Пластиды и их пигменты. Выделительные системы растений
Контрольная работа
Троицк, 2006 год
I.
ПЛАСТИДЫ И ИХ ПИГМЕНТЫ, ФОТОСИНТЕЗ, НЕОБХОДИМЫЕ ДЛЯ НЕГО УСЛОВИЯ, ДЕЛЕНИЕ
КЛЕТКИ
I.1 Пластиды и их пигменты
I.2 Фотосинтез, необходимые для него условия
I.3
Деление клетки
II. ВЫДЕЛИТЕЛЬНЫЕ СИСТЕМЫ
РАСТЕНИЙ (железистые волоски, выделительные ходы, млечники и т.д.).
ОХАРАКТЕРИЗУЙТЕ БАЛЬЗАМЫ, ЭФИРНЫЕ МАСЛА, СМОЛЫ, КАМЕДИ
II.1 Выделительные системы растений (железистые волоски,
выделительные ходы, млечники и т.д.)
II.2 Характеристика бальзамов, эфирных масел, смол,
камеди
III.
ПОЧКИ, ТИПЫ ПОЧЕК ПО ПРОИСХОЖДЕНИЮ. ПРИВЕДИТЕ РИСУНОК ВЕГЕТАТИВНОЙ ПОЧКИ В
ПРОДОЛЬНОМ РАЗРЕЗЕ IV. В КРУПНОМ МАСШТАБЕ НАРИСУЙТЕ СЕМЯЗАЧАТОК С
ЗАРОДЫШЕВЫМ МЕШКОМ В ПРОДОЛЬНОМ РАЗРЕЗЕ. ИЗ КАКИХ ЧАСТЕЙ СЕМЯЗАЧАТКА
РАЗВИВАЮТСЯ ТЕ ИЛИ ИНЫЕ ЧАСТИ СЕМЕНИ
V.
СПОСОБЫ РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН. ПРИВЕДИТЕ ПРИМЕРЫ. БИОЛОГОИЧЕСКАЯ РОЛЬ
РАСПРОСТРАНЕНИЯ ПЛОДОВ И СЕМЯН
VI.
ПОНЯТИЕ О ВИДЕ РАСТЕНИЙ. СОСТАВЛЕНИЕ ВИДОВЫХ НАЗВАНИЙ РАСТЕНИЙ СОГЛАСНО
БИНАРНОМУ МЕТОДУ К. ЛИННЕЯ. ВЫПИШЕТЕ ИЗ «СПИСКА ОСНОВНЫХ СЕМЕЙСТВ И ВИДОВ» 6
ВИДОВ (ИЗ НИХ 2 ОДНОГО РОДА) ИЗ РАЗНЫХ СЕМЕЙСТВ
VII.
ЖИЗНЕННЫЙ ЦИКЛ ПЛАУНА БУЛАВОВИДНОГО И СЕЛАГИНЕЛЛЫ НАРИСУЙТЕ КОЛОСКИ И ЗАРОСТКИ
ЭТИХ РАСТЕНИЙ
VIII.
ХАРАКТЕРИСТИКА СЕМ. БОБОВЫЕ (МОТЫЛЬКОВЫЕ). УКАЖИТЕ ВАЖНЕЙШИЕ ДИКОРАСТУЩИЕ И
КУЛЬТУРНЫЕ РАСТЕНИЯ ИЗ ЭТОГО СЕМЕЙСТВА (НЕ МЕНЕЕ 15 ВИДОВ)
IX. ПОНЯТИЕ О ЗОНАЛЬНОЙ И ИНТРАЗОНАЛЬНОЙ
РАСТИТЕЛЬНОСТИ. ПРИВЕДИТЕ ПРИМЕРЫ
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
Пластиды это органеллы,
характерные исключительно для растительных клеток. В них происходит первичный и
вторичный синтез углеводов. Формы, размеры, строение и функции пластид
различны. По окраске (наличию или отсутствию пигментов) Различают три типа
пластид:
1.
Зеленые
хлоропласты – это органеллы фотосинтеза, относительно крупные структуры (5-10
мкм. в длину при диаметре 2-4 мкм.) овальной или дисковидной формы. Содержимое
пластид называют стромой (матрикс). В строме содержатся белки, липиды, ДНК
(кольцевая молекула), РНК, рибосомы и запасные вещества (липиды, крахмальные и
белковые зерна). Кроме того, в строме содержатся также ферменты, участвующие в
фиксации диоксида углерода. Наружная мембрана гладкая. Внутренняя – образует
пластинчатые впячивания – тилакоиды, большая часть которых укладывается в виде
стопки монет и образует граны. Хлоропласты образуются из пропластид –
мельчайших сферических недеференцированных телец, котрые содержатся в растущих
частях растения (в клетках зародыша, образовательной ткани) и имеют зеленый
цвет, обусловленный присутствием фотосинтезирующим пигмента хлорофилла.
Существует несколько видов хлорофилла.
Наиболее распространен хлорофилл а (найден
у всех зеленых растений и цианобактерий) имеет голубовато-зеленый цвет
Хлорофилл б – желтовато-зеленый. Его
молекула содержит на один атом кислорода больше и на два атома водорода меньше.
В процессе фотосинтеза хлорофиллу
принадлежит ведущая роль. Он может поглощать солнечную энергию, запасать ее или
передавать ее другим молекулам.
Кроме того в состав хлоропластов
входят желтые пигменты – каротиноиды. Каротиноиды представляют собой высокомолекулярные
углеводороды: оранжевый каротин С40Н56 и желтый
ксантофилл С40Н56О2. Каротиноиды хлоропластов,
а также синие, красные, бурые пигменты хроматофоров водорослей называют
дополнительными, вспомогательными пигментами, поскольку энергия, поглощенная
ими, может передаватться на хлорофилл а. Хлорофилл использует энергию красной
части спектра, каротиноиды – синей. По своему строению каротиноиды являются
либо углеводородами (каротины), либо окисленными углеводородами, т. е.
Кислородсодержащими (каротинолы или ксантофиллы).Кроме того каротиноиды могут
защищать молекулы хлорофилла от чрезмерного фотоокисления на ярком свету.
2.
Лейкопласты
– бесцветные округлые пластиды, в которых обычно накапливаются запасные
питательные вещества, в основном крахмал. По строению лейкопласты мало
отличаются от протопластид, из которых он образуется: двумембранная оболочка
окружает бесструктурную строму. Внутренняя мембрана, врастая в строму, образует
немногочисленные тилакоиды. В лейкопластах имеются ДНК, рибосомы, а также
ферменты, осуществляющие синтез и гидролиз запасных веществ, в первую очередь
крахмала. Лейкопласты, в которых синтезируется и накапливается запасной
крахмал, называют амилопластами, белки – протеинопластами, масла –
элайопластами. В одено лейкопласте могут накапливаться разные вещества.
