводных видов есть редко сокращающиеся
пульсирующие вакуоли. Большинство видов лишено минерального скелета. Некоторые
морские солнечники имеют решётчатый сферический скелет из кремнезема. Бесполое
размножение путём деления. У некоторых видов описан половой процесс.
Виды, встречающиеся в сенных настоях: Amoeba proteus (рис. 20), Difflugia sp. (рис.
21), Actinophrys sol. (рис. 22, рис. 23).
3.2.3
Гастротрихи (Gastrotricha)
Мелкие, едва достигающие 1 -1,5 мм животные, вытянутой в длину,
червеобразной формы. Брюшная сторона тела покрыта ресничками, служащими для
передвижения животного по субстрату; по этому признаку Gastrotricha напоминают
турбеллярий. Встречаются как в морях, так и в пресных водах. В настоящее время
известно около 160 видов Gastrotricha.


Организация брюхоресничных червей относительно проста и свидетельствует о
принадлежности к низшим червям. У пресноводных форм передний конец тела большей
частью слегка обособлен в виде головного участка. Задний конец вытягивается в
два хвостика, или вилку, с открывающимися на них клейкими железами. На конце
тела имеются своеобразные «клеевые железы» грушевидной формы, открывающиеся на
вершине особых трубчатых придатков, по расположению и строению напоминающие
кожные железы прямокишечных турбеллярий (отряд Rhabdocoela).
Небольшая группа Gastrotricha представляет интерес благодаря той связи,
которую она позволяет установить между турбелляриями и некоторыми группами
Nemathelminthes. О близости Gastrotricha и Turbellaria говорят наличие на
некоторых частях тела мерцательного покрова, устройство кожных желез,
протонефридиальный аппарат и строение половой системы.
С другой стороны, тенденция к кутикулизации покровов (утрата части
ресничек), первичная полость тела, раздельнополость некоторых форм и строение
кишечника указывают на сближение брюхоресничных с нематодами.
Все встречающиеся гастротрихи принадлежат к отряду Chaetonotida (рис.
24). Мы не смогли определить этих червей до рода.
3.2.4
Коловратки (Rotatoria)


Коловратки - низшие первичнополостные животные, широко распространённые
во всевозможных водных средах. Форма тела коловраток чрезвычайно разнообразна,
что связано с образом жизни. Длина тела самки обычно не превышает 100-200 мкм,
самцы, если встречаются, заметно мельче самок. Форма тела сильно варьирует. В
редких случаях (Trochosphaera) тело шаровидное, тогда как у громадного
большинства оно продолговатое и разделяется на три участка: передний головной с
мерцательным аппаратом, туловищный, содержащий все внутренности, и задний,
ножной. Нога может и отсутствовать.
Головной отдел мало обособлен от туловищного, а внутренне и вовсе не
отделен от него. Позади венчика на брюшной стороне головы лежит рот, позади рта
- второй венчик ресниц, более мелких. Совокупность обоих венчиков образует
характерный для Rotatoria коловращательный аппарат. Реснички его находятся в
постоянном мерцании, чем и обусловливают движение животного в воде, его
плавание. Одновременно с этим движением ресничек в воде вызывается водоворот,
привлекающий мелкие пищевые частицы ко рту коловратки.


За исходную форму коловращательного аппарата всех Rotatoria в настоящее
время принимают слабо дифференцированный аппарат семейства Notommatidae. У этих
коловраток вместо двух венчиков мощных ресниц на брюшной стороне переднего
полюса тела имеется сплошная мерцательная площадка из мелких ресничек,
окружающая рот. Такой мерцательный аппарат примитивных коловраток напоминает
ресничный покров многих Gastrotricha.
При помощи особых мышц-ретракторов головной отдел может втягиваться
внутрь туловищного и затем снова выворачиваться.
