Рост и развитие растений
Приднестровский
государственный университет
им.
Т.Г.Шевченко.
Кафедра
«Ботаники и экологии».
Курсовая
работа
по
физиологии растений на тему:
«
Рост и развитие растений»
Регистрационный № ____ выполнила студентка III
курса
от ___________________ 302 группы
Допущена к защите: ____
естественно-географического
факультета
Защищена на «___» Нечаева Кристина.
«___» ____________2009г. Проверила:
преподаватель: Ионова
Людмила Григорьевна
Тирасполь,2009г.
Оглавление
Введение
I. Клеточное
строение
II.
Дифференциация органов
1. Морфогенез
побега
2. Рост
и развитие листа
3. Рост
и развитие стебля
4. Морфогенез
корня
III.
Влияние экологических факторов на рост и формообразование растений
Заключение
Литература
Введение
Одной из характерных черт многоклеточных
растений является их способность к неорганическому росту. Деревья в возрасте
200-300 лет продолжают увеличиваться в массе и размере, у них появляются новые
ветви и листья. Иными словами, рост многолетнего растения продолжается в
течении всей его жизни. Так как каждое растение состоит из клеток, слагающихся
в органы и ткани, то рост его выражается в изменениях, происходящих в клетках.
Во-первых, в растении увеличивается число клеток путем деления, а во-вторых, их
размеры увеличиваются путем растяжения.
Все изученные процессы в совокупности определяют
прежде всего осуществление основной функции растительного организма-роста,
образование потомства, сохранение видов. Эта функция осуществляется через
процессы роста и развития.
Онтогенез - индивидуальное развитие организма от
зиготы до естественной смерти. В ходе онтогенеза реализуется наследственная
информация организма (генотип) в конкретных условиях окружающей среды, в
результате чего формируется фенотип, т.е. совокупность всех признаков и свойств
данного индивидуального организма.
Рост - процесс необратимого новообразования
структурных элементов сопровождающихся увеличением массы и размером организма,
т.е. рост это количественное изменение .
Рост растения тесно связан с изменениями в самой
цитоплазме растения, приводящими, с одной стороны, к увеличению ее массы, а с
другой - к разнообразным дифференцировкам в самой клетке. Происходит
образование рибосом, хондриосом, пластид и рост оболочки.
Развитие - качественное изменение организма, при
котором одни имеют формы и функции превращаются в другие.[1]
На оба процесса оказывают влияние, как внешние
абиотические факторы окружающей среды, так и внутренние факторы самого
организма.
Одним из важнейших внутриклеточным регулятором
активации генов и развитие того или иного процесса, который связан с ростовым
процессом или переходом растения в следующую фазу развития является
фитогормоны. Изученные фитогормоны делятся на две
большие группы:
ü Стимуляторы роста.
ü Ингибиторы роста.
В свою очередь стимуляторы роста делятся на три
большие группы:
· Ауксины.
· Гиббереллины.
· Цитокинины.
Ауксины. При помощи этих веществ, влияющих на
цитоплазму клеток, в растении увеличивается пластичность клеточной оболочки.
Такова, по-видимому, роль ауксина и, в частности, гетероауксина.
Тургорное давление, которое не отличается
большой величиной у растущих клеток, вызывает процесс растяжения оболочек
клеток и увеличение их размеров. Таким образом, влияя на цитоплазму, ауксин
увеличивает пластичность оболочки, а тургорное давление является тем
механизмом, который обуславливает само растяжение клеток.
Действие ауксина в растении многообразно. Прежде
всего ауксин влияет на растяжение клеток, а также способен вызывать и клеточное
деление. Активная деятельность камбия весной зависит от количества ауксина,
поступающего из распускающихся почек. Ауксин обеспечивает коррелятивное
взаимодействие между органами растения в период роста. Он стимулирует рост
верхушечной, апикальной почки и тормозит рост боковых почек. Это свойство
получило даже название апикального доминирования. Ауксина влияет также на
формообразовательные процессы у растений, вызывая партенокарпию(отсутствие
семян) у плодов, активирует корнеобразование у черенков и взрослых растений.
В растениях гетероауксин образуется из
аминокислоты триптофана через ряд промежуточных продуктов, в частности
индолилпировиноградную кислоту и непосредственного предшественника
ИУК-индолилацетальдегида.
Гиббереллины. За последние несколько лет большое
внимание привлекли к себе физиологически активные вещества, получившие название
гиббереллины. Более ста лет известна болезнь риса- бакане, что в переводе
значит «дурные побеги». Наиболее характерным признаком бакане является
появление высоких тонких растений заметно обгоняющих в росте здоровые растения.
Больные растения имеют большие междоузлия и более тонкие и узкие листья.
