Кинетические закономерности ферментативных реакций
Курсовая
работа по физической химии
"Кинетические
закономерности ферментативных реакций"
Содержание
Введение
Задание №1. Какие
соединения получили название "ферменты", "энзимы", какова
их природа и внутренние строение. Два примера ферментативной реакции
1. Кинетические закономерности
односубстратных ферментативных реакций
1.1
Кинетические закономерности односубстратных ферментативных реакций
фермент односубстратный ингибирование
1.2
Исследование кинетики реакции окисления
сукцината натрия в фумарат натрия под действием сукциноимидазы
2.
Ингибирование ферментативных реакций
2.1
Ингибирование ферментативных реакций. Теоретические сведения
2.2
Исследование кинетической закономерности заданной
реакции, осложненной ингибированием( ингибирование напрексеном
простагландин-Н-синтетазы (PGH)
в ферментативной реакции, одним из субстратов которой является адреналин)
Вывод
Список
используемой литературы
Введение
В настоящее время химия одна из самых важных
наук, я бы даже сказал что она на первом месте среди всех остальных наук. Она
изучает много химических реакций, из которых наиболее важные являются
ферментативные реакции, так как ежедневно в нашем организме происходит
множество ферментативных реакций без которых мы не можем обойтись.
Физическая химия является одной из дисциплин
которая занимается изучением кинетический закономерностей ферментативных
реакций. В настоящее время физическая химия занимается также изучением строения
веществ, химическую термодинамику, химическую кинетику, термохимию, электрохимию
и т.д.
Ферменты широко используются для лечения
заболеваний, например при сердечной недостаточности, сахарном диабете.
Задание №1. Какие
соединения получили название "ферменты", "энзимы", какова
их природа и внутренние строение. Два примера ферментативной реакции
Ферментативные
реакции подчиняются общим законам катализа, но вследствие сложности состава и
строения ферментов образуют специфическую и весьма обширную область
каталитических реакций. Биологические катализаторы белковой природы, ускоряющие
каталитические реакции в живых системах, называются в отечественной литературе
ферментами, в англоязычной - энзимами. Не каждый белок является ферментом, но в
составе практически каждого белка находится определенный набор групп,
определяющих активность фермента.
Ферменты
представляют собой компактные молекулы с молекулярной массой от 104
и диаметром от 20 А и выше. Входящие в их состав глобулярные белки состоят из
расположенных в определенной последовательности α -аминокислотных
остатков, соединенных пептидными связями (-CO-NH-), причем установлено,
что в построении большинства ферментных белков участвуют 20 или 21 аминокислот.
Внутреннее строение
ферментов обычно характеризуют в биохимии наличием четырех типов структур:
первичная
определяется только генетическим кодом и указывает последовательность
аминокислот для каждого белка;
вторичная структура
определяет характер свертывания в спираль, или спирализации, цепи;
третичная
характеризует способ пространственной укладки частичного или полностью
свернутой в спираль полипептидной цепи;
четвертичная -
взаимное расположение и характер объединения структурно независимых глобул в
активный олигомерный фермент.
Строго говоря,
такие представления являются идеализацией, поскольку не существует независимых
вторичной и третичной структур: в процессе пространственной укладки
полипептидной цепи происходит как скручивание, обусловленное силами притяжения
и отталкивания различных её элементов, так и их укладка в компактную глобулу
фермента.
Но использование
указанных выше терминов, хотя и относится к приближенным методам описания
белковых молекул, является полезным при качественном анализе общих структурных
проблем в той области, которая соответствует разрешению ферментной глобулы
примерно 6-8 А.
Следует отметить,
что именно третичная структура играет большую роль в создании специфических
отличий ферментов от остальных катализаторов: при ее возникновении в глобуле
фермента образуется в виде "щели" адсорбционный участок активного
центра.
Понятие четвертичной
структуры приобретает ясный физический смысл, если объединение в глобулу не
изменяет строения объединяющихся частей. В действительности отдельные
субъединицы ферментов изменяют свои конформации при ассоциации, поэтому понятие
четвертичной структуры является еще менее строгим, чем двух предыдущих.
По-видимому, речь идет о зависимости пространственного строения белковой глобулы от всех
межмолекулярных взаимодействий в системе. В результате нестрогим понятие
вторичной, третичной и четвертичной структур.