Запасной белок может откладываться форме кристаллов или аморфных включений,
масла – в виде пластоглобул. Однако белки и масла в лейкопластах встречаются
довольно редко. В амилопластах в связи с тилакоидами в строме возникают
образовательные центры, вокруг которых в виде зерен откладывается вторичный
запасной крахмал из растворимых углеводов, образовавшихся в хлоропластах в
процессе фотосинтеза. Много амилопластов в клетках клубней картофеля, зерновок
ржи, пшеницы и других органах растений где откладываются запасные вещества. В
секреторных клетках они в комплексе с агранулярным ретикуломом участвую в
синтезе эфирных масел.
3.
Хромопласты
– пластиды оранжево-красного и желтого цвета, образующиеся из лейкопластов и
хлоропластов в результате накопления в их строме каротиноидов. Они встречаются
в клетках лепестков (лютик, нарцисс, тюльпан, одуванчик), зрелых плодов (томат,
тыква, арбуз, апельсин), редко – корнеплодов (морковь, кормовая свекла), а
также в осенних листьях.
Хромопласты – конечный этап в развитии
пластид. По форме накопления каротиноидов различают следующие типы
хромопластов: глобулярный пигменты растворены в липидных пластоглобулах;
фибриллярный – пигменты накапливаются в белковых нитях; кристалический –
пигменты откладываются в виде кристалов.
Косвенное биологическое значение хромопластов
в том, что ярко окрашенные плоды успешнее распространяются птицами и животными,
а выделяющиеся яркой желто-красной окраской цветки привлекают насекомых –
опылителей.
В филогенезе первичным исходным типом
пластид являются хлоропласты, из которых в связи со специализацией органов
произошли лейко- и хромопласты. В онтогенезе взаимопревращения пластид
происходит иными путями. Наиболее часто хлоропласты превращаются в хромопласты
при осеннем пожелтении листьев или созревании плодов. В природе этот процесс
необратим. Лейкопласты могут превращаться в хлоропласты (позеленение верхней
части корнеплода моркови, оказавшейся на поверхности почвы) или хромопласты.
Хлоропласты могут при помещении растения в темноту превратиться в лейкопласты.
Процесс этот обратим.
I.2 Фотосинтез, необходимые для него условия
Фотосинтез у зеленых растений – это
процесс преобразования света в химическую энергию органических соединений,
синтезируемых из диоксида углерода и воды. Процесс фотосинтеза представляет
собой цепь окислительно-восстановительных реакций, совокупность которых
подразделяют на две фазы – световую и темновую.
1.
Световая
фаза. Для
этой фазы характерно то, что энергия солнечной радиации, поглощенная пигментами
системы хлоропластов, преобразуется в электрохимическую.
При действии света на хлоропласт
начинается электронный поток по системе переносчиков – сложных органических
соединений, встроенных в мембраны тилакоидов. С переносом электронов по ЭТЦ
сопряжено активное поступление протонов через тилакоидную мембрану из стромы
внутрь тилакоида. В тилакоидном пространстве происходит увеличение концентрации
протонов за счет расщепления молекул воды и в результате окисления электронного
переносчика пластохинона на внутренней стороне мембраны. Когда протоны идут
обратно по градиенту из тилакоидного пространства в строму, на наружной
поверхности тилакоида с участием фермента АТФ-синтетазы из АДФ и фосфорной
кислоты синтезируется АТФ, т. е. происходит фотосинтетическое фосфореилирование
с запасанием энергии в АТФ, которая затем переходит в строму хлоропласта.
Заканчивается передача электронов
следующим образом. Достигнув внешней поверхности мембраны тилакоида, пара
электронов следует с ионом водорода, находящимся в строме. Оба электрона и ион
водорода присоеденяются к молекуле переносчика водорода – НАДФ+
(никатиномидадениндинуклетидфосфат), который при этом переходит в свою
востановленную форму
НАДФ•Н+Н+:
НАДФ++2Н++2е-→НАДФ•Н+Н+
Следовательно активированные световой
энергией электроны используются на присоедининие атома водорода к его
переносчику, т. е. на восстановление НАДФ+ в НАДФ•Н+Н+,
который с наружной поверхности фотосинтетической мембраны переходит в
строму.
В молекулах хлорофилла,
утративших свои электроны, образовавшиеся электронные «дырки» действуют как
сильный окислитель и отрывают электроны от молекул воды. Через ряд переносчиков
эти электроны передаются на молекулу хлорофилла и заполняют «дырку». Внутри
тилакоида происходит фотоокислние (фотолиз) воды, в результате которого
выделяется свободный кислород, а также накапливаются ионы водорода
2Н2О→4Н++4е-+О2
Таким образом, во время световой фазы
фотосинтеза происходят три процесса: образование кислорода вследствие
разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ•Н2.
Кислород диффундирует в атмосферу, а АТФ и НАДФ•Н2 транспортируются
в матрикс пластид и участвуют в процессе темновой фазы.
2.Темновая фаза фотосинтеза протекает в
матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд
последовательных преобразований СО2, поступаещего из воздуха.
Осуществляются реакции темновой фазы за счет энергии АТФ и НАДФ•Н2 и
использовании имеющихся в пластидах пятиуглеродных сахаров, один из которых –
рибулозодифосфат – является акцептором СО2. Ферменты связывают
пятиуглеродный сахар с углекислым газом воздуха. При этом образуются соединения
которые последовательно восстанавливаются до шестиуглеродной молекулы глюкозы.
Суммарная реакция фотосинтеза
6СО2+6Н2 энергия света С6Н12О6+6О2
 хлорофилл
хлорофилл
В процессе фотосинтеза кроме
моносахаридов (глюкоза и др.), которые превращаются в крахмал и запасаются
растением, синтезируются мономеры других органических соединений – аминокислоты,
глицерин и жирные кислоты. Таким образом, благодоря фотосинтезу растительные, а
точнее – хлорофиллсодержащие, клетки обеспечивают себя и все живое на Земле
необходимыми органическими веществами и кислородом.
Описано три способа деления
эукариотических клеток: амитоз (прямое деление), митоз (непрямое деление) и
мейоз (редукционное деление).
Амитоз – относительно редкий способ
деления клетки. При амитозе интерфазное ядро делится путем перетяжки,
равномерное распределение наследственного материала не обеспечивается. Нередко
ядро делится без последующего разделения цитоплазмы и образуются двухъядерные
клетки. Клетка, претерпевшая амитоз, в дальнейшем не способна вступать в
нормальный митотический цикл. Поэтому амитоз встречается, как правило, в
клетках и тканях, обреченных на гибель.
Митоз. Митоз, или непрямое деление,
- основной способ деления эукариотических клеток. Митоз – это деление ядра,
которое приводит к образованию двух дочерних ядер, в каждом из которых имеется
точно такой же набор хромосом, что и был в родительском ядре.
В непрерывном процессе митотического
деления различают четыре фазы: профазу, метафазу, анафазу и телофазу.
Профаза – самая длительная фаза
митоза, когда происходит перестройка всей структуры ядра для деления. В профазе
происходит укорочение и утолщение хромосом вследствие их спирализации. В это
время хромосомы двойные (удвоение происходит в S-периоде интерфазы), состоят из
двух хроматид, связанных между собой в области первичной перетяжки осбой
структурой – цетромерой. Одновременно с утолщением хромосом исчезает ядрышко и
фрагментируется (распадается на отдельные цистерны) ядерная оболочка. После
распада ядерной оболочки хромосомы свободно и беспорядочно лежат в цитоплазме.