Растения кажутся пораженными хлорозом. Колосья у больных растений обычно
меньше, и урожай риса ниже.
Сейчас известно более 40 гиббереллинов. Они
относятся к алициклическим карбоновым кислотам.
Ускоряют фазу растяжения клеток, регулируют
процессы цветения и плодоношения, транспорт происходит как сверху вниз так и
снизу вверх. Характеризуются стремительным ростом, вытягиванием, истончением.
Цитокинины. Производные пурина. Они стимулируют
деление клеток, вызывают заложение и рост стеблевых почек, а также вызывают
вторичное позеленение пожелтевших листьев. Усиливают синтез ДНК, РНК, белков.
Ингибиторы роста делятся на два класса:
· Абсцизовая кислота(АБК).
· Этилен.
АБК это гормон стресса ее количество
увеличивается при высоких температурах воздуха и недостатке воды. Это приводит
к закрыванию устьиц. АБК подавляет образование нуклеиновых кислот и белков.
Этилен. Это газообразный фитогормон, который
тормозит рост, но при этом ускоряет созревание плодов. Этот гормон выделяется с
созревающими органами этого же растения так и на растение растущее рядом.
Этилен ускоряет опадение листьев, цветов, плодов за счет образования
отделительного слоя у черешков.
I. Клеточные основы
роста
«Рост» и «развитие» растений - это разные
понятия.
Само слово «рост» говорит нам о каком-то
увеличении. если мы будем время от времени измерять одно и то же растение, то
обнаружим, что оно в течение своей жизни увеличивается в высоту, молодые листья
его и побеги становятся все больше и больше. Время от времени взвешивая
растение, начиная от его всходов и до плодоношения, можно обнаружить увеличение
его и в весе.
Увеличение растения в высоту, в объеме, в весе и
называют ростом растения. В это же время у растения появляются новые побеги и
новые листья.
Под термином рост у растений подразумевают
несколько процессов:
1. Рост клеток.
. Рост тканей.
3. Рост растительного организма в целом.
1. Рост
клеток.
Эмбриональная фаза. Клетка возникает в
результате деления из другой эмбриональной клетки. Затем она несколько
увеличивается главным образом за счет увеличения веществ протоплазмы, достигает
размеров материнской клетки и снова делится. Таким образом, эмбриональная фаза
делится на два периода. Период между делениями и собственно деление клетки.
Структура клетки в период между делениями (интерфаза) имеет ряд особенностей:
густая цитоплазма с хорошо развитой эндоплазматической сетью, каналы которой
узкие, с малым количеством расширений (цистерн), мелкие вакуоли; большое
количество рибосом, многие из которых свободно располагаются в цитоплазме и не
прикреплены к мембранам эндоплазматической сети; митохондрий много, но они еще
не достигли окончательного размера, с мало развитыми кристами и густым
матриксом. Имеются и промитохондрии и пропластиды деление которых можно
наблюдать. Ядро относительно небольшого размера, с крупными ядрышком. Первичная
клеточная оболочка пронизами плазмодесмами. В период между делениями в клетке
идут интенсивные процессы обмена веществ - активный синтез белка, высокая
интенсивность дыхания, сопровождаемая образованием АТФ. Именно в этот период в
ядре клетки происходит самовоспроизведение ДНК. Если процесс
самовоспроизведения ДНК почему-то приостановлен, деление клетки не происходит.
Таким образом основные синтетические и энергетические процессы в клетке
происходят именно в период между делениями.
Существует несколько гипотез, объясняющих
переход клетки к делению. Наиболее распространена гипотеза, согласно которой в
меристематической клетке должно быть определенное соотношение между размерами
ядра и цитоплазмы. Когда это отношение ниже определенного уровня, ядро как бы
уже может управлять возросшей массой цитоплазмы и клетка переходит к делению.
Перед делением происходят заметные изменения в
энергетическом состоянии клетки. Во время интерфазы клетка характеризуется
очень высоким энергетическим потенциалом. При переходе к митозе благодаря
глубокой структурной перестройке наступает как бы энергетическая разрядка и,
частично, энергия выделяется в виде коротковолного излучения. В период деления
интенсивность процессов обмена, в том числе и дыхания, падает.
Делению клетки предшествует деление ядра. Каждое
ядро дочерней клетки получает ровно такое же число хромосом и такое же
количество ДНК, как и материнская. Во время митоза движение цитоплазмы
прекращается, митохондрии и пластиды распределяются примерно поровну между
дочерними клетками.