Туловищный отдел, как правило, определяет внешний вид коловратки. В него
легко втягиваются голова и нога в момент опасности. Большинство ползающих форм
имеет удлинённое червеобразное, иногда ложносегментированное туловище. У
плавающих коловраток обычно туловище вздутое, овальное, колоколовидное,
мешковидное или шаровидное. Форма туловища зависит от ригидности покровов.
Коловратки с тонкими покровами могут сильно видоизменяться, особенно при
гибели, у многих видов покровы столь уплотняются, что туловище превращается в
своеобразный панцирь-футляр, куда втягиваются голова и нога. Панцирные
коловратки, напоминающие геометрические фигуры (овальные, многоугольные,
квадратные), нередко на передних и задних краях, реже на спине туловища, имеют
различные более или менее длинные выросты - шипы (чаще у сем. Brachionidae). У немногих коловраток от туловища
отходят выросты-придатки: игловидные (Filinia), листовидно-перистые (Polyarthra) или так называемые
"конечности" или "руки" (Hexarthra), выполняющие локомоторную функцию.


Ножной отдел тела, или нога, у ряда плавающих коловраток отсутствует.
Наоборот, большинство ползающих коловраток и особенно сидячие формы имеют очень
длинную, часто ложночленистую и легко сократимую ногу. Длинная нога характерна
также для колониальных плавающих коловраток, которые концами ног соединяются в
центре шаровидной колонии.
У большинства коловраток нога оканчивается 2 или 4 пальцами, реже - с
венчиком ресничек у плавающих форм или тонким стебельком с прикрепительной
пластинкой у сидячих.
Коловратки раздельнополы и отличаются половым диморфизмом. Самки
встречаются гораздо чаще самцов, и всё предыдущее описание относится именно к
ним. Самцы значительно меньше самок и часто заслуживают названия карликовых.
Сильная редукция многих органов у самцов объясняется кратковременностью их
жизни, которая заканчивается сразу после оплодотворения самки.
Для гетерогенных коловраток характерно чередование двуполого и
партеногенетического размножения. Экологическая специфика различных коловраток
определяет их сроки развития в природе.
Большинство коловраток живет в пресных водах, причём они делятся на
донные и планктонные формы. Донные коловратки большей частью ползают или
плавают у дна, но среди них имеется и целый ряд сидящих видов. Последние
прикрепляются выделением цементных желез ноги к субстрату, причём нога
изменяется в своего рода стебелёк. Большинство сидячих коловраток выделяет
вокруг себя защитную трубку из прозрачного студенистого вещества или склеенных
вместе в правильные комочки испражнений коловратки, которыми трубки обложены
как микроскопическими кирпичами. У типичных планктонных коловраток на теле
вырастают различного рода придатки, увеличивающие поверхность животного. У
некоторых это длинные неподвижные шипы панциря (Notholca), у других - подвижно
сочленённые с телом гребные щетинки (Polyarthra) или это особые, снабженные
мышцами и усаженные щетинками выросты самого тела (Pedalion). Пресноводные
коловратки играют немаловажную роль как пища различных, более крупных
пресноводных организмов. Морских коловраток значительно меньше.


Небольшое количество коловраток приспособилось к жизни на суше, главным
образом во влажном мху, среди лишайников и т. п. Некоторые из них (Callidina)
охотно поселяются в особых мешочках на слоевище печёночных мхов, в которых
долго удерживается вода.
Наземные, а также некоторые водные коловратки способны выдерживать полное
высыхание, не погибая. При этом они впадают в анабиотическое состояние (мнимая
смерть), но после осторожного смачивания оживают. Оживание коловраток было
впервые открыто Левенгуком в 1701 г. и с тех пор многократно исследовалось. При
применении ряда предосторожностей смоченные водой коловратки оживают после
многолетнего высушивания. Мало того, высушенные коловратки способны в течение 4
ч выдерживать понижение температуры до -270°С (в жидком гелии) и в течение 5
мин повышение температуры до + 100°С.
Встреченные в настоях коловратки были классифицированы как Euchlanis sp. (рис. 25), Adineta sp.
(рис. 26), Dicranophorus
sp. (рис. 27), Rotaria sp.(рис.