1. Кинетические закономерности
односубстратных ферментативных реакций
.1 Кинетика
ферментативной реакции с одним субстратом
Уже в ранних работах
было обнаружено, что начальная скорость ферментативной реакции при
фиксированной концентрации фермента и малых концентрациях субстрата линейно
зависит от концентрации субстрата, но для высоких концентраций субстрата такой
зависимости уже не наблюдалось.
Л.Анри (1903 г.) первым
предположил, что фермент образует промежуточное соединение с субстратом. Если
обозначить фермент Е, субстрат S,
то это соединение (интермедиат) обозначится ES.
Л.Михаэлис предложил для
количественного описания кинетики этой реакции механизм, состоящий из двух
стадий (первая - обратимая и быстрая, вторая -необратимая и медленная):


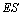

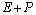 (2.1)
(2.1)
Система кинетических
уравнений для реакции (1) имеет довольно сложный вид:
 (2.2)
(2.2)
 (2.3)
(2.3)
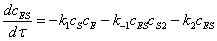 (2.4)
(2.4)
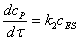 (2.5)
(2.5)
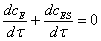 (2.6)
(2.6)
 (2.7)
(2.7)
Точное аналитическое
решение такой системы представляет большие сложности. Поэтому на практике очень
часто используют приближения квазиравновесных и стационарных концентраций.
Для данного механизма
скорость реакции определяется скоростью образования конечного продукта
 (2.8)
(2.8)
В свою очередь, скорость
образования промежуточного соединения ES,
согласно закону действующих масс, можно записать:
(2.9)
Уравнения материального
баланса для фермента и субстрата имеют вид:
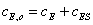 (2.10)
(2.10)
и
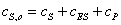 (2.11)
(2.11)
где 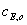 и
и
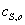 -
начальные концентрации фермента и субстрата;
-
начальные концентрации фермента и субстрата; 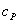 и
и  -
их текущие концентрации;
-
их текущие концентрации; и -
текущие концентрации промежуточного соединения и продукта.
и -
текущие концентрации промежуточного соединения и продукта.
Решение системы из трех
уравнений (9) - (11) находится при выполнении следующих условий:
1) на практике основные
параметры ферментативной реакции находят, изучая зависимость начальной скорости
реакции от начальной концентрации субстрата.
Поэтому можно считать,
что при  ,
,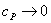 т.е.
т.е. 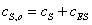 ;
;
'2) обычно >>,
поэтому с учетом условия (1) 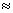
3) для ферментативных
реакций характерно следующее соотношение констант скоростей стадий 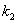 >>
>> ,
т.е промежуточное вещество ( ES ) является неустойчивым и его концентрация мала. В этом случае
используют приближение квазистационарных концентраций, суть которого выражена
соотношением:
,
т.е промежуточное вещество ( ES ) является неустойчивым и его концентрация мала. В этом случае
используют приближение квазистационарных концентраций, суть которого выражена
соотношением:

Подставляя значение 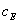 из
уравнения (10) в уравнение (9) и решая его относительно ,
получаем выражение для
из
уравнения (10) в уравнение (9) и решая его относительно ,
получаем выражение для
 (2.12)
(2.12)
которое после деления
числителя и знаменателя на принимает
вид:
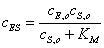 (2.13)
(2.13)
Величину
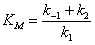
принято по решению ИЮПАК
называть константой Михаэлиса.
Подставляя выражение
(13) в (8), получаем уравнение для скорости образования продукта в начальные
моменты времени
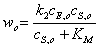 (2.14)
(2.14)
Анализ уравнения (14)
позволяет объяснить наблюдаемые на опыте закономерности.
Если <<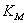 ,
тогда
,
тогда
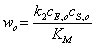 (2.15)
(2.15)
Если >>,
то из уравнения (14) получаем
 , (2.16)
, (2.16)
т.е. начальная скорость
реакции не зависит (как и в опыте при больших значениях концентрации субстрата)
от концентрации субстрата, порядок реакции по субстрату будет нулевой, по
ферменту - первый, а сама скорость достигает предельного значения, 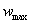 :
:
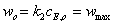
При достижении  значения
(кроме
предельной, ее называют еще максимальной скоростью) концентрация
фермент-субстратного комплекса изменяется от до
значения
(кроме
предельной, ее называют еще максимальной скоростью) концентрация
фермент-субстратного комплекса изменяется от до  ,
а из уравнения (13) следует, что =
. Это означает, что весь фермент переходит в состав промежуточного
соединения, т.е. фермент насыщен субстратом. Именно в этих условиях удобно
сравнивать скорости различных реакций, поэтому запишем уравнение (14) в виде:
,
а из уравнения (13) следует, что =
. Это означает, что весь фермент переходит в состав промежуточного
соединения, т.е. фермент насыщен субстратом. Именно в этих условиях удобно
сравнивать скорости различных реакций, поэтому запишем уравнение (14) в виде:
 (2.18)
(2.18)
где и
-
параметры уравнения Михаэлиса - Ментен
Следует отметить, что
уравнение вида (18) характерно для кинетики гомогенных, ферментативных и
гетерогенных каталитических реакций.