На конечной фазе митоза, когда образовались два
ядра, в экваториальной области клетки формируется клеточная стенка. Начало
образования клеточной стенки происходит в телофазе, когда хромосомы отошли к
полюсам клетки, и началась их деспирализация. На границе раздела двух клеток
скапливаются пузырьки, отделяющиеся от аппарата
Гольджи и эндоплазматической сети. Эти пузырьки
сливаются и образуют капельки, впоследствии преобразующиеся в плазмодесмы. В
результате образуется сплошной слой - межклеточная пластинка. Она удлиняется и
смыкается с продольными оболочками клетки. Затем начинается нарастание
межклеточной пластинки в толщину путем присоединения с обеих сторон новых
пузырьков, отщепляющиеся от аппарата Гольджи. Возникает трехслойное
образование, состоящее в основном из пектиновых веществ и гемицеллюлозы. Затем
в него включаются фибриллы. В результате в каждой из дочерних клеток возникает
первичная оболочка, состоящая из рикса. В связи с большой гидротированностью
клеточная оболочка в каждой из клеток первичной оболочки межклеточная пластинка
сохраняется в виде межклеточного вещества. Из мембран аппарата Гольджи формируется
плазмолемма.
Таким образом, в первой фазе роста увеличение
объема клетки происходит за счет деления и возрастания массы протоплазмы.
Одновременно идет формирование структурных компонентов клетки. Следовательно,
рост уже в этой фазе сопровождается формообразовательными процессами.
Образовавшаяся в результате деления клетка вновь увеличивается в объеме и снова
делится. После того как клетка разделится 3-5 раз, она переходит в вторую фазу
роста. Исключение составляют лишь инициальные клетки, которые продолжают
делиться в течении всего периода роста растительного организма.[3]
Фаза дифференциации. На этой фазе процесс
дифференцировки уже проявляется в определенных внешних признаках, т.е. меняются
форма и внешняя структура клетки. Протоплазма почти целиком расходуется на
утолщение клеточной оболочки. Вновь образовавшиеся слои фибрилл целлюлозы
накладываются на старые (аппозиция). При этом ориентация фибрилл целлюлозы в
каждом новом слое другая. В результате образуется вторичная клеточная оболочка.
Клетки теряют способность к делению и растяжению.
Процесс функциональной дифференциации клеток,
или накопление физиологических различий между ними, происходит на всех фазах
роста. Уже между появившимися в период деления дочерними клетками, из которых в
дальнейшем будут образовываться различные ткани, имеются определенные различия,
проявляющиеся в их химическом составе, морфологических особенностях ядер и
органелл.[3]
2. Рост тканей в зависимости от её специфичности
может проходить по какому - либо из типов:
v Апикальный или
верхушечный тип.
Расположены в окончаниях (верхушках) стебля,
растущих побегов и корня и обеспечивает верхушечный тип роста, а также рост
стебля в толщину, обеспечивают меристемы, которые располагаются между флоэмой и
ксилемой.
v Базальный тип.
Базальные меристемы расположены у основания
органа и характерны для листьев, которые растут основанием.
v Интеркалярный или вставочный тип.
Меристемы расположены между двумя закончившими
рост тканями, характерны для стебля (рост междоузлий) и для некоторых листьев.
Этим меристемам соответствует интеркалярный рост.[3]
2. Рост
растительного организма в целом.
Рост как отдельных клеток, так и тканей, органов
и целого организма характеризуется определённой динамикой, которую можно
отобразить S-образной (сигмовидной)
или так называемой большой кривой роста.[4] Она включает следующие фазы:
Lag фаза(1).
Отражает деление и эмбриональный рост клеток и свидетельствует о незначительной
интенсивности данного процесса. По скорости прохождения её можно судить о
степени готовности клетки к видимому росту.
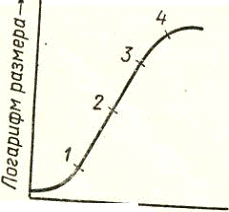

Log фаза(2).
Характеризует процесс растяжения клеток и говорит о наличии потенциальных
возможностях роста организма. К примеру, растянутость её во времени
свидетельствует о слабой способности клеток к росту из-за ограничений со
стороны внутренних или внешних факторов.
§ Фаза замедленного роста(3). Отражает постепенное
прекращение роста и начало дифференцировки. Её быстрый переход к стационарной
фазе свидетельствует о том, что высокой скоростью, которая обычно вызывается
недостатком влаги, высокой температурой или другими факторами.
§ Фаза старения и умирания (стационарная фаза)(4).
Характеризует полное прекращение роста и свидетельствует о качественных
изменениях, не отражающихся на нем.[2]
II.Дифференциация
органов
Термин "дифференцировка" происходит от
латинского differentia
- "разница", "различие". Это понятие было введено для
обозначения процесса приобретения различий между клетками (тканями, органами,
системами органов и т.д.). Предполагается, что есть начальное
недифференцированное состояние, когда наблюдатель не может установить различий
между клетками, а затем видимые различия приобретаются, и клетки становятся
дифференцированными.