Начинается формирование ахромативного веретена – веретена деления, которое
представляет систему нитей, идущих от полюсов клетки. Нити веретена имеют
диаметр около 25нм. Это пучки микротрубочек, состоящих из субъедениц белка
тубулина. Микротрубочки начинают формироваться со стороны центриолей либо со
стороны хромосом (в клетках растений).
Метафаза. В метафазе завершается
образование веретена деления, которое состоит из микротрубочек двух типов:
хромосомных, которые связываются с центромерами хромосом, и ценросомных (полюсных),
которые тянутся от полюса к полюсу клетки. Каждая двойная хромосома
прикрепляется к микротрубочкам веретена деления. Хромосомы как бы выталкиваются
микротрубочками в область экватора клетки, т.е. располагаются на равном
расстоянии от полюсов. Они лежат в одной плоскости и образуют так называемую
экваториальную, или метафазную пластинку. В метафазе отчетливо видно двойное
строение хромосом, соединенных только в области центромеры. Именно в этот
период легко подсчитать число хромосом, изучать их морфологические особенности.
Анафаза начинается делением
центромеры. Каждая из хроматид одной хромосомы становится самостоятельной
хромосомой. Сокращение тянущих нитей ахроматинового веретена увлекает их к
противоположным полюсам клетки. В результате у каждого из полюсов клетки
оказывается столько же хромосом, сколько было их в материнской клетке, причем
набор их одинаков.
Телофаза – последняя фаза митоза.
Хромосомы деспирализуются, становятся плохо заметными. На каждом из полюсов
вокруг хромосом воссоздается ядерная оболочка. Формируются ядрышки, веретено
деления исчезает. В образовавшихся ядрах каждая хромосома состоит теперь всего
из одной хроматиды, а не из двух.
Каждое из вновь образовавшихся ядер получило
весь объем генетической информации, которым обладала ядерная ДНК материнской
клетки. В результате митоза оба дочерних ядра имеют одинаковое количество ДНК и
одинаковое число хромосом, такое же, как в материнском.
Цитокинез – после образования в телофазе двух новых ядер
происходит деление клетки и формирование в экваториальной плоскости перегородки
– клеточной пластинки.
В ранней телофазе между двумя дочерними
ядрами, не достигая их, формируется цилиндрическая система волокон, называемая фрагмопластом,
которая также как и волокна ахроматинового веретена, состоит из микротрубочек и
связаны с ним. В центре фрагмопласта на экваторе между дочерними ядрами
скапливаются пузырьки Гольджи, содержащие пектиновые вещества. Они сливаются
друг с другом и дают начало клеточной пластинке, а их мембраны участвуют в
построении плазмолемм по обеим сторонам пластинки. Клеточная пластинка
закладывается в виде диска, взвешенного в фрагмопласте. Волокна фрагмопласта,
видимо, контролируют направление движения пузырьков Гольджи. Клеточная
пластинка растет центробежно по направлению к стенкам материнской клетки за
счет включения в нее полисахаридов все новых и новых пузырьков Гольджи.
Клеточная пластинка имеет полужидкую консистенцию, состоит из аморфного протопектина
и пектатов магния и кальция. В это время из трубчатого ЭР образуются
плазмодесмы. Расширяющийся фрагмопласт постепенно приобретает форму бочонка,
позволяя клеточной пластинке расти латерально, пока она не соединится со
стенками материнской клетки. Фрагмопласт исчезает, обособление двух дочерних
клеток заканчивается. Каждый протопласт откладывает на клеточную пластинку свою
первичную клеточную стенку.
Цитокинез с помощью клеточной пластинки
происходит у всех высших растений и некоторых водорослей. У остальных
организмов клетки делятся внедрением клеточной оболочки, которая постепенно
углубляется и разделяет клетки.
Биологическое значение митоза состоит в строго одинаковом распределении
между дочерними клетками материальных носителей наследственности – молекул ДНК,
входящих в состав хромосом. Благодаря равномерному разделению реплицированных
хромосом между дочерними клетками обеспечивается образование генетически
равноценных клеток и сохраняется преемственность в ряду клеточных поколений.
Это обеспечивает таки важные моменты жизнедеятельности, как эмбриональное
развитие и рост организмов, восстановление органов и тканей после повреждения.
Митотическое деление клеток является также цитологической основой бесполого
размножения организмов.
Мейоз. Мейоз – это особый способ деления клеток, в результате
которого происходит редукция (уменьшение) числа хромосом вдвое и переход клеток
из диплоидного состояния (2n) в гаплоидное (n). Мейоз – единый, непрерывный
процесс состоящий из двух последовательных делений, каждое из которых можно
разделть на те же, что и в митозе, четыре фазы: профазу, метафазу, анафазу и
телофазу. Обоим делениям предшествует одна интерфаза. В синтетическом периоде
интерфазы до начала мейоза удваивается количество ДНК и каждая хромосома
становится двухроматидной.
Первое мейотическое, или редукционное,
деление.
Профаза I продолжается
от нескольких часов до нескольких недель. Хромосомы спирализуются. Гомологичные
хромосомы коньюгируют, образуя пары – биваленты. Бивалент состоит из четырех
хроматид двух гомологичных хромосом. В бивалентах осуществляется кроссинговер –
обмен гомологичными участками гомологичных хромосом, что приводит к их
глубокому преобразованию. Во время коссинговера происходит обмен блоками генов,
что объясняет генетическое разнообразие потомства. К концу профазы исчезает
ядерная оболочка и ядрышко, формируется ахроматиновое веретено.
Метафаза I –
биваленты собираются в экваториальной плоскости клетки. Ориентирование
материнской и отцовской хромосомы из каждой гомологичной пары к одному или
другому полюсу веретена деления является случайным. К центромере каждой из
хромосом присоединяется тянущая нить ахроматинового веретена. Две сетринские
хроматиды не разделяются.
Анафаза I – происходит сокращение тянущих нитей, и к полюсам
расходятся двухроматидные хромосомы. Гомологичные хромосмы каждого из
бивалентов уходят к противоположным полюсам. Расходятся случайно
перераспределенные гомологичные хромосомы каждой пары (независимое
распределение), и на каждом из полюсов собирается половинное число (гаплоидный
набор) хромосом, образуется два гаплоидных набора хромосом.
Телофаза I – у полюсов веретена собирается одиночный, гаплоидный,
набор хромосом, в котором каждый вид хромосом представлен уже не парой, а одной
хромосомой, состоящей из двух хроматид. В короткой по продолжительности
телофазе I восстанавливается ядерная оболочка, после чего материнская клетка
делится на две дочернии.
Второе мейотическое деление следует сразу же после первого и сходно с
обычным митозом (поэтому его часто называют митозом мейоза), только клетки,
вступающие в него, несут гаплоидный набор хромосом.
Профаза II – непродолжительная.