Физический смысл
константы Михаэлиса можно пояснить, исходя из уравнения (18): она численно
равна концентрации субстрата (=),
при которой активность фермента составляет половину максимальной, т.е. =
0,5 .
Это наглядно видно из графика в координатах (, ),
представленного на рис. 1.
Константа Михаэлиса и
начальная концентрация субстрата имеют
одинаковую размерность концентраций: М (моль-л-1). Численные
значения обычно
лежат в пределах 10 -2-10-8 М, легко воспроизводятся и не
зависят от концентрации фермента. В то же время является
функцией температуры, рН среды, зависит от присутствия других веществ, играющих
роль ингибитора или активатора.
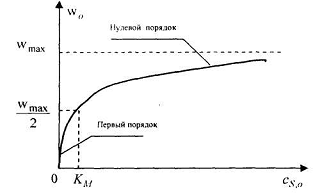
Рис.1. Зависимость
начальной скорости ферментативной реакции от начальной концентрации субстрата
Если для
рассматриваемого механизма реакции выполняется условие: 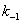 >>,
этом случае можно говорить, что на стадии
>>,
этом случае можно говорить, что на стадии
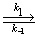 (2.19)
(2.19)
устанавливается
равновесие. Это позволяет использовать при анализе ферментативных реакций приближение
квазиравновесных концентраций. В этом случае выражение для константы Михаэлиса
упрощается и становится прозрачным ее физико-химический смысл:
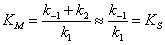 , (2.20)
, (2.20)
где 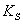 -
так называемая субстратная константа, т.е. константа диссоциации
фермент-субстратного комплекса на фермент и субстрат, характеризующая меру
связывания фермента с субстратом. В этом случае можно рассматривать как
величину, характеризующую сродство фермента к субстрату.
-
так называемая субстратная константа, т.е. константа диссоциации
фермент-субстратного комплекса на фермент и субстрат, характеризующая меру
связывания фермента с субстратом. В этом случае можно рассматривать как
величину, характеризующую сродство фермента к субстрату.
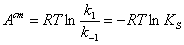 (2.21)
(2.21)
где 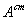 -
стандартное химическое сродство реакции (19).
-
стандартное химическое сродство реакции (19).
В общем случае ≥,
причем экспериментальное значение представляет
собой максимальное значение .
Иная картина наблюдается
только для ферментов разложения пероксида водорода, каталазы и пероксидазы, для
которых >>.
В этих случаях константа Михаэлиса, /
сильно отличается от субстратной константы (не
работает приближение квазиравновесных концентраций).
Величина ,
определяемая из экспериментальных, данных по уравнению (18), для ферментов с
неизвестной молекулярной массой является просто кинетическим параметром. Но для
ферментов с известной молекулярной массой, т.е. таких, для которых можно
рассчитать начальную концентрацию , эта величина
становится фундаментальным параметром, так как позволяет определить из
соотношения (11) значение константы :
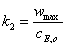 ,
(2.22)
,
(2.22)
характеризующей скорость
распада фермент- субстратного комплекса на продукт и фермент.
Величину называют
также числом оборотов фермента, так как она указывает на число моль субстрата,
превращенных в продукт одним молем фермента в единицу времени.
Для определения
параметров и
используют
несколько способов линеаризации уравнения Михаэлиса - Ментен, которые позволяют
выразить экспериментальные данные в удобной для анализа линейной графической
форме. Однако в рамках этой работы будет рассмотрен наиболее часто используемый
способ, носящий название способ Лайнуивера - Берка .
После записи уравнения
(18) в виде обратных величин, получается соотношение:
 , (2.23)
, (2.23)
которое удобно
использовать для построения линейного графика в координатах(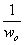 ,
,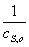 ).
Типичная зависимость, получаемая в подобных координатах приведена на рис. 2.