28).
3.2.5
Турбеллярии (Turbellaria)
Турбеллярии - примитивные первичноротые животные, относящиеся к типу Plathelminthes, в основном представленные морскими
видами, однако довольно разнообразные и в континентальных водах. В состав
класса турбеллярий входят 13 отрядов, 7 из которых в континентальных водах: это
Catenulida, Macrostomida, Lecithoepitheliata, Prolecithophora, Proseriata, Tricladida и Rhabdocoela (=Neorhabdocoela).


Рис. 29. Stenostomum leucops
Форма тела пресноводных турбеллярий червеобразная, как правило,
удлинённо-веретеновидная или каплевидная. Передний конец чаще несколько сужен и
закруглён, иногда заострён или, наоборот, расширен; задний конец обычно сужен и
заострён, иногда вытянут в хвостик. В поперечном сечении тело у небольших форм
округлое или слегка сплющенное, реже четырёхгранное, у крупных представителей (Tricladida) сильно сплющенное, листовидное.
Иногда имеются дополнительные образования, выполняющие прикрепительную или
осязательную функцию и располагающиеся на переднем или заднем конце тела.
Пресноводные турбеллярии не отличаются яркостью и разнообразием окраски. Обычно
они бесцветны или окрашены содержимым кишки в серые или коричневые тона. Реже
встречаются стекловидно прозрачные или сильно пигментированные виды. Пигмент
при этом может располагаться в эпителии или паренхиме. Некоторые турбеллярии
имеют полосатую окраску. Часто они бывают зелёного цвета, который им придают
симбиотические зелёные водоросли, содержащиеся в паренхиме или кишке.
Покровы пресноводных турбеллярий представлены мерцательным ресничным
эпителием, подостланным базальной мембраной. Реснички обеспечивают локомоцию
турбеллярий: мелкие формы плавают с их помощью в толще воды, более крупные
скользят над поверхностью субстрата. С возрастанием размеров тела двигательная
функция всё более переходит от ресничного покрова к мускулатуре кожно-мышечного
мешка. Защитную функцию выполняют находящиеся в эпителии рабдоиды -
палочковидно оформленный секрет эпителиальных желёз. Выбрасываемые на врага или
добычу, рабдоиды разбухают в воде и обволакивают их слизью.


Турбеллярии - хищники, обычно активно разыскивающие добычу, реже
поджидающие её возле своих слизистых сетей, к которым прилипают мелкие
организмы. Спектр питания пресноводных ресничных червей весьма широк - это
разнообразные водоросли (сине-зелёные, зелёные, диатомовые), жгутиконосцы,
саркодовые, инфузории, кишечнополостные (гидры), другие турбеллярии (обычен и
каннибализм), нематоды, коловратки, ракообразные, личинки водных насекомых (в
частности, комаров); имеются свидетельства о нападении крупных турбеллярий на
мальков рыб. Обнаруженная добыча обездвиживается выделяемой слизью, иногда
поражается стилетом и заглатывается целиком или высасывается. Непереваренные
остатки выбрасываются наружу через рот.
В связи с подвижным образом жизни турбеллярий у них наблюдается
определённая "цефализация" - обособление головной лопасти с
сосредоточенными на ней органами чувств.


Половой аппарат у турбеллярий устроен очень сложно и разнообразно и имеет
большое диагностическое значение. Ресничные черви - гермафродиты, у каждой
особи более или менее одновременно развиваются мужские и женские половые
органы. Для большинства характерна протерандрия - более раннее развитие мужской
части полового аппарата.
Турбеллярии обладают способностью к бесполому размножению поперечным
делением. Некоторые (отряд Macrostomida) способны к бесполому размножению посредством повторного поперечного
деления. Деление начинается поперечной перетяжкой тела позади глотки. Ещё до
полного отделения задней половины тела в последней замечается закладка органов,
которые впоследствии регенерируют, и образуется целое животное. Позади
перетяжки от эктодермы обособляется парная кучка клеток - первый зачаток
мозгового ганглия; над ним формируются глаза, а немного позади закладывается
зачаток глотки. Только после этого наступает деление. Так как закладка и
образование органов происходят заранее, то обе особи (получающиеся в результате
деления) сразу же становятся способными питаться, воспринимать раздражения и т.