Метафаза II – снова образуется веретено деления, хромосомы
выстраиваются в экваториальной плоскости и центормерами прикрепляются к
микротрубочкам веретена деления.
Анафаза II – осуществляется разделение их ценромер и каждая
хроматида становится самостоятельной хромосомой. Отделившиеся друг от друга
дочерние хромосомы направляются к полюсам веретена.
Телофаза II – завершается расхождение сестринских хромосом к
полюсам и наступает деление клеток: из двух гаплоидных клеток образуются 4
клетки с гаплоидным набором хромосом.
Значение. Редукционное деление является как бы регулятором,
препятствующим непрерывному увеличению числа хромосом при слиянии гамет. Не
будь такого механизма, при половом размножении число хромосом удваивалось бы в
каждом новом поколении. Т.е. благодаря мейозу поддерживается определенное и
постоянное число хромосом во всех поколениях каждого вида растений, животных,
протист и грибов. Другое значение заключается в обеспечении разнообразия
генетического состава гамет как в результате кроссинговера, так и в результате
различного сочетания отцовских и материнских хромосом при их расхождении в
анафазе I мейоза. Это обеспечивает появление разнообразного и
разнокачественного потомства при половом размножении организмов.
Выделительные системы в жизнедеятельности
растений играют важную, разнообразную роль. В процессе их жизнедеятельности
образуется ряд веществ с разнообразной химической природой, не участвующих в
дальнейшем метаболизме.
Выделительные структуры растений
характеризует, то что клетки выделительных тканей паренхимные, тонкостенные. Их
ультраструктура связана с секретирующим веществом. В тканях, где синтезируются
эфирные масла, смолы, каучук, имеется хорошо развитый агранулярный ЭР, слизи - аппарат
Гольджи.
Выделительные ткани подразделяются на ткани
внешней (наружной) и внутренней секреции.
Наружные выделительные структуры (внешняя секреция). Связаны эволюционно с
покровными тканями. Они представлены следующими структурами: железистые
волоски, желёзки, переваривающие желёзки, нектарники, осмофоры, гидатоды.
Железистые волоски и желёзки представляют
собой трихомы эпидермы. Они состоят из живых клеток, обычно имеют удлиненную
ножку из одной или нескольких клеток и одно- или многоклеточную головку. Клетки
головки выделяют секрет под кутикулу. При разрыве кутикулы вещество изливается
наружу, после чего может образовываться новая кутикула и накопиться новая капля
секрета. Железистые волоски цветков герани, листьев и цветков душистого табака
выделяют эфирные масла; сидячие головчатые волоски, образующие мучнистый налёт
на листьях мари и лебеды, - воду и соли. Желёзки отличаются от волосков
короткой ножкой из несекретирующих клеток и многоклеточной головкой. Они
характерны у мяты, лаванды, полыни, черной смородины.
Нектарники обычно образуются на частях цветка,
но могут встречаться и на других надземных органах растения. Они могут быть
представлены отдельными поверхностными железистыми клетками или находиться в
ямках (лютик), желобках, шпорцах (живокость), возвышаться в виде бугорков,
подушечек (тыква, ива, яснотка белая). Выделительные клетки нектарников
отличаются густой цитоплазмой и высокой активностью обмена веществ. К
нектарнику может подходить проводящий пучок.
Осмофоры представляют собой или
специализированные клетки эпидермы, или особые желёзки, где секретируются
ароматические вещества. Выделение летучего секрета происходит в течение
короткого времени и связано с использованием запасных веществ. Аромат цветка
создается секрецией сложной смеси органических соединений, главным образом
эфирных масел.
Гидатоды выделяют капельно-жидкую воду и
растворенные в ней соли. При избытке воды и ослаблении транспирации через гидатоды
происходит гуттация – выделение капель воды из внутренних частей листа на его
поверхность. Специальной секреторной ткани здесь, как правило нет. Гидатоды
могут иметь вид многоклеточных волосков, устьиц, потерявших способность
регулировать величину своей щели, и, наконец представлять собой специальное
образование из большого числа клеток, расположенных под водным устьицем.
Гидатоды в виде многоклеточных волосков имются
у фасоли огненно-красной, а в виде водных устьиц встречается у манжетки,
настурции. Наиболее сложно устроенные гидатоды находятся в зубцах листьев
камнеломки, земляники, шиповника, чая. Здесь под водными устьицами образуется
многоклеточная ткань эпитема, через которую вода передается от водопроводящих
элементов к отверстиям устьиц.
Преваривющие желёзки на листьях насекомоядных
растений, например росянки, венериной мухоловки и др., выделяют жидкость,
содержащую пищеарительные ферменты и кислоты.
Внутренние выделительные структуры (внутренняя секреция). Вырабатывают и
накапливают вещества, остающиеся внутри растения. Это могут быть отдельные
секреторные клетки, рассеяные среди других тканей, как идиобласты. Они содержат
различные вещества, особенно часто оксалат натрия в виде одиночных кристаллов,
друз или рафид, бальзамы, танины, слизи и др.
Секреторные вместилища разнообразны по форме и
происхождению. Схизогенные вместилища образуются вследствие расхождения клеток
и формирования межклетника, выстланного живыми эпителиальными клетками и
заполненного выделенными веществами. К ним относятся смоляные ходы хвойных
растений. Лизигенные вместилища возникают в результате растворения группы
клеток с продуктами секреции. Такие вместилища видны в кожуре плодов цитрусовых
(апельсина, лимона, мандарина).
Млечники – особый тип выделительной ткани. Это
живые клетки (нечленистые млечники) или ряды слившихся клеток (членистые
млечники), пронизывающие всё растение. В зрелом млечнике протопласт занимает
постенное положение, полость млечника занята млечным соком – латексом, млечные
клетки неодревесневающие, эластичные. Латекс представляет собой эмульсию
белого, реже оранжевого или красного цвета. Жидкая основа латекса – клеточный
сок, в котором растворены или взвешены углеводы (крахмальные зерна у
молочайных, сахар у астровых), белки (у фикуса), жиры танины, слизи, эфирные
масла, каучук (более чем у 12 500 растений). Среди каучуконосов промышленное
значение имеет тропическая гевея – Hevea (семейство Молочайные), в млечном соке
которой содержится 40-50% каучука.
II.2 Характеристика бальзамов, эфирных масел,
смол, камеди
Эфирные масла — летучие органические вещества
самого разнообразного химического состава. Это бесцветные, иногда окрашенные,
сильно летучие жидкости с весьма характерным, часто приятным ароматическим
запахом и жгучим вкусом. Плохо растворяются в воде и хорошо в эфире,
хлороформе, спирте и жирных маслах. Придают своеобразный запах различным частям
растений, могут привлекать насекомых-опылителей, отпугивать травоядных. В мире
известно 2500 эфиромасличных растений. Эфирные масла встречаются в различных
частях растений — в цветках, листьях, плодах, а иногда и в подземных частях.
Содержание их в растениях колеблется от следов до 20% (обычно 2—3%).
Смолы — твердые и полужидкие липкие
органические вещества разнообразного химического состава. По химическому
строению близки к эфирным маслам, часто содержатся в растениях вместе с ними.