).
Типичная зависимость, получаемая в подобных координатах приведена на рис. 2.
Очевидно, что отрезок,
отсекаемый на оси ординат при нулевом значении обратной начальной концентрации
субстрата, соответствует обратной величине максимальной скорости реакции (на
рис. 2 это отрезок О А).
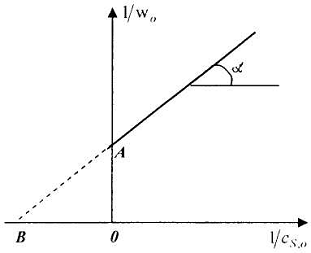
Рис.2. Определение
параметров и
по способу Лайнувера -
Берка
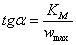 (2.24)
(2.24)
Из анализа уравнения
(23) также видно, что отрезок, отсекаемый продолжением линии зависимости
обратной начальной скорости реакции от обратной начальной концентрации
субстрата на оси абсцисс, равен -1/ -
обратной величине константы Михаэлиса, взятой с минусом (на рис. 2 это отрезок ОВ).
.2 Исследование реакции окисления
сукцината натрия в фумарат натрия под действием сукциноимидазы
Зависимость концентрации
субстрата от
времени
Таблица 1.
|
Время, мин
|
0
|
5
|
10
|
20
|
30
|
40
|
|
 ,
моль/л1,671,41,110,750,50,35 ,
моль/л1,671,41,110,750,50,35
|
|
|
|
|
|
|
Таблица 2.
|
Время, мин
|
0
|
3
|
6
|
10
|
20
|
30
|
|
,
моль/л0,830,70,550,40,250,17
|
|
|
|
|
|
|
Таблица 3.
|
Время, мин
|
0
|
3
|
6
|
10
|
20
|
30
|
|
,
моль/л0,560,440,320,240,120,06
|
|
|
|
|
|
|
Таблица 4.
|
Время, мин036102030
|
|
|
|
|
|
|
|
,
моль/л0,420,320,210,150,060,03
|
|
|
|
|
|
|
Таблица 5.
|
Время, мин
|
0
|
6
|
10
|
20
|
30
|
|
,
моль/л0,330,260,150,10,030,005
|
|
|
|
|
|
|
Находим области
линейного изменения концентрации субстрата во времени по графикам
1)  ,
моль/л
,
моль/л
) 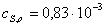 ,
моль/л
,
моль/л
) 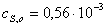 ,
моль/л
,
моль/л
)  ,
моль/л
,
моль/л
) 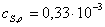 ,
моль/л
,
моль/л
Найдем значения
начальных скоростей по каждой кинетической кривой.
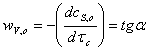 ,
,
где 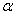 -
угол наклона прямой, который совпадает с прямолинейной областью зависимости
концентрации субстрата от времени.
-
угол наклона прямой, который совпадает с прямолинейной областью зависимости
концентрации субстрата от времени.
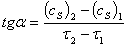 ,
,
где 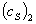 и
и
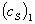 -
ординаты точек 1 и 2, лежащих в области прямолинейной зависимости концентрации
субстрата от времени;
-
ординаты точек 1 и 2, лежащих в области прямолинейной зависимости концентрации
субстрата от времени;  и
и
 -
их абсциссы
-
их абсциссы
Рассчитываем скорости по
значению концентрации субстрата и времени, приведенным в таблице.
1)  =
=
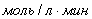 ;
;
)  =
=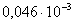 ;
;
) 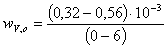 =
=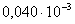 ;
;
) 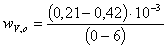 =
= ;
;
)  =
= ;
;
Полученные данные
занесем в таблицу.
Зависимость начальных
скоростей реакции от начальных концентраций субстрата
|
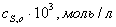 1,670,830,560,420,33 1,670,830,560,420,33
|
|
|
|
|
|
|
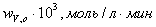 0,0560,0460,0400,0350,030 0,0560,0460,0400,0350,030
|
|
|
|
|
|
Построим график
зависимости начальной скорости реакции от начальной концентрации субстрата.
Отображаем данную
зависимость в двойных координатах, чтобы линеаризовать уравнение
Михаэлиса-Ментен для нахождния параметров и .