д. У многих турбеллярий отделение задней половины от передней задерживается
настолько, что каждый из будущих дочерних индивидов успевает начать подготовку
к новому делению. Таким путем у Microstomum и некоторых других
составляются цепочки из 4, 8 и даже 16 особей, расположенных в один ряд - так
называемые стробилы; впоследствии цепочка разрывается на отдельные особи. Такие
цепочки носят характер как бы временных колоний.
У турбеллярий хорошо выражена регенерация - восстановление утраченных
частей тела, это часто наблюдается и при бесполом размножении.
Мы имели некоторые трудности в классификации полученных в настое червей,
так как систематика этого класса разработана слабо. Нам удалось связаться с
японским специалистом в этой области (Masatsugu Yamazaki), который предоставил
ссылки на электронные версии статей, позволяющих определить виды рода Stenostomum (рис. 30). Определение также велось
путём сравнения фотографий и видеоматериалов зарубежных (итальянских,
французских, немецких, японских и испанских) источников и полученных нами в
ходе наблюдений. В результате оба, казалось бы разных внешне, вида, мы
определили как Stenostomum leucops (рис.
29, рис. 31).
3.2.6
Нематоды (Nematoda)


Нематоды - одна из самых распространенных групп беспозвоночных. Общее
число видов нематод превышает 15000, из которых 5000 - свободноживущие. Большая
часть свободноживущих видов входит в состав морских бентосных сообществ, как
правило, доминируя в них. Весьма многочисленны нематоды и в почвенных
биоценозах. В пресных водах отмечено более 100 родов (около 600 видов).
По широте приспособления к условиям обитания мы не имеем среди Metazoa
животных, равных нематодам. В этом отношении их можно сравнить только с
бактериями и одноклеточными организмами. Такая универсальная приспособленность
в значительной мере объясняется развитием у нематод плотной наружной кутикулы,
повышающей их жизненную стойкость. Кроме того, форма тела и характер движений
нематод оказались пригодными для жизни в различных средах - от донных осадков и
почвы до тканей растений и животных.
Несмотря на многообразие сред обитания, которыми овладели нематоды, эти
черви обладают единой довольно постоянной организацией.
Форма тела нематод цилиндрическая или, правильнее, веретеновидная, так
как по направлению к обоим концам тело обыкновенно сужается. Поперечное сечение
тела круглое. На переднем полюсе тела лежит рот, близ заднего конца животного,
на его брюшной стороне - порошица. На брюшной стороне тела находится, кроме
того, выделительное отверстие (недалеко позади рта), а у самки ещё и половое
отверстие. Как правило, никаких придатков, кроме небольших бугорков и щетинок,
тело не имеет.
Нематоды, как правило, раздельнополы и большей частью обладают ясным
внешним половым диморфизмом.
Большинство Nematoda откладывают яйца, но имеются и живородящие формы.
Жизненный цикл у большинства нематод сравнительно прост: нет чередования
поколений. Лишь изредка, например у паразитирующей в лёгких лягушки Rhabdias
bufonis, наблюдается гетерогония.
Нематоды делятся на два подкласса, различия между которыми сводятся,
прежде всего, к неодинаковому строению органов чувств и выделительной системы.
Оборудование лаборатории не позволило нам определить встреченных нематод
до рода (рис. 32).
3.3
Применение инфузорий в биотестировании
3.3.1
Нефтепродукты
Биотестирование водных экстрактов тяжелых НП дало следующую картину.
Просмотр проб в рабочем (неразведённом) растворе показал, что в первые минуты
опыта инфузория резко замедлила скорость движения вплоть до полной остановки.
Затем в течение 5 минут инфузория совершала круговые вращательные движения.
