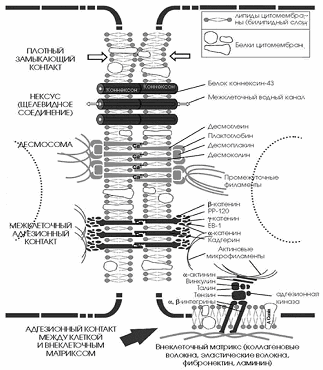
Классификация и ультраструктурное строение межклеточных соединений
Министерство
здравоохранения и социального развития РФ
Волгоградский
государственный медицинский университет
Кафедра
гистологии, эмбриологии, цитологии
Зав. каф.
д.м.н., профессор М. Ю. Капитонова
Учебно-исследовательская
работа студента
«Классификация
и ультраструктурное строение межклеточных соединений»
Выполнила
студентка I
курса 4 группы
медико-биологического факультета
Магомедова М.Н.
Проверил: асс. Загребин В. Л.
Волгоград
2011
Содержание
Введение
. Рыхлые, или простые соединения
. Сцепляющие контакты
.1 Адгезивный поясок
.2 Десмосома
.3 Полудесмосома
.4 Фокальный контакт
. Коммуникационные соединения
.1 Нексус
.2 Синапсы
. Запирающие соединения
.1 Плотный контакт
. Септированные соединения
.1 Плазмодесмы
Заключение
Литература
Ссылки
Классификация
контактов
.
Контакты простого типа - простые межклеточные соединения, интердигитации
(пальцевидные).
.
Контакты сцепляющего типа - десмосомы, адгезивные пояски.
.
Контакты запирающего типа - плотные соединения.
.
Контакты коммуникационного типа - нексусы, синапсы
Введение
Плазмалемма многоклеточных животных организмов
принимает активное участие в образовании специальных структур - межклеточных
контактов, или соединений (junctiones
intercellulares),
обеспечивающих межклеточные взаимодействия. Благодаря межклеточным
контактам осуществляется структурная и функциональная связь клеток в тканевых
системах и обеспечивается интегративная целостность органов. В зависимости от
выполняемой функции и ультраструктурной организации межклеточные контакты
подразделяются на несколько типов. В настоящей главе рассматриваются основные
типы контактов - простые и сложные.
Простые контакты
- между плазматическими мембранами соседних клеток имеется щель шириной 10-20
нм, заполненная гликокалликсом, специализированных структур на мембранах нет.
Сложные межклеточные соединения
представляют собой небольшие парные специализированные участки плазматических
мембран двух соседних клеток. Они подразделяются на запирающие(изолирующие),
сцепляющие (заякоривающие), обусловливающие механическое сцепление и
соединение клеток, и коммуникационные (объединяющие) контакты,
обеспечивающие химическую (метаболическую, ионную) и электрическую связь между
клетками. К запирающим относится плотный контакт.
К сцепляющим соединениям относятся
адгезивный (сцепляющий) поясок, десмосомы, полудесмосомы и фокальный контакт.
Общим для этой группы соединений является то, что к участкам плазматических
мембран со стороны цитоплазмы подходят фибриллярные элементы цитоскелета,
которые как бы заякориваются на их поверхности.
Коммуникационные соединения
в клетках животных представлены так называемыми щелевыми контактами и
синапсами. Особыми формами межклеточных контактов являются септированные,
к которым относятся плазмодесмы растительных клеток.
1.
Рыхлые или простые контакты
Простой контакт (junctiones
intercellulares
simplex) -
соединение клеток за счет пальцевидных впячиваний и выпячиваний
цитоплазматических мембран соседних клеток. Специфических структур, формирующих
контакт, нет.
Простые контакты занимают наиболее обширные
участки соприкасающихся клеток. Расстояние между билипидными мембранами
соседних клеток составляет 15-20 нм, а связь между клетками осуществляется за
счет взаимодействия макромолекул соприкасающихся гликокаликсов. Гликопротеиды
соседних клеток при образовании простого контакта «узнают» клетки одного типа.
Наличие этих белков-рецепторов (кадгерины, интегрины и др.) характерно для
определенных тканей. Они реагируют только с соответствующими им клетками.