Традиционно недифференцированными считают:
) делящиеся клетки эмбриона;
) меристематические клетки (в апикальных
меристемах корня и стебля, камбии, феллогене, интеркалярных меристемах и т.д.);
) клетки, неорганизованно делящиеся в
экспериментальных условиях (суспензионная и каллусная культура in
vitro).
В растительном организме клетки, покинувшие зону
деления, приступают к дифференцировке. Результат этого процесса можно увидеть,
например, при образовании проводящей системы. Сначала возникает прокамбий, из
которого дифференцируются флоэма, ксилема и камбий. Внутри флоэмы
дифференцируются ситовидные элементы и клетки-спутницы, в ксилеме - паренхимные
клетки и трахеиды. Проводящий пучок может быть усилен дифференцирующимися
механическими тканями и т.д. В приведенном примере клетки постепенно
приобретают анатомические различия в связи с выполняемыми функциями,
многообразие клеток растет.
Превращение эмбриональной клетки в
специализированную называют дифференциацией. Из меристематических клеток могут
образовываться различные клетки постоянных тканей: палисадных, паренхимных,
ситовидных трубок, склеренхимных волокон и др.
После прекращения фазы роста растяжением
меристематическая клетка может развиваться в разных направлениях. Одна
превращается в специализированную клетку, тогда как другая вновь делится,
оставаясь эмбриональной, а затем дифференцируется. Дифференциация клетки
(эмбриональной и специализированной) характеризуется определенной генной
моделью, одни гены которой могут быть в активном, другие в блокированном, третьи
в регулируемом состоянии. Тот или иной путь развития клетки определяется особым
набором белков-ферментов; синтез каждого белка, каждого фермента управляется
при посредстве мРНК одним геном ДНК. Образование различных наборов ферментов в
разных типах клеток - по-видимому, следствие дифференциальной активности генов.
В результате генной регуляции отмечается появление или исчезновение
белка-фермента. Генные регуляции вызываются и стимуляторами развития
(фитогормонами, светом и др.). так, укоренения черенков под воздействием
фитогормона ИУК, который, по-видимому, участвует в активации генов, образование
корней происходят из ткани перицикла, в клетках которого гены не блокируются.
Механизмы регуляции на уровне генов и ферментативной регуляции обуславливаются метаболитами:
активация и индукция - субстратом, торможение и репрессия - продуктами
метаболизма.
Дифференциация клеток становиться невозможной
после морфологических изменений в результате крайней степени дифференциации и
потери клеточного ядра. Явление, когда дифференцированная клетка вновь
становиться эмбриональной, т.е. происходит дедифференциация
(ремеристематизация), можно наблюдать у раневого каллуса, образовавшегося из
паренхимных тканей у травянистых и из камбия у древесных растений. Ткань
каллуса - это аморфная масса недифференцированных клеток, потенциально
способных к дифференциации.
Для изучения процессов роста и развития растений
большое значение имеет метод культивирования изолированных клеток и тканей на
исскуственных средах.[5]
Этапу анатомической дифференцировки предшествует
биохимическая дифференцировка. При биохимической дифференцировке видимых
различий между клетками мало, однако клетки не одинаковы по содержанию тех или
иных веществ. Наиболее удобно следить за появлением новых или снижением уровня
старых м-РНК и белков (дифференциальной экспрессией генов). Полученные данные
позволяют зарегистрировать различия между клетками раньше, чем они станут
видимыми на анатомическом уровне. Таким образом, дифференцировка начинается с
процесса изменения активности генома, экспрессии одних генов и с подавления
активности других.
При таком подходе делящиеся клетки меристемы
также придется считать дифференцированными, поскольку для прохождения
клеточного цикла необходима определенная активность генома, которая и будет
отличать эти клетки от других (т.е. у них будет отличие, differentia).
Любопытно, что еще анатомы обратили внимание на неоднородность клеток
меристемы. Можно сказать, что апикальная меристема корня дифференцирована на
каллиптроген (инициали чехлика), дерматоген (инициали эпидеральной ткани),
инициали коры, покоящийся центр и инициали осевого цилиндра. Для каждой из
групп делящихся клеток характерна определенная локализация, направление
веретена делений и тип производных клеток. Исследование меристемы методами
молекулярной генетики показывает, что обнаруженная анатомами дифференцировка
меристемы на зоны совпадает с зонами дифференциальной экспрессией определенных
генов. Более того, саму меристему вцелом можно достаточно четко выделить по
зонам дифференциальной экспрессии. Таким образом, меристема является
биохимически дифференцированной тканью.