Через 10 минут все инфузории превратились в цисты (рис. 33).



Реакция инфузорий на действие водных экстрактов НП в разведении 1:2 и 1:3
была идентичной. В течение нескольких минут после смешивания культуры Oxytricha
sp. с водными экстрактами НП последние замедляли своё движение. В следующие
10-15 минут они совершали круговые вращательные движения. При этом цитоплазма
инфузорий сокращалась. Размеры перистомального поля значительно уменьшились.
Хорошо просматривается изменение внутренней структуры организмов: уплотнение
макро- и микронуклеуса, нарушение функций сократительной вакуоли и др.
Изменение формы инфузории в течение первых 15 минут после начала опыта
представлено на рис. 34.




Спустя 30 минут во всех пробах инфузории Oxytricha sp.
превратились в цисты. Таким образом, все исследуемые пробы характеризуются
острой токсичностью.
Согласно литературным данным, токсичность нефти и нефтепродуктов зависит
от содержания ароматических и хлорсодержащих углеводородов, присутствия солей
тяжёлых металлов - меди, кадмия, свинца, ртути. Отмечено, что лёгкие НП менее
токсичны, чем тяжёлые (мазут, смазочные материалы). Экстрагируемые из нефтепродуктов
ядовитые вещества оказывают отрицательное влияние на водные организмы (Волкова
и др., 2009). Следует подчеркнуть, что пробы воды были взяты в прибрежной
полосе «Мурманской судоверфи». Результаты проведённого опыта наглядно
демонстрируют ответную реакцию инфузорий на загрязнение, а, следовательно, и
уровень токсичности водной среды.
3.3.2
Эпизодические наблюдения
Параллельно с наблюдениями за развитием сообщества в сенном настое мы
проводили различные опыты, чтобы оценить поведение представителей различных
таксономических групп в различных средах. В опытах применялись только те
животные, которые находились в настое достаточно долго.
Был поставлен ряд опытов на чувствительность инфузорий к различным
средам. В опыте использовалась вода различного происхождения: отстоянная,
водопроводная, родниковая и морская; экстракт нефтепродуктов.
В опыте с водой в чашки Петри добавляли 2мл исследуемой жидкости и 1 мл
сенного настоя, содержащего примерно одинаковое количество инфузории Stylonichia sp.
Отстоянная вода готовилась в лабораторных условиях в течение 1 суток из
водопроводной воды.
Водопроводная вода бралась из крана в лаборатории перед началом опыта.
Родниковая вода была отобрана из родника, находящегося в загородном парке
г. Североморска.
Морская вода была взята на мысе Белокаменном Кольского залива. Солёность
пробы составляла 25‰.
Концентрация нефтепродуктов составляла 0, 125 мг/л, что превышает ПДК в
2,5 раза (ПДК нефтепродуктов - 0, 05 мг/л).
Наблюдения проводились в момент добавления инфузорий, через 5, 15, 30
минут и 1 час.
Отстоянная вода. Через 5 минут после внесения, поведение инфузорий несколько
изменилось, их движения стали более хаотичными (возбужденными), однако уже
через 15 минут большинство из них пришло в норму, и к концу наблюдений в
поведении не наблюдалось отклонений.
Водопроводная вода. Через 5 минут после начала опыта практически все Stylonichia sp. изменили поведение, вращаясь вокруг собственной оси.
Через 15 минут примерно половина из них пришла в норму, а через 30 минут все
инфузории не проявляли каких-либо отклонений от обычного поведения. Однако
через 1 час их количество несколько уменьшилось, животные вели себя вяло,
медленно вращались вокруг оси.
Родниковая вода. В течение всего времени наблюдения каких-либо изменений по
сравнению с контролем замечено не было.
Морская вода. Сразу после внесения было отмечено замедление скорости
движения инфузорий, вращение вокруг собственной оси. Уже через 5 минут
наблюдался лизис животных. Через 15, 30 минут и 1 час ничего живого в пробе
замечено не было.