Например, Е-кадгерины участвуют в образовании контактов только между
эпителиальными клетками, обеспечивая их соединение практически по всей
поверхности контактирующих клеток.
Посредством простых контактов осуществляется
слабая механическая связь - адгезия, не препятствующая транспорту веществ в
межклеточных пространствах. Интегрины и обеспечивают адгезию клеток к
компонентам внеклеточного матрикса и иногда к другим клеткам, и представляют
собой поверхностные гетеродимерные белки. Утрата некоторых интегринов (при раке
молочной железы, раке предстательной железы, раке толстой кишки) или их избыток
(при меланоме, плоскоклеточном раке полости рта, носоглотки, гортани) сопряжены
с высокой степенью злокачественности опухоли. Взаимодействие интегринов с
белками внеклеточного матрикса в некоторых случаях препятствует апоптозу.
Клеточно-матриксные контакты, образованные с
помощью интегринов хорошо изучены в гладкомышечных клетках и в местах
прикрепления культивируемых фибробластов к внеклеточному матриксу.
Разновидностью простого контакта является
контакт "типа замка", когда плазмолеммы соседних клеток вместе с
участком цитоплазмы как бы впячивается друг в друга (интердигитация),
чем достигается большая поверхность соприкосновения и более прочная
механическая связь.
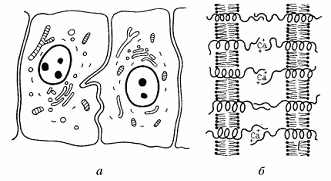
Рис. 1. Схема
простого межклеточного соединения
а - простое
соединение, без участия специальных структур; б - трансмембранные гликопротеиды
определяют связывание двух соседних клеток
2.
Сцепляющие (заякоривающие) контакты
Они соединяют не только плазматические мембраны
соседних клеток, но и связываются с фибриллярными элементами цитоскелета. Для
этого рода соединений характерным является наличие двух типов белков. Один из
них - это трансмембранные линкерные (связующие) белки, которые участвуют или в
собственно межклеточном соединении или в соединении плазмолеммы с компонентами
внеклеточного матрикса (базальная мембрана эпителиев, внеклеточные структурные
белки соединительной ткани). Второй - внутриклеточные белки, соединяющие или
заякоревающие за мембранные элементы такого контакта цитоплазматические
фибриллы цитоскелета.
К заякоревающим соединениям относятся
межклеточные сцепляющие точечные контакты, адгезивный поясок, фокальные
контакты или бляшки сцепления - все эти контакты связываются внутри клеток
с актиновыми микрофиламентами.
Другая группа заякоревающих межклеточных
соединений - десмосомы и полудесмосомы - связываются с другими
элементами цитоскелета, а именно с промежуточными филаментами.
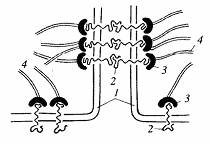
Рис. 2. Схема
строения заякоривающих адгезивных соединений
1 - плазматическая мембрана; 2 - трансмембранные
линкерные гликопротеиды; 3 - внутриклеточные белки сцепления; 4 - элементы
цитоскелета
2.1
Адгезионный поясок (zonula
adherens)
Это парное образование в виде ленты,
опоясывающей апикальную часть клетки однослойных эпителиев.
Он опоясывает весь периметр эпителиальной
клетки, подобно тому как это происходит в случае плотного соединения. Чаще
всего такой поясок или лента лежит ниже плотного соединения. В этом месте
плазматические мембраны не сближены, а даже несколько раздвинуты на расстояние
25-30 нм, и между ними видна зона повышенной плотности. Это ничто иное как
места взаимодействия трансмембранных гликопротеидов, которые специфически
сцепляются друг с другом и обеспечивают механическое соединение мембран двух
соседних клеток. Эти линкерные белки относятся к Е-кадгеринам - белкам,
обеспечивающим специфическое узнавание клетками однородных мембран.
Разрушение этого слоя гликопротеидов приводит к
обособлению отдельных клеток и разрушению эпителиального пласта.
С цитоплазматической стороны около мембраны
видно скопление плотного вещества, к которому примыкает слой тонких (6-7 нм)
филаментов, лежащих вдоль плазматической мембраны в виде пучка, идущего по
всему периметру клетки.