Они обладают характерным запахом, иногда сильно ароматическим. Некоторые смолы
обладают противомикробным, дезинфицирующим и ранозаживляющим свойствами, многие
являются балластными веществами. В медицинской практике применяются для
приготовления пластырей, настоек, назначаются внутрь как слабительное средство.
Бальзамы - природные
смолистые вещества, обогащенные эфирными маслами и др., которые образуются
преимущественно в тропических и субтропических растениях.
Камеди (гумми), высокомолекулярные углеводы,
составная часть соков и выпотов, выделяемых рядом растений при механических
повреждениях и некоторых заболеваниях. Используется в промышленности как клей,
стабилизатор суспензий и эмульсий и др., а также в виде слизей как
обволакивающее средство.

Почка (рис.1). Это зачаточный побег,
междоузлия которого очень малы. Внутри почки заключен апекс побега – его
меристематическая верхушка. Ниже него за счет выпячивания наружных слоев клеток
экзогенно формируются листовые бугорки, превращающиеся в зачатки листьев –
листовые примордии. Позже в их пазухах (обычно начиная с пазухи третьего-пятого
от апекса примордия листа) образуются бугорки (зачатки почек следующего порядка),
в связи с чем
Типы почек
В основу классификации почек положены различные
признаки.
I.
По наличию защитных
чешуй:
1.
Закрытые (защищенные)
2.
Открытые (голые)
II.
По составу и функции
почки бывают:
1.
вегетативные – состоят из
зачаточного стебля, чешуек, зачаточных листьев и конуса нарастания;
2.
генеративные – цветочные,
состоят из зачаточного стебля, чешуй и зачатков цветка или соцветия (бузина
красная);
3.
вегетативно-генеративные
(смешанные) – имеют зачатки листьев и цветков (яблоня, груша, вишня, слива и
т.д.);
4.
выводковые –
метаморфизированные почки с зачатками придаточных корней, предназначены для
вегетативного размножения. Они могут быть представлены луковичками
(бульбиллами), развивающимися в пазухах листьев надземных побегов (некоторые
лилии) или в соцветиях (некоторые луки); клубнелуковичками (гладиолусы); клубеньками
в пазухах листьев (горец живородящий); почками, формирующимися в углублениях
зубчиков листовой пластинки (каланхое).
III.
По месторасположению
на стебле:
1.
Верхушечные.
2.
Боковые.
1)
Пазушные – развиваются в
пазухах листьев и в свою очередь делятся на:
a)
Одиночные;
b)
сериальные почки,
образующие вертикальный ряд (свойственны двудольным). Последовательность их
заложения и развития:
·
восходящая (жимолость,
шалфей блестящий);
·
нисходящая (грецкий орех,
кирказон);
·
бисериальная (двурядная)
c)
коллатеральные почки,
образующие горизонтальный ряд (характерны, г.о., для однодольных – гладиолус,
крокус).
2)
Придаточные (адвентивные)
почки появляются на других органах: на листе (бриофиллюм, сердечник луговой,
бегония), у корнеотпрысковых растений на корнях (вишня, слива, малина, хрен и
т.д.).
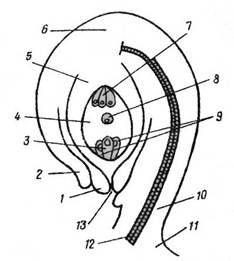
Рис. 2 Семязачаток
1, 2 – внутренний и наружный
ингументы; 3 – яйцеклетка;
4 – зародышевый мешок; 5 – нуцеллус;
6 – халаза;
7 – антиподы; 8 – вторичное ядро
(центральное); 9 – синергиды;
10 – фуникулюс; 11 – плацента;12 –
проводящий пучок;
13 – пыльцевход (микропиле)
Из каких частей семязачатка развиваются те или
иные части семени (рис. 2)
Зрелое семя состоит из зародыша,
запасающих тканей и семенной кожуры, которое развивается обычно после
оплодотворения из семязачатка.
Формирование зародыша происходит из зиготы
(оплодотворенной яйцеклетки) – 3. Первое деление приводит к формированию двух
клеток: базальной (со стороны макропиле) и терминальной (со стороны середины
зародышевого мешка). Базальная клетка делится в поперечном направлении и
формирует подвесок (суспензор), который прикрепляет зародыш к стенке
зародышевого мешка и вдвигает его в полость зародышевого мешка. Самая верхняя
крупная клетка-подвеска играет роль гаустория. Из терминальной клетки
развивается собственно зародыш, в результате трёх её делений развивается
шарообразное тело подзародыша, состоящее из мелких клеток. Далее зародыш на
верхушке становится плоским, и по обе стороны закладываются два бугорка. У
двудольных они развиваются симметрично, образуя две семядоли, у однодольных
одна семядоля развивается энергичнее, а другая отстает в росте. Асимметрично
развившаяся семядоля продолжает свой рост и занимает верхушечное положение, а
другая остается рудиментарной.
Формирование эндосперма – оплодотворенное
центральное ядро зародышевого мешка – 8, не проходя периода покоя делится и
дает начало триплоидному эндосперму. Существует два типа эндосперма: нуклеарный
и целлюлярный. При нуклеарном типе оплодотворенное централное ядро зародышевого
мешка многократно делится, формируя множество триплоидных ядер. Клеточные
стенки появляются поздно либо не формируются совсем. Нуклеарный тип эндосперма
наблюдается у однодольных и некоторых двудольных. Целлюлярный тип формирования
эндосперма отличается тем, что клеточные стенки образуются последовательно при
каждом делении. Такой тип встречается у некоторых двудольных.
Формирование семенной кожуры (спермодерма) развивается
из интегументов семязачатка. Она многослойна и присутствует в семени всегда.
Протопласты наружних клеток кожуры отмирают, и клетки заполняются смолистым
веществом, а стенки опробковевают, одревевсневают кутинизируются. Внутренние
слои клеток остаются тонкостенными. Главные функции спермодермы – защита
зародыша от механических повреждений, чрезмерного высыхания и преждевременного
насыщения влагой и прорастания.
Различают следующие способы
распространения плодов и семян:
·
без
участия посторонних агентов (семена и плоды крупных размеров);
·
с помощью
животных (сочные плоды, ягоду); ,
·
с помощью
ветра (плоды с крыльями и хохолками);
·
с помощью
воды (сухие плоды и семена);
·
с помощью
человека (все виды плодов и семян)
Автохория - распространение семян и
плодов без участия посторонних агентов (семена и плоды крупных размеров)
Осуществляется в двух основных
формах:
1) самопроизвольное опадение спелых
диаспор (плодов, семян и т.п.) под собственной тяжестью (барохория);
2) активное разбрасывание семян (в
радиусе от 1 до 15 м) при внезапном вскрывании плодов вследствии высокого и
неравномерного напряжения в околоплоднике (автомеханохория). Эта форма
автохории присуща и cочным плодам (некоторые тыквенные, бальзаминовые), но чаще
сухим (бобовые, гераниевые, виды фиалки и др.).