Экстракт нефтепродуктов. Через 5 минут наблюдения движение инфузорий
несколько замедлились. Спустя 15 минут некоторые Stylonichia двигались по спирали, некоторые
кружились вокруг собственной оси. Однако через 30 минут, после некоторого
уменьшения общего количества, поведение оставшихся инфузорий пришло в норму.
После изучения работы украинских учёных (Петросян, Дятлов, 2000), нами
был поставлен опыт на чувствительность инфузории Euplotes sp. на солёность, так как эти животные могут встречаться
в солоноватой воде. Для этого в 1 л отстоянной водопроводной воды растворили 25
г морской соли, приобретённой в аптеке, получив солёность 25‰. Опыт проводился
с разным разбавлением полученного раствора. Наблюдения проводились сразу после
внесения инфузорий, через 15, 30 минут, 1 час и спустя 1 сутки.
Солёная вода (25‰). Спустя 15 минут после начала, Euplotes были парализованы, не двигались.
Подобная картина наблюдалась через 30 минут и 1 час. Спустя сутки в растворе
обнаружены парализованные и живые, активно передвигающиеся Euplotes, а также некоторое количество цист.
Солёная вода, разбавленная в соотношении 1:1. Изменений в поведении инфузорий
замечено не было. Через сутки в растворе присутствовали активные Euplotes sp.
Солёная вода, разбавленная в соотношении 1:2. Через 15 минут после начала опыта
количество инфузорий визуально уменьшилось, но явных отклонений в поведении
замечено не было. Спустя сутки картина не отличалась, количество Euplotes sp.не изменилось.
Таким образом, мы можем сделать вывод, что даже Euplotes sp., изначально живущие в пресной воде, могут быть
использованы как тест-объекты в солёной воде.
Отдельно хотелось бы выделить опыты, поставленные на чувствительность
различных групп организмов, населяющих сенной настой, к водному раствору
моющего средства «Пемолюкс». Для этого в 100 мл воды добавляли 10 г порошка
«Пемолюкс» и размешивали, получая, таким образом, 10%-ный раствор. В растворе
образовывался осадок, так как в состав моющего средства входит нерастворимый в
воде карбонат кальция и малорастворимый сульфат натрия.
В 4 чашки Петри было внесено по 4 мл сенного настоя, содержащего
ресничных инфузорий Euplotes sp., амёб Amoeba sp., турбеллярий Stenostomum leucops, коловраток Dicranophorus sp, нематод Nematoda sp.
Опыт составлял 4 чашки Петри с различной концентрацией:
) 2 капли 10% раствора Пемолюкса на 4 мл воды;
) 1 капля 10% раствора Пемолюкса на 4 мл воды;
) 1 капля 5% раствора Пемолюкса на 4 мл воды;
) контроль.
В первой чашке Петри большинство Euplotes sp. были обездвижены, у коловраток наблюдались
«конвульсии» и гибель спустя 8-10 минут (рис. 35).
Во второй чашке Петри подобная картина наблюдалась через 1 час после
начала опыта.
В третьей чашке Петри инфузории не изменили поведения, в отношении
коловраток наблюдалась картина, подобная предыдущим опытам.
В первых трёх чашках Петри турбеллярии лизировали практически сразу после
добавления раствора «Пемолюкса».



Спустя сутки с начала опыта были получены следующие результаты:
В первой чашке Петри обнаружены коловратки в состоянии ангидробиоза,
несколько живых амёб, множество цист.
Во второй чашке Петри - активные Euplotes, присутствуют Dicranophorus (ангидробиоз), Amoeba и цисты.
В третьей чашке Петри - инфузории, коловратки, амёбы и цисты
соответственно контрольной пробе.
Таким образом, можно сделать вывод, что чувствительны к данному
токсиканту и пригодны к биотестированию коловратки Dicranophorus sp. и инфузории Euplotes sp.
Заключение
На основе проведённой работы можно сделать вывод о том, что, несмотря на
некоторые различия в условиях, в сенном настое прослеживается закономерность
смены организмов.