Тонкие филаменты относятся к актиновым
фибриллам, они связываются с плазматической мембраной посредством белка
катенина, образующего плотный около мембранный слой.
Функциональное значение такого ленточного
соединения заключается на только в механическом сцеплении клеток друг с другом:
при сокращении актиновых филаментов в ленте может изменяться форма клетки.
Считается, что кооперативное сокращение
актиновых фибрилл во всех клетках эпителиального пласта может вызвать изменение
его геометрии, например, сворачивание в трубку, подобно тому, что происходит
при образовании нервной трубки у эмбрионов позвоночных.
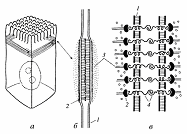
Рис 3. Адгезивный
(сцепляющий) поясок (лента)
а - расположение его в клетке; 6 - вид на
ультратонком срезе; в - схематическое изображение. 1
- плазматическая мембрана; 2 - слой винкулина; 3 - актиновые микрофиламенты; 4
- линкерные гликопротеиды
2.2
Десмосома (macula
adherens,
spot
desmosome)
Десмосома -
один из типов межклеточных контактов, обеспечивающих прочное соединение клеток.
Они обнаружены в различных тканях позвоночных и беспозвоночных животных -
эпителиях, эндотелии, мезотелии, в культуре печеночных клеток, а также между
отростками одной и той же клетки.
Десмосомы образуются между клетками тех тканей,
которые могут подвергаться трению, растяжению и другим механическим
воздействиям. В межклеточной щели в области десмосомы располагается
электронно-плотный слой, образованный взаимодействующими молекулами
интегральных гликопротеинов плазмолемм соседних клеток. Со стороны цитоплазмы к
десмосомам прикрепляются промежуточные филаменты, которые формируют в
цитоплазме сеть, обладающий большой прочностью на разрыв. Через десмосомы
промежуточные филаменты соседних клеток объединяются в непрерывную сеть,
охватывающую всю ткань.
Десмосома состоит из белков клеточной адгезии из
семейства кадгеринов и соединительных (адапторных) белков, которые соединяют их
с промежуточными филаментами. Белки клеточной адгезии, формирующие десмосомы-
десмоглеин и десмоколлин. Как и другие кадгерины, эти трансмембранные белки
имеют по пять внеклеточных доменов и являются кальцийсвязывающими. Они
обеспечивают гомофильное соединение клеток - между собой соединяются две
одинаковые по строению молекулы белка. Внутриклеточный белок десмоплакин (при
участии еще двух белков, плакофиллина и плакоглобина) соединяет внутриклеточные
домены десмоглеина с промежуточными филаментами.
Плакоглобин (белок с молекулярной массой 83 кД,
обнаруживающийся в адгезионных межклеточных контактах) возможно является
центральным пунктом в формировании десмосомы и прикреплении цитокератиновых
филаментов. Тип промежуточных филаментов зависит от типа клеток: в большинстве
эпителиальных клеток к десмосомам прикреплены кератиновые промежуточные
филаменты; в клетках сердечной мышцы - десминовые промежуточные филаменты.

Рис 4 Десмосомы. а - положение десмосом в
эпителиальных клетках.
б - структура десмосомы
ПД - полудесмосома
Д - десмосома
- промежуточные филаменты
- плазмалемма контактирующей клетки
- слой десмоглеина
- слой десмоплакина
Десмосомы обеспечивают механическую прочность,
необходимую для поддержания целостности эпидермиса. Система десмосом и
промежуточных филаментов в других тканях, по-видимому, имеет сходную роль.
Под световым микроскопом десмосомы (рис. 1)
имеют вид тонких мостиков, соединяющих противоположные края контактирующих
клеток. Существенными их элементами являются симметричные дифферентированные
участки цитоплазматических мембран протяженностью в среднем 0,2 - 0,5 мкм,
разделенных межклеточным пространством шириной 18 - 35 нм.
Рис 5. Десмосома. Электронная микрофотография
Общая площадь, занимаемая десмосомами, может составлять
до 5 % поверхности цитоплазматической мембраны. На микрофотографиях тонких
поперечных срезов демосом, по центру межклеточного пространства достаточно
часто выявляется зигзагообразный слой, который краями своих складок связывает
противоположные участки контактных мембран. При увеличении разрешения этот слой
выглядит фрагментированным - в виде выстроенных в ряд, нерегулярно
расположенных «глобул» диаметром 11 - 16 нм.