По-видимому, дифференциальная экспрессия генов
является фундаментальным проявляением дифференцировки, и, как это ни
парадоксально, недифференцированных клеток вообще не существует. Понятие
"недифференцированный" хорошо работает только там, где в соответствии
с задачами исследования исходные различия между клетками не учитываются (или
нет методов их обнаружить).
Таким образом, дифференцировкой можно назвать
процесс изменения профиля генной активности, приводящий к дальнейшему изменению
функции клеток.
Растения в отличии от животных в течении всей
жизни растут, образуя новые ткани и органы, которые закладываются в
эмбриональных зонах - меристемах, где все клетки делятся. Существование
меристем поддерживается инициальными клетками (инициалями), способными к
делению неопределенно длительное время. Апикальные (верхушечные) меристемы
расположены на концах побегов и корней. Латеральные (боковые) меристемы образуют
слои клеток вдоль каждого побега и корня (у двудольных растений). К ним
относятся первичные латеральные меристемы - прокамбий и перицикл и вторичные -
камбий и феллоген. В основании молодых междоузлий и листьев функционируют
интеркалярные (вставочные) меристемы. При повреждении у растений активируются
меристемы, за счет которых регенерируют ткани, органы и целые организмы. После
заложения тканей и органов в меристемах их рост происходит благодаря делению и
последующему растяжению клеток. Форма и размеры органов и растения в целом
определяются количеством и локализацией делящихся и растягивающихся клеток,
интенсивностью и ориентацией их деления и роста. В процессе роста тканей и
органов клетки, окруженные клеточными стенками и связанные плазмодесмами, не перемещаются
относительно друг друга(согласованный рост). Эта особенность также отличает
рост растительных организмов от животных, у которых рост зародыша
сопровождается перемещением клеток и целых клеточных слоев.
Таким образом, морфогенез, т.е. формообразование
у растений, включает в себя процессы заложения, роста и развития клеток
(цитогенез), тканей (органогенез), которые генетически запрограммированы и
скоординированы между собой. Апикальные меристемы побега и корня представляют
собой не только образовательные ткани, из которых формируется весь растительный
организм, но и главные координирующие (доминирующие) центры, влияющие на
морфогенетические процессы в целом растении.
Морфогенез побега.
Основные части побега - стебель, листья, почки,
цветки и др. - закладываются в апикальной меристеме побега, которая является
производной эмбриональной ткани дистального конца зародыша.
Апекс побега. Апекс (конус нарастания, точка
роста) вегетирующего побега семенного растения состоит из меристематических
клеток, которые по своему размеру, частоте и направленности делений, по
особенностям метаболизма могут быть разделены на несколько зон, прежде всего на
тунику и корпус. Туника, или мантия, - это один, два или более слоев клеток,
покрывающих апекс снаружи. Клетки туники делятся преимущественно антиклинально
( т.е. плоскость деления перпендикулярна поверхности апекса). Из наружного слоя
туники образуется эпидермис. Все остальные клетки, лежащие под туникой, входят
в состав корпуса, в котором при более дробной анатомо-физиологическом
разделении апикальной меристемы побега на зоны выделяют центральную,
периферическую и стержневую меристемы. Дистальная группа клеток в тунике и
центральной (аксиальная) зона выполняют функцию инициалей. Клетки этих участков
апекса относительно крупные и делятся сравнительно редко. Периферическая зона
(инициальное кольцо) состоит из мелких меристематических клеток, которые
интенсивно делятся. В них число рибосом выше чем у инициальных клеток. Клетки
этой зоны формируют примордии (зачатки) боковых органов побега - листьев и
почек. Граница между туникой и корпусом в этой зоне исчезает. Клетки все зон
апекса имеют крупные ядра, густую цитоплазму и не содержит вакуолей.
Стержневая (колончатая) зона состоит из
вакуолизированных клеток с относительно низким содержанием РНК. Клетки этой
зоны делятся главным образом антиклинально и дают начало продольным рядам
клеток первичной коры и сердцевины стебля. Границы между описанными зонами в
апексе побега весьма условны и далеко не всегда различимы.
Конус нарастания побега, обладая высокой
способностью к саморазвитию, тем не менее, нуждается в притоке не только
питательных веществ, но и фитогормонов. Изолированные апексы с двумя-тремя
листовыми примордиями нормально развиваются лишь в том случаи, если в питательной
инкубационной среде присутствуют цитокинин, а в ряде случаев - и ауксине.
Рост и развитие листа.
Формирующийся лист проходит четыре фазы: 1)
образование примордия; 2) формирование оси листа; 3) закладка пластинки листа
за счет боковой меристемы; 4) рост пластинки растяжением.