Изучаемый микрокосм можно отнести к замкнутой экосистеме, в
незначительной степени обменивающейся с окружающей средой.
По мере минерализации мертвой органики происходит осветление воды и
снижение источников энергии (пищи) для гетеротрофов.
Новаторство данной работы заключается в сравнительно продолжительном
наблюдении сообщества сенного настоя, в результате чего был получен следующий
сукцессионный ряд:
бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки → голые амёбы →
ресничные черви → нематоды.
Организмы, полученные нами в настое, принадлежат к 3 типам и 7 классам:
Protozoa - Rhisopoda, Heliozoa,Ciliata;- Turbellaria; - Nematoda, Rotatoria, Gastrotricha.
Однако видовое разнообразие в каждом варианте опытов отличалось. При
изменении условий существования (количество объектов питания, pH среды, наличие или отсутствие
определённых органических веществ) одни виды инцистируются, а другие
эксцистируются, занимая освободившуюся экологическую нишу. Можно выделить
определённые виды инфузорий, которые остаются в настое долгое время. Виноходов
(2007) отмечает, что в качестве тест-объекта можно использовать те виды
свободноживущих инфузорий, которые имеют в своём жизненном цикле стадию покоя и
способны образовывать цисты покоя. А отсутствие стадии конъюгации в жизненном
цикле и фазы старения в клональном цикле, специфика процесса деления
макронуклеуса, в результате которого генетический материал, в отличие от
большинства инфузорий, точно распределяется между дочерними вегетативными
ядрами, а также тетраплоидность макронуклеуса на постцистной стадии жизненного
цикла определяют относительное генетическое постоянство лабораторных штаммов
данной инфузории и однородность её культуры, полученной путём индуцированного
проращивания цист покоя. В нашем случае этими инфузориями являются Colpoda steinii и Euplotes sp.
В изученной нами литературе по гетеротрофной сукцессии, сукцессионный ряд
начинался с бактерий и заканчивался инфузориями (иногда отмечались коловратки).
Появление ресничных червей в сенном настое не отмечено в более ранних работах.
Фактически появление турбеллярий было отмечено нами впервые. То же самое
касается гастротрих и нематод.
) В разных вариантах опытов идентифицировано (за исключением бактерий и
жгутиконосцев) от 6 до 11 видов простейших организмов, принадлежащих к разным
таксономическим группам: трем типам и шести классам
) Несмотря на некоторые различия в структуре и времени смены одних
организмов другими в разных вариантах опытов, в сенном настое прослеживается
определенная закономерность смены организмов
) Исходя из полученных результатов был выстроен следующий сукцессионный
ряд: бактерии → жгутиконосцы → равноресничные инфузории →
брюхоресничные инфузории → коловратки → голые амёбы →
ресничные черви → нематоды.
Список
литературы
1. Виноходов
Д.О. Научные основы биотестирования с использованием инфузорий: автореф. дис.
докт. биол. наук: 03.02.23 [Текст]. - Санкт-Петербург, 2007. - 30 с.
. Волкова
И.В., Ершова Т.С., Шипулин С.В. Оценка качества воды водоемов
рыбохозяйственного назначения с помощью гидробионтов [Текст]. - М.: Колос,
2009. - 352 с.
. Временное
методическое руководство по нормированию уровней содержания химических веществ
в донных отложениях поверхностных водных объектов (на примере нефти) [Текст].
- М.: РЭФИА, НИА - Природа, 2002.
. Дажо Р.
Основы экологии [Текст]. - М.: Прогресс, 1975 - 415 с.
5. Дедю И.И. Экологический энциклопедический словарь [Текст]. - Кишинев:
Гл. ред. Молдав. Сов. Энциклопедии, 1990 - 408 с.
. Миркин Б.М., Розенберг Г.С., Наумова Л.Г. Словарь понятий и терминов
современной фитоценологии [Текст]. - М.: Наука, 1989. - 223 с.
7. Одум Ю.