2.3
Полудесмосома, или гемидесмосома
Они обнаружены в тканях млекопитающих, амфибий и
костистых рыб. Полудесмосома по своей структурной организации напоминает
десмосому, разрезанную по межклеточной щели Но в отличие от десмосом,
соединяющих мембраны соседних эпителиальных клеток, гемидесмосомы присоединяют
базальную поверхность эпителиальных клеток к подлежащей базальной мембране, тем
самым, однако, также, как и десмосомы , функционируя в качестве заклепок,
распределяющих силы натяжения или разрыва, но уже на подлежащую эпителий
соединительную ткань . В то время как промежуточные филаменты, ассоциированные
с десмосомами, латерально прикрепляются к десмосомным бляшкам, многие из
промежуточных филаментов, ассоциированных с гемидесмосомами, своими концами
погружены в бляшку. Внутриклеточные прикрепляющие белки гемидесмосом отличны от
подобных белков десмосом. Трансмембранные линкерные белки гемидесмосом
принадлежат к интегриновому семейству рецепторов внеклеточного матрикса.
Как и десмосомы, гемидесмосомы прикрепляют
промежуточные филаменты, однако основным адгезионным рецептором в данном случае
является альфа-6 бета-4-интегрин, прикрепляющий ламинин (на ранних этапах
развития базальная мембрана состоит в основном из сети ламинина и не содержит
(или содержит мало) коллагена типа IV); ламинин, адгезивный гликопротеин -
большой (молекулярная масса 850000) гибкий комплекс из длинных полипептидных
цепей, ассоциированных в форме асимметричного креста и удерживаемых вместе при
помощи дисульфидных связей. Содержит несколько функциональных доменов:
связывающиеся с коллагеном типа IV, с гепаран сульфатом, с энтактином, c
рецепторами ламинина на клеточной поверхности к базальной пластинке. Остальные
белки, составляющие гемидесмосому, также уникальны, хотя и отчасти гомологичны
десмосомальным белкам.

Рис 6. Схема строения полудесмосомы
2.4
Фокальные контакты
Они встречаются у многих клеток и
особенно хорошо изучены у фибробластов. Они построены по общему плану со
сцепляющими лентами, но выражены в виде небольших участков - бляшек на
плазмолемме. В этом случае трансмембранные линкерные белки-интегрины
специфически связываются с белками внеклеточного матрикса (например с
фибронектином). Со стороны цитоплазмы эти же гликопротеиды связаны с
примембранными белками, куда входит и винкулин, который в свою очередь связан с
пучком актиновых филаментов.
Функциональное значение фокальных контактов
заключается как в закреплении клетки на внеклеточных структурах, так и создании
механизма, позволяющего клеткам перемещаться.
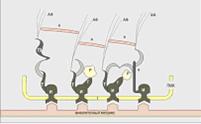
Рис 7. Упрощенная схема фокального контакта
клетки с внеклеточным матриксом
ПМК -
плазматическая мембрана клетки,
И - интегриновый
рецептор,
Т - талин, - FAK
(протеинкиназа фокального контакта),
Р- паксиллин,
В - винкулин,
ТЗ - тензин, а -
альфа-актинин,
АФ - пучки актиновых
микрофиламентов
3.
Коммуникационные соединения
Контакты коммуникационного типа позволяют
клеткам обмениваться веществами (нексусы) или сигналами (синапсы).
3.1
Щелевое соединение, или нексусы (nexus)
Представляет собой область протяженностью 0,5 -
3 мкм, где плазмолеммы разделены промежутком в 2 - 3 нм. Со стороны цитоплазмы
никаких специальных примембранных структур в данной области не обнаруживается,
но в структуре плазмалемм соседних клеток друг против друга располагаются
специальные белковые комплексы - коннексоны.
В зонах щелевого контакта может быть от 10 - 20
до нескольких тысяч коннексонов в зависимости от функциональных особенностей
клеток. Они состоят из шести субъединиц коннектина - трансмембранного белка с
молекулярным весом около 30 тыс. Объединяясь друг с другом, коннектины образуют
цилиндрический агрегат - коннексон, в центре которого располагается канал.