Каждый листовой примордий образуется в виде
бугорка в периферической меристеме апекса побега благодаря локальным
периклинальным делением клеток (плоскость деления параллельна поверхности
апекса). У многих видов периклинальные деления в зоне закладки примордия
происходит и в тунике. Примордий пазушной почки появляется несколько позже. В
нем затем формируется апикальная меристема, гомологичная апексу главного
побега.
Период времени между заложением двух листовых
зачатков называется пластохроном. Продолжительность его у разных видов и даже у
одного и того же вида при разных условиях сильно варьирует: от нескольких часов
до нескольких суток. Примордии листьев образуются на апексе в строго заданной
последовательности, предопределяя расположение листьев на зрелом побеге, или
филлотаксис. У растений распространен спиральный филлотаксис. Замечено, что на
апексах с многочисленными примордиями угол между ними близок к значению 137,5о
. при таком угле в идеальном случае ни один лист на стебле не располагается
точно под другим, что обеспечивает их минимальную затененность. По теории В.
Гофмейстера, такое листорасположение достигается путем возникновения новых
листовых зачатков в промежутках между уже существующими примордиями ( «теория
доступного пространства»).
Согласно «теории отталкивания», предложенной
Ю.Шоуте, когда детерминируется центр листового примордия, в нем продуцируется
специфические вещества, ингибирующие образование новых центров в
непосредственной близости от заложившегося. Соответственно новый примордий
развивается вне ингибиторных полей соседей.
Эти гипотезы довольно хорошо согласуются друг с
другом, так как «доступное пространство» может быть обусловлено не только
поверхностной зоной между примыкающими примордиями, но и зоной, от их
ингибирующего влияния. В этом месте и закладывается новая примордия. Возникшие
зачатки листьев оказывают влияние на нижележащие ткани, индуцируя
дифференциацию проводящих пучков. Это действие обусловлено делением ауксина,
который синтезируется в формирующихся примордиях.
Верхушечные клетки конуса листового примордия
делятся особенно интенсивно, превращая бугорок в пальцеобразный выступ. Этот
выступ состоит в основном из клеток будущей средней жилки и черешка листа. На
краях зоны средней жилки начинает функционировать маргинальная (краевая)
меристема, дающая начало листовой пластинке. Одновременно с этим прекращается
верхушечный рост листа.
Инициальные клетки маргинальной меристемы и
клетки самой этой меристемы делятся главным образом антиклинально, что приводит
к увеличению листовой пластинки, а не ее толщины. Поверхностные инициальные
маргинальные клетки образуют эпидермис, а субмаргинальные инициальные клетки -
внутренние ткани листа.
После 8-9 циклов делений клетки маргинальной
меристемы переходят к растяжению. Эпидермальные клетки кончают делиться
первыми, но продолжают расти растяжением. Клетки губчатой паренхимы прекращают
делиться растут раньше других тканей. Поэтому продолжающееся деление и
растяжение эпидермиса приводят к тому, что губчатые клетки отодвигаются друг от
друга, образуя большие межклетники. Палисадные клетки делятся и растут со
скоростью, близкой к росту эпидермиса. Этот процесс останавливается несколько
раньше окончания растяжения эпидермиса. Поэтому и палисадные клетки не сколько
отрываются друг от друга, образуя небольшие межклетники.
Особенность роста листа однодольных растений
состоит в том, что деления в возникшем на одной стороне апекса листовом бугорке
распространяются в обе стороны и охватывают всю окружность стебля. Возникающий
серповидный (у злаков) или кольцевой (у осок) меристематический валик дает
начало листу, растущему вверх. До формирования валика бугорок растет, как и у
двудольных, верхушкой. Листовая пластинка удлиняется путем интеркалярного
роста, который более продолжителен у основания пластинки.
На рост листьев большое влияние оказывают
периодичность, качество и интенсивность света. Свет сине-фиолетовой части
спектра тормозит рост междоузлий и способствует росту листьев ( у двудольных).
Интенсивное освещение способствует развитию палисадной ткани.
Гормональная регуляция роста листа изучена
недостаточно. Показано, что цитокинин и ауксин необходимы для формирования и
развития примордиев и тканей листа, ауксин принимает участие в образовании
жилок, гиббереллин способствует более интенсивному росту листовой пластинки в
длину.
Рост листьев ограничен в отличии от пазушных
почек, у которых заложенные апексы будут функционировать длительное время, если
эти почки дадут боковые побеги.
Рост и развитие стебля.