Экология [Текст]. В 2т. - М.: Мир, 1986. - Т.1. - 328 с.; Т.2. - 376 с.
. Пахомова
Н.А., Минченок Е.Е. Биоэкологические аспекты изучения сукцессии в микрокосме
сенного настоя [Текст]. Фундаментальные исследования. - 2011. - №5. - С.
201-205.
. Пахомова
Н.А., Минченок Е.Е. Перспективы использования организмов-гетеротрофов сенного
настоя для биотестирования нефтепродуктов [Текст]. Фундаментальные
исследования. - 2012. - №4 (часть 2). С. 396-400.
. Петросян
А.Г., Дятлов С.Е. Инфузории рода Euplotes как тест-организмы в морском
биотестировании // Экология моря. - 2000. - Вып. 50. - С. 105-108.
. Уиттекер Р.
Сообщества и экосистемы [Текст]. - М.: Прогресс, 1980. - 328 с.
12.
Dallinger, W.D. The president's address [Текст]. J. R. Microsc. Soc.,1887, №7, 184-199.
.
Eddy S. Succession of Protozoa in Cultures under Controlled Conditions [Текст]. Transactions of the American
Microscopical Society.- vol. 47, No. 3 (Jul., 1928), pp. 283-319.
.
Gause, G.F. The Struggle for Existence [Текст].- Dover., 1934. - 161 p.
.
Gorden, R.W. et al. Studies of a simple laboratory micro- ecosystem: bacterial
activities in a heterotrophic succession [Текст]. Ecology, 1969, №50, pp. 86-100.
.
Hairston, N.G. et al. The relationship between species diversity and stability:
an experimental approach with protozoa and bacteria [Текст]. Ecology, 1968, №49, pp.1091-1101.
.
Hardin, G. (1960) The competitive exclusion principle [Текст]. Science, 1960, №131, pp.
1292-1297.
.
Lott J. and Mertens Thomas R. Ecologic Succession in Hay Infusions: Search for
a One-Lab Presentation [Текст]. The American Biology
Teacher.- vol. 33, No. 2 (Feb., 1971), pp. 94-97.
.
Luckinbill, L.S. Coexistence in laboratory populations of Paramecium aurelia
and its predator Didinium nasutum [Текст]. Ecology, 1973, №54, pp. 1320-1327.
. Protist Images: Ciliophora [Электронный ресурс]. - Режим доступа: http://protist.i.hosei.ac.jp/PDB/Images/Protista/CiliophoraE.html. - Дата обращения 15.03.2012.
. Stenostomum - Encyclopedia of Life [Электронный ресурс]. - Режим доступа:
http://eol.org/pages/29696/overview. - Дата обращения 22.03.2012.
. Stenostomum leucops - YouTube
[Электронный ресурс] / Masatsugu Yamazaki. - Режим
доступа: http://www.youtube.com/watch?v= PVjOiz1KvvU&feature=plcp. - Дата обращения 14.04.2012.
23.
Tilman, D. Resource competition between planktonic algae: experimental and
theoretical approach [Текст]. Ecology, 1977, №58, pp.
338-348.
.
Turbellari, Forum Natura Mediterraneo [Электронный ресурс]. - Режим доступа:
http://www.naturamediterraneo.com/forum/topic.asp?TOPIC_ID=59981. - Дата обращения 15.03.2012.
.
Wimpergruben-Strudelwurm (Stenostomum leucops) [Электронный ресурс]. - Режим доступа:
http://www.plingfactory.de/Science/Atlas/KennkartenTiere/
Turbellaria/source/Stenostomum%20leucops.html. - Дата обращения 21.05.2012.
.
Woodruff, L.L. Observations on the origin and sequence of the protozoan fauna
of hay infusions [Текст]. J. Exp. Zool., 1912,
№12, pp. 205-264.
.
Vers plats - Naturalia - sciences naturelles [Электронный ресурс]. - Режим доступа:
http://naturalia.over-blog.com/article-28578180.html. - Дата обращения 21.05.2012.