Отдельные коннексоны встроены в плазматическую мембрану так, что прободают ее
насквозь. Одному коннексону на плазматической мембране клетки точно
противостоит коннексон на плазматической мембране соседней клетки так, что
каналы двух коннексонов образуют единое целое.
Коннексоны играют роль прямых межклеточных
каналов, по которым могут диффундировать вещества из клетки в клетку с молекулярным
весом не более 1-1,5 тыс. и размером не более 1,5 нм (у насекомых через щелевой
контакт могут проходить вещества с молекулярным весом до 2 тыс.). Среди этих
веществ были разные ионы, аминокислоты, нуклеотиды, сахара, витамины, стероиды,
гормоны, АМФ. В настоящее время известно о 20 генах, кодирующих образование
коннексинов у человека. Они находятся во многих хромосомах, образуя скопление
только в первой хромосоме.
Было обнаружено, что коннексоны могут
закрываться, изменяя диаметр внутреннего канала, и тем участвовать в регуляции
транспорта молекул между клетками. Ни белки, ни нуклеиновые кислоты через
щелевые контакты проходить не могут. Такая способность щелевых контактов
служить местом транспорта низкомолекулярных соединений используется в тех клеточных
системах, где нужна быстрая передача электрического импульса (волны
возбуждения) от клетки к клетке без участия нервного медиатора. Так, все
мышечные клетки миокарда сердца связаны с помощью щелевых контактов (кроме
того, клетки там связаны и адгезивными контактами). Это создает условие для
синхронного сокращения огромного количества клеток. При росте культуры
эмбриональных сердечных мышечных клеток (миокардиоциты) некоторые клетки в
пласте начинают независимо друг от друга спонтанно сокращаться с разной
частотой, и лишь только после образования между ними щелевых контактов они
начинают биться синхронно как единый сокращающийся пласт клеток. Таким же
способом обеспечивается совместное сокращение гладкомышечных клеток в стенке
матки.
Целостность и функционирование щелевых контактов
сильно зависит от уровня ионов Ca2+ внутри клетки. Ионы Ca2+
меняют конфигурацию коннексонов так, что просвет каналов закрывается. В норме
концентрация кальция в цитоплазме очень низка. Если Ca2+
инъецировать в одну из клеток пласта культуры тканей, то в соседних клетках
увеличения уровня Ca2+ в цитоплазме не происходит; клетки как бы
разобщаются с соседями, перестают проводить электрический ток и красители.
Через некоторое время, после того как введенный кальций будет аккумулирован
митохондриями, структура и функции щелевых контактов восстанавливаются. Такое
свойство очень важно для поддержания целостности и работы всего слоя клеток,
так как повреждение одной из них не передается на соседний через щелевые
контакты, которые перестают работать как межклеточные диффузионные каналы.

Рис 8. Структура канала щелевого контакта
Каждый ионный канал состоит из двух половинок -
коннексонов. Коннексон пронизывает мембрану лишь одной клетки и выступает в
межклеточную щель на 1-1,5 нм, где стыкуется со вторым коннексоном.
3.2
Синаптические соединения, или синапсы (synapsis)
Согласно классическому определению, синапсы
представляют собой специализированные функциональные контакты между клетками
возбудимых тканей. Термин «синапс» ввел Ч. Шеррингтон (1897).
Этот тип контактов характерен для нервной ткани
и встречается в специализированных участках контакта как между двумя нейронами,
так и между нейроном и каким-либо иным элементом - рецептором или эффектором
(например, нервно-мышечное окончание).
Синапсы - участки контактов двух клеток,
специализированных для односторонней передачи возбуждения или торможения от
одного элемента к другому. В принципе подобного рода функциональная нагрузка,
передача импульса может осуществляться и другими типами контактов (например,
щелевым контактом в сердечной мышце), однако в синаптической связи достигается
высокая эффективность в реализации нервного импульса.