Эпизоохория - распространение семян и плодов с помощью животных
на их покровах - прежде всего на шерсти млекопитающих. Основные приспособления
- наличие придатков, цепляющихся за шерсть:
Calamagrostis arundinacea - Вейник тростниковидный. Для этого вида лесных
злаков эпизоохория - дополнительный способ переноса семян. Перенос облегчается
наличием коротких жёстких волосков и короткой ости в цветке.
Pulsatilla patens - Сон - трава, Прострел (также, как и другие виды
этого рода). Длинные опушённые придатки у плодиков позволяют им как
прикрепляться к шерсти животных, так и, отчасти, распространяться по ветру. Geum
rivale, Geum urbanum, Bidens.
Клейкие семена, или выделения плодов
Linnea borealis - И у этого и у следующего видов - клейкие железистые
волоски; Circaea alpina; способность прилипать целиком - у всего растения Lemna
(сочные плоды, ягоды);
Анемохория - распространение семян и плодов, спор и
других зачатков растений воздушными течениями. У семенных растений А.
обеспечивается либо малыми размерами семян (орхидные, заразиховые, многие
вересковые и др.), либо наличием на семенах или плодах «парашютных»
приспособлений — волосков (ивы, тополи, многие сложноцветные и др.), крылатых
выростов (вязы, ясени, клёны, берёзы, щавели, сосны, ели и мн. др.), остей (ковыли
и др.), пузыревидно вздутых оболочек и т. п. Такие семена и плоды могут
переноситься ветром на большие расстояния (по некоторым данным, до 40 км). Анемохорами
являются также лишайники, мхи, папоротники, хвощи, плауны, многие грибы,
наземные водоросли, многие бактерии, которые распространяются ветром благодаря
ничтожной массе спор или других зачатков. К анемохорам могут быть отнесены и
растения группы перекати-поле (плоды с крыльями и хохолками).
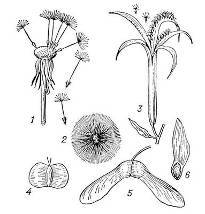
Рисунок 3
Плоды (1, 4, 5) и семена (2, 3, 6) с
приспособлениями для переноса их ветром: 1 — одуванчика; 2 — хлопчатника; 3 —
кипрея; 4 — берёзы; 5 — клёна; 6 -сосны.
Гидрохория – распространение плодов, семян и др. зачатков
растений водными течениями. Характерна преимущественно для болотных и водных
растений, водорослей и некоторых грибов. Приспособлениями для такого способа
переноса служат различные вздутия и выросты на плодовых или семенных оболочках
(или особые клетки — в спорах грибов), наполненные воздухом и действующие как
плавательные пузыри. Г. наблюдается у частухи, стрелолиста, сусака,
ежеголовника, рдеста и др. с помощью воды (сухие плоды и семена); с помощью
человека (все виды плодов и семян).
Антропохория – распространение семян и плодов в результате
деятельности человека. Антропохория может быть косвенной (диаспоры разносятся с
поливными водами, навозом или домашним скотом) и непосредственной: при
распространении диаспор с помощью транспорта (агестохория) — преднамеренная
антропохория (культивируемые растения) или непреднамеренная (попутно с другими
грузами распространяются диаспоры рудеральных, сорнополевых и дикорастущих
видов).
Последний способ наиболее быстрый и
эффективный по дальности.
Непосредственная антропохория
происходит при обработке почвы сельскохозяйственными орудиями и машинами —
эргазиохория (от греч. ergasia — обработка земли), она всегда непреднамеренна и
способствует возобновлению сорнополевых и сенокосных растений. Распространение
диаспор происходит и путём их высева — спейрохория (от греч. speiro — сею)
преднамеренная антропохория (культурные растения) или непреднамеренная
(сорнополевые растения). Эта форма антропохории привела к отбору
специализированных засорителей наиболее древних культур (пшеницы, риса, льна,
проса, гречихи). Диаспоры сорных видов, часто созревающих одновременно с
соответствующей культурой, настолько сходны с её семенами, что очень трудно
отсортировываются. К числу заносных сорных растений (антропохоров) в России
относятся виды амброзии, дурнишника, амаранта, а также новые — шалфей отогнутый
(Salvia reflexa), подсолнечник сорный.
Биологическая роль, благодаря вышеперечисленным способам
распространения, сводится к следующему: сохранение вида, завоевание новых
ареалов обитания, способствование распространению отдельных видов травоядных,
на тех территориях, на которых произрастает растительность служащая им пищей.
Вид – совокупность популяций особей, способных к скрещиванию с образованием
плодовитого потомства, населяющих определенную территорию, обладающих рядом
общих морфофизиологических признаков и типов взаимоотношений с абиотической и
биотической средой и отделенных от других таких же совокупностей особей полным
отсутсвием гибридных форм.
Вид – основная структурная единица в системе
живых организмов и особый качественный этап их эволюции. Вследствие этого вид
представляет собой основную таксономическую категорию в систематике.
Бинарная номенклатура – правило давать видам двойное название, введено К.
Линеем в 1753 г. Согласно этому методу название вида состоит из двух латинских
слов: родового названия и видового эпитета, например пшеница твердая – Triticum
durum, пшеница мягкая – Triticum aestivum.
6 Видов из разных семейств:
1.
Сем. Сельдерейные
(Зонтичные) – Apiaceae (Umellifrae)
Болиголов крапчатый – сonium maculatum L.
2.
Сем. Норичниковые – Scrophulariaceae
Наперстянка крупноцветковая - digitalis
grandiflora Mill.
3.
Сем. Лилейные – Liliaceae
Лук-порей – allium cepa L.
4.
Сем. Мятликовые (злаки) – Poaceae
(Gramineae)
 Рис посевной – oriza saliva L.
Рис посевной – oriza saliva L.
5.
Сем. Пасленовые – Solonoaceae
Белена черная – hyoscyamus niger L. Род Паслен
– Solanum
Картофель – solanum tuberosum L.
Плаун (Lycopodium)
Класс Плауновые
(Lycopodiopsida)
Порядок плауновые
(Lycopodiales)
Род плаун (Lycopodium)
Вид Плаун булавовидный – Lycopodium clavatum.
Относится к классу Равноспоровые плауны
Род плаун насчитывает около 400 видов. В
нашей стране произрастают 14 видов плаунов. Наиболее широкое распространение
имеет вид плаун булавовидный - Lycopodium
clavatum. Плаун булавовидный - вечнозеленое многолетнее травянистое
растение со стелющимся стеблем. На стебле образуются многочисленные
приподнимающиеся ветви длиной до 20 см. Побеги густо покрыты шиловидными
листочками. Стробилы со спорангиями сидят на облиственной ножке в числе 1-3.
Спорангии почковидной формы, содержат большое количество мелких спор желтого
цвета.
Споры после созревания опадают на почву
и прорастают в гаметофит (заросток). Гаметофит плаунов - многолетнее
бесхлорофильное растение в виде клубенька с ризоидами. Самостоятельно питаться
не может, поэтому его развитие начинает с проникновением в тело гиф грибы
(эндотрофная микориза). Заросток обоеполый, в глубине его ткани образуются
архегонии и антеридии. Оплодотворение происходит при наличии воды. После
оплодотворения из яйцеклетки развивается зародыш, вырастающий в многолетнее
вечнозеленое растение - спорофит.