Зависимость обратной
начальной скорости от обратной начальной концентрации субстрата
|
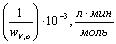 17,8621,502528,5733,33 17,8621,502528,5733,33
|
|
|
|
|
|
|
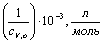 0,601,201,782,383,03 0,601,201,782,383,03
|
|
|
|
|
|
Найдем и
графическим
способом
Отрезок, отсекаемый на
оси ординат при нулевом значении обратной начальной концентрации субстрата,
соответствует обратной величине максимальной скорости реакции (отрезок ОА).
Отрезок отсекаемый
продолжением линии зависимости обратной начальной скорости реакции от обратной
начальной концентрации субстрата на оси абсцисс, равен обратной величине
константы Михаэлиса, взятой с минусом ( отрезок ОВ).
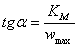 ;
;
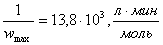 ;
;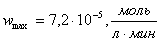
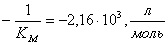 ;
;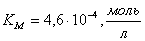
 ;
;
Уравнение
Михаэлиса-Ментен с найденными параметрами:
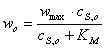 ;
;
2.
Ингибирование ферментативных реакций
.1 Ингибирование
ферментативных реакций. Теоретические сведения
Вещества, присутствие
которых в системе понижает активность фермента, т. е. уменьшает скорость
реакции, называются ингибиторами.
Если для ферментативного
катализа необходимы предварительная адсорбция субстрата и его строгая
ориентация относительно активных групп каталитического центра, то для
ингибирования достаточно не только взаимодействия со всем адсорбционным
участком, но и простого связывания ингибитора с отдельными участками
адсорбционного центра.
Соединения проявляют
свойства ингибиторов либо благодаря возможности образовывать с каталитическим
центром прочные комплексы (сероводород, цианиды), либо из-за воздействия на
карбонильную группу фермента, либо благодаря своей способности денатурировать
белки (например, соли серебра, меди, ртути).
Эффект ингибирования
может иметь место по разным причинам. Среди возможных типов обратимого
ингибирования остановимся на трех.
1.Ингибитор I конкурирует с субстратом
за активный центр, т.к. по строению близок к нему и является псевдосубстратом,
образуя с ферментом неактивный комплекс (EI). Если при увеличении концентраций субстрата в растворе активность
фермента восстанавливается, такое ингибирование называется конкурентным.
2.Ингибитор не конкурирует с субстратом за активный центр, так как
присоединяется к другой части молекулы белка с образованием неактивных
комплексов (EI и ESI), что приводит к снижению активности фермент но не изменяет его
сродство к субстрату. Если достигается обратимость ингибирования, т.е.
восстановление активности фермента под действием отличных от субстрата веществ,
такое ингибирование называется неконкурентным.
З.Если при
ингибировании по типу 2 образуется только один неактивный комплекс ESI (фермент- субстрат- ингибитор), имеет место бесконкурентное ингибирование.
Реакция
конкурентного ингибирования протекает по следующему механизму:
(3.25)
 ( неактивный
комплекс)(3.26)
( неактивный
комплекс)(3.26)
Величина, определяемая
как
 ,(3.27)
,(3.27)
Называется константой
ингибирования ( )
или константой диссоциации комплекса EI.
)
или константой диссоциации комплекса EI.
Для математического
решения задачи используется следующие допущения:
) рассматривается, как и
ранее начальный этап реакции, т.э. при 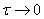 ,
,  ,
т.э.
,
т.э. 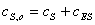 ;
;
) поскольку  >>
>> ,
то
,
то  ;
;
) как и ранее
предполагается, что работает приближение квазистационарных концентраций, т.э. 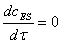 ;
;
) 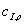 >>,
т.э
>>,
т.э  - начальная
концентрация ингибитора.
- начальная
концентрация ингибитора.
Скорость реакции
определяется скоростью образования конечного продукта
 (3.28)
(3.28)
В присутствии ингибитора
уравнение материального баланса для фермента примет вид:
 (3.29)
(3.29)
Используя (27)
преобразуем выражение (29)
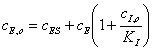 ,(3.30)
,(3.30)
Для нахождения
концентрации ES
используем принцип стационарных концентраций:
 ,(3.31)
,(3.31)
откуда
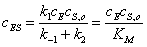 ,(3.32)
,(3.32)
 ,(3.33)
,(3.33)
Полученное выражение
подставляем в уравнение (30), которое решаем относительно ;
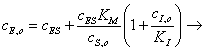
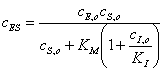 (3.34)
(3.34)
Из (28) с учетом (34)
можно получить выражение для начальной скорости реакции, ускоряемой ферментом в
присутствии конкурентного ингибитора:
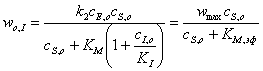 ,(3.35)
,(3.35)
где 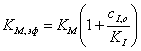 ,(3.36)
,(3.36)
Эффективная константа,
зависящая от 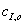 и
и
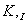
Сравнение уравнений
Михаэлиса - Ментен (23) и (36) показывает, что начальная скорость реакции в
присутствии ингибитора всегда меньше скорости реакции в его отсутствие.