В синапсе различают несколько составных частей
(рис 9):
) пресинаптическая мембрана (ПреМ):
представляет собой расширенное окончание клетки (нейрона). Именно здесь
располагаются многочисленные синаптические пузырьки (везикулы), окруженные
мембраной структуры диаметром от 10 до 90 нм, содержащие химическое вещество (медиатор
или нейромедиатор). Пресинаптическая мембрана представляет собой участок
плазмалеммы, непосредственно контактирующий с соседней клеткой;
) синаптическая щель: участок
межклеточного пространства, отделяющий пресинаптическую клетку от
постсинаптической;
) постсинаптическая мембрана (ПоМ):
образована участком плазматической мембраны другой клетки, содержит встроенные
белковые молекулы - рецепторы, способные обратимо связываться с
нейромедиатором, вызывая впоследствии генерацию электрического импульса в
постсинаптическом нейроне.
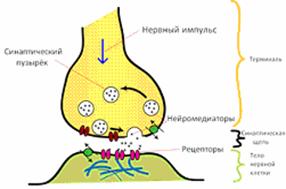
Рис 9. Основные
элементы синапса
Обычно сигнал предается химическим веществом -
медиатором: последний диффундирует от ПреМ и воздействует на специфические
рецепторы в ПоМ.
В зависимости от морфологии (рис. 10)
контактирующих пре- и постсинаптических мембран выделяют синапсы двух типов:
асимметричные,
1-го типа, и симметричные, 2-го типа.

Рис 10. синапсы 1-го и 2-го типов: а -
асимметричный синапс; б - симметричный синапс.
Они отличаются друг от друга поряду признаков:
синаптическая щель
в синапсах 1-го типа шире синаптической щели синапсов 2-го типа
постсинаптическая
мембрана синапсов 1-го типа толще и плотнее;
синапсы 1-го типа длиннее, а синапсы 2-го типа
короче
синаптические пузырьки многочисленны в синапсах
1-го типа, имеют
округлую форму (30-60 нм в диаметре), а в
синапсах 2-го типа они
овальной или дисковидной формы, менее
многочисленны, их размер составляет 10-30 нм;
в синаптической щели синапсов 1-го типа (ближе к
постсинаптической мембране) расположена бляшка из внеклеточного вещества.
4.
Запирающий контакт (zona
occuludens)
Плотное соединение характерно для однослойных
эпителиев. Это зона, где внешние слои двух плазматических мембран максимально
сближены. Часто видна трехслойность мембраны в этом контакте: два внешних
осмофильных слоя обеих мембран как бы сливаются в один общий слой толщиной 2-3
нм. Слияние мембран происходит не по всей площади плотного контакта, а
представляет собой ряд точечных сближений мембран.
Было обнаружено, что точки соприкосновения
мембран представляют собой ряды глобул. Это белки окклудин и клаудин,
специальные интегральные белки плазматической мембраны, встроенные рядами.
Такие ряды глобул или полоски могут пересекаться так, что образуют на
поверхности скола как бы решетку или сеть.
Этот тип соединений характерен для эпителиев,
особенно железистых и кишечных. В последнем случае плотный контакт образует
сплошную зону слияния плазматических мембран, опоясывающую клетку в апикальной
(верхней, смотрящей в просвет кишечника) ее части. Таким образом, каждая клетка
пласта как бы обведена лентой этого контакта. Такие структуры при специальных
окрасках можно видеть и в световом микроскопе. Они получили у морфологов
название замыкающих пластинок. Оказалось, что в данном случае роль замыкающего
плотного контакта заключается не только в механическом соединении клеток друг с
другом.
Эта область контакта плохо проницаема для
макромолекул и ионов, и следовательно, она запирает, перегораживает
межклеточные полости, изолируя их (и вместе с ними собственно внутреннюю среду
организма) от внешней среды (в данном случае - просвет кишечника).
Плотный контакт
встречается между всеми типами однослойного эпителия (эндотелий, мезотелий,
эпендима). Основной функцией которой является создание барьера, ограничивающего
диффузию веществ между наружной средой и межклеточным пространством.
Трехмерное схематическое изображение фрагмента
плотного контакта представлено на рис.7
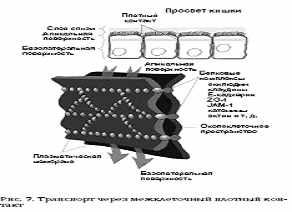
5.