В цикле развития плаунов преобладает
спорофит. Мейоз происходит в спорангии при образовании спор.
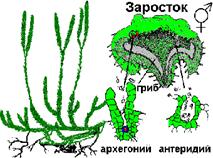
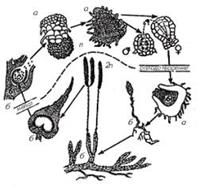
Рисунок 4. Чередование поколений в жизненном цикле плауна: а
– гаметофит; б – спорофит
Селягинелла (Selaginella)
Класс полушниковые (Isoetopsida)
Порядок селягинелловые
(Selaginellales)
Относится к классу – Разноспоровые плауны
Размножение
бесполое при помощи спор.Наибольшее распространение в нашей стране имеет
селягинелла селаговидная (Selaginella
selaginoides). Это мелкие многолетние растения, 5-12 см высотой, с
короткими ползучими стеблями, покрытыми удлиненно-яйцевидными листьями,
образующие рыхлую дерновинку. На верхушке побегов образуются спороносные
колоски. В стробилах находятся спорангии двух типов. В микроспорангиях
созревают микроспоры, которые при прорастании дают заросток (гаметофит), на
котором развиваются антеридии со сперматозоидами. В мегаспорах находятся 4
крупных мегаспоры, одна из которых прорастает в женский заросток с архегониями.
Созревшая в архегонии яйцеклетка оплодотворяется сперматозоидом с помощью воды
и образует зиготу. Из зиготы вырастает взрослое растение - спорофит. Заростки
селягинелл сильно редуцированы и обычно развиваются, не покидая оболочек
рассеившихся спор. В этом случае из микроспоры выходят уже сформированные
сперматозоиды. Даже после оплодотворениязародыш спорофита долгое время
продолжает использовать для роста запасные питательные вещества мегаспоры.
Распространены селягинеллы во влажных тропических лесах, мшистых лесах, на
торфяниках.

Рисунок 5.
Селагинелла обыкновенная: А – спорофит: 1 – придаточный корень, 2 – стебель с
листьями, 3
– стробил, 4 – молодой спорофит; Б – стробил со спорангиями: 5 – микроспорангий, 6 –
мегаспорангий, 7– микроспора, 8 – мегаспора
Семейство бобовые объединяет огромное
количество видов - около 18 тысяч. В современной систематике бобовые растения
отнесены к отделу Magnoliophita, классу Magnoliatae, подклассу Rosidae, порядку
Fabales Nakai, семейству Fabaceae L. Это семейство одно из наиболее филогенетически
молодых, но очень разнообразных по формам, включает около 490 родов и 12000
видов, широко распространенных от тропиков до холодных районов севера.
Крупнейшее подсемейство бобовых –
мотыльковые – содержит огромное число культурных (соя, горох), кормовых
(люцерна, вика) лекарственных (кассия) и декоративных растений.
Порядок бобовые – Fabales содержит
одно семейство бобовые. Семейство бобовые – Fabaceae объединяет 17000-18000
видов и 650 родов. Семейство принято делить на 3 подсемейства: цезальпиниевые,
мимозовые, мотыльковые. Крупнейшее семейство мотыльковые.
К бобовым принадлежит крупнейший в
мире род цветковых растений - астрагал (Astragalus) – около 2200 видов; широко
известны виды рода клевер (Trifolium), объединяющего 320 видов; акация (Acacia)
1200 тропических и субтропических видов; мимоза (Mimosa) 500 американских
тропических видов и другие. После злаков бобовые - важнейшая по практической
значимости для человека группа цветковых растений. В пределах своего ареала
бобовые занимают очень разнообразные местообитания.
Среди бобовых известны почти все
жизненные формы, исключая паразитов и эпифитов. Некоторые древесные
представители достигают 60-80 метров высоты и в тоже время существуют
крохотные, всего в несколько сантиметров однолетние травы. На корнях
значительной части бобовых имеются клубеньки, которые возникают как разрастания
паренхимной ткани их корней вследствие внедрения и эндогенного расселения
азотфиксирующих бактерий рода ризобиум.
Листья бобовых разнообразны, но
всегда сложные. Прилистники всегда имеются, однако часто рано опадают, либо
иногда сильно разрастаются (горох). Листья и отдельные листочки способны
совершать разного рода круговые движения или складываться на ночь.
Цветки обоеполые, нередко довольно
крупные яркие. У мотыльковых и цезальпиниевых они зигоморфные. Околоцветник
всегда двойной. Чашечка состоит из 5, реже из 4 срастающихся чашелистиков,
правильная или неправильная. Лепестков 5 (всё подсемейство мотыльковых, часть
цезальпиниевых и мимозовых). Венчик мотыльковых устроен характерно. Верхний,
наружный по своему положению в почке и обычно самый крупный лепесток получил
название флага (паруса). Он способствует привлечению насекомых опылителей. Рис.
6
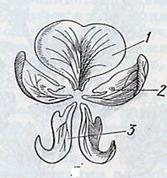
Рисунок 6 1. Флаг
(парус) 2. Крылья (весла) 3. Лодочка
Боковые лепестки называются крыльями (веслами)
и используются насекомыми в качестве посадочной площадки. Самые внутренние по
положению в почке лепестки обычно срастаются вдоль нижнего края, образуя
лодочку (киль). Лодочка подобно защитному футляру, предохраняет тычинки и
пестик и препятствует посещению цветков мелкими насекомыми, которые поедают
пыльцу или нектар, но не являются эффективными опылителями.
Андроцей у всех бобовых состоит из 10 тычинок
и может быть одно- и двубратственным. Для определения родов в рамках
рассматриваемого семейства важное значение имеет строение андроцея. У одних
родов андроцей многобратственный, когда все 10 тычинок свободные (роды: софора,
термопсис). У других — однобратственный, когда все 10 тычинок срастаются, между
собой тычиночными нитями, образуя так называемую тычиночную трубку внутри
которой располагается пестик (роды люпин и др.). Наконец, большинство родов
семейства имеет двубратственный андроцей: 9 тычинок срастаются тычиночными
нитями в трубку, а одна тычинка свободная (роды: горох, люцерна, вика, чина).
Форма тычиночной трубки также бывает разнообразной. В одних случаях она срезана
прямо и свободные концы тычиночных нитей одинаковой длины (чина), в других
случаях она срезана косо. Мотыльковые очень часто имеют гипантий (специфичная
деталь цветка, возникшая в результате срастания оснований чашелистиков,
лепестков и тычинок с тканями цветоложа). Гинецей почти всегда монокарпный, с
верхней одногнездной, редко двугнездной завязью.
Цветки собраны в простые соцветия кисть,
зонтик, головку.