Предельные значения скорости в обоих случаях для больших концентраций субстрата
одинаковы и равны:
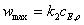 (3.37)
(3.37)
Из выражения (36) можно
получить соотношение для вычисления константы ингибирования:
 (3.38)
(3.38)
Таким образом, для
определения константы ингибирования кроме знания начальной концентрации
ингибитора, необходимо найти эффективную константу реакции в присутствии
ингибитора и константу Михаэлиса в его отсутствие.
Графическую обработку
результатов опытов удобно проводить в двойных обратных координатах (
координатах Лайнуивера - Берка).

Рис. 3. Обработка экспериментальных
данных ферментативной реакции, осложненной конкурентным ингибированием:
1 - в отсутствие
ингибитора;
2-с ингибитором
На рис.3 приведено
графическое изображение опытных данных по изучению кинетики ферментативной
реакции, осложненной конкурентным ингибированием.
Обратная величина
предельного значения скорости легко найдется графически по величине отрезка,
отсекаемого прямой в двойных обратных координатах на оси ординат (на рис. 3 это
отрезок ОА). Располагая значением предельной скорости реакции и
определив величины угловых коэффициентов прямых в двойных обратных координатах,
по формуле (24) можно определить величины константы Михаэлиса в отсутствие
ингибитора (по линии 1) и эффективной константы реакции в присутствии
ингибитора (по линии 2).
Константу ингибирования
находят по формуле (38) или графически, так как отношение тангенсов углов
наклона прямых в точке пересечения с осью ординат равно 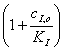
Простейший механизм
процесса при ферментативной реакции, осложненной неконкурентным
ингибированием может быть представлен схемой
(3.39)
(3.40)
 (3.41)
(3.41)
Здесь имеются две
константы диссоциации комплексов
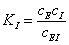 иK’=
иK’=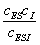 (3.42)
(3.42)
Если оба комплекса
неактивны, т.е. не дают продукта, а константы их диссоциации равны между собой:
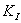 =К',
то это равносильно дополнительному условию: ингибитор не мешает
взаимодействию субстрата с ферментом, но уменьшает скорость реакции из-за
неактивности образующихся комплексов на поверхности фермента.
=К',
то это равносильно дополнительному условию: ингибитор не мешает
взаимодействию субстрата с ферментом, но уменьшает скорость реакции из-за
неактивности образующихся комплексов на поверхности фермента.
Решая задачу аналогично
предыдущей, получаем выражение для начальной скорости реакции в следующем виде:
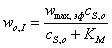 ,(3.43)
,(3.43)
где 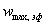 -
эффективная предельная скорость реакции, причем
-
эффективная предельная скорость реакции, причем
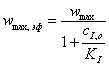 (3.44)
(3.44)
Из выражения (43) видно,
что при таком типе ингибирования изменяется предельная скорость процесса, а
константа Михаэлиса остается постоянной. Из соотношения (44) можно получить
выражение для константы ингибирования:
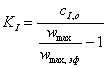 ,(3.45)
,(3.45)
которое показывает, что
константу ингибирования легко вычислить при известной начальной концентрации
ингибитора, измеряя скорости реакции в отсутствие и в присутствии ингибитора.
Обработка
экспериментальных данных для неконкурентного ингибирования в двойных обратных
координатах (рис.4) позволяет найти все параметры уравнения Михаэлиса:
величина, обратная предельной скорости определяется по отрезку, отсекаемому на
оси ординат прямой в опыте без ингибирования (на рис.4 это отрезок ОА1),
величина, обратная эффективной предельной скорости находится по отрезку на
оси ординат, отсекаемому прямой в опыте с ингибированием ( на рис. 4 это
отрезок ОА2), а взятая с минусом обратная величина константы
Михаэлиса по отрезку, в котором сходятся на оси абсцисс обе прямые (на рис. 4
это отрезок ОВ), или по формуле (24) через угловые коэффициенты обеих
прямых.