Септированнные соединения
5.1
Плазмодесмы
Этот тип межклеточных связей встречается у
растений. Плазмодесмы представляют собой тонкие трубчатые цитоплазматические
каналы, соединяющие две соседние клетки. Диаметр этих каналов обычно составляет
20-40 нм. Ограничивающая эти каналы мембрана непосредственно переходит в
плазматические мембраны соседствующих клеток. Плазмодесмы проходят сквозь
клеточную стенку, разделяющую клетки.
Таким образом, у некоторых растительных клеток
плазмодесмы соединяют гиалоплазму соседних клеток, поэтому формально здесь нет
полного разграничения, отделения тела одной клетки от другой, это скорее
представляет собой синцитий: объединение многих клеточных территорий с помощью
цитоплазматических мостиков.
Внутрь плазмодесм могут проникать мембранные
трубчатые элементы, соединяющие цистерны эндоплазматического ретикулума
соседних клеток. Образуются плазмодесмы во время деления клетки, когда строится
первичная клеточная оболочка (см. ниже). У только что разделившихся клеток
число плазмодесм может быть очень велико (до 1000 на клетку), при старении
клеток их число падает за счет разрывов при увеличении толщины клеточной
стенки.
Функциональная роль плазмодесм очень велика: с
их помощью обеспечивается межклеточная циркуляция растворов, содержащих
питательные вещества, ионы и другие соединения. По плазмодесмам могут
перемещаться липидные капли.
Через плазмодесмы происходит заражение клеток
растительными вирусами. Однако эксперименты показывают, что свободный транспорт
через плазмодесмы ограничивается частицами с массой не более 800 дальтон.

Рис 11.
Схематическая структура плазмодесмы.
1 - клеточная
стенка
- плазмолемма
- десмотубула
-
эндоплазматический ретикулум
5 - белки
плазмодесмы
механизм структура
клетка соединение
Заключение
Морфологические структуры, возникающие в местах
соприкосновения клеток в тканях, носят название межклеточных контактов.
В зависимости от выполняемой функции их можно классифицировать:
1. Функция герметизации отсеков
межклеточного пространства между соседними клетками.
В результате мелкие водорастворимые молекулы не способны легко и быстро
перемещаться во внеклеточном матриксе.
В организме позвоночных этот тип соединений
представлен плотными контактами. Здесь плазмолеммы прилегают друг к другу
вплотную, сцепляясь с помощью специальных белков. Места такого плотного
прилегания образуют на контактирующих поверхностях подобие ячеистой сети. Тем
самым обеспечивается надежность отграничения двух сред, находящихся по разные
стороны от пласта клеток.
. Функция скрепления клеток друг с
другом. В результате ткани приобретают механическую
прочность, а различные клеточные типы не смешиваются в пределах одного органа
(ткани).
Наиболее просто устроенными типами подобных
соединений являются рыхлые (простые) контакты, в англоязычной литературе
именуемые слипающимися контактами (adherens junction).
Это просто сближение плазмалемм соседних клеток
на расстояние 15-20нм. без образования специальных структур.
При этом плазмолеммы взаимодействуют друг с
другом с помощью специфических адгезивных гликопротеинов - кадгеринов,
интегринов и др.
3. Функция коммуникации между
клетками как в пределах одной ткани, так и между разными
типами тканей. Благодаря этим соединениям осуществляется транспорт веществ и
передача сигналов. Представлены щелевыми контактами и синапсами, плазмодесмами.
Литература
. А. В. Сидоров Физиология
межклеточной коммуникации : учеб. пособие / А. В. Сидоров. - Минск: БГУ, 2008.
- 215 с.
. С. Л. Кузнецов, Н. Н. Мушкамбаров
К89 Гистология, цитология и эмбриология: Учебник для медицинских вузов. - М. :
ООО «Медицинское информационное агенство», 2007. - 600 с. ; ил., табл.
. Гистология: Учебник/ Ю. И.
Афанасьевой, Н. А. Юрина, Е. Ф. Котовский и др.; Под ред. Ю. И. Афанасьева, Н.
А. Юриной. - 5-е изд., перераб. и доп. - М.: Медицина, 2001. - 744с.: ил.