Рисунок 7 Диаграммы цветка
Faboideae
Плод
— монокарпий: одно-, дву- или многосемянный боб, вскрывающийся, невскрывающийся
или членистый. Может быть опушенным или нет. У мотыльковых семена относительно
крупные, с эндоспермом или без него, зародыш с хорошо развитыми семядолями,
согнутый.
У большинства мотыльковых опыление
перекрестное, осуществляется крупными насекомыми — шмелями и пчелами, которых
привлекают обильные запасы нектара или пыльцы. Встречаются самоопыляющиеся
виды, например горох. Плоды распространяются под действием собственной тяжести,
ветром, водой. Семена вскрывающихся бобов разбрасываются силой раскручивающихся
створок.
В семенах бобовые содержат в качестве
запасного вещества белок, а также крахмал и жирное масло, что определяет их
большую пищевую и кормовую ценность. У значительного числа видов найдены
алкалоиды, тритерпеновые гликозиды, антраценпроизводные и т. д. В качестве
пищевых продуктов широко используются семена сои (Glycine max), фасоли (Phaseolus
vulgaris), гороха (Pisum sativum), чечевицы (Lens culinaris), арахиса, или
земляного ореха (arachis hypogaea), и др. Лучшими кормовыми травами считаются
люцерна посевная (Medicato sativa), люцерна серповидная, или желтая (M/
falcata), и разные виды клеверов. Одновременно бобовые заметно улучшают
плодородие почвы. Ежегодно виды бобовых, живущих в симбиозе с бактериями,
возвращают в почву не менее 100—140 кг/га азота.
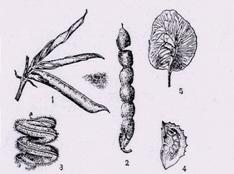
Примеры плодов бобовых (рисунок 8): 1 – типичный плод вики, 2 – разламывающийся
боб копеечника, 3 – спирально закрученный боб люцерны, 4 – односемянной боб
эспарцета, 5 – крылатый одно–двусемянной боб красного сандалового дерева.
Многие бобовые — ценные лекарственные
растения, например солодки голая и уральская (С1усуrrhiza glabra и С.
uralensis), корни и корневища которых содержат тритерпеновые сапонины и
флавоноиды. Кассии остролистная и узколистная (Cassia acutifolia и С.
angustifolia ) дают так называемый «александрийский лист» — классическое
слабительное средство. Многие акации и астрагалы выделяют камеди
(полисахариды), используемые в технике и медицине. Софора японская (Styphnolobium
japonicum) — промышленный источник получения флавоноида рутина, обладающего
Р-витаминной активностью. Многие бобовые высоко декоративны и культивируются
как в тропиках, так и в странах умеренного климата.
В дополнение к вышеназванным видам
семейства мотыльковые:
Клевер луговой -Trifolium pratense L.
Клевер гибридный -Trifolium hibridum
L.
Клевер ползучий -Trifolium repens L.
Люцерна посевная, или синяя -Medicago sativa L.
Горошек мышиный -Vicia cracca L.
Лядвенец рогатый -Lotus corniculatus L.
Чина посевная -Lathyrus sativus L.
Фасоль обыкновенная -Phasoeolus vulgaris
L.
фотосинтез плод растение
биологический
ИНТРАЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ,
естественная растительность, которая не образует самостоятельной зоны, а входит
в состав одной, нескольких или всех основных растительных зон. Напр.,
растительность речных наносов, различных водоемов, береговых дюн. Иногда
интразональная растительность преобладает над зональной, напр., болота Западной
Сибири в таежной подзоне.
ЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ,
естественная растительность,
характеризующая соответствующую
растительную зону (тундра, лес, степь, пустыня).
Зональная растительность занимает
наиболее типичные местообитания: ровные, возвышенные, хорошо дренированные
водораздельные плато (плакоры). Обычно зональная растительность преобладает над
другими (незональными) типами растительности. Лишь в некоторых случаях,
например в обширных, слабо дренированных понижениях с изолированными
возвышенностями или при резких сменах условий увлажнения в зависимости от
микрорельефа, зональная растительность выражена слабо или отсутствует.
Зональная растительность включает
большое число растительных формаций и ассоциаций, сменяющихся в пределах зоны с
С. на Ю. соответственно изменению климата (на этом основании зоны, имеющие
большую протяжённость, делят на подзоны). Большую часть земель, занятых прежде
зональной растительностью (главным образом в степи и лесостепи), используют
теперь как с.-х. угодья.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. Общая биология, учебник для 10-11
классов образовательных учреждений. Под редакцией Д.К.Беляева, профессора Г.М.Дымшица
и профессора Рувинского. Москва "Просвещение" 1996.
2. Общая биология, учебник для 10-11
классов образовательных учреждений. Под редакцией члена-корреспондента
Ю.И.Полянского. Москва "Просвещение" 1998.
3. Общая биология. Мамонтов С.Г.,
Захаров В.Б. Москва, "Высшая школа" 2002 г.
4. Биологический энциклопедический
словарь. Главный редактор М.С.Гиляров. Москва, научное изда-тельство
"Большая Российская энциклопедия" 1995.
5. "Биология для поступающих в
вузы" под редакцией академика РАМН, профессора В. Н. Ярыгина. Мо-сква
"Высшая школа" 1995 г.
6. Анастасова Л.П. Самостоятельные
работы учащихся по общей биологии. Москва, просвещение 1978 г.
7. Воронцов Н.Н., Сухорукова Л.Н.
Эволюция органического мира. Факультативный курс, учебное пособие для 10-11 классов
средней школы. Москва "Наука" 1996 г.
8. Гончаров О.В., Пименов А.В.
Пособие по биологии для поступающих в вузы. Саратов, 2001 г.
9. Грин Н., Стаут У., Тейлор Д.
"Биология" в трех томах. Москва, "Мир" 1996 г.
10. Гуляев Г.В. Генетика. - Москва,
Колос, 1984 г.
11. Киселева Э.А. Книга для чтения по
дарвинизму. Москва, "Просвещение", 1970 г.
12. Корсунская В.М., Мироненко Г.Н.,
Мокеева З.А., Верзилин Н.М. Уроки общей биологии. Пособие для учителей.
"Просвещение", 1977 г.
13. Миркин Б.М., Наумова Л.Г. Экология
России. Москва, АО МДС, Юнисам 1995 г.
14. Пехов А.П. Биология с основами
экологии. Санкт-Петербург "Лань" 2000 г.
15. Приходченко Н.Н., Шкурат Т.П.
Основы генетики человека. - Ростов н/Д, "Феникс", 1997 г.
16. Радкевич В.А. Экология. Минск,
"Вышэйшая школа", 1983 г.
17. Рейвн П., Эверт Р., Айкхорн С.
Современная ботаника. В 2 т. Пер. с англ. - М.: Мир, 1990 г.
18. Чернова Н.М., Былова А.М.
Экология. Москва, "Просвещение", 1981 г.
19. Яковлев Г.П., Аверьянов Л.В.
Ботаника для учителя. В 2 ч. - М.: Просвещение, 1996, ч.1; 1997 г, ч.2.
20. Ярыгин В.Н. Биология для
поступающих в вузы. Москва "Высшая школа" 1995 г.