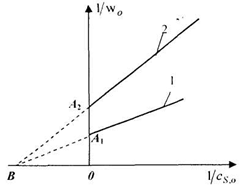
Рис. 4. Обработка
экспериментальных данных ферментативной реакции с неконкурентным
ингибированием:
- в отсутствие
ингибитора; 2-с ингибитором
При бесконкурентном
ингибировании механизм реакции имеет вид:
(3.46)
(3.47)
В этом случае образуется
один неактивный комплекс ES1
за счет присоединения ингибитора к промежуточному соединению ES с
константой равновесия:
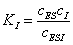 (3.48)
(3.48)
Решение задачи
аналогично разобранным ранее примерам и приводит к следующему выражению
скорости реакции в начальный период времени:
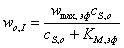 ,(3.49)
,(3.49)
причем 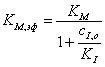 ,(3.50)
,(3.50)
а величина эффективной
максимальной скорости совпадает со случаем неконкурентного ингибирования, т.е.
рассчитывается по уравнению (44).
Обработка
экспериментальных данных в двойных обратных координатах, приведенных на рис. 5,
позволяет определить значения всех констант уравнения Михаэлиса: величина,
обратная предельной скорости определяется по отрезку, отсекаемому на оси
ординат прямой в опыте без ингибирования (на рис.5 это отрезок ОА1);
величина, обратная эффективной предельной скорости находится по отрезку на оси
ординат, отсекаемому прямой в опыте с ингибированием (на рис. 5 это отрезок ОА2);
из отрезков, отсекаемых на оси абсцисс соответствующими прямыми легко найти
величины констант Михаэлиса и эффективной константы реакции (на рис. 5 это
отрезки ОВ1 и ОВ2).

Рис. 5. Обработка
экспериментальных данных ферментативной реакции с бесконкурентным ингибированием:
1 - в отсутствие
ингибитора; 2-с ингибитором
2.2
Исследование кинетической закономерности заданной реакции, осложненной
ингибированием(
ингибирование напрексеном простагландин-Н-синтетазы (PGH) в
ферментативной реакции, одним из субстратов которой является адреналин).
Зависимость начальной скорости реакции
(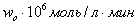 )) от
концентрации субстрата в различных опытах для реакции С.
)) от
концентрации субстрата в различных опытах для реакции С.
|
Номер опыта
|
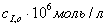 Концентрация
субстрата, Концентрация
субстрата, 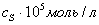
|
|
|
|
6,25
|
7
|
8
|
10
|
12,5
|
16,7
|
25
|
50
|
|
48
|
0
|
1,0
|
1,09
|
1,19
|
1,49
|
1,67
|
2,0
|
2,38
|
|
52
|
40
|
0,88
|
0,96
|
1,03
|
1,15
|
1,26
|
1,41
|
1,63
|
1,9
|
|
56
|
80
|
0,78
|
0,85
|
0,9
|
0,98
|
1,05
|
1,18
|
1,3
|
1,5
|
|
61
|
130
|
0,67
|
0,71
|
0,75
|
0,82
|
0,85
|
0,95
|
1,0
|
1,12
|
|
66
|
200
|
0,6
|
0,64
|
0,66
|
0,70
|
0,76
|
0,8
|
0,85
|
0,92
|
Кривые зависимости
начальных скоростей реакции от начальных концентраций субстрата при различных
концентрациях ингибитора.