Стержневая меристема апекса и прокамбий,
образование которого индуцируется растущими примордиями листьев, формируют
основные ткани стебля. Выходя из меристематической хоны, клетки начинают
растягиваться, что приводит к быстрому удлинению побега. Зона роста растяжением
у побегов в отличие от корней достигает больших размеров (нескольких
сантиметров). Рост растяжением стебля активируется гиббереллинами ,
стимулирующими переход большого количества клеток к этому типу роста, и
ауксином, который непосредственно индуцирует удлинение клеток. Гиббереллины
главным образом из листьев, и это позволяет регулировать скорость и
продолжительность роста верхних междоузлий.
Стебель у двудольных утолщается за счет
деятельности камбия, для активации которого необходима ИУК, поступающая из
верхушки побега, а также благодаря пробковому камбию - феллогену, который
образуется из различных слоев наружных клеток стебля.
Рост пазушных почек (ветвление) находится под
двойным контролем: их рост и развитие ингибируют верхушечная почка побега и
листья, в пазухах которых они находятся.
Морфогенез корня.
Апекс корня. У высших растений апикальная
меристема корня имеет сравнительно простое строение. Это зона длинной 1-2 мм. В
ней не образуются боковые органы, как в верхушечной меристеме побега. Корнева
меристема формирует ткани корня и корневой чехлик, защищающий корень при его
продвижении в почве.
Кроме активно делящихся клеток, в меристеме
корня находится группа клеток, расположенных между корневыми чехликом и
активной меристематической зоной, для которых характерны низкий уровень синтеза
ДНК и очень редкие деления клеток. Эта группа клеток получила название
«покоящегося центра». Предполагается что « покоящийся центр» является
промеристемой активной апикальной меристемы корней, восстанавливающей
численность быстро делящихся специализированных инициальных клеток
При их естественном изнашивании или повреждении.
В этом смысле функции клеток «покоящегося центра» сходны с аналогичной ролью
центральной зоны («зоны ожидания») апекса побега.
Одна группа инициальных клеток локализована на
дистальном конце апекса и продуцирует клетки ризодермы и корневого чехлика.
Другая инициаль связана с репродуцированием клеток первичной коры. Третья
ответственная за поддержание меристематической активности клеток,
дифференцирующихся затем в различные клетки и ткани проводящего пучка.
Образование рядов специализированных клеток в корне можно проследить
непосредственно от их клеток -инициалей. Таким образом, апикальная меристема
корня, как и апекс побега, продолжают ту ткане-и органообразующую деятельность,
которая началась еще при формировании зародыша.
Изолированные «покоящиеся центры» при
культировании на питательной среде нуждаются в присутствии ИУК и циткинина.
Изолированные кончики корней однодольных растут при добавлении ауксина, а у многих
двудольных кончики корней развиваются и без экзогенных фитогормонов. Очевидно,
в последнем случае ИУК синтезируется базальной частью отрезка корня[1].
растение организм развитие
III. Влияние
экологических факторов на рост и формообразование растений
На рост растений оказывают влияние многие
факторы внешней среды. Прежде всего это физические факторы: свет(его
интенсивность, качество, продолжительность и периодичность),
температура(величина и периодичность), сила тяжести, газовый состав, магнитное
поле, влажность, питательные вещества(минеральные и органические) и
механические воздействия(например ветер). Кроме того, находясь в составе
растительных сообществ, растение испытывает влияние продуктов жизнедеятельности
других растении(аллелопатия), а также физиологически активных веществ
микроорганизмов(антибиотиков, ростовых веществ).[1]
Свет .
Рост растений может происходить на свету и в
темноте. Зеленые органы высших растений в темноте приобретают ряд
морфологических особенностей, отличающих их от растений , выращиваемых на
свету. Такие растения называются этиолированными. У двудольных при росте в
темноте сильно вытягиваются междоузлия, а листовые пластинки недоразвиты. У
этиолированных растений, как правило, недоразвиты механические ткани и устьица.
Растения лишены хлорофилла и имеют бледно-желтый цвет из-за присутствия
каротиноидов. Явление этиоляции не связано с отсутствием хлорофилла или
недостатком питания: этиоляция отмечена у многих грибов и у проростков хвойных,
способных зеленеть в темноте. Для устранения этиоляции достаточно
кратковременного освещения растений один раз в сутки, что не влияет на
образование хлорофилла. Предотвращение этиоляции регулируется фитохромом -
флавопротеин.
При освещении этиолированных растений торможение
роста растяжением наступает через несколько минут, остальные эффекты
света(развертывание первого листа, закладка следующих листьев, рост волосков и
т.д.) проявлется лишь через несколько часов.
Температура.
Как и другие процессы, рост растений зависит от
температуры. В пределах от 0 до 35оС влияние температуры подчиняется правилу
Вант-Гоффа, но свыше 35-40оС скорость роста снижается. В зависимости от
приспособленности к действию температур различают растения теплолюбивые с
минимальными точками для роста выше 10оС и оптимальными при 30-40оС и
холодостойкие с минимальными точками от 0-5оС и оптимальными при 25-31оС.
Максимальные температуры для большинства растений находятся в пределах 35-45оС.
Оптимальной называют температуру, при которой рост осуществляется наиболее
быстро. Но высокая скорость роста не всегда благоприятна для формирования
растения. При «выгонке» растений умеренного пояса при высоких температурах
стебель становится слабым, а листья тонкими. В поле такие растения погибают.
Для роста растений необходимо присутствие
кислорода. Однако кратковременное снижение наполовину его содержание
значительно не сказывается на росте растений. Даже в условиях длительного
недостатка кислорода в зоне корней при затоплении рост продолжается, хотя и с
более низкой скоростью. Это связано, как уже отмечалось, с включением
адаптационных механизмов, позволяющих использовать О2 нитратов, воздухоносных
тканей и т.д.
Избыток СО2 в атмосферном воздухе приводит к
увеличению растяжимости клеточных стенок и кратковременному усилению роста
тканей(эффект «кислого роста»). Выяснено, что влияние углекислого газа на рост
основано на способности снижать рН клеточных стенок и таким образом
индуцировать рост клеток. Эффект углекислого газе не зависит от присутствия
кислорода. Именно с эффектом «кислого роста» наряду с затенением может быть
связано чрезмерное удлинение нижних междоузлий злаков в загущенных посевах и
вследствие этого полегание растений.[1]
Водный режим.
Процесс роста клеток растений растяжением
осуществляется путем вакуолизации при поступлении в клетки воды. Поэтому
недостаточное снабжение клеток водой задерживает рост. Корни способны расти
только в достаточно влажной почве, почти насыщенной водяным паром, с
осмотическим давлением почвенного раствора не более 1-1,5 МПа. Надземные части
растений всегда находятся в более сухом воздухе с влажностью 50-70%. От потери
воды ткани неземных органов защищены кутикулярно-эпидермальным слоем. Поэтому в
мезофилле листьев упругость водяного пара обычно не бывает ниже 98-99%
относительной влажности. При длительном стрессе, что приводит к укорочению
стебля и корня, к уменьшению размеров листьев, к их мелкоклеточности и т.д.
Недостаток воды до начала и в период стеблевания злаков(осуществляемого за счет
роста растяжением) особенно резко снижает урожай.
Минеральное питание.
На росте растений благоприятно сказывается
высокое содержание в почве минеральных элементов, особенно азота. Однако
способствующие быстрому росту высокие концентрации азота задерживают процессы
дифференцировки, в частности закладку цветков. Высокий минеральный фон приводит
к разрастанию вегетативных органов и необходим при наращивании зеленой массы
кормовых растений. Но чрезмерное удобрение снижает урожай плодов и зерна.[1]
Заключение
Как уже отмечалось, в основе роста растений
лежит процесс новообразования элементов структуры (органы, такни, клетки и
отдельные их части). Это процесс осуществляется непрерывно на протяжении всей
жизни растения. Лишь у некоторых видов происходит довольно медленно или с
возрастом растения затухает. Неограниченность роста растений обуславливается
наличием у них меристематических зон, сохраняющих активность вплоть до
завершения онтогенеза. Апикальные меристематические зоны обеспечивают
образование побегов, листьев, цветков, молодых участков корня, рост осевых
органов в длину, а латеральные - формирование камбия, феллогена и нарастание
осевых органов в ширину.
Новообразование элементов структуры происходит
поэтапно. Причем по мере появления тканей и органов между ними, как уже
отмечалось, устанавливаются определённые взаимосвязи, выражающиеся в
закономерной зависимости роста одних тканей и органов от роста других.
На рост растения оказывают существенное влияние
внешние факторы. Они, как известно, претерпевают периодические изменения и в
отдельные моменты становятся неблагоприятными для растения. В результате
длительной эволюции растения приспособились переживать такие периоды путем
перехода в состояние покоя: у них резко снижается активность физиологических
процессов и они становятся практически неуязвимыми для вредных воздействий.
Литература
1. Полевой
В.В. «Физиология растений». М.: Высшая школа. 1989.
2. Э.Ф.
Шабельская. « Физиология растений». Минск.: Высшая школа.1987.
. Н.И.Якушкина.
«Физиология растений». М.: Просвещение.1980.
. П.А.
Генкель. «Физиология растений». М.: Просвещение. 1975.
. С.И.
Лебедев «Физиология растений». М.: ВО «Агропромиздат» 1988.