а) 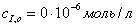 ;б)
;б)
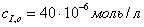 ;в)
;в)
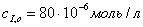
г)  д)
д)
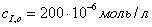
Кривая зависимости
начальной скорости от начальной концентрации ингибитора при 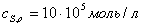
Переведем
экспериментальные данные о начальных скоростях реакции и начальных
концентрациях субстрата в обратные величины
Зависимость обратной
величины начальной скорости от обратной величины начальной концентрации
субстрата для разных концентраций ингибитора
|
Номер опыта
|
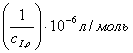 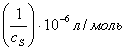
|
|
|
|
1,6
|
1,4
|
1,3
|
1
|
0,8
|
0,6
|
0,4
|
0,2
|
|
48
|
0
|
1,0
|
0,91
|
0,84
|
0,75
|
0,67
|
0,60
|
0,5
|
0,42
|
0,025
|
1,14
|
1,04
|
0,97
|
0,87
|
0,79
|
0,71
|
0,61
|
0,53
|
|
56
|
0,013
|
1,28
|
1,18
|
1,11
|
1,02
|
0,95
|
0,85
|
0,77
|
0,67
|
|
61
|
0,008
|
1,49
|
1,41
|
1,33
|
1,22
|
1,18
|
1,05
|
1,0
|
0,89
|
|
66
|
0,005
|
1,67
|
1,56
|
1,52
|
1,43
|
1,32
|
1,25
|
1,18
|
1,09
|
По полученному
расположению прямых мы можем сослаться на совпадение данных с соответствующим
рисунком в теоретической части 3.1 рис. 5 обработка экспериментальных данных
ферментативной реакции с бесконкурентным ингибированием. а) -
в отсутствие ингибитора; б) , в) ,
г) ,
д) -
с ингибитором
Обработка
экспериментальных данных в двойных обратных координатах, приведенных на нашем
графике, позволяет определить значения всех констант уравнения Михаэлиса:
величина, обратная предельной скорости определяется по отрезку, отсекаемому на
оси ординат прямой в опыте без ингибирования (отрезок ОА1);
величина, обратная эффективной предельной скорости находится по отрезку на оси
ординат, отсекаемому прямой в опыте с ингибированием ( отрезок ОА2,
ОА3, ОА4, ОА5); из отрезков,
отсекаемых на оси абсцисс соответствующими прямыми легко найти величины
констант Михаэлиса и эффективной константы реакции (отрезки ОВ1 и
ОВ2, ОВ3, ОВ4, ОВ5).
Находим  и
графическим
способом:
и
графическим
способом:
 ;
;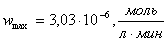
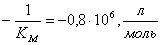 ;
;
Уравнение
Михаэлиса-Ментен для данной реакции
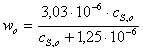
Находим 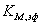 и
и
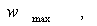 графическим
способом:
графическим
способом:
б) 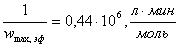 ;
;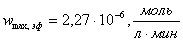
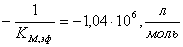 ;
;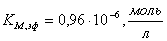
в) 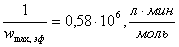 ;
;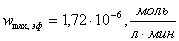
 ;
;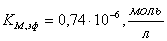
г) 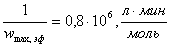 ;
;
 ;
;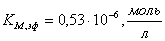
д) 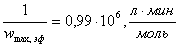 ;
;
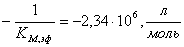 ;
;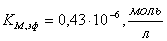
Находим по
формуле
б) 
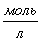
в) 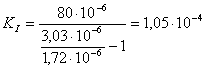
г) 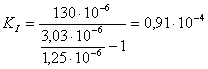
д) 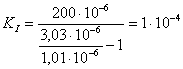
Рассчитаем среднюю
константу ингибирования 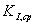
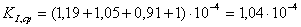
Уравнение
Михаэлиса-Ментен:
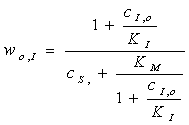

Вывод
В данной курсовой работе
мною были изучены закономерности ферментативных реакций с помощью литературы по
физической химии, биокинетики, основам химической кинетики и катализа.
В результате
исследования кинетики ферментативной реакции окисления сукцината натрия в
фумарат натрия под действием сукциноимидазы получено уравнение Михаэлиса-Ментен
со следующими параметрами:
где 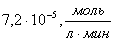 -
это максимальная скорость реакции ;
-
это максимальная скорость реакции ; 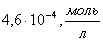 -
это константа Михаэлиса
-
это константа Михаэлиса
Найдена средняя
константа ингибирования:

Уравнение
Михаэлиса-Ментен принимает вид:
Список используемой
литературы
1. Степановских Е. И., Макурин Ю.Н.
2005 г. "Кинетические закономерности ферментативных реакций".
. Байрамов В.М. Основы химической
кинетики и катализа / под ред. В.В.Лунина. М.: изд.центр "Академия".
. Ашмор П. Катализ и ингибирование
химических реакций. М.: Мир, 1966.
. Физическая химия. 2 кн./под ред.
К.С.Краснова М.: Высшая школа, 1995.
. Стромберг А.Г., Семченко Д.П.,
Физическая химия М.: Высшая школа, 1999.
. Ипполитов Е.Г., Артемов А.В.,
Батраков В.В. Физическая химия. М.: изд. Центр "Академия", 2005.